
Abstract
The global public health threat of antimicrobial resistance has led the scientific community to highly engage into research on alternative strategies to the traditional small molecule therapeutics. Here, we review one of the most popular alternatives amongst basic and applied research scientists, synthetic antimicrobial peptides. The ease of peptide chemical synthesis combined with emerging engineering principles and potent broad-spectrum activity, including against multidrug-resistant strains, has motivated intense scientific focus on these compounds for the past decade. This global effort has resulted in significant advances in our understanding of peptide antimicrobial activity at the molecular scale. Recent evidence of molecular targets other than the microbial lipid membrane, and efforts towards consensus antimicrobial peptide motifs, have supported the rise of molecular engineering approaches and design tools, including machine learning. Beyond molecular concepts, supramolecular chemistry has been lately added to the debate; and helped unravel the impact of peptide self-assembly on activity, including on biofilms and secondary targets, while providing new directions in pharmaceutical formulation through taking advantage of peptide self-assembled nanostructures. We argue that these basic research advances constitute a solid basis for promising industry translation of rationally designed synthetic peptide antimicrobials, not only as novel drugs against multidrug-resistant strains but also as components of emerging antimicrobial biomaterials. This perspective is supported by recent developments of innovative peptide-based and peptide-carrier nanobiomaterials that we also review.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Background: the antibiotic resistance global threat and current molecular design approaches
Historical perspective
The first documented failure of antimicrobial therapy in humans due to acquired microbial resistance was reported in the 1940s for penicillin, which occurred only a few years after its commercialisation (Wright 2010). The therapeutic revolution initiated by penicillin motivated the discovery of varied molecular design approaches in the following few decades (Table 1).
Gramicidin was the first reported antimicrobial peptide (AMP), isolated from the soil bacteria Bacillus brevis in 1939, while defensin was the first evidenced animal AMP, isolated from rabbit leukocytes. Despite these early discoveries, it was only after the 1960s that global interest and research efforts were devoted to antimicrobial peptides, due to the rise of multidrug-resistant microbial pathogens. Presently, over 5000 AMPs have been reported (Zhang and Gallo 2016; Travkova et al. 2017).
Microbial evolution towards acquiring resistance to marketed drugs kept increasing over the twentieth century and early twenty-first century (Ventola 2015a; Aslam et al. 2018; Kennedy and Read 2018; Peterson and Kaur 2018). Decades of fundamental research unravelled four main types of molecular resistance mechanisms: secretion of inactivating enzymes, secretion of efflux pumps, immunity and bypass, and target modification. A number of excellent reviews have compiled these mechanisms for the antimicrobial agents currently on the market (Table 1, Fig. 1) (Davies and Davies 2010; Malmir et al. 2018; Ribeiro da Cunha et al. 2019; Magana et al. 2020). Two main factors are identified as favouring the increase of antimicrobial resistance: the generalized use and misuse of antimicrobial therapies and the insufficient discovery of new antimicrobial therapies (Table 1) (Ventola 2015a; Aslam et al. 2018; Kennedy and Read 2018; Peterson and Kaur 2018).

Main molecular mechanisms of action and resistance for marketed antibiotics. Modified from Wright 2010
Worldwide, numerous organizations, like the Centre for Disease Control and Prevention (CDC), the Infectious Diseases Society of America, the World Economic Forum, and the World Health Organization (WHO) have recognized antibiotic resistance as a “global public health concern”. Numerous attempts have been made to delineate the diverse aspects of antibiotic resistance (Li et al. 2012; Crofts et al. 2017; Sierra et al. 2017). WHO published several documents to guide the development of new antimicrobials, such as the list of global priority pathogens and corresponding target diseases (WHO 2017). In 2015, WHO launched the Global Antimicrobial Resistance and Use Surveillance System (GLASS) as the first global system to collect official national antimicrobial resistance data in selected bacterial pathogens (WHO 2015). In a span of 4 years, 91 countries and territories had already enrolled in the system. Based on worldwide data, the GLASS report of May 2020 revealed a worrying number of common bacterial infections increasingly resistant to available antibiotics (WHO 2020b).
Recently, concerns have been raised regarding an aggravation of the inappropriate use of antibiotics during the COVID-19 pandemic. Consequently, the WHO issued guidance to “NOT recommend antibiotic therapy or prophylaxis for patients with mild or moderate COVID-19 unless signs and symptoms of a bacterial infection exist” (Getahun et al. 2020).
Global priority pathogens and infectious diseases
The WHO priority list contains 12 bacterial families, including the ESKAPE pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species), which are known to represent a global threat to human health (WHO 2017). Mycobacteria was not included in the catalogue, because it is already considered a global priority for which innovative new treatments are urgently needed. The main objective of the list was to identify the most important resistant bacteria at a global level for which there is an urgent need for new treatments, in order to encourage the prioritisation of funding and incentives, to align research and development priorities of public health and to garner global coordination in the fight against those pathogens. The WHO priority pathogens were selected through a multi-criteria decision analysis technique contemplating both expert opinion and evidence-based data, following ten criteria: all-cause mortality, healthcare and community burden, prevalence of resistance, 10-year trend of resistance, transmissibility, preventability in hospital and community settings, treatability and current pipeline (Santajit and Indrawattana 2016; WHO 2017; Parish 2019; De Oliveira et al. 2020).
In May 2020, WHO released a report comprising the target product profiles (TTP) for needed antimicrobial agents for priority diseases, namely enteric fever, gonorrhoea, neonatal sepsis, and urinary tract infections. Table 2 compiles the diseases considered as priority targets, together with their available treatments and reported antimicrobial resistance (WHO 2020c).
The increasing occurrence of fungal infections that are resistant to the limited number of commonly used antifungal classes, such as triazoles, echinocandins, and polyenes, has lead the WHO to develop a priority list of fungal pathogens. This list is programmed to be launched at the end of 2021 with the main purpose of defining R&D priorities with identified public health needs. In April 2020, a first meeting of the WHO antifungal expert group took place to establish the parameters for pathogen selection. The priority fungal pathogens initially set out include: Candida auris; azole-resistant Candida spp.; azole-resistant Aspergillus fumigatus; Cryptococcus neoformans, Cryptococcus gattii; Pneumocystis jirovecii and Mucorales. Further evaluation will be made based on limitations of treatment options due to resistance and/or existing treatability issues for those pathogens (WHO 2020a).
Molecular design approaches in the pre-clinical and clinical pipelines
Since the publication of the first priority list by WHO in 2017 and until September 2019, eight new antibiotics were approved by the US and European authorities (FDA/EMA) with activity against the priority pathogens. However, most of these products are not very innovative, being derivatives of known classes, such as the tetracycline derivatives eravacycline, omadacycline, or sarecyclin, the cephalosporin derivative cefiderocol and the combination therapy imipenem-cilastatin-relebactam. Sarecycline is indicated for moderate to severe acne caused by Cutibacterium acnes while the combination therapy imipenem-cilastatin-relebactam and cefiderocol were approved for complicated urinary tract infections (Andrei et al. 2019; EMA 2020; FDA 2020).
As of September 2019, the WHO listed 49 small molecules and 10 biological molecules in clinical development that target priority pathogens, Mycobacterium tuberculosis and Clostridium difficile. Besides this high number, only six of these therapies did not display cross-resistance to existing antibiotics and between those six only two were active against multidrug-resistant Gram negative bacteria. This can be explained by the general lack of chemical diversity. Indeed, most of the new antimicrobial small molecules in clinical trials are combinatory therapies of beta lactams with beta-lactamase inhibitors or tetracycline derivatives. The six innovative small molecule antibiotics include two boronate beta-lactam inhibitors (taniborbactam-cefepime and VNRX-7145—ceftibuten), two new topoisomerase inhibitors (zoliflodacin and gepotidacin), a new FabI inhibitor (afabicin), and a FtsZ inhibitor (TXA709) (WHO 2019a). Antimicrobial biological medicines in clinical development include mono/polyclonal antibodies and endolysins, which all can be considered innovative, as they target new structures through new modes of action (classes: anti-Staphylococcus aureus immunoglobulin M (IgM) monoclonal antibody; anti-Staphylococcus aureus IgG monoclonal antibody; phage endolysin; anti-Pseudomonas aeruginosa IgG monoclonal antibody; anti-Pseudomonas aeruginosa serotype O11 IgG monoclonal antibody; anti-Staphylococcus aureus IgG monoclonal antibody; anti-Staphylococcus aureus IgG monoclonal antibody; anti-Clostridium difficile polyclonal antibody). However, the high cost of biologicals compared to small molecules limits their potential use as alternative treatments to current therapies (Peterson and Kaur 2018; WHO 2019a).
The first comprehensive global overview of the antibacterial pre-clinical pipeline was published by the WHO in 2019, compiling 252 antibacterial products being developed in 145 institutions around the world. The review identified relatively innovative products, with many projects including direct-acting small molecules (n = 108, 42.9%), antimicrobial peptides (n = 27, 10.7%), and phage/phage-derived peptides (n = 28, 11.1%) (WHO 2019b). Considering the WHO priority pathogens list, examination of the pre-clinical pipeline projects indicates that a significant number of products were focused on single pathogenic species, representing a consistent shift rather than broader spectrum agents. Almost one-third of these products target bacterial cell wall synthesis or directly act on the bacterial membrane because of the less likely occurrence of resistance to these mechanisms of action (Kennedy and Read 2018). However, for these new non-traditional approaches, the failure rate in clinical trials is considerably higher than that of proven pathways for new agents of existing antibiotic classes. Finding innovative antimicrobial entities is scientifically complex and less successful than drug discovery in other fields. The challenges include (i) in order to avoid single-step resistance, find compounds that have more than one binding site; (ii) discover molecules that penetrate the outer layers of Gram negative cell walls without being pumped out immediately by efflux pumps; and (iii) overcome toxicity problems due to the usual high concentrations of the antimicrobials required to kill bacteria (Ventola 2015b; Kennedy and Read 2018; Peterson and Kaur 2018).
Antimicrobial drug discovery: latest approaches
Historically, the most successful means of antibacterial discovery have been the screening of natural products obtained from microbial sources. To date, most of the antibiotics available on the market are natural or semi-synthetic/synthetic compounds derived from natural molecules (Simmons et al. 2010; Renwick et al. 2016; Parish 2019). Several approaches can be employed in the discovery of natural antimicrobials, such as diversity-based approaches (culturing the “uncultured” bacteria; co-culture; examine rare bacterial taxa and unusual habitats), ribosome engineering, genetic engineering, and genome mining, as developed below (Goodfellow and Fiedler 2010; Renwick et al. 2016; Wohlleben et al. 2016). These approaches are supported by recent advances in computational models and combinatorial chemistry to rationally discover new drugs and improve current antimicrobials (Pucci 2006; Simmons et al. 2010; Renwick et al. 2016; Blaskovich et al. 2017).
Natural antibiotics discovery: diversity-based approaches
Less than 1% of the microbes present in the environment can be cultivated in the laboratory. In support of the global effort to cultivate new microbial taxa, nutrient media and diffusion chambers have been successfully used in high-throughput cultivation for several microbes such as marine isolates (Amann et al. 1995; Nichols et al. 2010; Wohlleben et al. 2016). Soil Actinomycetes, particularly Streptomyces species, have been the source of most natural antibiotics in use nowadays. In recent years, after decades of exploiting terrestrial Streptomycetes, the search has shifted to rare Actinomycetes and other taxa of bacteria such as Cyanobacteria and Proteobacteria. Novel Actinomycetes species and their novel natural products have been discovered in deep ocean sediments, hyper-arid desert soils, and hot springs to name a few unusual habitats. Endophytic bacteria present in plant tissues and plant rhizospheres, and symbiotic bacteria such as the actinobacteria living in mutualistic association with fungal growing attine ants, bacterial-nematode associations, or even human commensals are being sourced for novel antibiotics. Goodfellow and Fiedler have outlined a bioprospecting strategy where species from extreme or unusual habitats are cultivated with selective isolation methods, novel taxa are recognized by dereplication, and then screened for natural products (Hosaka et al. 2009; Goodfellow and Fiedler 2010; Wohlleben et al. 2016). In the environment, microorganisms are rarely found in isolation. Several reports have shown that microorganisms that grow in co-culture with another species result in induction of antibiotic production, often related to previously silent gene clusters. Co-cultures can induce silent antibiotic genes because of competition (Hosaka et al. 2009; Wohlleben et al. 2016).
Ribosome engineering, genetic engineering and genome mining
Ribosome engineering is an approach to discover microbes with spontaneous mutations in their ribosome or RNA polymerase, through screening antibiotic-resistant mutants. Some selected mutants may have elevated secondary metabolite production or produce new series of natural products with interesting biological activities. A simple way to introduce ribosomal mutations is by exposing strains to aminoglycoside antibiotics. Strains that become resistant to these antibiotics harbour mutations in the ribosome. Resistance to other ribosome targeting antibiotics such as gentamicin or erythromycin could also stimulate antibiotic production (Hosaka et al. 2009; Zhu et al. 2019).
Eliminating genes for one secondary metabolite can stimulate the production of silent pathways. This was verified when ten new amexanthomycins were produced after the deletion of rifamycin synthesis genes in the Amycolatopsis mediterranei. Furthermore, by introducing or altering the expression of a heterologous global regulator, the secondary metabolome can change unpredictably. Many biosynthetic gene clusters include pathway-specific regulators that when overexpressed can increase the yield of the natural product or stimulate a previously silent cluster (e.g., production of stambomycin by Streptomyces ambofaciens) (Laureti et al. 2011; Li et al. 2018; Rajeev 2018).
Genome mining can be applied to predict the synthesis of new compounds. Genes synthesizing natural products are contained in biosynthetic gene clusters. Those clusters are modular, following an assembly line method with the number of modules correlating to the number of molecular building blocks incorporated in the product. Normally, there is a core cluster that models the product and adjacent genes that change the product such as acyltransferases and genes that confer self-resistance (Blin et al. 2017; Rajeev 2018). Genome mining tools can connect biosynthetic gene clusters to their corresponding natural products, being empowered to find the accessory tailoring domains and use the information to predict structural features and physicochemical properties of the final product. The most important of these are PRISM (PRediction Informatics for Secondary Metabolomes) (Skinnider et al. 2017) and antiSMASH (antibiotics and Secondary Metabolite Analysis SHell) (Blin et al. 2017; Rajeev 2018). Those computational tools can foresee the number of biosynthetic gene clusters in a selected genome and the structures of the natural products. Other important tools which are restricted to peptide natural products are Pep2Path (Medema et al. 2014), NRPQuest (for non-ribosomal peptides) (Mohimani et al. 2014a) and RiPPQuest (for ribosomal peptides) (Mohimani et al. 2014b).
Bioactive-guided screening and target-based drug discovery
Two main principles have been applied as the basis for effective drug discovery programmes: (i) target-oriented screening: aiming to identify compounds that are compatible with a known and validated molecular target; (ii) bioactive-guided screening: after the identification of the active substance, efforts are made to analyse the target and the mode of action of the compound (Goodfellow and Fiedler 2010; Jackson et al. 2018; Parish 2019).
Target-based drug discovery has a great potential. This approach starts with identifying compounds or pathways that are essential to the pathogen. Genome sequencing can help to identify unique targets specific to bacteria, providing high specificity over human cells, and targets can be selected that are either common to all bacteria or specific to certain species. After selection of the target, they are characterized, validated and screened against a panel of small molecules or chemical libraries to identify those that affect them. Then, the selected molecules are evaluated for their suitability as drugs (low toxicity, good bacterial penetration and bioavailability) and later attempts are made to improve their potency and pharmacokinetic properties (Simmons et al. 2010; Parish 2019). Structure-based drug discovery (SBDD) is an example for target-based drug discovery. An antimicrobial drug target should be essential have a unique function in the pathogen and exhibit an activity that can be altered by a small molecule. After, protocols for high-throughput screening and de novo design are used to establish new drugs and their antimicrobial structure–activity relationship (Simmons et al. 2010).
Antimicrobial peptides as promising new drugs
Antimicrobial peptides (AMPs) are small proteins usually composed of up to 50 amino acid residues, which, depending on their composition, size and conformation, can be arranged in different groups such as cationic peptides (largest), non-cationic peptides, aromatic peptides or those derived from oxygen-binding proteins (Vizioli and Salzet 2002). Apart from the peptide charge, the features mentioned above may be essential for their antimicrobial activity (Friedrich et al. 2000). AMPs have exhibited broad-spectrum activity against many Gram negative and Gram positive strains as well as fungi (Chung and Khanum 2017) and are produced by both animals and plants (Zasloff 2002). They have been used for the treatment of Candidiasis (Danesi et al. 2002) and have also been useful against replication of HIV1 by eliminating viral gene expression (Wachinger et al. 1998). Antimicrobial peptides are part of the innate immune defence of organisms (Peschel and Sahl 2006); they may present antibacterial, antifungal, antiparasitic and antiviral activity (Hancock and Diamond 2000). They are one of the strategies used by leukocytes to kill microorganisms after being ingested (Otto 2010). AMPs also have a signalling function and can activate immune cells (Yang et al. 1999). AMPs bind to the anionic bacterial surface and can integrate into the cytoplasmic membrane, making pores to kill the microorganism, since most of them are cationic (CAMPs) and show amphipathic properties (Brown and Hancock 2006). In humans, most AMPs belong to the beta defensin family (Ganz 2003) while the LL-37 peptide belongs to the cathelicidin family (Dürr et al. 2006).
Most of the naturally occurring antimicrobial peptides have rather long sequences, which can compromise their application as commercial drugs due to the high cost of protein production at industrial scale. Hence, research is focussing on identifying short active fragments as a base to develop new AMPs, using a range of molecular engineering strategies, which are the topic of this review. Short AMPs represent a chemical class of promising new drugs due to their robust biomimetic mechanisms of action, their relative ease of synthesis, and low production cost when compared to biologicals. With a clearer understanding of AMP pharmacokinetics and structural biology, these peptides are increasingly gaining attention as therapeutic agents (da Costa et al. 2015; Magana et al. 2020; Mookherjee et al. 2020).
Molecular targets and mechanisms of action involved in peptide antimicrobial activity
The bactericidal and/or bacteriostatic activity of AMPs primarily depends upon their ability to interact with bacterial membranes or cell walls, as a direct cell death mechanism or means to reach intracellular targets (Kang et al. 2014; da Costa et al. 2015; Zhang and Gallo 2016; Le et al. 2017; Mookherjee et al. 2020). Mechanisms of action that target the microbial membrane are amongst the preferred ones to develop new agents, as less likely to engender resistance (Kennedy and Read 2018). For these reasons, the interactions of antimicrobial peptides with the microbial lipid membrane have been the focus of intense research, with molecular mechanisms being unravelled. Lipopolysaccharides (LPS) are other essential components of the Gram negative bacterial cell membrane. Given the negative charge of LPS, cationic AMPs strongly interact with these compounds, which constitute another antimicrobial target of choice. Interestingly, immunomodulatory activity has been reported through the interaction of AMPs with LPS, and also with microbial nucleic acids or other immune ligands. Various microbial intracellular targets are emerging from fundamental research, with growing evidence that potent AMPs target multiple functions. Bacterial biofilms recently gained attention as extracellular AMP targets to prevent infection spread.
Lipid membrane
Lipid membrane disruption
Several modes of action have been suggested for bacterial membrane disruption by peptides, including carpet, barrel-stave, toroidal pore formations and aggregate mechanism (Chan et al. 2006; Kang et al. 2014; Bechinger and Gorr 2017; Nagarajan et al. 2018; Mookherjee et al. 2020) (Fig. 2). In the carpet model, the peptides align and accumulate parallel to the surface of the membrane until they reach a critical concentration, leading to changes in the membrane fluidity and/or reduction in membrane barrier properties resulting in membrane permeabilization. The membrane is disrupted in a detergent-like way, with no need of peptide insertion into the hydrophobic core of the bilayer. For the barrel-stave model, peptides interact laterally, resulting in the formation of a structure that behaves like a protein ion channel. In the toroidal pore model, the peptides interact only with the head groups of the lipids, affecting the local curvature of the membrane and resulting in the formation of high curvature peptide-lipid toroids. Another model is the aggregate mechanism, when peptides cross the membrane without causing significant membrane disruption. Once inside the cell, AMPs aim at intracellular targets (da Costa et al. 2015; Le et al. 2017; Mookherjee et al. 2020).

