
Abstract
This paper investigates agricultural management choices of farmers at the Neolithic site of Kouphvouno, southern Greece. Previous stable isotopic analysis of charred plant remains and bone animal collagen showed that throughout the Neolithic occupation of this site, farmers employed species-specific strategies to cultivate crops and herd domestic animals. Additional analyses of charred plant remain carried out in this study (including einkorn, a cereal species not measured before) expanding our understanding of the diversity and flexibility of early crop cultivation on a local scale. Furthermore, sequential tooth enamel carbonate isotopic analyses are used to assess the seasonal dietary and grazing patterns of domestic sheep and goat, providing a more nuanced picture of the roles of these animals in the subsistence economy of this community. The results show that the species-specific cultivation system was dictated by the crops’ ecological adaptations. Based on a small number of individuals available for analysis, the findings suggest that animal management was also likely driven by cultural choices, and involved foddering of goats managed for milk and local grazing of sheep managed for meat.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Applications of stable isotopic analysis in the field of Neolithic archaeology have made insightful contributions to our understanding of how early farming systems functioned, shedding light on the scale and intensity of ancient crop cultivation as well as seasonal and multi-annual patterns in livestock herding management (Balasse et al. 2012; Bogaard et al. 2013; Makarewicz et al. 2017). At the Middle–Late Neolithic site of Kouphovouno, southern Greece, an investigation that integrated stable isotopic measurements of archaeological plant and animal remains provided some of the first direct evidence for species-specific management strategies employed by Neolithic farmers (Vaiglova et al. 2014a). Furthermore, it presented preliminary insight into subtle differences between long-term average diets of domestic sheep and goats. These findings support the model—based on results of systematic archaeobotanical and archaeozoological analyses (Halstead 1981, 2011; Bogaard 2004, 2005) — that the management techniques employed by these ancient agropastoralists were small-scale and intensive. The pilot study raised further questions regarding the differential management of crops and seasonal herding patterns of animals that formed the basis of this mixed farming economy.
This paper presents the results of a second phase of crop and animal isotopic analyses carried out at Kouphovouno. Additional samples (including a crop species not measured before) became available with the completion of the archaeobotanical analysis, enabling a more thorough assessment of the crop cultivation practices developed by the Neolithic farmers. Furthermore, sequential analysis of tooth enamel carbonate of sheep and goats from both phases of the site’s occupation was used to assess the animals’ seasonal and multi-annual patterns in grazing behavior. The combined crop and animal stable isotopic results further our understanding of the symbiotic relationship between crop cultivation and animal husbandry in an early farming context. This information provides direct insight into the roles that plants and animals played in the local subsistence economy, as well as the ways that people adapted their management strategies to their environmental constraints and opportunities.
The archaeological site of Kouphovouno
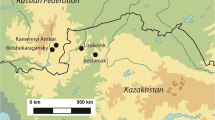
Kouphovouno is a Middle–Late Neolithic (5800–5000 cal BC, Mee et al. 2014) site located about 2.5 km southwest of modern-day Sparta, southern Greece (see Fig. 1). The tell site rises to an elevation of 5 m above the surrounding Sparta basin and spreads over an area of 4–5 ha. The mound is bordered in the north by a seasonal stream, Parori, which originates in a system of alluvial fans to the west and feeds into the Evrotas river 3.5 km to the east (Cavanagh et al. 2004, 2007). To the south, the mound is met by a perennial pond (Fouache et al. 2007). The alluvial fans occupy the piedmont zone of the Taygetos Mountains, a limestone massif that reaches an elevation of 2404 masl. Geomorphological reconstruction suggests that in the Neolithic, water was abundant close to the surface of the Sparta basin, causing annual flooding (Fouache et al. 2007).

Map of the Peloponnesian peninsula, southern Greece, showing the location of the Neolithic site of Kouphovouno (from Rivals et al. 2011)
The excavated portions of the Neolithic village indicate that the settlement was nucleated and may have been divided into neighborhoods. There were refuse disposal areas located in close proximity to the habitation spaces. A possible garden was identified in Area G. No human remains dating to the Neolithic period were found (Cavanagh et al. 2004, 2007). The transition from Middle Neolithic (MN) to Late Neolithic (LN) (~ 5400 cal BC) was marked by a shift from more permanent to more ephemeral architecture, accompanied by a change from a more uniform ceramic assemblage to one that is more diverse and contains black ware pottery (Mee et al. 2014). The data from Kouphovouno suggests that the Middle to Late Neolithic transition in southern Greece was not as abrupt as previously thought (Mee et al. 2014). Cavanagh and Renard (2014) argue that the diverse collection of figurines, together with obsidian and flint blades, suggests that the Neolithic inhabitants participated in a network of exchange that extended across the Adriatic zone (the Balkans to the north and Italy to the west) and into the eastern Aegean.