Membrane disruption molecular models and some intracellular targets of AMPs. Reproduced from Mookherjee et al. 2020. Copyright © 2020, Springer Nature Limited
Negative Gaussian curvature model
A body of research has focussed on the link between the mechanism of action of AMPs and the ability of these peptides to generate negative Gaussian curvature (or saddle-splay curvature) in the bacterial membrane, which is known to destabilize the membrane (Schmidt et al. 2011; Schmidt and Wong 2013; Lee et al. 2016). This effect is strongly linked to the membrane lipid composition—the presence of anionic lipids, and lipids of intrinsic negative curvature including phosphatidylethanolamine (PE)-based lipids and cardiolipin increase the susceptibility to AMPs (Som et al. 2009; Schmidt et al. 2011). PE is the main phospholipid found in the plasma membrane of Gram negative bacteria. It has been shown that the presence of these negative curvature lipids may reduce the energy required to form membrane pores associated with disruption of the bilayer by AMPs (Zimmerberg and Kozlov 2006). In contrast, cholesterol, which is more typical of eukaryotic membranes and acts to rigidify the lipid bilayer, appears to inhibit the effect of AMPs (Verly et al. 2008).
The role of negative Gaussian curvature in the AMP mechanism of action has also been demonstrated using peptide-mimicking phenylene ethynylenes (Yang et al. 2008). The ability of these compounds to induce permeation in bacterial membranes was again shown to require the presence of phosphatidylethanolamine (PE)-based lipids, which promote negative Gaussian curvature in membranes. A mutant PE-knockout strain of E. coli was shown to out-survive the wild type strain following addition of phenylene ethynylenes (Yang et al. 2008).
Lee et al. used a machine learning approach to predict whether a series of membrane-permeating α-helical peptides of various sequence homologies were likely to be antimicrobial (Lee et al. 2016). Sequence homologies were restricted to peptides 20–25 residues in length, with homology to known AMPs, and high α-helical content. A support vector machine (SVM)-based classifier was used to generate a metric σ based on the distance from the SVM hyperplane—this metric was not correlated with the peptide’s minimum inhibitory concentration, but instead with the ability of the peptide to generate negative Gaussian curvature in a lipid membrane. Small-angle x-ray scattering (SAXS) experiments on unilamellar vesicles following the addition of selected peptides showed the formation of coexisting bicontinuous cubic lipid phases of Pn3m and Im3m symmetry. These phases are based around a fundamental lipid bilayer structure with a non-positive Gaussian curvature (although they maintain zero mean curvature) (Lee et al. 2016).
Peptide secondary structure and membrane activity
Membrane active AMPs comprise peptides of varied secondary structures commonly classified into α-helical, β-sheet, or unstructured (random coil) conformations. Examples of α-helical antimicrobial peptides include magainin, temporins and melittin, which undergo a conformational change from unstructured in solution to an amphipathic helix when in interaction with the lipid membrane. All these peptides are cationic and disrupt the membrane of Gram positive and Gram negative bacteria. They also exhibit activity against fungal species (Lombardi et al. 2019). Membrane active β-sheet antimicrobial peptides can already be structured in solution, due to rigid conformations eventually stabilised by disulphide bridges, as for instance the natural host defense peptides defensins and their designed synthetic cyclic derivatives (Scudiero et al. 2015). Pioneering work by Ghadiri’s group showed that de novo designed cyclic D,L-peptides can form pores within membranes and exert antimicrobial activity (Fernandez-Lopez et al. 2001). Membrane active unstructured/extended antimicrobial peptides can adopt an amphipathic fold within membranes to induce disruption or simply be translocated to the intracellular space. The activity of such AMPs generally involves intracellular targets via specific amino acid motifs, including proline-rich AMPs originated from insects, or tryptophan and arginine rich peptides such as indolicidin (Le et al. 2017).
Toxicity: membrane composition selectivity
Due to differences in the membrane composition of different microbes/organism and cell types (Table 3), most AMPs can be cell-selective, by discriminating between the target membranes.
Membrane selectivity is governed by phospholipid composition and the resulting net charge of the membrane (Yeaman and Yount 2003), parameters which can differ significantly between prokaryotic and eukaryotic cells. Though not always the case, significant levels of antibacterial activity are achievable with minimal haemolysis and cytotoxicity, resulting in a high therapeutic index, which can be gauged by the ratio of minimum haemolytic concentration (MHC) to minimum inhibitory concentration (MIC).
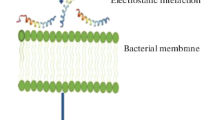
Numerous studies have found that antimicrobial peptide charge (which is typically cationic) plays a considerable role in the mechanism through which AMPs achieve cell selectivity. In mammalian cells, the outer lipid leaflet is typically comprised of zwitterionic phosphatidylcholine and sphingomyelin species, with anionic lipids distributed asymmetrically within the inner leaflet (Verkleij et al. 1973). In contrast, bacterial cell membranes contain an abundance of anionic phospholipids such as phosphatidylglycerol and cardiolipin. Other structural molecules, such as the negatively charged lipopolysaccharides in Gram negative bacteria, and teichoic acids in Gram positive bacteria, further contribute to the net negative charge of bacterial cell membranes. As a result, electrostatic interactions lead to preferential binding between AMPs and bacterial membranes (Christensen et al. 1988; Matsuzaki et al. 1997; Silvestro et al. 1997).
However, significant differences in a given peptide’s antimicrobial activity are observed when assayed against different bacterial species, pointing to separate mechanisms beyond simple membrane charge. Aside from fundamental structural differences between species, several studies have indicated that amongst anionic phospholipids, head group structure and hydrocarbon chain length further regulate peptide-membrane interactions. As assessed via DSC, the AMP protegrin-1 was found to influence the thermotropic phase behaviour of exposed DPPG vesicles significantly more than those comprised of DPPA, despite net negative charge of both lipid species being equal (Jing et al. 2005). It was additionally found that while the peptide bound to both DMPG and DSPG liposomes, penetration of protegrin-1 into the hydrophobic region of the bilayer only occurred for DMPG. This second observation was explained by the lipid tail packing, which is tighter in DSPG due to increased van der Waals forces between the longer hydrocarbon chains, resulting in an increased energy barrier for peptide insertion.
Further contributing to cell selectivity are the hydrophobic interactions occurring between the hydrophobic portions of amphipathic peptides and the cell membrane. It has been demonstrated that peptides with high MHC values exhibit strong interactions with membranes comprised of the zwitterionic phosphatidylcholine, while non-haemolytic peptides do not. With increasing insight into the parameters responsible for activity and selectivity, attempts have been made to modify or engineer existing antimicrobial peptides, with an aim to increase efficacy. It has been noted that increases in cationic charge up to approximately + 10 increase antimicrobial activity with minimal impact on haemolytic activity (Zelezetsky and Tossi 2006).
The effect on antimicrobial and haemolytic activity of peptide hydrophobicity, hydrophobic moment and angle subtended by the positively charged helix face were examined by Dathe et al. using a series of magainin and model peptides (Dathe et al. 1997). It was found that increases in each of these three parameters increased both activity against Gram positive bacteria and haemolytic activity to a similar extent. It was suggested that such increases in activity were driven by hydrophobic peptide–membrane interactions, particularly given that the erythrocyte membrane is largely comprised of zwitterionic PC and PE phospholipids. MIC values for the Gram positive S. aureus remained below the EC50 for haemolysis, and the model KLA peptides (+ 6 net charge) displayed lower MICs than magainins (+ 4 net charge), indicating that Gram positive activity remains dependent on peptide charge as well (Dathe et al. 1997). Activity against Gram negative bacteria showed little change as these parameters were altered, and it was reasoned that Gram negative activity was governed predominantly by electrostatic interactions. This observation would appear to explain the prokaryotic specificity of magainin, while more hydrophobic AMPs such as melittin exhibit activity against both bacterial and mammalian cell membranes (Dathe et al. 1997).
If one considers the action of AMPs as a combination of electrostatic and hydrophobic interactions with a membrane, then for more neutral membranes, where electrostatic peptide-lipid interactions are minimized, the hydrophobic effect plays a more predominant role regarding host cell toxicity. Hydrophobicity is an essential feature for AMP-membrane interactions. However, the increase of the hydrophobic interaction is strongly correlated with mammalian cell toxicity and loss of antimicrobial specificity. Therefore, many AMPs are moderately hydrophobic, to optimise selective toxicity towards microbial cell membranes. Selective antimicrobial activity results from a delicate balance of hydrophobic and electrostatic interactions between an antimicrobial peptide and the targeted membrane (Giuliani and Rinaldi 2011; Kang et al. 2014; Ong et al. 2014; Sierra et al. 2017).
Other cellular targets: lipopolysaccharides and intracellular targets
Lipopolysaccharides (LPS) are hallmark components of Gram negative bacteria outer membranes. LPS can be described as containing a lipid domain (lipid A or endotoxin) covalently bound to a “core” oligosaccharide and an antigen polysaccharide (O-antigen). Endotoxins exhibit pro-inflammatory activity via binding to innate immunity receptors, the Toll-like receptors present in animal immune cells. Hence, targeting LPS can result in Gram negative bactericidal activity via outer membrane destabilisation and in reducing endotoxin activity (Raetz and Whitfield 2002). Given the LPS amphipathic nature and negative charge, cationic amphipathic peptides are expected to significantly interact with LPS. This is the case for natural AMPs such as melittin, magainin and protegrin (Ding et al. 2003), but also synthetic peptides, which have been shown to destabilise the Gram negative bacterial outer membrane and to exhibit anti-toxin activity (Lakshminarayanan et al. 2016; Heinbockel et al. 2018).
However, endogenous antimicrobial peptides have also been shown to exhibit pro-inflammatory activity by complexation of immune ligands, including LPS and nucleic acids. Indeed, complexes of microbial ligands by endogenous AMPs are currently thought to be potentially immune-stimulant to signal microbial invasion to the immune system (Lee et al. 2019a). The interplay between the immune system and antimicrobial peptides is complex and is currently being investigated on a case by case basis, with excellent reviews available on the topic (Hilchie et al. 2013; Mookherjee et al. 2020).
Intracellular microbial targets identified for natural AMPs span from binding nucleic acids, the ribosome or protein chaperones, to inhibiting protein metabolism, cell division or cell wall synthesis. Over the past decade, an increasing number of natural AMPs have been shown to inhibit multiple microbial functions (Le et al. 2017; Mookherjee et al. 2020). For instance, it was evidenced that the short cationic peptide indolicidin, a cathelicidin with high tryptophan and arginine content, induces non-bactericidal membrane permeabilisation and binds intracellular duplex DNA via the peptide motif PWWP to inhibit microbial DNA replication and transcription (Ghosh et al. 2014). Although intracellular targets may provide an interesting addition or alternative for the molecular engineering of AMPs, few synthetic peptides have been specifically designed to target such microbial components to date and few of these peptide motifs have been identified (Le et al. 2017).
Targeting microbial colonies: biofilm components and bacterial adhesion
Bacterial biofilms are resilient drug-resistant communities of surface-adhered bacteria encapsulated within a sturdy extracellular matrix that acts as a physical barrier (Dunne Jr. 2002). Medical implants such as catheters are often compromised due to the formation of bacterial biofilms on the surface of the device; the rate of biofilm formation being affected by the material of the device (Quirynen and Bollen 1995; Garrett et al. 2008; Song et al. 2015). While this is typically remedied by simply replacing the medical device, an infection forming a resilient biofilm on a more permanent implant such as a prosthetic joint is disastrous. Biofilms may also form on living tissue such as P. aeruginosa colonisation of the lungs in immunocompromised cystic fibrosis patients (Lam et al. 1980). This is mainly due to the innate resistance of bacterial biofilms to antibiotics and disinfectants (Oie et al. 1996). Biofilm formation is also associated with several diseases and medical complications such as diabetic foot ulcer infections (Neut et al. 2011), ear infections, and urinary tract infections (Vestby et al. 2020).
Aside from the cells themselves, microbial biofilms are primarily composed of exopolysaccharides such as the teichoic acids of Gram positive Staphylococci and several from Gram negative species such as P. aeruginosa (Byrd et al. 2009; Colvin et al. 2012). These polysaccharides are involved in surface adhesion during the initial stage of biofilm formation (Flemming 1993). Proteins and extracellular microbial DNA are also major components of biofilm systems (Whitchurch et al. 2002; Flemming and Wingender 2010). Exopolysaccharides effect a local environment like that of an extracellular matrix, allowing for microbial cells to remain attached to the surface and resist shear forces (Donlan and Costerton 2002). Biofilm-residing microbial cells are less metabolically active than planktonic phase cells, allowing for perseverance in nutrient-deficient environments, and may contribute to higher resistance to antibiotics targeting metabolic pathways (Walters 3rd et al. 2003). In bacterial biofilms, the cells participate in communication via quorum sensing (QS) (Li and Tian 2012). QS arises from the bacterial generation and detection of signalling molecules called autoinducers (AIs), which are typically acyl homoserine lactones and other compounds biosynthesised from S-adenosylmethionine (Parveen and Cornell 2011; Papenfort and Bassler 2016). While Gram negative species utilise small molecule AIs, Gram positive bacteria such as S. aureus employ cyclic oligopeptides as autoinducer peptides (AIPs), which may be mimicked by synthetic peptides in order to block the quorum sensing pathway and the generation of virulence factors (Mayville et al. 1999; Thoendel et al. 2011; Zhu and Lau 2011).
The design of anti-biofilm peptides is a more nuanced approach compared to that of AMP inhibitors of planktonic cells as membrane disruption is not the sole mechanism sought after for anti-biofilm peptides (Sakala and Reches 2018; Dostert et al. 2019). However, it has been suggested that AMPs are more suitable as anti-biofilm agents compared to traditional antibiotics since the membrane remains a target irrespective of the metabolic state of the cell (Hurdle et al. 2011). The downside of this is of course that higher than usual concentrations of AMPs may be required, which may also induce haemolytic or cytotoxic effects. The distinct growth stages of biofilms may be targeted such as by inhibiting bacterial adhesion in the first place or interfering with growth and quorum sensing (Table 4).
Antimicrobial peptide self-assembly and activity
A correlation between peptide self-assembly and antimicrobial activity was suggested by the discovery that (i) amyloidogenic sequences can exhibit antimicrobial properties and (ii) endogenous antimicrobial proteins can form amyloid-like nanofibrillar aggregates (Sood et al. 2008; Jang et al. 2011; Kagan et al. 2012). Similarly, two molecular design approaches supported the interplay between peptide self-assembly and antimicrobial activity: (i) peptide sequences firstly designed to self-assemble into nanostructures, then discovered to exhibit antimicrobial activity; (ii) or the opposite, peptide sequences designed towards antimicrobial activity—essentially by membrane disruption—then discovered to self-assemble into hydrogels and/or supramolecular nanostructures (Tian et al. 2015; Shi et al. 2021). Although the requirement of amphipathic sequences is an obvious common physicochemical property shared by both molecular designs, the mechanisms of action by which peptide oligomers or nanostructures can damage microbial cells emerged relatively recently. Interestingly, the microbial membrane is not the only target that can be affected by peptide self-assembled species, as reviewed below.
Peptide amyloidogenic properties and antimicrobial activity
The amyloid beta (Aβ) peptide, which self-assembles into the so-called amyloid nanofibrillar aggregates in the brain of Alzheimer’s disease patients, was shown to exhibit broad-spectrum antimicrobial activity against bacteria and fungi (Soscia et al. 2010; Bourgade et al. 2016). Unexpectedly, the amyloid beta (Aβ) peptide was found to be of equivalent or greater antimicrobial potency as the human host defence peptide LL-37, suggesting unrecognised normal functions of the Aβ peptide in the innate immune response to microbial infections (Soscia et al. 2010). Mechanistic studies support that Aβ peptide antimicrobial activity is caused by the formation of ion channels within the microbial membranes, e.g. similar mechanism of action as its toxicity to brain cells (Kagan et al. 2012). A number of mechanisms of membrane disruption were unravelled to explain amyloid toxicity towards brain cells, including the insertion of toxic oligomers and membrane interactions with mature nanofibrils (Dharmadana et al. 2017). These molecular interactions can be hypothesised to apply to microbial membranes too (Fig. 3).

Main models of membrane disruption by amyloid oligomers. Reproduced from Dharmadana et al. 2017. Copyright © 2017, The Royal Society Publishing
Further supporting a correlation between self-assembly and antimicrobial activity, endogenous antimicrobial proteins and peptides were shown in vitro to undergo self-assembly into amyloid-like nanofibrillar aggregates, e.g. the human peptide LL-37 (Sood et al. 2008), protegrin-1 (Jang et al. 2011), or human α-defensin peptides (Chairatana et al. 2016). Two main mechanisms of action related to self-assembly were identified for these natural AMPs: either microbial membrane permeabilization by cytotoxic oligomers (Sood et al. 2008) and/or the entanglement of bacteria within “nanonets” formed by antimicrobial peptide nanostructures (Chairatana et al. 2016; Chairatana and Nolan 2017), which relates to biofilm targeting.
Membrane disruption upon peptide self-assembly
A few excellent reviews compile examples of self-assembling synthetic peptides that exhibit antimicrobial activity (Tian et al. 2015; Malekkhaiat Häffner and Malmsten 2018; Lombardi et al. 2019; Shi et al. 2021). The large majority of systems are amphipathic cationic peptides that affect the microbial membrane, either by lipid phase destabilisation or pore formation, eventually leading to cell lysis and death. In most cases, the peptide amphipathic design is responsible for intrinsic self-assembly, while hydrophobic and cationic charges ensure interactions with phospholipids and membrane insertion leading to antimicrobial activity. Hence, the amphipathic cationic design is sufficient to drive self-assembly while ensuring activity. By introducing rational variations in the amphipathic design in a single peptide system, various research groups succeeded in fine-tuning self-assembly into supramolecular structures of different morphologies and/or hydrogels while conserving antimicrobial activity (Shi et al. 2021). Recently, Gazit et al. exploited the self-assembly of the Phe-Phe dipeptide into precipitated nanotubes to create solid nanotube arrays on surfaces. Such nanostructured surfaces exhibit antibacterial activity by mechanical disruption of the bacterial membrane (Schnaider et al. 2017). This mechanism is directly inspired from the natural antibacterial properties of micro-spikes on dragonfly wings, which paved the way to diverse research on antimicrobial nanostructured surfaces of various chemical compositions (Elbourne et al. 2017).
Few studies report on the direct characterisation of bacterial membrane disruption of monomeric versus self-assembled peptides for comparable sequences. Although one would expect synergy between self-assembly and membrane disruption, current reports support subtlety and system specificity of this structure-function relationship. For instance, rationally designed cationic amphipathic “multidomain peptides” (MDPs) self-assemble into nanofibers, which have been shown to directly interact with lipid membranes as assembled peptides. However, when the nanofibers form hydrogels, antimicrobial activity is affected by rheological properties (Jiang et al. 2015). Interestingly, the same group reported an increase in melittin selective toxicity towards model bacterial membranes over mammalian membranes when assembled on MDP nanofibers. This result was assigned to the different conformations adopted by melittin in its monomeric and assembled forms when presented to the lipid membrane (Chen et al. 2019). Another example is the rationally designed family of ultrashort battacin-inspired peptides, with some peptides shown to self-assemble into nanostructures and hydrogels while other sequences remain as small oligomeric species in solution. These peptides act by targeting the microbial membrane. Although there is a clear difference in self-assembling properties for these related sequences, it appeared difficult to draw a direct correlation with the antimicrobial activity measured on various strains (Glossop et al. 2019). The understanding of the direct interplay between self-assembly and antimicrobial activity requires more research to enunciate generic rules.
Other targets than membranes affected by peptide self-assembly
Rational design of peptide sequences to self-assemble into toxic aggregates within bacterial cells was reported for a few examples, although this approach remains marginal and mechanistic details still need to be unravelled. A smart design involving dephosphorylation induced self-assembly of a tripeptide derivative into toxic intracellular nanofibers was for instance reported (Yang et al. 2007). Such a peptide antibiotic was shown to be bacteriostatic. Another approach to induce intracellular peptide aggregation focused on bacterial genome screening to identify aggregation-prone sequences (Bednarska et al. 2016). Interestingly, antibacterial peptide sequences were obtained, including against resistant strains and with good indications of specific toxicity. Intracellular aggregation resulted in bacterial cell death.
Extracellular peptide self-assembly, especially when resulting into hydrogels, can lead to bacteria entrapment and prevent cell mobility. This effect was reported for both natural antimicrobial sequences (Chairatana et al. 2016; Chairatana and Nolan 2017) and synthetic self-assembling peptides (Jiang et al. 2015). It is noteworthy that the matrix structure of bacterial biofilms relies on amyloid-like protein self-assembly into nanofibers. Hence, the current research effort on amyloid nanofibril self-assembly inhibition may lead to the discovery of compounds that chemically hinder biofilm formation (Andreasen et al. 2019).