The plant assemblage from Kouphovouno is dominated by domestic crop species, mainly cereals: one-seeded einkorn wheat (Triticum monococcum), emmer wheat (Triticum dicoccum), free-threshing wheat (Triticum aestivum/Triticum durum), lentil (Lens culinaris), pea (Pisum sativum), and bitter vetch (Vicia ervilia). The high incidence of free-threshing wheat at Kouphovouno presents a contrast to the situation in northern Greece, where farmers are argued to have rejected this crop by choice (Valamoti and Jones 2003). At Kouphovouno, free-threshing wheat is not only found in rich storage contexts, but previous stable isotope analysis suggests that it was cultivated under more intensively maintained growing conditions (likely using the application of farmyard manure or other organic waste) than hulled barley. Furthermore, it may have been grown exclusively for human consumption (Vaiglova et al. 2014a). The stable isotopic values of einkorn were not previously measured, and the opportunity to do so in this study provides a chance to expand our understanding of crop cultivation systems employed by the Neolithic farmers at the site. The previous isotopic work also suggests that peas were grown at a high intensity, possibly in rotation with free-threshing wheat but under a higher water status than the cereals, likely using hand-watering.
The faunal assemblage is dominated by domestic species, namely sheep (Ovis aries), goats (Capra sp.), pigs (Sus scrofa) and cattle (Bos taurus). Analysis of the mortality profiles of the ovicaprids (sheep and goats) suggests that either the farmers practiced a mixed primary and secondary product exploitation strategy during both periods of occupation or that the exploitation strategy shifted focus from secondary products in the Middle Neolithic to meat in the Late Neolithic (Cantuel et al. 2008). The mortality profiles of the cattle could not be determined due to the low numbers of preserved individuals. Analysis of dental micro- and meso-wear patterns did not indicate any dietary differences between sheep and goats (distinguished using morphological criteria) in the last 2–3 weeks of their lives (Rivals et al. 2011). Stable isotope analysis of the animals’ bone collagen, identified to species using Zooarchaeology Mass Spectrometry (ZooMS), however, suggests that there was a chronological shift in the long-term average diets of the ovicaprids, which points to a change in the management strategy over time (Vaiglova et al. 2014a).
Principles of stable isotope analysis
Isotopic ratios of organic (bone collagen, charred grains) and inorganic (tooth enamel) materials recovered from archaeological sites contain information about past climatic conditions, animal seasonal dietary behavior and plant growing conditions (Balasse et al. 1999; Bogaard et al. 2007; Lee-Thorp 2008). Stable carbon isotopic values (13C/12C, δ13C) of plants are mainly determined by their photosynthetic pathway (C3 or C4) and to a smaller degree by environmental factors including temperature, humidity, light and air pressure (Ehleringer et al. 1991; Tieszen 1991; Cerling et al. 1997; Hartman and Danin 2010).
C3 plants include domestic crops such as Triticum monococcum and have δ13C values ranging from − 34 to − 21‰, with an average around − 27‰. C4 plants include arid-adapted grasses such as Cynodon dactylon and have δ13C values ranging from − 17 to − 9‰, with an average around − 12‰ (Vogel and van der Merwe 1977; Cerling et al. 1997). Plant δ13C values are negatively correlated with moisture availability, so plants growing in regions with higher mean annual rainfall have more negative δ13C values compared to plants growing in drier regions (Hartman and Danin 2010). Physiological differences in carbon assimilation of plants utilizing the same photosynthetic pathway also cause small differences in δ13C values (Hartman and Danin 2010). For example, barley has been found to have consistently lower δ13C values than wheat grown under the same moisture availability (Araus et al. 1997a, b). In Mediterranean climates, the δ13C values of plants are highest during the warm/dry summers due to increased evapotranspiration which causes 13C enrichment, and lowest during the cold/wet winters (Hartman and Danin 2010).
Stable nitrogen isotope values (15N/14N, δ15N) values of plants are primarily determined by the source from which the plants obtain nitrogen (Sharp 2007; Szpak 2014). Nitrogen-fixers (such as members of the Leguminosae family) host mycorrhizae fungi that fix N2 directly from the atmosphere. As a result, they have values close to 0‰, which is the δ15N value of AIR (atmospheric inhalable reservoir) (Craine et al. 2009; Szpak 2014). Nitrogen non-fixers (such as Triticum and Hordeum) assimilate nitrogen from soil and are thus influenced by a host of natural and anthropogenic soil 15N enrichment factors. Higher temperatures and lower precipitation create conditions that are more prone to N loss, and thus cause 15N enrichment (Szpak 2014). Mean annual precipitation is negatively correlated with plant δ15N values, so plants growing in more arid regions tend to have higher δ15N values compared to plants growing in wetter regions (Heaton et al. 1986; Ambrose 1991; Craine et al. 2009; Hartman and Danin 2010). However, certain chemical transformations that occur in soil, such as denitrification (which involves the transformation of inorganic nitrate, NO3−, into gaseous N2) also cause soil 15N enrichment (Tiedje et al. 1982; Högberg 1997), so an understanding of a site’s hydrological history is crucial for the interpretation of past soil and plant δ15N values.