The self-assembly of endogenous antimicrobial peptides in complex with nucleic acids (DNA or RNA) has been shown to control Toll-like receptors activation and subsequent pro-inflammatory responses. Interestingly, it was recently supported that structural specificity of these complexes might be essential for triggering an immune response: the peptides self-assemble into nanoscaffolds that present ordered nucleic acids to immune receptors (Schmidt et al. 2015; Lee et al. 2017b; Lee et al. 2019b). Although this recently emerged area of research is directly relevant to auto-immune diseases, it may hold promise for unravelling novel mechanisms that promote antimicrobial activity and/or modulate immunogenicity to support novel peptide designs.
Mechanisms of resistance to antimicrobial peptides
Microbial pathogens have developed distinct systems to resist the effect of antimicrobial peptides. These systems involve the destruction of antimicrobial peptides, change of their target and removal of peptides from their site of action (Gunn 2001; Joo and Otto 2015). Some of the defence mechanisms of bacteria versus AMPs include sequestration or degradation by secreted proteases (Mattiuzzo et al. 2014), impedance by biofilm formation and exopolymers (Bechinger and Gorr 2017), surface modification (Gunn 2001), cytoplasmic membrane alteration and export by multidrug efflux pumps (Nizet 2006). Excellent reviews have provided details on these mechanisms and are presented in Table 5 and Fig. 4 (Guilhelmelli et al. 2013; Joo et al. 2016) (Table 5, Fig. 4).

Mechanisms of bacterial resistance to AMPs. (1) Extracellular proteases perform proteolytic degradation; (2) sequestration can occur by extracellular matrix or extracellular proteins; (3) alanylated teichoic acids create electrostatic repulsion; (4) aminoacylated peptidoglycan also create electrostatic repulsion; (5) lack of lipid II-binding AMPs by pentapeptide alteration; (6) AMPs rejection by efflux pumps; (7) proteolytic cleavage by cytosolic protease after uptake by transporters; (8) sequestration or steric hindrance by O-antigen of LPS; (9) amine compound-added lipid A creates electrostatic repulsion; (10) lipid A acylation creates increased rigidity. Figure modified from Joo et al. 2016
Extracellular proteolytic degradation
Extracellular proteins are the first mechanism of defence and include proteases that degrade AMPs. Some of these proteases are SepA, aureolysin (metalloproteases), V8 protease and SepB that can protect against the human cathelicidin LL-37 (Schmidtchen et al. 2002; Barańska-Rybak et al. 2005; Frick et al. 2011). It has been observed that the interaction of SpeB and a host protein: alpha2M-binding (GRAB) creates a complex with a proteinase inhibitor bound on the surface that increases the resistance to LL-37 (Nyberg et al. 2004). Also, SpeB-degraded host proteoglycans release dermatan sulphate that neutralizes human alpha defensin HNP-1 (Schmidtchen et al. 2001) amongst others (Schmidtchen et al. 2002).
Within Gram negative bacteria, the omptin family are aspartate proteases located in the enterobacterial outer membrane (OM) that cleave LL-37 and cathelicidin-related antimicrobial peptide (CRAMP) (Stumpe et al. 1998; Guina et al. 2000; Galván et al. 2008). Metalloproteases are also important for Gram negative bacteria since several AMPs such as LL-37, human beta defensin HBD-1 and the porcine AMP protegrin-1 are neutralized by the metalloprotease ZapA (Belas et al. 2004). This metalloprotease is produced by P. mirabilis and is not specific to AMPs as it also degrades components of the cell matrix (actin, collagen) and antibodies (Belas et al. 2004). The degradation of alpha and beta defensins has also been observed in Porphyromonas gingivalis (Carlisle et al. 2009).
Similarly, various cationic AMPs (CAMPs) can be degraded by ZmpA and ZmpB produced by B. cenocepacia (Kooi and Sokol 2009). The degradation can also occur intracellularly with the help of transport proteins such as the ABC transporter that is encoded by the sapABCDFZ (Groisman et al. 1992; Mason et al. 2005). Moreover, an increased expression of genes in the sap operon is caused by the binding of the CAMPs by the SapA that leads to their proteolytic degradation (Mason et al. 2005; Mason et al. 2006; Shelton et al. 2011) as observed with H. influenzae with several AMPs such as LL-37, HBD-2, HBD-3 and HNP-1 and melittin (Mason et al. 2011). It is also important to note that inactivating AMPs highly depends on the structure of the target peptide. Linear structures are easier to degrade because the proteolytic areas are more exposed to cleavage than those with disulfide bonds. Bacteriocins such as lantibiotics contain sophisticated post-translational modification, presenting a resistance mechanism towards proteolytic degradation by the host or competing microorganisms (Bierbaum and Sahl 2009).
Another resistance mechanism is the sequestration of AMPs. Staphylokinase is an important extracellular AMP-sequestering molecule that targets alpha defensins (HNP1 and 2) (Bokarewa and Tarkowski 2004). Another streptococcal sequester is streptococcal inhibitor of complement (SIC). It protects S. pyogenes from defensins, LL-37 and the membrane attack complex (Åkesson et al. 1996; Frick et al. 2003).
Also, some streptococcal cell surface-attached proteins such as M1 and PilB from S. pyogenes and S. agalactiae respectively bind cathelicidins (LL-37 and CRAMP) providing resistance to AMPs (Maisey et al. 2008; Lauth et al. 2009).
Exopolymers and biofilms
A bacterial biofilm exhibits higher resistance to AMPs and antibiotics compared to planktonic bacteria (Nickel et al. 1985; Mah and O’Toole 2001). This is partially due to AMPs not being able to penetrate the biofilm matrix since EPS and capsular polysaccharides (CPS) can adsorb incoming antimicrobials, hinder their traversal into the biofilm, and limit their local concentration at the cells (Sugano et al. 2016; Singh et al. 2017). This property also applies especially to cationic AMPS (CAMPs), which have been found to bind to the anionic exopolysaccharide alginate from P. aeruginosa biofilms. This sequestering of CAMPs arises from the same mechanism by which they target anionic components of bacterial membranes. Negatively charged extracellular DNA (eDNA) is another major component of biofilms that may bind CAMPs (Mulcahy et al. 2008; Montanaro et al. 2011).
eDNA is important for biofilm regulation and is secreted or released by lysed cells during biofilm growth, allowing for horizontal gene transfer within the bacterial population and also acting as a potential source of nutrients (Spoering and Gilmore 2006; Flemming and Wingender 2010; Mulcahy et al. 2010; Okshevsky and Meyer 2015) It has been proposed that this sharing of genetic material may enhance antimicrobial resistance, adaptability, and pathogenicity in the biofilm (Sykes 2010; Vorkapic et al. 2016) Furthermore, the chelation of cations such as magnesium (Mg2+) by eDNA may trigger virulence responses in P. aeruginosa leading to AMP resistance (Mulcahy et al. 2008; Lewenza 2013; Wilton et al. 2016; Ibanez de Aldecoa et al. 2017).
Polysaccharide intercellular adhesin (PIA) provides AMP resistance in different forms: it is efficient against LL-37, cationic HDB-3 and anionic dermicidin (Vuong et al. 2004b; Wang et al. 2004b), its deacetylation increases its positive net charge and repels CAMPs (Vuong et al. 2004a), but is also crucial for biofilm formation and enhanced resistance (Heilmann et al. 1996; Rupp et al. 1999; Vuong et al. 2004b). The CPS of Klebsiella pneumoniae or P. aeruginosa impedes CAMPs by electrostatic trapping (Campos et al. 2004; Llobet et al. 2008). Other capsules such as M protein and the hyaluronic acid capsule of group A streptococci also sequester AMPs (Cole et al. 2010).
Bacteria (P. aeruginosa and S. pyogenes) can also exploit the host polysaccharide to sequester AMPs after degrading their host proteoglycan matrix with bacterial proteases (Schmidtchen et al. 2001). Exopolysaccharides play a major role in the resistance to LL-37 in P. aeruginosa and other lung pathogens (Herasimenka et al. 2005; Foschiatti et al. 2009). Also, in Pseudomonas species, alginate plays an important role in AMP resistance in biofilms (Hentzer et al. 2001). Alginate acts by mimicking the microbial membrane inducing changes in AMPs and prevents their diffusion since it binds and induces peptide aggregation (Chan et al. 2004; Chan et al. 2005). Poly-gamma-glutamic acid (PGA) exists only in Gram positive bacteria and coagulase-negative staphylococci and forms an extracellular capsule that protects from phagocytosis by leukocytes (Ogunleye et al. 2015). Although it protects from dermcidin, LL-37 and HBD-3 like PIA, it does not facilitate biofilm formation (Kocianova et al. 2005; Otto 2008).
For these reasons, eradication of matured biofilms by AMPs is a complicated task, especially in vivo, where there may be limitations to the concentration of AMP allowed in treatments within a therapeutic window. Recent efforts to overcome the antibiotic resistance of biofilms involve the use of DNase to counter the effects of eDNA. DNase has been reported to disperse or destabilise bacterial biofilms of several medically relevant pathogens including S. aureus, P. aeruginosa, E. coli and C. jejuni (Tetz and Tetz 2010; Kaplan et al. 2012; Baelo et al. 2015; Brown et al. 2015) This gives credence to the further development of potential co-administration treatments of DNases and AMPs as a means of overcoming the resistance granted to bacteria by the biofilm matrix and its components (Jones et al. 2013; Fleming and Rumbaugh 2017).
Surface modification
The critical molecules on the cell surface involved in the resistance to AMPs are the anionic polymers attached to the outermost cell surface, teichoic acids (TA) in Gram positive cell wall and lipopolysaccharides (LPS) in the Gram negative outer membrane. TA represent 60% of the cell wall in Gram positive bacteria and provide a negative cell charge (Kojima et al. 1985; Bera et al. 2007) but alanylation on free hydroxyls of the repeating sugars confers a positive charge to TA and this in turn decreases the attraction of CAMPs as observed with S. aureus (Peschel et al. 1999). D-alanylation also increases cell density suggesting a reduction of the permeability and electrostatic repulsion (Saar-Dover et al. 2012). Gram negative Bordetella pertussis uses D-alanylation via Dra (a Dlt homologue) to decrease the negative charge on the OM and increases resistance to LL-37, HNP-1, HNP-2 and polymyxin B (Taneja et al. 2013). Another mechanism involves the L-rhamnosylation of WTA in L. monocytogenes that increases the steric impediment in the cell wall (Carvalho et al. 2015).
Modification of lipid II (which is an important cell wall building block and peptidoglycan precursor) is also performed by many bacteria to evade AMPs. The modification consists in the replacement of the terminal D-alanine with D-lactate or D-serine to prevent vancomycin binding to the D-Ala-D-Ala dipeptide and blocking the transpeptidation required for cell wall synthesis (Kahne et al. 2005). Lipid II with D-lactate or D-serine is found on resistant strains conferring them a 1000-fold increased resistance to vancomycin (Bugg et al. 1991). Bacteriocins and human AMPs HNP-1 and HBD-3 also use lipid II for pore formation (Brötz et al. 1998) and block cell wall biosynthesis respectively (de Leeuw et al. 2010).
In Gram negative bacteria, LPS provides a negative charge (as TA in Gram positive bacteria) and its alteration is required for resistance to AMPs. Thus, amine containing molecules such as glycine, phosphoethanolamine (PEA) and 4-amino-4-deoxy-T-arabinose (Ara4N) increase the positive charge of LPS by modifying lipid A. Acinetobacter baumannii and Bordetella species perform the modification of lipid A with galactosamine or glucosamine (Llewellyn et al. 2012; Pelletier et al. 2013; Shah et al. 2014). The attachment of PEA onto phosphates in lipid A by S. typhimurium, Neisseria gonorrhoea and A. baumanii also decreases the anionic properties of LPS (Lee et al. 2004a; Lewis et al. 2013; Pelletier et al. 2013). Burkholderia and N. meningitidis constitutively express PEA-attached lipid A conferring them a higher resistance to AMPs (Tzeng et al. 2005; Loutet and Valvano 2011). The removal of anionic phosphate groups from lipid A provides the same effect of adding positive molecules to LPS; this is done by phosphatases in F. novicida (Wang et al. 2004a; Wang et al. 2006). In addition to the mentioned mechanisms, the O-antigen (outermost long polysaccharide chain in LPS) provides an extra barrier to Gram negative bacteria. A couple of studies have confirmed it is essential for AMP resistance in Brucella abortus and B. cenocepacia (Allen et al. 1998; Loutet et al. 2006).
Alteration of the cytoplasmic membrane structure
Since the cytoplasmic membrane is the major target of AMPs, bacteria modify it with different strategies. One of them is the amino acylation of the phosphatidyl glycerol (PG) head group, which masks anionic phosphates with cationic primary amines. This prevents CAMPs attraction to anionic head groups and results in electrostatic repulsion (Peschel et al. 2001). The protein responsible for this amino acylation is the multipeptide resistance factor (MprF) and was first described in S. aureus (Peschel et al. 2001). The C-terminal synthase domain of MprF uses PG and aminoacyl-tRNA as substrates, the resulting aminoacyl-PG is sent to the outer leaflet of the cytoplasmic membrane by MprF (Ernst et al. 2009). A different combination of substrates has been used by other bacteria such as Ala-PG in P. aeruginosa (Klein et al. 2009), Lys-PG and Lys-CL in Listeria monocytogenes (Thedieck et al. 2006) and Lys-PG and Ala-PG in Clostridium perfringes (Johnston et al. 2004). Lys-PG has also been reported in Gram negative bacteria such as Caulobacter crescentus (Jones and Smith 1979) or Rhizobium tropici (Sohlenkamp et al. 2007). Finally, the increase of saturated acyl chains can provide a higher membrane rigidity and higher resistance to nisin and pediocin in L. monocytogenes and E. faecalis respectively (Mazzotta and Montville 1997; Kumariya et al. 2015). Similarly, the addition of carotenoid phyloxanthin has been reported to stabilize the acyl chains in the membrane and increase its rigidity in S. aureus (Mishra et al. 2011).
Efflux pumps
Efflux pumps can remove the AMPs even when they have attached to the cytoplasmic membrane. They are energy-dependent transporters that expel not only AMPs but also other antibiotics (Poole 2007). Gram negative bacteria contain a resistance-nodulation-cell division (RND) family of transporters (Piddock 2006; Delmar et al. 2014). A typical AMP efflux pump of this family has three components: (1) inner membrane proton/ AMP antiporter, (2) an outer membrane (OM) transporter and (3) a periplasmic accessory protein that provides stability to the whole complex. Some of examples of efflux pumps in Gram negative bacteria include AcrAB-TolC in K. pneumoniae, which is efficient against polymyxin B and HNP-1, HBD-1 (human defensins), and VexAB-TolC of V. cholerae that also protects from polymyxin B (Bina et al. 2008). RosA, RosB in Y. enterocolotica and K. pneumoniae is a major facilitator superfamily (MFS) efflux pump that also confers resistance to polymyxin B (Bengoechea and Skurnik 2000; Padilla et al. 2010).
Within Gram positive bacteria, AMP-exporting efflux pumps are ATP-binding cassette (ABC) transporters that can secrete or produce immunity against newly synthesized AMPs (Gebhard 2012). NisT of Lactococcus lactis exports nisin (Qiao and Saris 1996) and Epi FEG of S. epidermidis exports epidermin and gallidermin (Otto et al. 1998). The BceAb type are another group of two-component ABC transporters that are active on lantibiotics, cyclic AMPs and defensins, glycopeptides and cathelicidin and have a wider range of action vs. AMPs (Gebhard 2012). The transporter VraFG in S. aureus provides resistance to several AMPs (nisin, bacitracin, vancomycin, indolicidin, LL-37 and HBD-3 (Li et al. 2007a; Falord et al. 2012; Gebhard 2012). The BceAb-type transporters have also been detected in B. subtilis, S. pneumoniae and other Gram positive bacteria (Becker et al. 2000; Ohki et al. 2003; Collins et al. 2010; Gebhard 2012). The MefE/Mel is a unique efflux pump in S. pneumoniae of the MFS and ABC families that confers resistance to LL-37 (Ambrose et al. 2005; Zähner et al. 2010).
Sensing systems
The AMP resistance mechanisms are regulated by sensors and regulators in order to save energy. The antimicrobial peptide sensor (Aps) was first observed in S. epidermidis and is also known as GraRs or GraRSX (Li et al. 2007b). It has three components: a membrane-bound AMP sensing histidine kinase (ApsS), DNA-binding response regulator (ApsR) and another component (ApsS). The expression of genes involved in AMP resistance mechanisms is regulated by the Aps and one of them is the dlt operon for TA alanylation, the mprF gene for PG lysinylation and the vraFG ABC transporter.
Systems homologous to Aps are also found in Gram positive pathogens such as Bacillus anthracis, Clostridium difficile, L. monocytogenes, S. pneumoniae and Sthaphylococcus haemolyticus (Li et al. 2007b). Other systems that sense and regulate resistance mechanisms are BceSR of B. subtilis, BraSRVraSR in S. aureus and LiaFSR in Streptococci (Suntharalingam et al. 2009).
Within Gram negative bacteria, the PhoPQ acts as a central regulator (Groisman et al. 1989; Miller et al. 1989). PhoPQ is located in the cytoplasmic membrane and is a histidine kinase sensor while PhoP is the corresponding DNA-binding response regulator. PhoPQ and PmrAB (another two-component system) regulate lipid A modifications involved with AMP resistance (Gunn et al. 1998; Herrera et al. 2010). Some PhoPQ homologous are found in several strains such as S. typhimurium, Y. pestis and Shigella flexneri (Moss et al. 2000; Oyston et al. 2000; Rebeil et al. 2004; Bader et al. 2005).
Molecular engineering approaches for antimicrobial peptide motif design
AMPs isolated from natural sources typically exhibit common sequence motifs: the peptides are 12–50 amino acid long, are highly cationic (net charge between + 2 and + 9) and amphipathic, with around 50% of hydrophobic residues in their sequence (Hancock and Sahl 2006). Starting from these general design rules, recent strategies for AMP molecular engineering include the use of high-throughput screening, machine learning, modelling tools and combinatorial libraries to aid in the identification and initial characterization of lead sequences (Kang et al. 2014; Zhang and Gallo 2016; Bechinger and Gorr 2017).
Bioinformatic tools
Considering the structural diversity of AMPs, bioinformatics has significant advantages in the computational enumeration of all possible peptide sequences (Travkova et al. 2017; Nagarajan et al. 2018). Online databases that contain large AMP libraries and several bioinformatic functions based on classification algorithms and machine learning have been built in the past decade to assist on the rational design of antimicrobial peptides, allowing the identification and optimization of new lead sequences in a time and cost-effective manner. A classic application of those tools is the use of artificial neural network (ANN), quantitative matrices (QM) and support vector machine (SVM) to predict antibacterial peptides (Wang and Wang 2004; Kang et al. 2014; Wang et al. 2016a; Zhang and Gallo 2016). Computer-assisted AMP design is useful for estimating the desired biological activity from the primary peptide structure. There are five types of prediction methods for AMPs; (1) prediction based on mature peptide sequences only (2) prediction based on precursor sequences only, (3) prediction based on both mature and precursor sequences, (4) prediction based on sequence similarity of the modifying enzymes and (5) prediction based on genomic information. The databases can also indicate related information such as specific target, antimicrobial activity and cytotoxicity (Hammam and Fliss 2010). Commonly used online databases are compiled in Table 6. Online peptide modelling tools are also available to assess physicochemical parameters, such as aggregation in aqueous media and pharmacokinetic profiles. Examples of modelling tools include (i)TANGO to predict amyloid cross-beta aggregation (Rousseau et al. 2006; Goldschmidt et al. 2010); (ii) ZipperDB to evaluate the peptide fibrillogenic propensity profile (Rousseau et al. 2006; Goldschmidt et al. 2010); (iii) GROMACS to perform molecular dynamics simulations (Pronk et al. 2013); and (iv) SWISSADME to predict pharmacokinetic properties, drug-like nature and medicinal chemistry friendliness (Daina et al. 2017).
Machine learning has recently been applied as a high-throughput molecular design approach for AMPs, fed by the increased depth of molecular understanding of AMP activity, especially membrane activity, and by the immense molecular dataset compiled by researchers over the past decades (Lee et al. 2017a; Lee et al. 2018a; Lee et al. 2018b). Although in its early ages, such high-throughput methods hold promises for a boost to the development of lead candidates for clinical translation of rationally designed minimal antimicrobial peptides.