Anthropogenic factors that drive soil 15N enrichment include methods to improve soil fertility such as tillage, burning and application of animal manure or decomposing midden material (Bateman et al. 2005; Bogaard et al. 2007; Fraser et al. 2011; Hobbie and Högberg 2012; Szpak 2014). Tillage promotes mineralization of soil organic matter and brings soil from deeper in a soil profile (and more enriched in 15N) closer to the surface (Szpak 2014). Manure/compost undergo increased ammonia volatilization, which causes loss of the lighter 14N in the form of gaseous NH3, leaving the substrate enriched in 15N. Plants that grow in heavily managed soil thus have higher δ15N values than plants growing in unmanaged soils.
Animals assimilate C and N from food into their hard and soft tissues and their isotopic values become enriched at every level of the food chain. Thus, carnivores have higher δ15N values compared to herbivores, and herbivores have higher δ15N compared to the plants they consume (DeNiro and Epstein 1981; Minagawa and Wada 1984; Hedges and Reynard 2007). The diet to tissue offset is between 3 and 6‰ in δ15N values of bone collagen (Schoeninger and DeNiro 1984; Bocherens and Drucker 2003; O’Connell et al. 2012), 3–6‰ for δ13C in bone collagen and 9–15‰ for δ13C in enamel bioapatite; the actual offset depending on the animals’ digestive physiology (van der Merwe and Vogel 1978; Krueger and Sullivan 1984; Lee-Thorp et al. 1989; Cerling and Harris 1999). Animals that consume mixed C3 and C4 diets have tooth enamel values above − 8‰ (Cerling et al. 1997). Animal bones remodel themselves throughout the life of individuals at varying rates (DeNiro and Epstein 1981). The isotopic composition of bone collagen, which is more diagenetically robust than the mineral portion of bone (Hare 1980; Collins et al. 2002), thus reflects the dietary inputs over the last several years of the individuals’ lives. Carbon is preferentially routed from the protein component of diets and together, δ13C and δ15N values of bone collagen from archaeological specimens serve as indicators of their long-term dietary protein intake (Krueger and Sullivan 1984; Ambrose and Norr 1993; Tieszen and Fagre 1993).
Animals incorporate oxygen into hard tissues like tooth enamel mainly from ingested food and water, which in turn reflect the isotopic composition of meteoric, surface and plant leaf water (Longinelli 1984; Luz et al. 1984; Bryant et al. 1996). Stable oxygen isotopic values (18O/16O, δ18O) of meteoric water are driven by fractionation during the hydrological cycle, so that precipitation closer to the coast is isotopically lighter compared to precipitation in upland areas (Dansgaard 1964; Bowen and Wilkinson 2002; Hoefs 2009). At mid to high latitudes, higher temperatures and increased rates of evapotranspiration favor 18O enrichment in surface and plant leaf water, resulting in higher δ18O values in the summer and lower values in the winter (Gat 1980; Allison and Hughes 1983; Rozanski et al. 1993).
Enamel mineralizes sequentially along the axis of tooth growth (starting at the tooth crown and ending at the enamel root junction), incorporating oxygen from ingested water in equilibrium with body water (Land et al. 1980; Luz et al. 1984). The second molars (M2) of herbivores like sheep and goats complete mineralization during the first 12–13 months of the individuals’ lives (Brown et al. 1960; Weinreb and Sharav 1964; Balasse et al. 2001; Hillson 2005; Towers et al. 2014). Once mineralized during the developmental years, tooth enamel does not remodel itself, and its isotopic composition thus reflects the dietary and water inputs consumed during the mineralization period. δ18O values of enamel samples taken along the axis of tooth growth reflect an entire annual cycle of seasonal changes in δ18O values. Sequential enamel δ18O and δ13C values thus provide information on the seasonal changes in the animals’ dietary and mobility patterns and serve as short-term indicators of their grazing behavior (Bocherens et al. 2001; Balasse 2002; Balasse et al. 2002).
Individuals that eat fresh vegetation and do not migrate across altitudes during the tooth mineralization period typically have sinusoidal intra-tooth δ13C and δ18O value sequences, with the maximum values reflecting composition of plants and water ingested during the summer and minimum values reflecting the composition of plants and water ingested during the winter (Balasse et al. 2002, 2013; Kirsanow et al. 2008). Animals that eat collected fodder for part of the year tend to have flattened δ13C value sequences, since the consumed vegetation does not reflect the seasonal changes in carbon isotopic discrimination (Makarewicz 2014). However, if they are obligate drinkers (like sheep and goats), their δ18O sequences will still fluctuate predictably due to the seasonal changes in temperature and evapotranspiration. Animals that migrate across altitudinal boundaries, such as those that partake in transhumant pastoralism and spend the dry season at higher altitudes, can show dampened δ18O value sequences due to consumption of water that does not reflect the annual extremes (Longinelli and Selmo 2003; Britton et al. 2009; Henton et al. 2010).