Top-down molecular design
In top-down molecular design, an AMP sequence, generally from a natural source, is used as a starting template for systematic sequence truncation, amino acid substitution, hybridization and cyclization in order to obtain important information on the structural activity correlation and further design shorter AMPs with improved antimicrobial activity and selectivity (Ong et al. 2014; da Costa et al. 2015). Evolutionary conserved AMPs typically have long sequences of amino acids with redundancy in the sequence required for potent antimicrobial activities, thus trimming the natural AMP sequence allows a shorter active motif to be acquired, which can have reduced cost involved in the industrial scale production of the synthetic AMP for therapeutic use. Furthermore, the removal of naturally occurring peptide regions with undesired properties could also yield synthetic derivatives with reduced toxicity or immunogenicity (Ong et al. 2014; da Costa et al. 2015). Applying the principle of template modification, it is also possible to construct hybrid peptides composed by truncated fragments from multiple types of naturally occurring AMPs, so-called multidomain peptides. The resulting sequences could offer an effective strategy to capitalize on the desirable properties of the individual parent peptides. Such an approach typically involves the amalgamation of a sequence from a potent, but comparatively toxic AMP with a less active AMP having lower toxicity (Bianco et al. 2013; Ong et al. 2014; Zhu et al. 2017). Alanine scanning is another typical top-down approach to studying the structure-activity relationship (SAR) of an AMP to in turn optimize the peptide. Exemplary AMP alanine scanning studies include that of polymyxin B, which elucidated the SAR but also gave credence to optimized structures. More recently, alanine scanning on tridecaptin A1 (Kanazawa et al. 2009; Cochrane et al. 2014; Ballantine et al. 2019) revealed favourable modifications to retain activity but at a reduced synthetic cost, whereas alanine scanning of the linear battacin octapeptide revealed a truncated pentapeptide sequence that retained most of its antimicrobial activity (De Zoysa et al. 2015). Additional top-down designs include NSAID drug-peptide conjugates exhibiting antimicrobial, anti-inflammatory and self-assembling hydrogelator properties all in one due to the modification of a self-assembling peptide (McCloskey et al. 2016). Further techniques include the reorientation of a peptide structure without changing the amino acid composition, such as in the case of retro, inverso and retro-inverso peptide modifications (Chorev and Goodman 1995; Li et al. 2010; Parthsarathy et al. 2013).
Bottom-up molecular design
Bottom-up molecular design relies on the fundamental understanding of the impact of key chemical peptide features for antimicrobial activity (Frecer et al. 2004; Zelezetsky and Tossi 2006). A simplified approach is typically applied such as combining (i) cationic amino acids for electrostatic interactions with the microbial membranes and (ii) non-polar amino acids for insertion or permeation of the microbial membrane (Sforça et al. 2004; Matsuzaki 2009; Oddo and Hansen 2017). This approach is highly attractive for drug development as it can lead to the identification of the smallest motif or pharmacophore essential for antimicrobial effects, which could in turn increase the ease of optimization through systematic modifications to the short peptide sequence (Gordon et al. 2005; Ong et al. 2014) (Fig. 5).

Three-stage de novo peptide design workflow diagram. (1) design inputs and sequence selection; (2) fold specificity; (3) approximate binding affinity. Modified from Smadbeck et al. 2014
Combined with bioinformatics, a complete de novo peptide design framework usually consists in three stages: an optimization-based sequence selection, fold specificity calculation and approximate binding affinity calculation (Fig. 5) (Klepeis et al. 2004; Smadbeck et al. 2014). The first stage of the method is a sequence selection stage which uses a set of design inputs such as biologically relevant constraints in an integer linear optimization model to produce a rank ordered list of sequences with the lowest potential energy in a given template structure. The comparative model in peptides is based on the fact that sequence similarity implies structural similarity, thus the structure of an active peptide can be predicted by comparing the amino acid sequence to that of known native 3D structures. However, similar structures can be found for peptides with different sequences. The second stage collects the top sequences from the sequence selection stage and determines the specificity of candidates for the target. It determines how well-designed sequences fold into the desired template structure compared to the native sequence. The sequences with the top fold specificity values are then run through a computationally rigorous third stage to calculate the approximate binding affinity of the sequences to the target. Those peptides with the highest predicted binding affinity are then validated experimentally (Klepeis et al. 2004; Smadbeck et al. 2014).
Multivalent ligand design is another technique for AMP motif design, wherein multiple copies of an AMP are appended to a branched core. Branched multivalent peptide systems have also been investigated extensively in antimicrobial peptide design. Early study on multiple antigen peptide (MAP) systems was eventually translated into the field of antimicrobial peptide chemistry due to the inherent proteolytic stability of MAP ligands (Tam 1988; Bracci et al. 2003). This technique typically involves appending identical AMPs to a branched multi-lysine core to yield a multivalent peptide array and was utilised by Pini et al. to form a tetrabranched M33 antimicrobial peptide system with high proteolytic stability (Pini et al. 2010).
Antimicrobial peptide motifs and their rationale
A number of peptide motifs have been identified to promote antimicrobial activity and pharmacokinetic properties that aid molecular engineering translatable AMPs, as developed below.
Necessary peptide features for antimicrobial activity
Several studies have established that strong antimicrobial potencies can be obtained with engineered synthetic AMPs consisting of a cationic amino acid combined with a non-polar amino acid. Thus, AMPs generally contain positively charged amino acid residues (Dab, Orn, Lys and Arg) and hydrophobic residues (Phe, Trp, Tyr, Met) in a ratio varying from 1:1 to 1:2. Their primary structure is often characterized by a net positive charge of + 2 to + 9. Hydrophobic and non-charged peptides such as the aromatic peptide diphenylalanine have also shown antimicrobial activity due mainly to hydrophobic interactions (Strom et al. 2002; Klepeis et al. 2004; Ong et al. 2014).
It has been reported that increasing the positive charge of the peptide results in increasing antibacterial activity. However, there is a limit since a very high net charge can lead to an enhanced haemolytic propensity and a loss of antimicrobial activity. Additionally, the most common motif found in AMPs is the secondary structures; however, many of them also exist as unstructured conformers (Galdiero et al. 2013; Falanga et al. 2017).
Studies have suggested that a free N-terminal amino group and an amidated C-terminal on a peptide are correlated to a high antimicrobial activity. Thus, the balance between cationic residues, aromatic residues, an amidated C-terminal end and a free N-terminal amino group constitutes a highly efficient motif in antimicrobial peptides (Strom et al. 2002; Chen et al. 2010; Deslouches et al. 2013; Ong et al. 2014).
Choice of specific amino acids
Arginine and lysine are the most common cationic residues in AMP sequences as they are protonated under physiological conditions and have been found to be related strong antimicrobial activities. While the incorporation of arginine residues in AMPs confers greater antimicrobial activity, their presence is frequently associated with relatively higher haemolytic activities as compared to the use of lysine residues.
Due to their common occurrence in many naturally occurring AMPs such as cathelicidins, the combination of cationic arginine with hydrophobic tryptophan residues represents one of the most extensively investigated designs for synthetic AMPs. This combination is believed to be extremely efficient as the guanidinium side chain of arginine undergoes hydrogen bonding with negatively charged microbial membranes while participating in cationic–π interactions with the aromatic indole side chain of tryptophan, making AMP penetration into the hydrophobic lipid bilayer more energetically favoured. The bulky amphipathic character of the indole side chain in Trp is also believed to aid in the interfacial association of AMP with lipid bilayers and to mediate cohesive hydrophobic interactions with the lipid acyl chains during membrane penetration (Strom et al. 2002; Klepeis et al. 2004; Ong et al. 2014).
Secondary and supramolecular structures
In addition to the amphipathic content of the peptide primary sequence, their folding into distinct secondary structures either in the presence of bacterial membranes (e.g. cathelicidins and magainins) or constitutively (e.g. the defensins) is also a well-studied motif for antimicrobial activity. Generally, a basic repeating sequence pattern of amino acids that can be arranged into stabilized α-helical, β-sheet and/or hairpin structures are selected (Chen et al. 2010; Deslouches et al. 2013; Ong et al. 2014). Due to charge repulsion between cationic entities, linear AMPs often remain as free unstructured molecules in aqueous solutions, which facilitate their diffusion across the bacterial capsule and/or cell wall to the membrane lipid bilayers. Various peptide motifs were reported to enhance the stability of specific secondary structures, including for AMPs. For instance, the high proline content of insect AMPs has been correlated to a high helical propensity favouring membrane insertion (Chen et al. 2010; Ong et al. 2014).
Although self-assembly may inherently arise from amphipathic primary and/or secondary structures, simple aromatic motifs recently emerged as chemical tricks to induce the intrinsic formation of peptide supramolecular structures in solution via mainly pi-stacking interactions. Notable examples include (i) fluorenylmethoxycarbonyl (Fmoc) driving self-assembly when conjugated to peptides, generally resulting in the formation of hydrogels, and (ii) the amyloid-derived diphenylalanine motif inducing self-assembly into nanofibers or nanotubes when present in a sequence. These motifs were incorporated into numerous de novo designed peptides, including self-assembling AMPs (Schnaider et al. 2017; Glossop et al. 2019).
Non-proteinogenic amino acids and peptide termini
Unnatural residues, such as D-amino acids, improve proteolytic stability because proteases have poor affinity for unnatural side chains (Weinstock et al. 2012). Another common modification is N-alkylation of amino acids, typically N-methylation, which can improve pharmacokinetic properties. Studies on the cyclic peptide somatostatin yielded analogues with up to 10% improved oral bioavailability when N-methylated at certain points (Biron et al. 2008). While N-terminal acylation reduces the overall charge of the peptide, it improves proteolytic stability depending on the sequence and may also drastically improve antimicrobial activity of AMPs or grant activity to inactive sequences (Malina and Shai 2005; Radzishevsky et al. 2005; Glossop et al. 2018). Amidation is a common C-terminal modification employed to improve the antimicrobial potency of peptides in two ways, by increasing the net charge of the peptide and by enhancing proteolytic stability, thus improving half-life times in vivo (Stromstedt et al. 2009; Kumar et al. 2014; Mura et al. 2016). N-terminal acetylation and amidation may also enhance helicity by stabilising hydrogen bonding (Fairman et al. 1989; Stromstedt et al. 2009).
Fluorination
Fluorination is a frequent modification in medicinal chemistry and as such has been translated to peptide and protein engineering (Yoder and Kumar 2002). A small subset of AMPs feature naturally halogenated residues such as bromotryptophan in hagfish cathelicidins and hedistin (Uzzell et al. 2003; Tasiemski et al. 2007). Fluorination has been employed to enhance AMP activity through stabilised conformation and increased proteolytic resistance. Most notably, analogues of melittin substituting every leucine residue for 5,5,5-trifluoroleucine exhibited enhanced binding to DOPC vesicle membranes due to improved self-association in solution from hydrophobic interactions from trifluoroleucine (Niemz and Tirrell 2001). Furthermore, fluorinated buforin and magainin analogues were found to exhibit modest resistance to trypsin proteolytic degradation while retaining or improving in antimicrobial efficacy (Meng and Kumar 2007). However, this came at the cost of greater hemolysis in some fluorinated analogues. Studies on longer helical sequences by Lee et al. have also reported on the benefits of fluorination, in this case by hexafluoroleucine, in enhancing helical protein stability (Lee et al. 2004b). Further study of substitutions with hexafluoroleucine generated the magainin analogue fluorogainin-1, which resisted proteolytic degradation under the same conditions that degraded the unfluorinated parent peptide, while exhibiting improved antimicrobial activity towards K. pneumoniae and S. aureus (Gottler et al. 2008). Overall, fluorination has generally positive outcomes when designing synthetic AMPs.
Antimicrobial peptide lipidation
Lipopeptide motifs and antimicrobial activity
There are several examples of inactive peptides that gained antimicrobial activity upon conjugation of a hydrophobic acyl group (Malina and Shai 2005). Structure-activity studies were conducted by generating lipopeptides of varying fatty acid lengths (Nasompag et al. 2015; Glossop et al. 2018). Over improving lipid membrane insertion, fatty acid conjugation may enhance secondary structure characteristics: magainin-2 analogues showed tuneable helical, β-sheet, and oligomeric conformations depending on the size of attached fatty acid, from heptanoyl to palmitoyl in length (Avrahami and Shai 2002). Typically, longer fatty acids improve lipopeptide antimicrobial activity, but at a certain threshold also contribute to cytotoxicity. The acyl group may vary widely in structure from saturated fatty acyl moieties to unsaturated, branched or aromatic groups (Chooi and Tang 2010).
Prime examples of lipopeptide antibiotics that have undergone extensive structure-activity studies are the polymyxins and daptomycin. These lipopeptides are cyclic in structure and contain a majority of cationic and anionic charged side chains, respectively, and a fatty acyl chain in addition to hydrophobic residues. Both of these drugs have potent antimicrobial activity, even against multidrug-resistant strains (Dixon and Chopra 1986; Steenbergen et al. 2005; Zavascki et al. 2007). Structure-activity studies on polymyxins revealed that acyl chain length is also important for the spectrum of activity against pathogens. Polymyxin B analogues were found to have ideal acyl chains of 7–9 carbon length (Naoki et al. 2004). Studies on magainin analogues indicate that the fatty acyl moiety can modulate solution conformation and cell selectivity as well (Avrahami and Shai 2002). The primary sequence of the peptide component has the greatest influence over peptide hydrophobicity, stability, net charge and secondary structure. Lipopeptides may be cyclic or linear and may or may not have an exocyclic peptide region to which the fatty acyl chain is conjugated. An example of important primary peptide sequence is the hydrophobic dipeptide motif observed in polymyxin B (D-Phe-6, L-Leu-7) and E (D-Leu-6, L-Leu-7). The hydrophobic region provided by this dipeptide in the macrocycle is considered to contribute significantly to the amphipathicity of the lipopeptide as well as its biological interaction with Gram negative bacterial outer membranes (Pristovsek and Kidric 1999). The same phenylalanine and leucine combinations are seen in octapeptins (Velkov et al. 2017).
Charged amino acids are important determinants of lipopeptide surfactant and antimicrobial activity as they contribute to the hydrophilicity and electrostatic interactions of the peptide at physiological pH. Residues such as lysine and its relatives Dab or ornithine (2,5-diaminovaleric acid) are common cationic moieties found in natural antimicrobial lipopeptides such as the polymyxins, octapeptins and tridecaptins, which target anionic bacterial membranes, whereas daptomycin and its congeners feature anionic residues involved in their calcium-directed mechanisms (Kato et al. 1979; Baltz et al. 2005; Kanazawa et al. 2009; Velkov et al. 2017). Rarely will membrane-lytic lipopeptide antibiotics have neutral net charge at physiological pH such as in the case of iturin A, a potent antifungal agent and surfactant (Besson et al. 1984).
The incorporation of negatively charged residues into the peptide sequence is important for acidic lipopeptides such as daptomycin, amphomycins, tsushimycin, laspartomycins and friulimicins (Tanaka et al. 1977; Bunkoczi et al. 2005; Steenbergen et al. 2005; Borders et al. 2007; Schneider et al. 2009). These anionic lipopeptides often undergo conformational changes by interacting with calcium ions, effecting a stronger amphipathic structure and enhanced activity, whereas cationic lipopeptides, such as the polymyxins, have weaker antibiotic activity in vitro at higher concentrations of divalent cations such as Mg2+ (Daugelavicius et al. 2000; Ho et al. 2008).
Lipopeptide motifs for self-assembly into different colloidal morphologies
Self-assembly into various types of colloids can be programmed by conjugating a peptide sequence to an acyl chain. The resulting morphologies range from nanofibers and nanoribbons to micelles and vesicles (Cui et al. 2010; Webber et al. 2011; Hendricks et al. 2017). The influence of both the peptide sequence and acyl chain on the resulting colloidal morphology has been thoroughly investigated by Stupp’s group (Hendricks et al. 2017). For instance, a minimum of 10 carbon long acyl chain was found necessary to induce the hydrophobic effect needed to direct one-dimensional self-assembly into elongated morphologies. However, variation in the peptide residues closer to the acyl chain can still induce spherical micelles through altering hydrogen bonding capacity, while assembly into flat structures is favoured by alternating hydrophobic and hydrophilic residues in the peptide sequence (Hendricks et al. 2017). The conjugation of various bioactive peptide sequences to acyl chains has been reported to achieve the combination of nanostructure formation and biological/therapeutic activity in a single molecule (Silva et al. 2004; Webber et al. 2011; Hendricks et al. 2017).
Table 7 compiles the main peptide motifs reported to enhance antimicrobial activity and other properties important in the molecular engineering of such peptide drug molecules.
Translation opportunities of antimicrobial peptide–based formulations and biomaterials
Antimicrobial peptide drug discovery nowadays focuses on short synthetic peptides to overcome drawbacks inherent to naturally isolated AMPs, such as high production costs of long sequences and off-target cytotoxicity (Ong et al. 2014). A recent SWOT analysis identified critical weaknesses associated with the development of peptides as therapeutics: (1) chemically and physically instable, (2) prone to hydrolysis and oxidation, (3) tendency for aggregation, (4) short half-life and fast elimination, (5) not orally available and (6) low membrane permeability (Fosgerau and Hoffmann 2015). Besides the discovery and design of thousands of AMPs to date, very few AMP drugs have advanced to clinical development to date (Magana et al. 2020). In addition to molecular engineering principles developed in the previous sections, a number of formulation approaches are currently being investigated to overcome the challenge to reach clinical stage. An overview of peptide formulations is here provided with a focus on the latest advances in peptide and lipid nanotechnology, which are identified as the most promising formulations for translation (Magana et al. 2020).
Biomedical applications of antimicrobial peptide self-assembled nanostructures
Peptide hydrogels result from the self-assembly of amphipathic and/or aromatic rich sequences into elongated nanostructures, including nanofibers and nanotubes, which reticulate the aqueous media into a gel. In recent years, peptide hydrogels have gained attention for broad biomedical applications, from therapeutic formulations to tissue engineering (Mondal et al. 2020). The intrinsic self-assembly of antimicrobial peptide sequences into elongated nanostructures was reported for various sequences, opening the door to hydrogel applications for self-assembling AMPs (Shi et al. 2021). A remarkable example is the molecular engineering of short AMP sequences into self-assembling peptides that form hydrogels. For instance, the antibacterial sequence (KIGAKI)3-NH2 was incorporated into an engineered beta-hairpin sequence, resulting in hydrogels active against E. coli (Liu et al. 2013). Peptide sequences designed to self-assemble into nanostructured hydrogels also showed intrinsic antimicrobial properties. For instance, the synthetic sequence of MAX1 peptide was designed as an amphiphilic beta-hairpin of alternating lysine and valine residues, in order to induce self-assembly. This strategy resulted in the formation of rigid nanostructured hydrogels (Schneider et al. 2002). This elegantly designed hydrogel was shown to be active against both Gram negative and Gram positive bacteria when directly in contact with colonies, with a proposed mechanism of action involving disruption of the bacterial lipid membrane (Salick et al. 2007). Within the broad landscape of applications being currently explored for AMP hydrogels, one of the most promising ones appears to be their use as topical treatments, to cure skin infections or promote wound healing (Pfalzgraff et al. 2018).
In the current context of a viral pandemic, self-assembling AMPs could be of use as vaccine adjuvants or nanocarriers. Subunit vaccines, which are composed of synthetic antigen peptides or proteins, require adjuvants to stimulate an appropriate immune response. Amongst the several immunological adjuvants that have been developed so far (such as nanoparticles, liposomes, and polymers), an interesting category could be self-assembling AMP nanostructures (Hudalla et al. 2014; Wang et al. 2016b). Self-assembling AMPs could be used as delivery carriers for antigenic proteins and/or to boost immunogenicity, especially considering the immunomodulatory properties of some AMP sequences.
The deposition of arrays of solid-state peptide nanostructures was shown to provide antibacterial properties to the surface. The vertically aligned arrays of diphenylalanine nanotubes showed activity against E. coli, through their topological capacity to disrupt bacterial membranes (Schnaider et al. 2017). This original bionanomaterial deposition augurs well for a future use of self-assembling AMPs as medical coatings.
Formulations of antimicrobial peptides with lipid nanomaterials
The encapsulation of antimicrobial peptides in lipid nanoparticles (LNPs) confers two main advantages: Firstly, LNPs provide encapsulated peptides protection from enzymatic and chemical degradation. Such protection is otherwise difficult to achieve via modifications to the peptides chemical structure, due to the inherently fragile constituent peptide bonds. Secondly, increases in efficacy and bioavailability are possible via further functionalization of lipid nanoparticles, enabling the formulation of targeted delivery vehicles through techniques such as surface conjugation of antibodies, or the design of environment-responsive or triggered release particles. A wide range of lipid-based nanoparticle species have been investigated as delivery vehicles for antimicrobial peptides, ranging from liposomes to liquid crystalline cubic nanoparticles (Table 8).
Given the lipid bilayer structure inherent in these materials, it is unsurprising that membrane disruptive peptides have the potential to influence phase behaviour and structural parameters. Thus, careful characterization is necessary to determine any changes in material properties that come as a result of peptide loading and resulting peptide-membrane interactions. For example, the encapsulation of gramicidin A in cubic phase lipid materials has repeatedly revealed a shift towards structures with high negative curvature, such as hexosomes, with increasing peptide concentration (Liu and Caffrey 2005; Meikle et al. 2016; Meikle et al. 2017). The cationic charge of many AMPs must also be considered; electrostatic repulsion between membrane associated peptides appears to influence lipid phase behaviour as well as encapsulation efficiency (Boge et al. 2017; Meikle et al. 2017; Meikle et al. 2021). Manipulation of these interactions via the addition of anionic lipids to the cubosome formulation, or the inclusion of a charge screening buffer condition, can lead to favourable structural properties and increased peptide loading (Boge et al. 2017). Antimicrobial activity of cubosome encapsulated AMPs is seen to increase in some cases, decreasing in others. Work is currently underway exploring the fusion and uptake mechanisms involved in cubosome-bacteria interactions (Dyett et al. 2019; Meikle et al. 2020; Meikle et al. 2021).
Liposomes have long been utilized as nanocarriers for various drugs, as their structure allows for both hydrophobic and amphiphilic drug encapsulation within the membrane, while hydrophilic drugs can be contained within the liposome interior. They are also highly biocompatible and non-toxic; the first FDA approved nanodrug was Doxil, a liposomal doxorubicin formulation (Barenholz 2012). Design considerations include maximising both stability and peptide loading efficiency, achieved via careful selection of lipid formulations. Accordingly, lipids for which AMPs display selectivity or increased activity, such as those containing anionic head groups, are often avoided due to complete disruption of the liposomal bilayer.
A wide range of antimicrobial peptides have since been formulated with liposomes. In a recent study, DSPC/DSPE-PEG liposomes were used to encapsulate the AMP LL-37 (Ron-Doitch et al. 2016). Characterisation and activity experiments showed that the liposomal formulation possessed increased antiviral activity against HSV-1, with reduced toxicity over the free peptide. Cantor et al. have also demonstrated increased antimicrobial activity following encapsulation of the peptide alyteserin-1c in polymer-coated liposomes (Cantor et al. 2019).
Other lipid-based nanoparticle formulations are under investigation for AMP delivery, including phospholipid micelles, recently investigated for the encapsulation of polymyxin B (Brandenburg et al. 2012). In this study, encapsulation of the peptide in DSPE-PEG(2000) micelles was high; however, the in vitro activity against P. aeruginosa was significantly reduced compared to free peptide, presumably due to reduced electrostatic interactions between the encapsulated peptide and the bacterial membrane as a result of the PEG (200) coating.
Solid lipid nanoparticle solutions are colloidal dispersions of solid-state lipids, often stabilized with surfactants. They are formulated using lipids with increased melting points, such as mono- and tri-glycerides, steroids and fatty acids. Various production methods have been proposed, and parameters such as drug structure and solubility, final particle size and production conditions such as increased temperature must be considered. They have been used for the encapsulation of polymyxin B (Brandenburg et al. 2012), cyclosporin A (Zhang et al. 2000; Müller et al. 2006; Kim et al. 2009) and nisin (Prombutara et al. 2012). To increase wound healing, Fumika et al. fabricated solid lipid nanoparticles comprised of glyceryl monostearate and phosphatidylcholine co-encapsulated with two AMPs, LL-37 and serpin A1 (Fumakia and Ho 2016). The nanoparticles were found to accelerate wound healing and closure via BJ fibroblasts and keratinocytes, and the two peptides were found to possess synergistic antimicrobial activity against S. aureus and E. coli.
Conclusion
Worldwide health institutions generally project multidrug-resistant infections to cause several million deaths annually by 2050. In the search for new antimicrobial compounds and biomaterials to counter the alarming progression of such infections, antimicrobial peptides (AMPs) are considered as promising candidates by the research community. The increasing interest in these compounds is indicated by the exponential number of scientific articles published on AMPs, which led this review to cite close to 400 references to provide a global overview of this accelerating research area.
AMPs present the advantage of typically targeting microbial lipid membranes, a site of action less prone to resistance. AMPs often concomitantly act on other microbial sites, either intracellular and/or extracellular targets. Such multi-target mechanisms of action further limit the occurrence of resistance.
The recent years have seen the emergence of molecular engineering principles for the creation of optimised synthetic AMPs. Thanks to intense fundamental research in peptide design, it is now possible to engineer and optimise short peptide sequences that exhibit selective antimicrobial activity while limiting toxicity. Peptide molecular engineering is also now capable to optimise sequences towards biomaterial applications via controlling peptide self-assembling properties, including into nanostructured peptide hydrogels.
Challenges still remain to translate this global research effort to marketed products, especially improving pharmacokinetic properties of peptide-based therapeutics and continuing to decrease the costs of synthesis. However, recent advances in peptide synthesis and bio-nanotechnology augur well for overcoming these issues and providing AMP-based formulations and biomaterials with optimised pharmacokinetic properties at a reasonable cost.
References
Ahmad I, Perkins WR, Lupan DM, Selsted ME, Janoff AS (1995) Liposomal entrapment of the neutrophil-derived peptide indolicidin endows it with in vivo antifungal activity. Biochim Biophys Acta 1237:109–114
Åkesson P, Sjöholm AG, Björck L (1996) Protein SIC, a novel extracellular protein of Streptococcus pyogenes interfering with complement function. J Biol Chem 271:1081–1088
Alipour M, Suntres ZE, Halwani M, Azghani AO, Omri A (2009) Activity and interactions of liposomal antibiotics in presence of polyanions and sputum of patients with cystic fibrosis. PLoS One 4:e5724
Allen CA, Adams LG, Ficht TA (1998) Transposon-derived Brucella abortus rough mutants are attenuated and exhibit reduced intracellular survival. Infect Immun 66:1008–1016
Amann R, Ludwig W, Schleifer K (1995) Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol Rev 59:43–69
Ambrose KD, Nisbet R, Stephens DS (2005) Macrolide efflux in Streptococcus pneumoniae is mediated by a dual efflux pump and is erythromycin inducible. Antimicrob Agents Chemother 49:4203–4209
Andreasen M et al. (2019) Physical determinants of amyloid assembly in biofilm formation. mBio 10:e02279–02218
Andrei S, Droc G, Stefan G (2019) FDA approved antibacterial drugs: 2018-2019. Discoveries (Craiova) 7:e102
Aslam B et al (2018) Antibiotic resistance: a rundown of a global crisis. Infect Drug Resist 11:1645–1658
Avrahami D, Shai Y (2002) Conjugation of a magainin analogue with lipophilic acids controls hydrophobicity, solution assembly, and cell selectivity. Biochemistry 41:2254–2263
Bader MW et al (2005) Recognition of antimicrobial peptides by a bacterial sensor kinase. Cell 122:461–472
Baelo A et al (2015) Disassembling bacterial extracellular matrix with DNase-coated nanoparticles to enhance antibiotic delivery in biofilm infections. J Control Release 209:150–158
Ballantine RD, McCallion CE, Nassour E, Tokajian S, Cochrane SA (2019) Tridecaptin-inspired antimicrobial peptides with activity against multidrug-resistant Gram-negative bacteria. MedChemComm 10:484–487
Baltz RH, Miao V, Wrigley SK (2005) Natural products to drugs: daptomycin and related lipopeptide antibiotics. Natural Product Reports 22:717–741
Barańska-Rybak W, Sonesson A, Nowicki R, Schmidtchen A (2005) Glycosaminoglycans inhibit the antibacterial activity of LL-37 in biological fluids. J Antimicrob Chemother 57:260–265
Barenholz Y (2012) Doxil - the first FDA-approved nano-drug: lessons learned. J Control Release 160:117–134
Bechinger B, Gorr SU (2017) Antimicrobial peptides: mechanisms of action and resistance. J Dent Res 96:254–260
Becker S, Boger P, Oehlmann R, Ernst A (2000) PCR Bias in ecological analysis: a case study for quantitative Taq nuclease assays in analyses of microbial communities. Appl Environ Microbiol 66:4945–4953
Bednarska NG et al (2016) Protein aggregation as an antibiotic design strategy. Mol Microbiol 99:849–865
Belas R, Manos J, Suvanasuthi R (2004) Proteus mirabilis ZapA metalloprotease degrades a broad spectrum of substrates, including antimicrobial peptides. Infect Immun 72:5159–5167
Benech RO, Kheadr EE, Laridi R, Lacroix C, Fliss I (2002) Inhibition of Listeria innocua in cheddar cheese by addition of nisin Z in liposomes or by in situ production in mixed culture. Appl Environ Microbiol 68:3683–3690
Benech RO, Kheadr EE, Lacroix C, Fliss I (2003) Impact of nisin producing culture and liposome-encapsulated nisin on ripening of Lactobacillus added-cheddar cheese. J Dairy Sci 86:1895–1909
Bengoechea JA, Skurnik M (2000) Temperature-regulated efflux pump/potassium antiporter system mediates resistance to cationic antimicrobial peptides in Yersinia. Mol Microbiol 37:67–80
Bera A, Biswas R, Herbert S, Kulauzovic E, Weidenmaier C, Peschel A, Götz F (2007) Influence of wall teichoic acid on lysozyme resistance in Staphylococcus aureus. J Bacteriol 189:280–283
Bernegossi J, Calixto GMF, Sanches PRS, Fontana CR, Cilli EM, Garrido SS, Chorilli M (2016) Peptide KSL-W-loaded mucoadhesive liquid crystalline vehicle as an alternative treatment for multispecies oral biofilm. Molecules 21:37
Besson F, Peypoux F, Quentin MJ, Michel G (1984) Action of antifungal peptidolipids from Bacillus subtilis on the cell membrane of Saccharomyces cerevisiae. J Antibiot (Tokyo) 37:172–177
Bianco A, Venanzi M, Aleman C (2013) Peptide materials: from nanostructures to applications. John Wiley & Sons, Technology & Engineering
Bierbaum G, Sahl HG (2009) Lantibiotics: mode of action, biosynthesis and bioengineering. Curr Pharm Biotech 10:2–18
Bina XR, Provenzano D, Nguyen N, Bina JE (2008) Vibrio cholerae RND family efflux systems are required for antimicrobial resistance, optimal virulence factor production, and colonization of the infant mouse small intestine. Infect Immun 76:3595–3605
Biron E et al (2008) Improving oral bioavailability of peptides by multiple N-methylation: somatostatin analogues. Angew Chem Int Ed Engl 47:2595–2599
Blaskovich MA, Butler MS, Cooper MA (2017) Polishing the tarnished silver bullet: the quest for new antibiotics. Essays Biochem 61:103–114
Blin K et al (2017) antiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res 45:W36–W41
Boge L et al (2016) Lipid-based liquid crystals as carriers for antimicrobial peptides: phase behavior and antimicrobial effect. Langmuir 32:4217–4228
Boge L et al (2017) Cubosomes post-loaded with antimicrobial peptides: characterization, bactericidal effect and proteolytic stability. Int J Pharmaceutics 526:400–412
Boge L et al (2019a) Peptide-loaded cubosomes functioning as an antimicrobial unit against Escherichia coli. ACS Appl Mater Interf 11:21314–21322
Boge L et al (2019b) Cubosomes for topical delivery of the antimicrobial peptide LL-37. Eur J Pharm Biopharm 134:60–67
Bokarewa M, Tarkowski A (2004) Human α-defensins neutralize fibrinolytic activity exerted by staphylokinase. Thrombosis and Haemostasis 91:991–999
Borders DB et al (2007) Laspartomycin, an acidic lipopeptide antibiotic with a unique peptide core. J Nat Prod 70:443–446
Bourgade K, Dupuis G, Frost EH, Fülöp T (2016) Anti-viral properties of amyloid-β peptides. J Alzheimers Dis 54:859–878
Bracci L et al (2003) Synthetic peptides in the form of dendrimers become resistant to protease activity. J Biol Chem 278:46590–46595
Brandenburg KS, Rubinstein I, Sadikot RT, Önyüksel H (2012) Polymyxin B self-associated with phospholipid nanomicelles. Pharm Dev Tech 17:654–660
Brötz H, Josten M, Wiedemann I, Schneider U, Götz F, Bierbaum G, Sahl H-G (1998) Role of lipid-bound peptidoglycan precursors in the formation of pores by nisin, epidermin and other lantibiotics. Mol Microbiol 30:317–327
Brown KL, Hancock REW (2006) Cationic host defense (antimicrobial) peptides. Curr Op Immun 18:24–30
Brown HL, Hanman K, Reuter M, Betts RP, van Vliet AH (2015) Campylobacter jejuni biofilms contain extracellular DNA and are sensitive to DNase I treatment. Front Microbiol 6:699
Bugg TDH, Wright GD, Dutka-Malen S, Arthur M, Courvalin P, Walsh CT (1991) Molecular basis for vancomycin resistance in Enterococcus faecium BM4147: biosynthesis of a depsipeptide peptidoglycan precursor by vancomycin resistance proteins VanH and VanA. Biochemistry 30:10408–10415
Bunkoczi G, Vertesy L, Sheldrick GM (2005) Structure of the lipopeptide antibiotic tsushimycin. Acta Cryst D 61:1160–1164
Byrd MS et al (2009) Genetic and biochemical analyses of the Pseudomonas aeruginosa Psl exopolysaccharide reveal overlapping roles for polysaccharide synthesis enzymes in Psl and LPS production. Mol Microbiol 73:622–638
Campos MA, Vargas MA, Regueiro V, Llompart CM, Albertí S, Bengoechea JA (2004) Capsule polysaccharide mediates bacterial resistance to antimicrobial peptides. Infect Immun 72:7107–7114
Cantor S, Vargas L, Rojas AOE, Yarce CJ, Salamanca CH, Oñate-Garzón J (2019) Evaluation of the antimicrobial activity of cationic peptides loaded in surface-modified nanoliposomes against foodborne bacteria. Int J Mol Sci 20:680
Carlisle MD, Srikantha R, Brogden K (2009) Degradation of human α-and β-defensins by culture supernatants of Porphyromonas gingivalis strain 381. J Innate Immun 1:118–122
Carvalho F et al (2015) L-Rhamnosylation of Listeria monocytogenes wall teichoic acids promotes resistance to antimicrobial peptides by delaying interaction with the membrane. PLOS Pathogens 11:e1004919
Chairatana P, Nolan EM (2017) Human α-defensin 6: a small peptide that self-assembles and protects the host by entangling microbes. Acc Chem Res 50:960–967
Chairatana P, Chu H, Castillo PA, Shen B, Bevins CL, Nolan EM (2016) Proteolysis Triggers Self-Assembly and Unmasks Innate Immune Function of a Human α-Defensin Peptide. Chem Sci 7:1738–1752
Chan C, Burrows LL, Deber CM (2004) Helix induction in antimicrobial peptides by alginate in biofilms. J Biol Chem 279:38749–38754
Chan C, Burrows LL, Deber CM (2005) Alginate as an auxiliary bacterial membrane: binding of membrane-active peptides by polysaccharides. J Pep Res 65:343–351
Chan DI, Prenner EJ, Vogel HJ (2006) Tryptophan- and arginine-rich antimicrobial peptides: structures and mechanisms of action. Biochim Biophys Acta 1758:1184–1202
Chen C et al (2010) Antibacterial activities of short designer peptides: a link between propensity for nanostructuring and capacity for membrane destabilization. Biomacromolecules:402–411
Chen W et al (2019) Self-assembled peptide nanofibers display natural antimicrobial peptides to selectively kill bacteria without compromising cytocompatibility. ACS Appl Mater Interf 11:28681–28689
Cho J, Choi H, Lee J, Kim MS, Sohn HY, Lee DG (2013) The antifungal activity and membrane-disruptive action of dioscin extracted from Dioscorea nipponica. Biochim Biophys Acta 1828:1153–1158
Chooi YH, Tang Y (2010) Adding the lipo to lipopeptides: do more with less. Chem Biol 17:791–793
Chorev M, Goodman M (1995) Recent developments in retro peptides and proteins--an ongoing topochemical exploration. Trends Biotechnol 13:438–445
Christensen B, Fink J, Merrifield R, Mauzerall D (1988) Channel-forming properties of cecropins and related model compounds incorporated into planar lipid membranes. Proc Natl Acad Sci USA 85:5072–5076
Chung PY, Khanum R (2017) Antimicrobial peptides as potential anti-biofilm agents against multidrug-resistant bacteria. J Microbiol Immun Infect 50:405–410
Cochrane SA, Findlay B, Vederas JC, Ratemi ES (2014) Key Residues in octyl-tridecaptin A1 analogues linked to stable secondary structures in the membrane. ChemBioChem 15:1295–1299
Cole JN, Pence MA, von Köckritz-Blickwede M, Hollands A, Gallo RL, Walker MJ, Nizet V (2010) M Protein and hyaluronic acid capsule are essential for in vivo selection of covRS mutations characteristic of invasive serotype M1T1 group A streptococcus. mBio 1:e00191–00110
Collins B, Curtis N, Cotter PD, Hill C, Ross RP (2010) The ABC transporter AnrAB contributes to the innate resistance of Listeria monocytogenes to nisin, bacitracin, and various β-lactam antibiotics. Antimicrob Agents Chemother 54:4416–4423
Colvin KM et al (2012) The Pel and Psl polysaccharides provide Pseudomonas aeruginosa structural redundancy within the biofilm matrix. Environ Microbiol 14:1913–1928
Crofts TS, Gasparrini AJ, Dantas G (2017) Next-generation approaches to understand and combat the antibiotic resistome. Nat Rev Microbiol 15:422–434
Cui H, Webber MJ, Stupp SI (2010) Self-assembly of peptide amphiphiles: from molecules to nanostructures to biomaterials. Biopolymers 94:1–18
da Costa JP, Cova M, Ferreira R, Vitorino R (2015) Antimicrobial peptides: an alternative for innovative medicines? Appl Microbiol Biotechnol 99:2023–2040
da Silva Malheiros P, Sant’Anna V, Micheletto YMS, da Silveira NP, Brandelli A (2011) Nanovesicle encapsulation of antimicrobial peptide P34: physicochemical characterization and mode of action on Listeria monocytogenes. J Nanoparticle Res 13:3545–3552
Daina A, Michielin O, Zoete V (2017) SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci Rep 7:42717
Danesi R, Senesi S, Wout JWVT, JTv D, Lupetti A, Nibbering PH (2002) Antimicrobial peptides: therapeutic potential for the treatment of Candida infections. Exp Opi Invest Drugs 11:309–318
Dathe M et al (1997) Hydrophobicity, hydrophobic moment and angle subtended by charged residues modulate antibacterial and haemolytic activity of amphipathic helical peptides. FEBS Lett 403:208–212
Daugelavicius R, Bakiene E, Bamford DH (2000) Stages of polymyxin B interaction with the Escherichia coli cell envelope. Antimicrob Agents Chemother 44:2969–2978
Davies J, Davies D (2010) Origins and evolution of antibiotic resistance. Microbiol Mol Biol Rev 74:417–433