Materials and methods
In the first part of this study, we measured the δ13C and δ15N values of samples from the archaeobotanical assemblage that were not available during the pilot study. In the second part, we obtained incremental samples of tooth enamel carbonate of sheep and goats, and measured the seasonal fluctuations in δ13C and δ18O values. Permission to carry out the scientific analyses was granted by the excavation directors (Prof William Cavanagh, Prof Christopher Mee, Prof Josette Renard) and supported by the Ephorea of Antiquities of Laconia, Greece (permit ΥΠΠΟΑ/ΣΥΝΤ/Φ44/234210/6186).
Stage 1: additional charred plant isotope analysis
δ13C and δ15N values were measured from 19 samples of charred grains/seeds (see Table 1). Each sample was taken from a discrete contextual unit and included between 3 and 13 whole grains/seeds or fragments thereof. The chronological assignments were made on the basis of stratigraphic associations and radiocarbon dating. Five contexts analyzed in this study and by Vaiglova et al. (2014a) (C0854, C0848, C0844, G2003, G1139) have been directly dated using radiocarbon (Mee et al. 2014). Only samples that were charred in the ‘optimal charring window’ (i.e. at charring conditions that do not obscure the original isotopic signatures, cf. Nitsch et al. 2015) were measured. All samples were chemically pre-treated using a gentle acid-only treatment with 0.5 M HCl at 80 °C for 30 min (Vaiglova et al. 2014b).
Stable carbon and nitrogen isotopic compositions were determined using a SerCon 20/22 continuous flow mass spectrometer coupled to a Callisto elemental analyzer at the Research Laboratory for Archaeology and the History of Art, University of Oxford. Analysis was carried out separately for δ13C and δ15N values, as the target weights differed for the two sets of measurements. δ13C and δ15N values were calibrated relative to VPDB and AIR, respectively. Measurement uncertainty for δ13C was monitored using international reference standards: CH-6 (δ13C = − 10.45 ± 0.03‰) and CH-7 (δ13C = − 32.15 ± 0.05‰). Measurement uncertainty for δ15N was monitored using international reference standards N2 (δ15N = 20.3 ± 0.02‰) and caffeine (δ15N = 2.9 ± 0.03‰). Precision (u(Rw)) was determined to be ± 0.48 for δ13C and ± 0.29 for δ15N on the basis of repeated measurements of calibration standards and check standards. Accuracy (u(bias)) was determined to be ± 0.38 for δ13C and ± 0.34 for δ15N on the basis of the difference between the observed and known δ values of the check standards and the long-term standard deviations of the check standards. Using the equations from Szpak et al. (2017), the total analytical uncertainty was estimated to be ± 0.62 for δ13C and ± 0.45 for δ15N.
All plant isotope values were corrected for an average charring-induced isotopic enrichment of 0.3‰ in δ15N values and 0.1‰ in δ13C values (the confidence intervals for these average values are 0.003–0.22‰ for δ13C and 0.05–0.57‰ for δ15N) (Nitsch et al. 2015). The previously measured plant data from this assemblage (n = 28) were corrected using the more recent charring offset (the offset at the time of the first publication was 1.0‰ in δ15N, cf. Fraser et al. 2013) and will be presented here alongside the new measurements. This new charring correction did not change the interpretation of the previous results, as the distinction between the free-threshing wheat δ15N values and the hulled barley δ15N values remains the same (even though all the absolute corrected values are 0.7‰ higher).
The Δ13Cair for the Neolithic period was estimated to be − 6.5‰ using the AIRCO2_LOESS data calibrator (Cleveland 1979; Leuenberger et al. 1992; Indermühle et al. 1999; Ferrio et al. 2005). One sample of hulled barley (KFO34) and one sample of free-threshing wheat (KFO50), both from the MN, only yielded reliable δ15N measurements. One sample of free-threshing wheat (KFO52) only yielded a reliable δ13C measurement.
Stage 2: tooth enamel carbonate analysis
Sequential samples of tooth enamel were obtained from the second molars (M2) of sheep (n = 4) and goats (n = 4) from Kouphovouno. M2s mineralize within 12–13 months of the animals’ birth (Brown et al. 1960; Balasse et al. 2001). Because enamel does not remodel once mineralized, the isotopic values of M2s thus reflect the dietary and water inputs during the first year of the animals’ lives. Three sheep (KFO209, KFO217, KFO210) and two goats (KFO204, KFO207) date to the Middle Neolithic and one sheep (KFO212) and two goats (KFO214, KFO216) date to the Late Neolithic. The chronological assignments were made on the basis of stratigraphic associations and radiocarbon dating (Mee et al. 2014). Zooarchaeology Mass Spectrometry (ZooMS) (Buckley et al. 2010), carried out at the University of Manchester, was used to confirm the species identification of the samples. Six out of the eight samples were extracted from fully preserved mandibles, and these were aged using tooth wear stages established by Payne (1973). However, due to the state of preservation of the teeth, most of these age estimates are uncertain (see Table 2).