de la Fuente-Nunez C, Reffuveille F, Haney EF, Straus SK, Hancock RE (2014) Broad-spectrum anti-biofilm peptide that targets a cellular stress response. PLoS Pathogens 10:e1004152
de la Fuente-Nunez C et al (2015) D-enantiomeric peptides that eradicate wild-type and multidrug-resistant biofilms and protect against lethal Pseudomonas aeruginosa infections. Chem Biol 22:196–205
de Leeuw E et al (2010) Functional interaction of human neutrophil peptide-1 with the cell wall precursor lipid II. FEBS Lett 584:1543–1548
De Oliveira DMP et al (2020) Antimicrobial resistance in ESKAPE Pathogens. Clin Microbiol Rev 33:e00181–e00119
De Zoysa GH, Sarojini V (2017) Feasibility study exploring the potential of novel battacin lipopeptides as antimicrobial coatings. ACS Appl Mater Interfaces 9:1373–1383
De Zoysa GH, Cameron AJ, Hegde VV, Raghothama S, Sarojini V (2015) Antimicrobial peptides with potential for biofilm eradication: synthesis and structure activity relationship studies of battacin peptides. J Med Chem 58:625–639
Degnan AJ, Luchansky JB (1992) Influence of beef tallow and muscle on the antilisterial activity of pediocin AcH and liposome-encapsulated pediocin AcH. J Food Protect 55:552–554
Degnan AJ, Buyong N, Luchansky JB (1993) Antilisterial activity of pediocin AcH in model food systems in the presence of an emulsifier or encapsulated within liposomes. Int J Food Microbiol 18:127–138
Delmar JA, Su C-C, Yu EW (2014) Bacterial multidrug efflux transporters. Ann Rev Biophys 43:93–117
Deslouches B, Steckbeck JD, Craigo JK, Doi Y, Mietzner TA, Montelaro RC (2013) Rational design of engineered cationic antimicrobial peptides consisting exclusively of arginine and tryptophan, and their activity against multidrug-resistant pathogens. Antimicrob Agents Chemother 57:2511–2521
Dhand A, Snydman DR (2009) Mechanism of resistance in metronidazole. In: Mayers DL (ed) Antimicrobial drug resistance. Infectious Disease Humana Press, pp 223–227
Dharmadana D, Reynolds NP, Conn CE, Valéry C (2017) Molecular interactions of amyloid nanofibrils with biological aggregation modifiers: implications for cytotoxicity mechanisms and biomaterial design. Interface Focus 7:20160160
Ding L, Yang L, Weiss TM, Waring AJ, Lehrer RI, Huang HW (2003) Interaction of antimicrobial peptides with lipopolysaccharides. Biochemistry 42:12251–12259
Dixon RA, Chopra I (1986) Polymyxin B and polymyxin B nonapeptide alter cytoplasmic membrane permeability in Escherichia coli. J Antimicrob Chemother 18:557–563
Donlan RM, Costerton JW (2002) Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev 15:167–193
Dostert M, Belanger CR, Hancock REW (2019) Design and assessment of anti-biofilm peptides: steps toward clinical application. J Innate Immun 11:193–204
Dunne WM Jr (2002) Bacterial adhesion: seen any good biofilms lately? Clin Microbiol Rev 15:155–166
Dürr UHN, Sudheendra US, Ramamoorthy A (2006) LL-37, the only human member of the cathelicidin family of antimicrobial peptides. Biochim Biophys Acta 1758:1408–1425
Duval RE, Grare M, Demore B (2019) Fight against antimicrobial resistance: we always need new antibacterials but for right bacteria. Molecules:24
Dyett BP, Yu H, Strachan J, Drummond CJ, Conn CE (2019) Fusion dynamics of cubosome nanocarriers with model cell membranes. Nat Comm 10:1–13
Elbourne A, Crawford RJ, Ivanova EP (2017) Nano-structured antimicrobial surfaces: from nature to synthetic analogues. J Colloid Interface Sci 508:603–616
EMA (2020) Medicines. https://www.ema.europa.eu/en/medicines. Accessed 2020
Ernst CM et al (2009) The bacterial defensin resistance protein MprF consists of separable domains for lipid lysinylation and antimicrobial peptide repulsion. PLoS Pathogens 5:e1000660
Fairman R, Shoemaker KR, York EJ, Stewart JM, Baldwin RL (1989) Further studies of the helix dipole model: effects of a free alpha-NH3+ or alpha-COO- group on helix stability. Proteins 5:1–7
Falanga A, Nigro E, De Biasi MG, Daniele A, Morelli G, Galdiero S, Scudiero O (2017) Cyclic peptides as novel therapeutic microbicides: engineering of human defensin mimetics. Molecules 22
Falord M, Karimova G, Hiron A, Msadek T (2012) GraXSR proteins interact with the VraFG ABC transporter to form a five-component system required for cationic antimicrobial peptide sensing and resistance in Staphylococcus aureus. Antimicrob Agents Chemother 56:1047–1058
Farrell LJ, Lo R, Wanford JJ, Jenkins A, Maxwell A, Piddock LJV (2018) Revitalizing the drug pipeline: antibioticDB, an open access database to aid antibacterial research and development. J Antimicrob Chemother 73:2284–2297
FDA (2020) Drugs@FDA: FDA-approved drugs. Food and Drug Administration. https://www.accessdata.fda.gov/. Accessed 2020
Fernandez-Lopez S et al. (2001) Antibacterial agents based on the cyclic d,l-α-peptide architecture. Nature 412:452–455
Fleming D, Rumbaugh KP (2017) Approaches to dispersing medical biofilms. Microorganisms 5:15
Flemming HC (1993) Biofilms and environmental protection. Water Sci Tech 27:1–10
Flemming HC, Wingender J (2010) The biofilm matrix. Nat Rev Microbiol 8:623–633
Fong IW, Shlaes DM, Drlica K (2018) Antimicrobial resistance in the 21st century, Second edn. Springer, Cham, Switzerland
Foschiatti M, Cescutti P, Tossi A, Rizzo R (2009) Inhibition of cathelicidin activity by bacterial exopolysaccharides. Mol Microbiol 72:1137–1146
Fosgerau K, Hoffmann T (2015) Peptide therapeutics: current status and future directions. Drug Discov Today 20:122–128
Frecer V, Ho B, Ding JL (2004) De novo design of potent antimicrobial peptides. Antimicrob Agents Chemother 48:3349–3357
Frick I-M, Åkesson P, Rasmussen M, Schmidtchen A, Björck L (2003) SIC, a secreted protein of Streptococcus pyogenes that inactivates antibacterial peptides. J Biol Chem 278:16561–16566
Frick I-M, Nordin SL, Baumgarten M, Mörgelin M, Sørensen OE, Olin AI, Egesten A (2011) Constitutive and inflammation-dependent antimicrobial peptides produced by epithelium are differentially processed and inactivated by the commensal finegoldia magna and the pathogen Streptococcus pyogenes. J Immunol 187:4300–4309
Friedrich CL, Moyles D, Beveridge TJ, Hancock REW (2000) Antibacterial action of structurally diverse cationic peptides on gram-positive bacteria. Antimicrob Agents Chemother 44:2086–2092
Fumakia M, Ho EA (2016) Nanoparticles encapsulated with LL37 and serpin A1 promotes wound healing and synergistically enhances antibacterial activity. Mol Pharmaceutics 13:2318–2331
Galdiero S, Falanga A, Cantisani M, Vitiello M, Morelli G, Galdiero M (2013) Peptide-lipid interactions: experiments and applications. Int J Mol Sci 14:18758–18789
Galván EM, Lasaro MAS, Schifferli DM (2008) Capsular antigen fraction 1 and Pla modulate the susceptibility of Yersinia pestis to pulmonary antimicrobial peptides such as cathelicidin. Infect Immun 76:1456–1464
Ganz T (2003) Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol 3:710–720
Gao G et al (2011) The biocompatibility and biofilm resistance of implant coatings based on hydrophilic polymer brushes conjugated with antimicrobial peptides. Biomaterials 32:3899–3909
Garrett TR, Bhakoo M, Zhang Z (2008) Bacterial adhesion and biofilms on surfaces. Prog Nat Sci 18:1049–1056
Gebhard S (2012) ABC transporters of antimicrobial peptides in Firmicutes bacteria – phylogeny, function and regulation. Mol Microbiol 86:1295–1317
Getahun H, Smith I, Trivedi K, Paulin S, Balkhy HH (2020) Tackling antimicrobial resistance in the COVID-19 pandemic. Bulletin of the World Health Organization 98:442–442A
Ghosh A et al (2014) Indolicidin targets duplex DNA: structural and mechanistic insight through a combination of spectroscopy and microscopy. ChemMedChem 9:2052–2058
Giuliani A, Rinaldi AC (2011) Beyond natural antimicrobial peptides: multimeric peptides and other peptidomimetic approaches. Cell Mol Life Sci 68:2255–2266
Glossop HD, Pearl E, De Zoysa GH, Sarojini V (2018) Chapter eleven - linear analogues of the lipopeptide battacin with potent in vitro activity against S. aureus. In: Donev R (ed) Advances in protein chemistry and structural biology, vol 112. Academic Press, pp 385–394
Glossop HD et al (2019) Battacin-inspired ultrashort peptides: nanostructure analysis and antimicrobial activity. Biomacromolecules 20:2515–2529
Goldschmidt L, Teng PK, Riek R, Eisenberg D (2010) The amylome, all proteins capable of forming amyloid-like fibrils. Proc Natl Acad Sci USA 107:3487–3492
Gomaa AI, Martinent C, Hammami R, Fliss I, Subirade M (2017) Dual coating of liposomes as encapsulating matrix of antimicrobial peptides: development and characterization. Front Chem 5:103
Gontsarik M, Buhmann MT, Yaghmur A, Ren Q, Maniura-Weber K, Salentinig S (2016) Antimicrobial peptide-driven colloidal transformations in liquid-crystalline nanocarriers. J Phys Chem Lett 7:3482–3486
Goodfellow M, Fiedler HP (2010) A guide to successful bioprospecting: informed by actinobacterial systematics. Antonie Van Leeuwenhoek 98:119–142
Gordon YJ, Romanowski EG, McDermott AM (2005) A review of antimicrobial peptides and their therapeutic potential as anti-infective drugs. Curr Eye Res 30:505–515
Gottler LM, Lee H-Y, Shelburne CE, Ramamoorthy A, Marsh ENG (2008) Using fluorous amino acids to modulate the biological activity of an antimicrobial peptide. ChemBioChem 9:370–373
Groisman EA, Chiao E, Lipps CJ, Heffron F (1989) Salmonella typhimurium phoP virulence gene is a transcriptional regulator. Proc Natl Acad Sci USA 86:7077–7081
Groisman EA, Parra-Lopez C, Salcedo M, Lipps CJ, Heffron F (1992) Resistance to host antimicrobial peptides is necessary for Salmonella virulence. Proc Natl Acad Sci USA 89:11939–11943
Guilhelmelli F, Vilela N, Albuquerque P, Derengowski L, Silva-Pereira I, Kyaw C (2013) Antibiotic development challenges: the various mechanisms of action of antimicrobial peptides and of bacterial resistance. Front Microbiol 4:353
Guina T, Yi EC, Wang H, Hackett M, Miller SI (2000) A PhoP-regulated outer membrane protease of Salmonella enterica serovar typhimurium promotes resistance to alpha-helical antimicrobial peptides. J Bacteriol 182:4077–4086
Gunn JS (2001) Bacterial modification of LPS and resistance to antimicrobial peptides. J Endotoxin Res 7:57–62
Gunn JS, Lim KB, Krueger J, Kim K, Guo L, Hackett M, Miller SI (1998) PmrA–PmrB-regulated genes necessary for 4-aminoarabinose lipid A modification and polymyxin resistance. Mol Microbiol 27:1171–1182
Hammam R, Fliss I (2010) Current trends in antimicrobial agent research: chemo- and bioinformatics approaches. Drug Discov Today 15:540–546
Hammami R, Hamida JB, Vergoten G, Fliss I (2008) PhytAMP: a database dedicated to plant antimicrobial peptides. Nucleic Acids Res 37:D963–D968
Hancock RE, Diamond G (2000) The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol 8:402–410
Hancock RE, Sahl HG (2006) Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat Biotechnol 24:1551–1557
Heilmann C, Schweitzer O, Gerke C, Vanittanakom N, Mack D, Götz F (1996) Molecular basis of intercellular adhesion in the biofilm-forming Staphylococcus epidermidis. Mol Microbiol 20:1083–1091
Heinbockel L et al (2018) Inhibition of Lipopolysaccharide- and Lipoprotein-induced inflammation by antitoxin peptide Pep19–2.5. Front Immunol 9:1704–1704
Hendricks MP, Sato K, Palmer LC, Stupp SI (2017) Supramolecular assembly of peptide amphiphiles. Acc Chem Res 50:2440–2448
Hentzer M, Teitzel GM, Balzer GJ, Heydorn A, Molin S, Givskov M, Parsek MR (2001) Alginate overproduction affects Pseudomonas aeruginosa biofilm structure and function. J Bacteriol 183:5395–5401
Herasimenka Y, Benincasa M, Mattiuzzo M, Cescutti P, Gennaro R, Rizzo R (2005) Interaction of antimicrobial peptides with bacterial polysaccharides from lung pathogens. Peptides 26:1127–1132
Herrera CM, Hankins JV, Trent MS (2010) Activation of PmrA inhibits LpxT-dependent phosphorylation of lipid A promoting resistance to antimicrobial peptides. Mol Microbiol 76:1444–1460
Hilchie AL, Wuerth K, Hancock RE (2013) Immune modulation by multifaceted cationic host defense (antimicrobial) peptides. Nat Chem Biol 9:761–768
Ho SW et al (2008) Effect of divalent cations on the structure of the antibiotic daptomycin. Eur Biophys J 37:421–433
Hosaka T et al (2009) Antibacterial discovery in actinomycetes strains with mutations in RNA polymerase or ribosomal protein S12. Nat Biotechnol 27:462–464
Hudalla GA, Sun T, Gasiorowski JZ, Han H, Tian YF, Chong AS, Collier JH (2014) Gradated assembly of multiple proteins into supramolecular nanomaterials. Nat Mater 13:829–836
Hurdle JG, O'Neill AJ, Chopra I, Lee RE (2011) Targeting bacterial membrane function: an underexploited mechanism for treating persistent infections. Nat Rev Microbiol 9:62–75
Ibanez de Aldecoa AL, Zafra O, Gonzalez-Pastor JE (2017) Mechanisms and regulation of extracellular DNA release and its biological roles in microbial communities. Front Microbiol 8:1390
Jackson N, Czaplewski L, Piddock LJV (2018) Discovery and development of new antibacterial drugs: learning from experience? J Antimicrob Chemother 73:1452–1459
Jang H, Arce FT, Mustata M, Ramachandran S, Capone R, Nussinov R, Lal R (2011) Antimicrobial protegrin-1 forms amyloid-like fibrils with rapid kinetics suggesting a functional link. Biophys J 100:1775–1783
Jiang L, Xu D, Sellati TJ, Dong H (2015) Self-assembly of cationic multidomain peptide hydrogels: supramolecular nanostructure and rheological properties dictate antimicrobial activity. Nanoscale 7:19160–19169
Jing W, Prenner EJ, Vogel HJ, Waring AJ, Lehrer RI, Lohner K (2005) Headgroup structure and fatty acid chain length of the acidic phospholipids modulate the interaction of membrane mimetic vesicles with the antimicrobial peptide protegrin-1. J Pep Sci 11:735–743
John T, Thomas T, Abel B, Wood BR, Chalmers DK, Martin LL (2017) How kanamycin A interacts with bacterial and mammalian mimetic membranes. Biochim Biophys Acta 1859:2242–2252
Johnston NC, Baker JK, Goldfine H (2004) Phospholipids of Clostridium perfringens: a reexamination. FEMS Microbiol Lett 233:65–68
Jones DE, Smith JD (1979) Phospholipids of the differentiating bacterium Caulobacter crescentus. Canadian J Biochem 57:424–428
Jones EA, McGillivary G, Bakaletz LO (2013) Extracellular DNA within a nontypeable Haemophilus influenzae-induced biofilm binds human beta defensin-3 and reduces its antimicrobial activity. J Innate Immun 5:24–38
Joo H-S, Otto M (2015) Mechanisms of resistance to antimicrobial peptides in staphylococci. Biochim Biophys Acta 1848:3055–3061
Joo H-S, Fu C-I, Otto M (2016) Bacterial strategies of resistance to antimicrobial peptides. Philos Trans R Soc Lond B Biol Sci 371:20150292
Kagan BL, Jang H, Capone R, Teran Arce F, Ramachandran S, Lal R, Nussinov R (2012) Antimicrobial properties of amyloid peptides. Mol Pharm 9:708–717
Kahne D, Leimkuhler C, Lu W, Walsh C (2005) Glycopeptide and lipoglycopeptide antibiotics. Chem Rev 105:425–448
Kanazawa K, Sato Y, Ohki K, Okimura K, Uchida Y, Shindo M, Sakura N (2009) Contribution of each amino acid residue in polymyxin B(3) to antimicrobial and lipopolysaccharide binding activity. Chem Pharm Bull (Tokyo) 57:240–244
Kang SJ, Park SJ, Mishig-Ochir T, Lee BJ (2014) Antimicrobial peptides: therapeutic potentials. Expert Rev Anti Infect Ther 12:1477–1486
Kang X, Dong F, Shi C, Zheng H et al (2019) DRAMP 2.0, an updated data repository of antimicrobial peptides. Scientific Data 6:148
Kaplan JB, LoVetri K, Cardona ST, Madhyastha S, Sadovskaya I, Jabbouri S, Izano EA (2012) Recombinant human DNase I decreases biofilm and increases antimicrobial susceptibility in staphylococci. J Antibiot (Tokyo) 65:73–77
Kato T, Sakazaki R, Hinoo H, Shoji J (1979) The structures of tridecaptins B and C (studies on antibiotics from the genus Bacillus. XXV). J Antibiot (Tokyo) 32:305–312
Kennedy DA, Read AF (2018) Why the evolution of vaccine resistance is less of a concern than the evolution of drug resistance. Proc Natl Acad Sci U S A 115:12878–12886
Kim ST et al (2009) Topical administration of cyclosporin A in a solid lipid nanoparticle formulation. Die Pharmazie 64:510–514
Klein S et al (2009) Adaptation of Pseudomonas aeruginosa to various conditions includes tRNA-dependent formation of alanyl-phosphatidylglycerol. Mol Microbiol 71:551–565
Klepeis JL, Floudas CA, Morikis D, Tsokos CG, Lambris JD (2004) Design of peptide analogs with improved activity using a novel de novo protein design approach. Ind Eng Chem Res 43:3817–3826
Kocianova S, Vuong C, Yao Y, Voyich JM, Fischer ER, DeLeo FR, Otto M (2005) Key role of poly-γ-dl-glutamic acid in immune evasion and virulence of Staphylococcus epidermidis. J Clin Invest 115:688–694
Kojima N, Araki Y, Ito E (1985) Structure of the linkage units between ribitol teichoic acids and peptidoglycan. J Bacteriol 161:299–306
Kooi C, Sokol PA (2009) Burkholderia cenocepacia zinc metalloproteases influence resistance to antimicrobial peptides. Microbiol (Reading) 155:2818–2825
Kumar D, Eipper BA, Mains RE (2014) Amidation. In: Reference module in biomedical sciences. Elsevier, Amsterdam
Kumariya R, Sood SK, Rajput YS, Saini N, Garsa AK (2015) Increased membrane surface positive charge and altered membrane fluidity leads to cationic antimicrobial peptide resistance in Enterococcus faecalis. Biochim Biophys Acta 1848:1367–1375
Lai J, Lu Y, Yin Z, Hu F, Wu W (2010) Pharmacokinetics and enhanced oral bioavailability in beagle dogs of cyclosporine A encapsulated in glyceryl monooleate/poloxamer 407 cubic nanoparticles. Int J Nanomed 5:13
Lakshminarayanan R et al (2016) Branched peptide, B2088, disrupts the supramolecular organization of lipopolysaccharides and sensitizes the Gram-negative bacteria. Sci Rep 6:25905
Lam J, Chan R, Lam K, Costerton JW (1980) Production of mucoid microcolonies by Pseudomonas aeruginosa within infected lungs in cystic fibrosis. Infect Immun 28:546–556
Lange CF, Hancock RE, Samuel J, Finlay WH (2001) In vitro aerosol delivery and regional airway surface liquid concentration of a liposomal cationic peptide. J Pharm Sci 90:1647–1657
Laridi R, Kheadr E, Benech R-O, Vuillemard J, Lacroix C, Fliss I (2003) Liposome encapsulated nisin Z: optimization, stability and release during milk fermentation. Int Dairy J 13:325–336
Laureti L, Song L, Huang S, Corre C, Leblond P, Challis GL, Aigle B (2011) Identification of a bioactive 51-membered macrolide complex by activation of a silent polyketide synthase in Streptomyces ambofaciens. Proc Natl Acad Sci U S A 108:6258–6263
Lauth X et al (2009) M1 Protein allows group A streptococcal survival in phagocyte extracellular traps through cathelicidin inhibition. J Innate Immun 1:202–214
Le C-F, Fang C-M, Sekaran SD (2017) Intracellular targeting mechanisms by antimicrobial peptides. Antimicrob Agents Chemother 61:e02340–e02316