Prior to sampling, the external surface of the teeth was cleaned using a laboratory aluminum oxide sandblaster and a Dremel tool with a tungsten drill bit. A total of 5–10-mg sub-samples of powdered enamel were removed at 1 mm intervals along the axis of growth from the buccal side of each tooth, starting at the occlusal surface and ending at the enamel root junction (erj), following the protocol established by Balasse (2002) (see Fig. 2 for an image of a sampled tooth). The number of sub-samples per tooth ranged from 15 to 16 in sheep (total n = 61) and 11–22 in goats (total n = 65). The crown lengths ranged from 22 to 30 mm in sheep and 24–30 mm in goats.

Tooth enamel sampling of ovicaprid second molars carried out in stage 2 of this study
All samples were pre-treated using 1 M Ca-buffered acetic acid for 30 min at room temperature in order to remove possible exogenous carbonate contamination (Snoeck and Pellegrini 2015). Stable carbon and oxygen isotopic compositions were determined using a Thermo Gas Bench II device coupled to a Thermo Delta V Advantage mass spectrometer at the School of Archaeological Sciences, University of Bradford. δ13C and δ18O values were calibrated relative to VPDB. Measurement uncertainty was monitored using international reference standards: CO-1 (δ13C = 2.49 ± 0.03‰, δ18O = 2.40 ± 0.1‰), CO-8 (δ13C = − 5.76 ± 0.03‰, δ18O = − 22.7 ± 0.2‰) and NBS-19 (δ13C = 1.95‰, δ18O = −2.20. Precision (u(Rw)) was determined to be ± 0.33 for δ13C and 0.43 for δ18O on the basis of repeated measurements of calibration standards and check standards. Accuracy (u(bias)) was determined to be ± 0.21 for δ13C and 0.4 for δ18O on the basis of the difference between the observed and known δ values of the check standards and the long-term standard deviations of the check standards. Using the equations from Szpak et al. (2017), the total analytical uncertainty was estimated to be ± 0.39 for δ13C and ± 0.43 for δ18O.
All graphs presented in this paper were prepared using the software R (version 3.5.1).
Results
Stage 1: additional plant δ13C and δ15N values
Figure 3 shows all plant and animal δ13C and δ15N values from Kouphovouno, including previously published plant (n = 28) and animal (n = 68) samples (Vaiglova et al. 2014a). Table 3 presents the new raw data and Table 4 shows the summary statistics of the combined plant datasets. This new set of analyses revealed an even larger variability in cereal δ15N values than seen previously (δ15N range: 6.2‰ for free-threshing wheat, n = 17; 7.3‰ for barley, n = 15; 6.0‰ for einkorn, n = 6, see Fig. 4). When an einkorn outlier (KFO35), which lies in the middle of the sheep/goat cluster, is removed, the δ15N value range of einkorn is reduced to 2.6‰. With reported δ15N values below 0‰, this dataset provides some of the lowest plant δ15N values measured from any archaeological site to date. Only one additional sample of a pulse crop (grass pea) was available for analysis, which provided a significantly lower δ15N value (− 2.5‰) compared to the seven pea samples measured previously.

All plant and animal δ13C and δ15N values from Neolithic Kouphovouno. Samples KFO34 and KFO50 are not included, because they only yielded a reliable δ15N value. a Individual measurements, b means and standard deviations of each species. The dataset includes samples measured previously (28 plants and 68 animals, Vaiglova et al. 2014a); the previous plant measurements were corrected here using the more recent charring correction (see “Materials and methods”). Measurement error is shown in the bottom-right of a

All plant δ15N values from Kouphovouno. a All individual measurements, including KFO34 and KFO50, which are excluded from Fig. 3 because they did not produce reliable δ13C measurements. b Means and standard deviations of each species. The dataset includes samples measured previously (n = 28, Vaiglova et al. 2014a) and corrected using the more recent charring correction (see “Materials and methods”). Measurement error is shown in the bottom-right of a
Multivariate statistics were used to test the difference between the mean δ15N values of the species where n > 1 (free-threshing wheat, hulled barley, einkorn, pea). The einkorn outlier (KFO35) was not included in the analysis. The data are normally distributed (Shapiro-Wilk test, W = 0.95, p = 0.06), but not homogenous (Levene’s test, F(3,39) = 3.02, p = 0.041), so the non-parametric Kruskall-Wallis test was used. The results show that there are significant differences between the mean δ15N values of the four groups (H(3) = 27.21, p < 0.01) and a post hoc Bonferroni test reveals that the differences are between the free-threshing wheat and each of the other three species (p < 0.01 in all pairs), but not between the pairs of the other species (hulled barley and einkorn, p = 0.14, einkorn and pea, p = 0.13, hulled barley and pea, p = 1.0).
To calculate a measure of plant carbon discrimination irrespective of the concentration of atmospheric CO2, all δ13C values of all plant measurements were converted to Δ13C using the following equation (Farquhar et al. 1989):
The ranges in Δ13C values of the crops also increased with the additional measurements: 1.9‰ for free-threshing wheat, 2.7‰ for hulled barley, 4.4‰ for einkorn (see Fig. 5 and Table 4). The data for the four main species (where n < 1) are normally distributed (Shapiro-Wilk test, W = 0.95, p = 0.09) and homogenous (Levene’s test, F(3,38) = 1.12, p = 0.35), so an ANOVA test was used to assess statistically significant differences between the mean δ13C values of the four groups. The results indicate that there are significant differences (F(3,38) = 25.26, p < 0.01) and a post hoc Bonferroni test reveals that the differences are between free-threshing wheat and hulled barley (p < 0.01), free-threshing wheat and pea (p < 0.01) and einkorn and pea (p = 0.004).

All plant δ13C values from Kouphovouno. a All individual measurements, including KFO52, which is excluded from Fig. 3 because it did not produce a reliable δ15N measurement. b Means and standard deviations of each species. The dataset includes samples measured previously (n = 28, Vaiglova et al. 2014a) and corrected using the more recent charring correction (see “Materials and methods”). Measurement error is shown in the bottom-right of a
Figure 6 shows all plant and animal δ13C and δ15N values divided by chronological phase (free threshing wheat: MN = 14, LN = 3; hulled barley: MN = 8, LN = 7; einkorn: MN = 1 (outlier), LN = 5); pea: MN = 4, LN = 3). Barley is the only species represented equally in the two phases. The mean δ13C values of barley in the two periods are comparable, but the range is narrower in the Middle Neolithic (− 24.4 ± 0.3‰) compared to the Late Neolithic (− 24.2 ± 0.9‰). The δ15N values are similarly variable, but are overall higher in the MN (2.3 ± 2.0‰) compared to the LN (1.0 ± 1.8‰). A Student’s t test showed that there is no statistically significant difference between the mean δ15N values of MN and LN barley (t = − 1.35, p = 0.02).

All plant δ13C and δ15N values from Kouphovouno divided according to phase of occupation
Stage 2: tooth enamel carbonate δ13C and δ18O values
Figure 7 shows the sequential δ13C and δ18O values of sheep and goat teeth from Kouphovouno. Table 5 lists the individual measurements and Table 6 presents the summary statistics. δ13C values vary from − 13.7 to − 9‰ in sheep and − 13.3 to − 11.4‰ in goats, with mean intra-tooth variation of 3.2 (from 1.1 to 4.6‰) in sheep and 1.1‰ (from 0.4 to 1.6‰) in goats. δ18O values vary from − 5.3 to + 1.2‰ in sheep and − 4.0 to + 2.2‰ in goats, with mean intra-tooth variation of 3.7‰ (from 2.6 to 5.0‰) in sheep and 4.5‰ (from 3.1 to 5.3‰) in goats.

Sequential tooth enamel carbonate δ13C and δ18O values of sheep (a–d) and goats (e–h) from Middle (MN) and Late Neolithic (LN) levels at Kouphovouno. Positions of sub-samples are recorded as distances (in mm) from the enamel root junction (erj)
None of the animals exhibit notable consumption of C4 vegetation during the first year of their lives, as their δ13C values lie below the − 8‰ threshold for a mixed C3-C4 diet (cf Cerling et al. 1997) (see Fig. 8 with carbon sequences only). Three out of the four sheep exhibit sinusoidal intra-tooth carbon and oxygen isotopic sequences that reflect seasonal fluctuation in moisture and temperature, recording higher values in the summer and lower values in the winter (cf Gat 1980; Allison and Hughes 1983; Rozanski et al. 1993). The intra-tooth δ13C-value sequences of all four goats and one of the sheep (from the MN) are nearly flat. One sheep (KFO212 from the LN) records the highest δ13C values of all the individuals, but these values are still below the − 8‰ cut-off for C4 input in mixed diets (cf Cerling et al. 1997). This sheep also has the flattest intra-tooth δ18O-value sequence of all the animals measured: it has an amplitude (Δ18O value) of 2.6‰, while all the other individuals have Δ18O values between 3.1 and 5.0‰ (see Fig. 9 with oxygen sequences only).