Lee H, Hsu F-F, Turk J, Groisman EA (2004a) The PmrA-regulated pmrC gene mediates phosphoethanolamine modification of lipid A and polymyxin resistance in Salmonella enterica. J Bacteriol 186:4124–4133
Lee KH, Lee HY, Slutsky MM, Anderson JT, Marsh EN (2004b) Fluorous effect in proteins: de novo design and characterization of a four-alpha-helix bundle protein containing hexafluoroleucine. Biochemistry 43:16277–16284
Lee EY, Fulan BM, Wong GC, Ferguson AL (2016) Mapping membrane activity in undiscovered peptide sequence space using machine learning. Proc Natl Acad Sci U S A 113:13588–13593
Lee EY, Lee MW, Fulan BM, Ferguson AL, Wong GCL (2017a) What can machine learning do for antimicrobial peptides, and what can antimicrobial peptides do for machine learning? Interface Focus 7:20160153
Lee EY, Takahashi T, Curk T, Dobnikar J, Gallo RL, Wong GCL (2017b) Crystallinity of double-stranded RNA-antimicrobial peptide complexes modulates toll-like receptor 3-mediated inflammation. ACS Nano 11:12145–12155
Lee EY, Wong GCL, Ferguson AL (2018a) Machine learning-enabled discovery and design of membrane-active peptides. Bioorg Med Chem 26:2708–2718
Lee MW, Lee EY, Ferguson AL, Wong GCL (2018b) Machine learning antimicrobial peptide sequences: some surprising variations on the theme of amphiphilic assembly. Curr Opin Colloid Interface Sci 38:204–213
Lee EY, Lee MW, Wong GCL (2019a) Modulation of toll-like receptor signaling by antimicrobial peptides. Semin Cell Dev Biol 88:173–184
Lee EY et al (2019b) Helical antimicrobial peptides assemble into protofibril scaffolds that present ordered dsDNA to TLR9. Nat Comm 10:1012
Lewenza S (2013) Extracellular DNA-induced antimicrobial peptide resistance mechanisms in Pseudomonas aeruginosa. Front Microbiol 4:21
Lewis LA, Shafer WM, Dutta Ray T, Ram S, Rice PA (2013) Phosphoethanolamine residues on the lipid A moiety of Neisseria gonorrhoeae lipooligosaccharide modulate binding of complement inhibitors and resistance to complement killing. Infect Immun 81:33–42
Li YH, Tian X (2012) Quorum sensing and bacterial social interactions in biofilms. Sensors (Basel) 12:2519–2538
Li M, Cha DJ, Lai Y, Villaruz AE, Sturdevant DE, Otto M (2007a) The antimicrobial peptide-sensing system aps of Staphylococcus aureus. Mol Microbiol 66:1136–1147
Li M, Lai Y, Villaruz AE, Cha DJ, Sturdevant DE, Otto M (2007b) Gram-positive three-component antimicrobial peptide-sensing system. Proc Natl Acad Sci U S A 104:9469–9474
Li C et al (2010) Limitations of peptide retro-inverso isomerization in molecular mimicry. J Biol Chem 285:19572–19581
Li Y, Xiang Q, Zhang Q, Huang Y, Su Z (2012) Overview on the recent study of antimicrobial peptides: origins, functions, relative mechanisms and application. Peptides 37:207–215
Li X, Wu X, Zhu J, Shen Y (2018) Amexanthomycins A-J, pentangular polyphenols produced by Amycolatopsis mediterranei S699rifA. Appl Microbiol Biotechnol 102:689–702
Liu W, Caffrey M (2005) Gramicidin structure and disposition in highly curved membranes. J Struct Biol 150:23–40
Liu Y, Yang Y, Wang C, Zhao X (2013) Stimuli-responsive self-assembling peptides made from antibacterial peptides. Nanoscale 5:6413–6421
Liu X et al (2016) Novel antimicrobial peptide–modified azithromycin-loaded liposomes against methicillin-resistant Staphylococcus aureus. International Journal of Nanomedicine 11:6781
Llewellyn AC et al (2012) NaxD is a deacetylase required for lipid A modification and Francisella pathogenesis. Mol Microbiol 86:611–627
Llobet E, Tomás JM, Bengoechea JA (2008) Capsule polysaccharide is a bacterial decoy for antimicrobial peptides. Microbiology 154:3877–3886
Lombardi L, Falanga A, Del Genio V, Galdiero S (2019) A new hope: self-assembling peptides with antimicrobial activity. Pharmaceutics 11
Loutet S, Valvano M (2011) Extreme antimicrobial peptide and polymyxin B resistance in the genus Burkholderia. Front Cell Infect Microbiol 2
Loutet SA, Flannagan RS, Kooi C, Sokol PA, Valvano MA (2006) A complete lipopolysaccharide inner core oligosaccharide is required for resistance of Burkholderia cenocepacia to antimicrobial peptides and bacterial survival in vivo. J Bacteriol 188:2073–2080
Magana M et al (2020) The value of antimicrobial peptides in the age of resistance. The Lancet Infect Dis 20:e216–e230
Mah T-FC, O’Toole GA (2001) Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol 9:34–39
Maisey HC, Quach D, Hensler ME, Liu GY, Gallo RL, Nizet V, Doran KS (2008) A group B streptococcal pilus protein promotes phagocyte resistance and systemic virulence. FASEB J 22:1715–1724
Maity S, Nir S, Zada T, Reches M (2014) Self-assembly of a tripeptide into a functional coating that resists fouling. Chem Commun (Camb) 50:11154–11157
Malekkhaiat Häffner S, Malmsten M (2018) Influence of self-assembly on the performance of antimicrobial peptides. Curr Opin Colloid Interface Sci 38:56–79
Malina A, Shai Y (2005) Conjugation of fatty acids with different lengths modulates the antibacterial and antifungal activity of a cationic biologically inactive peptide. Biochem J 390:695–702
Malmir S, Bahreinian M, Yeganeh SZ, Mirnejad R, Saberi MMMF (2018) Molecular mechanisms of resistance to conventional antibiotics in bacteria. Int J Med Rev 5:118–129
Mason KM, Munson RS, Bakaletz LO (2005) A Mutation in the sap Operon attenuates survival of nontypeable Haemophilus influenzae in a chinchilla model of otitis media. Infect Immun 73:599–608
Mason KM, Bruggeman ME, Munson RS, Bakaletz LO (2006) The non-typeable Haemophilus influenzae Sap transporter provides a mechanism of antimicrobial peptide resistance and SapD-dependent potassium acquisition. Mol Microbiol 62:1357–1372
Mason KM, Raffel FK, Ray WC, Bakaletz LO (2011) Heme utilization by nontypeable Haemophilus influenzae is essential and dependent on sap transporter function. J Bacteriol 193:2527–2535
Matsuzaki K (2009) Control of cell selectivity of antimicrobial peptides. Biochim Biophys Acta 1788:1687–1692
Matsuzaki K, Sugishita K-I, Harada M, Fujii N, Miyajima K (1997) Interactions of an antimicrobial peptide, magainin 2, with outer and inner membranes of Gram-negative bacteria. Biochim Biophys Acta 1327:119–130
Mattiuzzo M, De Gobba C, Runti G, Mardirossian M, Bandiera A, Gennaro R, Scocchi M (2014) Proteolytic activity of Escherichia coli oligopeptidase B against proline-rich antimicrobial peptides. J Microbiol Biotechnol 24:160–167
Mayville P, Ji G, Beavis R, Yang H, Goger M, Novick RP, Muir TW (1999) Structure-activity analysis of synthetic autoinducing thiolactone peptides from Staphylococcus aureus responsible for virulence. Proc Natl Acad Sci U S A 96:1218–1223
Mazzotta AS, Montville TJ (1997) Nisin induces changes in membrane fatty acid composition of Listeria monocytogenes nisin-resistant strains at 10°C and 30°C. J Appl Microbiol 82:32–38
McCloskey AP et al (2016) Self-assembling ultrashort NSAID-peptide nanosponges: multifunctional antimicrobial and anti-inflammatory materials. RSC Advances 6:114738–114749
Medema MH, Paalvast Y, Nguyen DD, Melnik A, Dorrestein PC, Takano E, Breitling R (2014) Pep2Path: automated mass spectrometry-guided genome mining of peptidic natural products. PLoS Comput Biol 10:e1003822
Meikle TG, Conn CE, Separovic F, Drummond CJ (2016) Exploring the structural relationship between encapsulated antimicrobial peptides and the bilayer membrane mimetic lipidic cubic phase: studies with gramicidin A. RSC Advances 6:68685–68694
Meikle TG, Zabara A, Waddington L, Separovic F, Drummond C, Conn CE (2017) Incorporation of antimicrobial peptides in nanostructured lipid membrane mimetic bilayer cubosomes. Colloids Surf B, Biointerf, pp 143–151
Meikle TG, Dyett BP, Strachan JB, White J, Drummond CJ, Conn CE (2020) Preparation, characterization, and antimicrobial activity of cubosome encapsulated metal nanocrystals. ACS Appl Mater Interfaces 12:6944–6954
Meikle TG, Dharmadana D, Hoffmann SV, Jones NC, Drummond CJ, Conn CE (2021) Analysis of the structure, loading and activity of six antimicrobial peptides encapsulated in cubic phase lipid nanoparticles. J Colloid Interface Sci 587:90–100
Meng H, Kumar K (2007) Antimicrobial activity and protease stability of peptides containing fluorinated amino acids. J Am Chem Soc 129:15615–15622
Miller SI, Kukral AM, Mekalanos JJ (1989) A two-component regulatory system (phoP phoQ) controls Salmonella typhimurium virulence. Proc Natl Acad Sci U S A 86:5054–5058
Mishra NN, Liu GY, Yeaman MR, Nast CC, Proctor RA, McKinnell J, Bayer AS (2011) Carotenoid-related alteration of cell membrane fluidity impacts Staphylococcus aureus susceptibility to host defense peptides. Antimicrob Agents Chemother 55:526–531
Mizukami S, Hosoda M, Satake T, Okada S, Hori Y, Furuta T, Kikuchi K (2010) Photocontrolled compound release system using caged antimicrobial peptide. J Am Chem Soc 132:9524–9525
Mohan A, Rajendran SR, Thibodeau J, Bazinet L, Udenigwe CC (2018) Liposome encapsulation of anionic and cationic whey peptides: influence of peptide net charge on properties of the nanovesicles. LWT 87:40–46
Mohimani H et al (2014a) Automated genome mining of ribosomal peptide natural products. ACS Chem Biol 9:1545–1551
Mohimani H, Liu WT, Kersten RD, Moore BS, Dorrestein PC, Pevzner PA (2014b) NRPquest: coupling mass spectrometry and genome mining for nonribosomal peptide discovery. J Nat Prod 77:1902–1909
Mondal S, Das S, Nandi AK (2020) A review on recent advances in polymer and peptide hydrogels. Soft Matter 16:1404–1454
Montanaro L, Poggi A, Visai L, Ravaioli S, Campoccia D, Speziale P, Arciola CR (2011) Extracellular DNA in biofilms. Int J Artif Organs 34:824–831
Mookherjee N, Anderson MA, Haagsman HP, Davidson DJ (2020) Antimicrobial host defence peptides: functions and clinical potential. Nat Rev Drug Discov 19:311–332
Moss JE, Fisher PE, Vick B, Groisman EA, Zychlinsky A (2000) The regulatory protein PhoP controls susceptibility to the host inflammatory response in Shigella flexneri. Cell Microbiol 2:443–452
Mulcahy H, Charron-Mazenod L, Lewenza S (2008) Extracellular DNA chelates cations and induces antibiotic resistance in Pseudomonas aeruginosa biofilms. PLoS Pathogens 4:e1000213
Mulcahy H, Charron-Mazenod L, Lewenza S (2010) Pseudomonas aeruginosa produces an extracellular deoxyribonuclease that is required for utilization of DNA as a nutrient source. Environ Microbiol 12:1621–1629
Müller RH, Runge S, Ravelli V, Mehnert W, Thünemann AF, Souto EB (2006) Oral bioavailability of cyclosporine: solid lipid nanoparticles (SLN®) versus drug nanocrystals. Int J Pharmaceutics 317:82–89
Mura M, Wang J, Zhou Y, Pinna M, Zvelindovsky AV, Dennison SR, Phoenix DA (2016) The effect of amidation on the behaviour of antimicrobial peptides. Eur Biophys J 45:195–207
Nagarajan D et al (2018) Computational antimicrobial peptide design and evaluation against multidrug-resistant clinical isolates of bacteria. J Biol Chem 293:3492–3509
Naoki S et al (2004) The contribution of the N-terminal structure of polymyxin B peptides to antimicrobial and lipopolysaccharide binding activity. Bull Chem Soc Japan 77:1915–1924
Nasompag S et al (2015) Effect of acyl chain length on therapeutic activity and mode of action of the CX-KYR-NH2 antimicrobial lipopeptide. Biochim Biophys Acta 1848:2351–2364
Neut D, Tijdens-Creusen EJ, Bulstra SK, van der Mei HC, Busscher HJ (2011) Biofilms in chronic diabetic foot ulcers--a study of 2 cases. Acta Orthop 82:383–385
Nichols D et al (2010) Use of ichip for high-throughput in situ cultivation of “uncultivable” microbial species. Appl Environ Microbiol 76:2445–2450
Nickel JC, Ruseska I, Wright JB, Costerton JW (1985) Tobramycin resistance of Pseudomonas aeruginosa cells growing as a biofilm on urinary catheter material. Antimicrob Agents Chemother 27:619–624
Nicolosi D, Scalia M, Nicolosi VM, Pignatello R (2010) Encapsulation in fusogenic liposomes broadens the spectrum of action of vancomycin against Gram-negative bacteria. Int J Antimicrob Agents 35:553–558
Nielsen JE, Bjornestad VA, Lund R (2018) Resolving the structural interactions between antimicrobial peptides and lipid membranes using small-angle scattering methods: the case of indolicidin. Soft Matter 14:8750–8763
Niemz A, Tirrell DA (2001) Self-association and membrane-binding behavior of melittins containing trifluoroleucine. J Am Chem Soc 123:7407–7413
Nizet V (2006) Antimicrobial peptide resistance mechanisms of human bacterial pathogens. Curr Issues Mol Biol 8:11
Nyberg P, Rasmussen M, Björck L (2004) α2-Macroglobulin-Proteinase complexes protect Streptococcus pyogenes from killing by the antimicrobial peptide LL-37. J Biol Chem 279:52820–52823
Oddo A, Hansen PR (2017) Hemolytic activity of antimicrobial peptides. Methods Mol Biol 1548:427–435
Ogunleye A, Bhat A, Irorere VU, Hill D, Williams C, Radecka I (2015) Poly-γ-glutamic acid: production, properties and applications. Microbiology 161:1–17
Ohki R, Giyanto TK, Masuyama W, Moriya S, Kobayashi K, Ogasawara N (2003) The BceRS two-component regulatory system induces expression of the bacitracin transporter, BceAB, in Bacillus subtilis. Mol Microbiol 49:1135–1144
Oie S, Huang Y, Kamiya A, Konishi H, Nakazawa T (1996) Efficacy of disinfectants against biofilm cells of methicillin-resistant Staphylococcus aureus. Microbios 85:223–230
Okshevsky M, Meyer RL (2015) The role of extracellular DNA in the establishment, maintenance and perpetuation of bacterial biofilms. Crit Rev Microbiol 41:341–352
Ong ZY, Wiradharma N, Yang YY (2014) Strategies employed in the design and optimization of synthetic antimicrobial peptide amphiphiles with enhanced therapeutic potentials. Adv Drug Deliv Rev 78:28–45
Otto M (2008) Staphylococcal biofilms. In: Romeo T (ed) Bacterial biofilms. Springer, Heidelberg, Berlin, pp 207–228
Otto M (2010) Staphylococcus colonization of the skin and antimicrobial peptides. Exp Rev Derm 5:183–195
Otto M, Peschel A, Götz F (1998) Producer self-protection against the lantibiotic epidermin by the ABC transporter EpiFEG of Staphylococcus epidermidis Tü3298. FEMS Microbiol Lett 166:203–211
Oyston PCF, Dorrell N, Williams K, Li S-R, Green M, Titball RW, Wren BW (2000) The response regulator PhoP is important for survival under conditions of macrophage-induced stress and virulence in Yersinia pestis. Infect Immun 68:3419–3425
Padilla E, Llobet E, Doménech-Sánchez A, Martínez-Martínez L, Bengoechea JA, Albertí S (2010) Klebsiella pneumoniae AcrAB efflux pump contributes to antimicrobial resistance and virulence. Antimicrob Agents Chemother 54:177–183
Papenfort K, Bassler BL (2016) Quorum sensing signal-response systems in Gram-negative bacteria. Nat Rev Microbiol 14:576–588
Parish T (2019) Steps to address anti-microbial drug resistance in today’s drug discovery. Expert Opin Drug Discov 14:91–94
Parthsarathy V et al (2013) A novel retro-inverso peptide inhibitor reduces amyloid deposition, oxidation and inflammation and stimulates neurogenesis in the APPswe/PS1ΔE9 mouse model of Alzheimer’s disease. PLoS One 8:e54769
Parveen N, Cornell KA (2011) Methylthioadenosine/S-adenosylhomocysteine nucleosidase, a critical enzyme for bacterial metabolism. Mol Microbiol 79:7–20
Pelletier MR et al (2013) Unique structural modifications are present in the lipopolysaccharide from colistin-resistant strains of Acinetobacter baumannii. Antimicrob Agents Chemother 57:4831–4840
Peschel A, Sahl H-G (2006) The co-evolution of host cationic antimicrobial peptides and microbial resistance. Nat Rev Microbiol 4:529–536
Peschel A, Otto M, Jack RW, Kalbacher H, Jung G, Götz F (1999) Inactivation of the dlt Operon in Staphylococcus aureus confers sensitivity to defensins, protegrins, and other antimicrobial peptides. J Biol Chem 274:8405–8410
Peschel A et al (2001) Staphylococcus aureus resistance to human defensins and evasion of neutrophil killing via the novel virulence factor Mprf is based on modification of membrane lipids with l-lysine. J Exp Med 193:1067–1076
Peterson E, Kaur P (2018) Antibiotic resistance mechanisms in bacteria: relationships between resistance determinants of antibiotic producers, environmental bacteria, and clinical pathogens. Front Microbiol 9:2928
Pfalzgraff A, Brandenburg K, Weindl G (2018) Antimicrobial peptides and their therapeutic potential for bacterial skin infections and wounds. Front Pharmacol 9:281
Piddock LJV (2006) Multidrug-resistance efflux pumps ? Not just for resistance. Nat Rev Microbiol 4:629–636
Pini A et al (2010) A novel tetrabranched antimicrobial peptide that neutralizes bacterial lipopolysaccharide and prevents septic shock in vivo. The FASEB Journal 24:1015–1022
Pirtskhalava M et al (2016) DBAASP v.2: an enhanced database of structure and antimicrobial/cytotoxic activity of natural and synthetic peptides. Nucl Acids Res 44:1104–1112
Poole K (2007) Efflux pumps as antimicrobial resistance mechanisms. Annals of Medicine 39:162–176
Pristovsek P, Kidric J (1999) Solution structure of polymyxins B and E and effect of binding to lipopolysaccharide: an NMR and molecular modeling study. J Med Chem 42:4604–4613
Prombutara P, Kulwatthanasal Y, Supaka N, Sramala I, Chareonpornwattana S (2012) Production of nisin-loaded solid lipid nanoparticles for sustained antimicrobial activity. Food Control 24:184–190
Pronk S et al (2013) GROMACS 4.5: a high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics:845–854
Pucci MJ (2006) Use of genomics to select antibacterial targets. Biochem Pharmacol 71:1066–1072
Qiao M, Saris PEJ (1996) Evidence for a role of NisT in transport of the lantibiotic nisin produced by Lactococcus lactis N8. FEMS Microbiol Lett 144:89–93
Quirynen M, Bollen CM (1995) The influence of surface roughness and surface-free energy on supra- and subgingival plaque formation in man. A review of the literature. J Clin Periodontol 22:1–14
Qureshi A, Thakur N, Himani T, Kumar M (2014) AVPdb: a database of experimentally validated antiviral peptides targeting medically important viruses. Nucl Acids Res 42:1147–1153
Radzishevsky IS, Rotem S, Zaknoon F, Gaidukov L, Dagan A, Mor A (2005) Effects of acyl versus aminoacyl conjugation on the properties of antimicrobial peptides. Antimicrob Agents Chemother 49:2412–2420
Raetz CRH, Whitfield C (2002) Lipopolysaccharide endotoxins. Annual Rev Biochem 71:635–700
Ragioto DA, Carrasco LD, Carmona-Ribeiro AM (2014) Novel gramicidin formulations in cationic lipid as broad-spectrum microbicidal agents. Int J Nanomedicine 9:3183
Rajeev L (2018) Antibiotic discovery. Mater Methods 8:2671
Rebeil R, Ernst RK, Gowen BB, Miller SI, Hinnebusch BJ (2004) Variation in lipid A structure in the pathogenic yersiniae. Mol Microbiol 52:1363–1373
Renwick MJ, Simpkin V, Mossialos E (2016) Targeting innovation in antibiotic drug discovery and development. In. Health Policy Series, vol 45. European Observatory on Health Systems and Policies., Copenhagen (Denmark),