Sequential tooth enamel carbonate δ13C values of sheep (a) and goats (b) from Middle (MN) and Late Neolithic (LN) levels at Kouphovouno. Measurement error is shown in the bottom-right of a

Sequential tooth enamel carbonate δ18O measurements of sheep (a) and goats (b) from Middle (MN) and Late Neolithic (LN) levels at Kouphovouno. Measurement error is shown in the bottom-right of a
Discussion
Previous stable isotopic analysis of plant and animal remains from Neolithic Kouphovouno indicated that farmers used distinct strategies for cultivating different species of cereals and that they did not herd ovicaprids (sheep and goats) together (Vaiglova et al. 2014a). The results raised more questions about the dynamics of a small-scale intensive management system, and these questions formed the basis of additional stable isotopic work carried out in this study. The following discussion will:
- (a)
draw on a larger plant stable isotopic dataset to expand our understanding of the range of cultivation regimes employed by the ancient farmers, and
- (b)
paint a more nuanced picture of the seasonal and multi-annual patterns of caprine grazing behavior.
Together, these narratives will further our understanding of the way Neolithic farmers at Kouphovouno made use of their environmental resources and adapted their food production systems to their culinary preferences and socio-economic choices.
Crop management
Based on a sample size of 13 free-threshing wheat and 7 hulled barley bulk charred grain samples, previous measurements of δ13C and δ15N values showed that throughout the Middle to Late Neolithic, these two cereals were not cultivated in the same soil conditions. Free-threshing wheat was consistently sown in more 15N-enriched soils. Because this distinction was species-specific rather than eco-systemic, it was argued that anthropogenic rather than natural factors were responsible for the differential soil enrichment (Vaiglova et al. 2014a). Notably, this distinction in δ15N values exists for free-threshing wheat and hulled barley samples that come from the same archaeological contexts, suggesting that the higher δ15N values were not caused by localized soil-diagenetic factors.
The most likely anthropogenic cause of soil enrichment was more intensive soil management, including application of farmyard manure, midden material, or other decomposing organic material, and tillage. The previous results suggest that while free-threshing wheat was cultivated using more intensive treatments, the soils in which barley grew may also have been manured/managed, but to a lesser extent than free-threshing wheat. As the δ15N values of the animals do not lie a trophic level above the values of the δ15N values of free-threshing wheat grain, it was argued that this crop was grown exclusively for human consumption. This left open the possibility that barley was cultivated (at least partially) as a fodder crop.
The expanded dataset provides further support for distinct treatment of free-threshing wheat and barley, but it reduces the uniformity of the soil conditions in which the two crops grew. In the original dataset, the range of Δ13C values of barley was narrow (1.2‰), and this was taken to suggest that this crop was grown under restricted water availability. With the addition of eight new barley samples, the range of Δ13C values increased to 2.6‰; the original indication of restricted soil wetness was thus a construct of the smaller sample size. Barley that was grown in rainfed conditions at modern-day organic farms in Morocco exhibited within-field ranges of values between 0.5‰ (Bellota farm) and 5.0‰ (Agda farm) (both farms receive 703 mm of rainfall per year) (Styring et al. 2016). This suggests that Δ13C values of crops grown under uniform environmental conditions can be variable, and that ranges below 2‰ should not necessarily be interpreted as indicative of more restricted water availability. Araus et al. (1997a, b) argue that there are physiological differences in 13C assimilation between wheat and barley, accounting for differences of 1–2‰ (with barley having higher Δ13C values than wheat grown under the same moisture availability). At Kouphovouno, the mean Δ13C values of barley (18.3 ± 0.7‰) are 1.7‰ lower compared to the mean δ13C values of free-threshing wheat (16.6 ± 0.5‰), suggesting that the two crops were grown under similar moisture availability, likely in rain-fed conditions.
The range of δ15N values of barley also increased with the addition of new samples, but none of the samples lie in the cluster where the majority of the free-threshing wheat sample lie (above + 4.9‰). While the difference between the δ15N values of free-threshing wheat and barley remains statistically significant, the new data provides further evidence that that free-threshing wheat was cultivated in more intensively managed soils. Similar higher-intensity management of free-threshing wheat was recorded at Aceramic Neolithic Knossos, Crete, situated in an environment that receives similar rainfall inputs as Kouphovouno in the Sparta Basin. The Neolithic samples from Knossos (n=3) had significantly higher δ15N values (between +5.1‰ to +5.7‰) compared to Late Bronze Age emmer samples from the same site (n=4, δ15N: +3.6‰ to +5.5‰) (Nitsch et al. 2019). Apart from the archaeobotanical assemblage from 7th millennium BC Knossos (Sarpaki 1995), free-threshing wheat does not occur in significant quantities in Neolithic contexts in Greece. At Kouphovouno, it mostly occurs in grain-rich deposits, suggesting that the farmers had distinct reasons to cultivate it in the first place. It is for this reason that they may have placed higher value on it, and cultivated it under higher management intensity.