Ribeiro da Cunha B, Fonseca LP, Calado CRC (2019) Antibiotic discovery: where have we come from, where do we go? Antibiotics (Basel) 8:45
Ron-Doitch S et al (2016) Reduced cytotoxicity and enhanced bioactivity of cationic antimicrobial peptides liposomes in cell cultures and 3D epidermis model against HSV. J Control Release 229:163–171
Rousseau F, Schymkowitz J, Serrano L (2006) Protein aggregation and amyloidosis: confusion of the kinds? Curr Opin Struct Biol 16:118–126
Rupp ME, Ulphani JS, Fey PD, Bartscht K, Mack D (1999) Characterization of the Importance of polysaccharide intercellular adhesin/hemagglutinin of Staphylococcus epidermidis in the pathogenesis of biomaterial-based infection in a mouse foreign body infection model. Infect Immun 67:2627–2632
Saar-Dover R et al (2012) D-Alanylation of lipoteichoic acids confers resistance to cationic peptides in group B streptococcus by increasing the cell wall density. PLoS Pathogens 8:e1002891
Sakala GP, Reches M (2018) Peptide-based approaches to fight biofouling. Advanced Materials Interfaces 5:1800073
Salick DA, Kretsinger JK, Pochan DJ, Schneider JP (2007) Inherent antibacterial activity of a peptide-based beta-hairpin hydrogel. J Am Chem Soc 129:14793–14799
Santajit S, Indrawattana N (2016) Mechanisms of antimicrobial resistance in ESKAPE pathogens. Biomed Res Int 2016:2475067
Schmidt NW, Wong GC (2013) Antimicrobial peptides and induced membrane curvature: geometry, coordination chemistry, and molecular engineering. Curr Opin Solid State Mater Sci 17:151–163
Schmidt NW et al (2011) Criterion for amino acid composition of defensins and antimicrobial peptides based on geometry of membrane destabilization. J Am Chem Soc 133:6720–6727
Schmidt NW et al (2015) Liquid-crystalline ordering of antimicrobial peptide–DNA complexes controls TLR9 activation. Nat Mater 14:696–700
Schmidtchen A, Frick I-M, Björck L (2001) Dermatan sulphate is released by proteinases of common pathogenic bacteria and inactivates antibacterial α-defensin. Mol Microbiol 39:708–713
Schmidtchen A, Frick I-M, Andersson E, Tapper H, Björck L (2002) Proteinases of common pathogenic bacteria degrade and inactivate the antibacterial peptide LL-37. Mol Microbiol 46:157–168
Schnaider L et al (2017) Self-assembling dipeptide antibacterial nanostructures with membrane disrupting activity. Nat Comm 8:1365
Schneider JP, Pochan DJ, Ozbas B, Rajagopal K, Pakstis L, Kretsinger J (2002) Responsive hydrogels from the intramolecular folding and self-assembly of a designed peptide. J Am Chem Soc 124:15030–15037
Schneider T, Gries K, Josten M, Wiedemann I, Pelzer S, Labischinski H, Sahl HG (2009) The lipopeptide antibiotic Friulimicin B inhibits cell wall biosynthesis through complex formation with bactoprenol phosphate. Antimicrob Agents Chemother 53:1610–1618
Schwarz S, Shen J, Kadlec K, Wang Y, Brenner Michael G, Fessler AT, Vester B (2016) Lincosamides, streptogramins, phenicols, and pleuromutilins: mode of action and mechanisms of resistance. Cold Spring Harb Perspect Med 6:a027037
Scudiero O et al (2015) Design and activity of a cyclic mini-β-defensin analog: a novel antimicrobial tool. Int J Nanomedicine 10:6523–6539
Sekyere JO (2018) Genomic insights into nitrofurantoin resistance mechanisms and epidemiology in clinical Enterobacteriaceae. Future Sci OA 4
Severino P et al (2017) Antimicrobial activity of polymyxin-loaded solid lipid nanoparticles (PLX-SLN): characterization of physicochemical properties and in vitro efficacy. Eur J Pharm Sci 106:177–184
Sforça ML et al (2004) How C-terminal carboxyamidation alters the biological activity of peptides from the venom of the eumenine solitary wasp. Biochemistry 43:5608–5617
Shah NR, Hancock REW, Fernandez RC (2014) Lipid A glucosamine modification confers resistance to cationic antimicrobial peptides and increases resistance to outer membrane perturbation. Antimicrob Agents Chemother 58:4931–4934
Shelton CL, Raffel FK, Beatty WL, Johnson SM, Mason KM (2011) Sap transporter mediated import and subsequent degradation of antimicrobial peptides in haemophilus. PLoS Pathogens 7:e1002360
Shi Y, Wareham DW, Phee LM, Azevedo HS (2021) Self-assembled peptide nanostructures for antibacterial applications. In: Guler MO (ed) Peptide-based biomaterials, Soft Matter Series, vol 11. The Royal Society of Chemistry, Cambridge, UK, pp 395–428
Shrestha S, Grilley M, Fosso MY, Chang CW, Takemoto JY (2013) Membrane lipid-modulated mechanism of action and non-cytotoxicity of novel fungicide aminoglycoside FG08. PLoS One 8:e73843
Sierra JM, Fuste E, Rabanal F, Vinuesa T, Vinas M (2017) An overview of antimicrobial peptides and the latest advances in their development. Expert Opin Biol Ther 17:663–676
Silva GA, Czeisler C, Niece KL, Beniash E, Harrington DA, Kessler JA, Stupp SI (2004) Selective differentiation of neural progenitor cells by high-epitope density nanofibers. Science 303:1352–1355
Silvestro L, Gupta K, Weiser JN, Axelsen PH (1997) The concentration-dependent membrane activity of cecropin A. Biochemistry 36:11452–11460
Simmons KJ, Chopra I, Fishwick CW (2010) Structure-based discovery of antibacterial drugs. Nat Rev Microbiol 8:501–510
Singh S, Singh SK, Chowdhury I, Singh R (2017) Understanding the Mechanism of bacterial biofilms resistance to antimicrobial agents. Open Microbiol J 11:53–62
Skinnider MA, Merwin NJ, Johnston CW, Magarvey NA (2017) PRISM 3: expanded prediction of natural product chemical structures from microbial genomes. Nucleic Acids Res 45:W49–W54
Smadbeck J et al (2014) De novo peptide design and experimental validation of histone methyltransferase inhibitors. PLoS One 9:e90095
Sohlenkamp C et al (2007) The lipid lysyl-phosphatidylglycerol is present in membranes of Rhizobium tropici CIAT899 and confers increased resistance to polymyxin B under acidic growth conditions. Mol Plant Microbe Interact 20:1421–1430
Som A, Yang L, Wong GC, Tew GN (2009) Divalent metal ion triggered activity of a synthetic antimicrobial in cardiolipin membranes. J Am Chem Soc 131:15102–15103
Song F, Koo H, Ren D (2015) Effects of material properties on bacterial adhesion and biofilm formation. J Dent Res 94:1027–1034
Sood R, Domanov Y, Pietiäinen M, Kontinen VP, Kinnunen PK (2008) Binding of LL-37 to model biomembranes: insight into target vs host cell recognition. Biochim Biophys Acta 1778:983–996
Soscia SJ et al (2010) The Alzheimer's disease-associated amyloid beta-protein is an antimicrobial peptide. PLoS One 5:e9505
Spoering AL, Gilmore MS (2006) Quorum sensing and DNA release in bacterial biofilms. Curr Opin Microbiol 9:133–137
Steenbergen JN, Alder J, Thorne GM, Tally FP (2005) Daptomycin: a lipopeptide antibiotic for the treatment of serious Gram-positive infections. J Antimicrob Chemother 55:283–288
Strom MB, Rekdal O, Svendsen JS (2002) Antimicrobial activity of short arginine- and tryptophan-rich peptides. J Pept Sci 8:431–437
Stromstedt AA, Pasupuleti M, Schmidtchen A, Malmsten M (2009) Evaluation of strategies for improving proteolytic resistance of antimicrobial peptides by using variants of EFK17, an internal segment of LL-37. Antimicrob Agents Chemother 53:593–602
Stumpe S, Schmid R, Stephens DL, Georgiou G, Bakker EP (1998) Identification of OmpT as the protease that hydrolyzes the antimicrobial peptide protamine before it enters growing cells of Escherichia coli. J Bacteriol 180:4002–4006
Sugano M, Morisaki H, Negishi Y, Endo-Takahashi Y, Kuwata H, Miyazaki T, Yamamoto M (2016) Potential effect of cationic liposomes on interactions with oral bacterial cells and biofilms. J Liposome Res 26:156–162
Suntharalingam P, Senadheera MD, Mair RW, Lévesque CM, Cvitkovitch DG (2009) The LiaFSR system regulates the cell envelope stress response in Streptococcus mutans. J Bacteriol 191:2973–2984
Sykes R (2010) The 2009 Garrod lecture: the evolution of antimicrobial resistance: a Darwinian perspective. J Antimicrob Chemother 65:1842–1852
Taha MN, Saafan AE, Ahmedy A, El Gebaly E, Khairalla AS (2019) Two novel synthetic peptides inhibit quorum sensing-dependent biofilm formation and some virulence factors in Pseudomonas aeruginosa PAO1. J Microbiol 57:618–625
Tam JP (1988) Synthetic peptide vaccine design: synthesis and properties of a high-density multiple antigenic peptide system. Proc Natl Acad Sci U S A 85:5409–5413
Tanaka H, Iwai Y, Oiwa R, Shinohara S, Shimizu S, Oka T, Omura S (1977) Studies on bacterial cell wall inhibitors. II. Inhibition of peptidoglycan synthesis in vivo and in vitro by amphomycin. Biochim Biophys Acta 497:633–640
Taneja NK, Ganguly T, Bakaletz LO, Nelson KJ, Dubey P, Poole LB, Deora R (2013) Alanine modification of a protease-susceptible outer membrane component by the Bordetella pertussis locus promotes resistance to antimicrobial peptides and polymorphonuclear leukocyte-mediated killing. J Bacteriol 195:5102–5111
Tasiemski A, Schikorski D, Le Marrec-Croq F, Pontoire-Van Camp C, Boidin-Wichlacz C, Sautière P-E (2007) Hedistin: a novel antimicrobial peptide containing bromotryptophan constitutively expressed in the NK cells-like of the marine annelid, Nereis diversicolor. Dev Comp Immun 31:749–762
Tetz VV, Tetz GV (2010) Effect of extracellular DNA destruction by DNase I on characteristics of forming biofilms. DNA Cell Biol 29:399–405
Thedieck K et al (2006) The MprF protein is required for lysinylation of phospholipids in listerial membranes and confers resistance to cationic antimicrobial peptides (CAMPs) on Listeria monocytogenes. Mol Microbiol 62:1325–1339
Thoendel M, Kavanaugh JS, Flack CE, Horswill AR (2011) Peptide signaling in the staphylococci. Chem Rev 111:117–151
Tian X, Sun F, Zhou X-R, Luo S-Z, Chen L (2015) Role of peptide self-assembly in antimicrobial peptides. J Pep Sci 21:530–539
Travkova OG, Moehwald H, Brezesinski G (2017) The interaction of antimicrobial peptides with membranes. Adv Colloid Interface Sci 247:521–532
Tzeng Y-L, Ambrose KD, Zughaier S, Zhou X, Miller YK, Shafer WM, Stephens DS (2005) Cationic antimicrobial peptide resistance in Neisseria meningitidis. J Bacteriol 187:5387–5396
Uzzell T, Stolzenberg ED, Shinnar AE, Zasloff M (2003) Hagfish intestinal antimicrobial peptides are ancient cathelicidins. Peptides 24:1655–1667
Velkov T, Roberts KD, Li J (2017) Rediscovering the octapeptins. Natural Product Reports 34:295–309
Ventola CL (2015a) The antibiotic resistance crisis: part 1: causes and threats. Pharm Ther 40:277–283
Ventola CL (2015b) The antibiotic resistance crisis: part 2: management strategies and new agents. Pharm Ther 40:344–352
Verkleij A, Zwaal R, Roelofsen B, Comfurius P, Kastelijn D, Van Deenen L (1973) The asymmetric distribution of phospholipids in the human red cell membrane. A combined study using phospholipases and freeze-etch electron microscopy. Biochim Biophys Acta 323:178–193
Verly RM et al (2008) Effect of cholesterol on the interaction of the amphibian antimicrobial peptide DD K with liposomes. Peptides 29:15–24
Vestby LK, Gronseth T, Simm R, Nesse LL (2020) Bacterial biofilm and its role in the pathogenesis of disease. Antibiotics (Basel) 9
Vizioli J, Salzet M (2002) Antimicrobial peptides from animals: focus on invertebrates. Trends in Pharmacological Sciences 23:494–496
Vorkapic D, Pressler K, Schild S (2016) Multifaceted roles of extracellular DNA in bacterial physiology. Curr Genet 62:71–79
Vuong C, Kocianova S, Voyich JM, Yao Y, Fischer ER, DeLeo FR, Otto M (2004a) a crucial role for exopolysaccharide modification in bacterial biofilm formation, immune evasion, and virulence. J Biol Chem 279:54881–54886
Vuong C, Voyich JM, Fischer ER, Braughton KR, Whitney AR, DeLeo FR, Otto M (2004b) Polysaccharide intercellular adhesin (PIA) protects Staphylococcus epidermidis against major components of the human innate immune system. Cell Microbiol 6:269–275
Wachinger M et al (1998) Antimicrobial peptides melittin and cecropin inhibit replication of human immunodeficiency virus 1 by suppressing viral gene expression. J Gen Virol 79:731–740
Waghu FH, Barai RS, Gurung P, Idicula-Thomas S (2016) CAMPR3: a database on sequences, structures and signatures of antimicrobial peptides. Nucleic Acids Res 44:D1094–D1097
Walters MC 3rd, Roe F, Bugnicourt A, Franklin MJ, Stewart PS (2003) Contributions of antibiotic penetration, oxygen limitation, and low metabolic activity to tolerance of Pseudomonas aeruginosa biofilms to ciprofloxacin and tobramycin. Antimicrob Agents Chemother 47:317–323
Wang Z, Wang G (2004) APD: the antimicrobial peptide database. Nucleic Acids Res 32:D590–D592
Wang X, Karbarz MJ, McGrath SC, Cotter RJ, Raetz CRH (2004a) MsbA Transporter-dependent Lipid A 1-dephosphorylation on the periplasmic surface of the inner membrane: topography of francisella novicida LpxE expressed in Escherichia coli. J Biol Chem 279:49470–49478
Wang X, Preston JF, Romeo T (2004b) The pgaABCD Locus of Escherichia coli promotes the synthesis of a polysaccharide adhesin required for biofilm formation. J Bacteriol 186:2724–2734
Wang X, McGrath SC, Cotter RJ, Raetz CRH (2006) Expression, cloning and periplasmic orientation of the Francisella novicida lipid A 4′-phosphatase LpxF. J Biol Chem 281:9321–9330
Wang G, Li X, Wang Z (2016a) APD3: the antimicrobial peptide database as a tool for research and education. Nucleic Acids Res 44:D1087–D1093
Wang H et al (2016b) Enzyme-catalyzed formation of supramolecular hydrogels as promising vaccine adjuvants. Adv Funct Mater 26:1822–1829
Webber MJ, Tongers J, Newcomb CJ, Marquardt KT, Bauersachs J, Losordo DW, Stupp SI (2011) Supramolecular nanostructures that mimic VEGF as a strategy for ischemic tissue repair. Proc Natl Acad Sci U S A 108:13438–13443
Weinstock MT, Francis JN, Redman JS, Kay MS (2012) Protease-resistant peptide design-empowering nature's fragile warriors against HIV. Biopolymers 98:431–442
Whitchurch CB, Tolker-Nielsen T, Ragas PC, Mattick JS (2002) Extracellular DNA required for bacterial biofilm formation. Science 295:1487
WHO (2015) Global Antimicrobial Resistance Surveillance System (GLASS). https://www.who.int/glass/en/. Accessed 2020
WHO (2017) Global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. . https://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/. Accessed 2019
WHO (2019a) 2019 antibacterial agents in clinical development: an analysis of the antibacterial clinical development pipeline. ISBN 9789240000193. Geneva. https://apps.who.int/iris/handle/10665/330420
WHO (2019b) Antibacterial agents in preclinical development: an open access database. WHO/EMP/IAU/2019.12. Geneva. https://apps.who.int/iris/bitstream/handle/10665/330290/WHO-EMP-IAU-2019.12-eng.pdf
WHO (2020a) First meeting of the WHO Antifungal Expert Group on Identifying Priority Fungal Pathogens: meeting report. ISBN 978-92-4-000635-5. Geneva. https://www.who.int/publications/i/item/9789240006355
WHO (2020b) Record number of countries contribute data revealing disturbing rates of antimicrobial resistance. https://www.who.int/news-room/detail/01-06-2020-record-number-of-countries-contribute-data-revealing-disturbing-rates-of-antimicrobial-resistance. Accessed 2020
WHO (2020c) Target product profiles for needed antibacterial agents: enteric fever, gonorrhea, neonatal sepsis, urinary tract infections and meeting report. ISBN 978–92–4-000389-7. Geneva. https://www.who.int/publications/i/item/9789240003897
Wilton M, Charron-Mazenod L, Moore R, Lewenza S (2016) Extracellular DNA acidifies biofilms and induces aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother 60:544–553
Wohlleben W, Mast Y, Stegmann E, Ziemert N (2016) Antibiotic drug discovery. Microb Biotechnol 9:541–548
Wright GD (2010) Q&A Antibiotic resistance where does it come from and what can we do about it. BMC Biology 8:123
Yang D et al (1999) β-Defensins: linking innate and adaptive immunity through dendritic and T Cell CCR6. Science 286:525–528
Yang Z, Liang G, Guo Z, Guo Z, Xu B (2007) Intracellular hydrogelation of small molecules inhibits bacterial growth. Angew Chem Int Ed Engl 46:8216–8219
Yang L et al (2008) Mechanism of a prototypical synthetic membrane-active antimicrobial: efficient hole-punching via interaction with negative intrinsic curvature lipids. Proc Natl Acad Sci U S A 105:20595–20600
Yeaman MR, Yount NY (2003) Mechanisms of antimicrobial peptide action and resistance. Pharmacol Rev 55:27–55
Yoder NC, Kumar K (2002) Fluorinated amino acids in protein design and engineering. Chem Soc Rev 31:335–341
Zabara M et al (2019) Multifunctional nano-biointerfaces: cytocompatible antimicrobial nanocarriers from stabilizer-free cubosomes. Adv Funct Mater 29:1904007
Zähner D, Zhou X, Chancey ST, Pohl J, Shafer WM, Stephens DS (2010) Human antimicrobial peptide LL-37 induces MefE/Mel-mediated macrolide resistance in Streptococcus pneumoniae. Antimicrob Agents Chemother 54:3516–3519
Zasloff M (2002) Antimicrobial peptides of multicellular organisms. Nature 415:389–395
Zavascki AP, Goldani LZ, Li J, Nation RL (2007) Polymyxin B for the treatment of multidrug-resistant pathogens: a critical review. J Antimicrob Chemother 60:1206–1215
Zelezetsky I, Tossi A (2006) Alpha-helical antimicrobial peptides-using a sequence template to guide structure-activity relationship studies. Biochim Biophys Acta 1758:1436–1449
Zetterberg MM, Reijmar K, Pränting M, Engström Å, Andersson DI, Edwards K (2011) PEG-stabilized lipid disks as carriers for amphiphilic antimicrobial peptides. J Control Release 156:323–328
Zhang LJ, Gallo RL (2016) Antimicrobial peptides. Curr Biol 26:R14–R19
Zhang Q, Yie G, Li Y, Yang Q, Nagai T (2000) Studies on the cyclosporin A loaded stearic acid nanoparticles. Int J Pharmaceutics 200:153–159
Zhao X, Wu H, Lu H, Li G, Huang Q (2013) LAMP: a database linking antimicrobial peptides PLoS One 8:66557
Zhu L, Lau GW (2011) Inhibition of competence development, horizontal gene transfer and virulence in Streptococcus pneumoniae by a modified competence stimulating peptide. PLoS Pathogens 7:e1002241
Zhu M, Liu P, Niu Z-W (2017) A perspective on general direction and challenges facing antimicrobial peptides. Chinese Chemical Letters 28:703–708
Zhu S, Duan Y, Huang Y (2019) The application of ribosome engineering to natural product discovery and yield improvement in streptomyces. Antibiotics (Basel) 8:133
Zimmerberg J, Kozlov MM (2006) How proteins produce cellular membrane curvature. Nat Rev Mol Cell Biol 7:9–19
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Cardoso, P., Glossop, H., Meikle, T.G. et al. Molecular engineering of antimicrobial peptides: microbial targets, peptide motifs and translation opportunities. Biophys Rev 13, 35–69 (2021). https://doi.org/10.1007/s12551-021-00784-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12551-021-00784-y