Three new free-threshing wheat samples (one of which, KFO50, only yielded a δ15N measurement) fall below the + 4.9‰ line that separates the intensively grown free-threshing wheat and less intensively grown barley. This suggests that even though the main strategy was to cultivate the two crops separately under varying degrees of intensity, occasionally, the farmers sowed free-threshing wheat in the same soils as the barley, perhaps as a risk-buffering strategy.
Einkorn is a glume wheat that played an important role in the Neolithic farming systems of northern and southern Greece (Sarpaki 1995; Valamoti and Kotsakis 2007). At Kouphovouno, it was found in contexts generally dating to the Late Neolithic. The δ15N values of einkorn obtained in this study are systematically lower than all the δ15N values of free-threshing wheat. They overlap with some of the barley samples, but most of them are systematically lower still. With values reaching below 0‰, the results indicate that this crop was grown in even less 5N-enriched soils than barley, likely in soils that were not manured at all. It may have been intended as a fodder crop or to provide reserves (for both animals and humans) in case of shortages of the other crops.
Nowadays, wheat is considered more demanding in terms of its soil growing requirements than barley. It can withstand drier conditions, poorer soils and a degree of salinity (Zohary et al. 2012). Einkorn (also a wheat), is now often cultivated in soils with low nutrient quality, such as in modern farms in Provence, France (Bogaard et al. 2016). Considering these nutritional requirements, the fact that free-threshing wheat was consistently cultivated in more 15N-enriched soils compared to both hulled barley and einkorn suggests that the farmers were aware of similar ecological adaptations and catered to the crops’ distinct growing requirements. Plant stable isotopic analyses have shown, however, that this is not always the case. At the Late Bronze Age site of Archontiko in northern Greece, higher δ15N values of hulled barley suggest that this crop was grown under higher intensity compared to free-threshing wheat (Nitsch et al. 2017). This indicates that early production systems were not always optimized to the crops’ nutritional requirements, and other reasons, possibly stemming from culinary or economic preferences, played a role in determining the crop management of the different cereals.
The sample sizes of the crops investigated in this study are still too small to allow for robust investigation of chronological shifts in crop management between the Middle and the Late Neolithic at Kouphovouno. Free-threshing wheat is only represented by three samples in the Late Neolithic and einkorn is only represented by one sample in the Middle Neolithic; and the latter sample may be an outlier. Even though barley presents more samples for comparison and even though the mean δ15N values of barley from the two phases are different, there is no statistical significance to this difference. It is noteworthy that the samples which widen the range of δ13C values of barley as well as all the barley and einkorn samples whose δ15N values lie below 0‰ date to the Late Neolithic. This may mean that in the latter period of occupation, farmers had to seek new plots of land that were more distant and/or less naturally enriched in 15N. These soil conditions may have accentuated the differences in soil wetness on any given year. However, due to the limited numbers of samples in the two chronological groups, caution must be exercised when interpreting these differences.
Sheep and goat management
The mortality profiles of sheep and goats from Neolithic Kouphovouno suggest that their exploitation strategy either (1) changed from focus on secondary products in the Middle Neolithic to focus on meat procurement in the Late Neolithic or (2) the farmers practiced a mixed strategy during both periods of occupation (Cantuel et al. 2008). Previous bulk bone collagen δ13C and δ15N measurements (on a small number of individuals) indicated subtle chronological differences in multi-annual dietary averages of these two animals (Vaiglova et al. 2014a). The data showed significant differences in δ13C values between sheep and goats during the Middle Neolithic and significant differences between their δ15N values in the Late Neolithic. Although the Middle Neolithic trend is statistically significant (keeping in mind the small sample sizes), it is no longer considered worthy of discussion in this paper, because the mean δ13C values of the two animals differ by only 0.3‰. The Late Neolithic pattern of varying δ15N values, however, is still considered meaningful, as the difference between the average δ15N values is 1.1‰. What the bone collagen data thus show is that during the Middle Neolithic, the two ovicaprid species consumed isotopically similar diets, while in the Late Neolithic, they subsisted on vegetation variably enriched in 15N.
One possible explanation for the variable δ15N values is that in the Late Neolithic, the animals were kept in smaller herds closer to the arable landscape, and their dietary differences were thus a result of their variable browsing/grazing adaptations. Another possibility is that the animals grazed in distinct parts of the landscape (Vaiglova et al. 2014a). The assessment of seasonal dietary and grazing patterns of these animals provides an opportunity to not only better understand this chronological shift. It will also allow us to better understand the role of the animals in the subsistence economy of the Neolithic farmers.
The results of sequential tooth enamel carbonate δ13C and δ18O analyses indicate that sheep and goats at this site were managed differently in their first year of life. Two sheep from the Middle Neolithic exhibit sinusoidal δ13C and δ18O value sequences, which are reflective of seasonal fluctuations in moisture and temperature (Gat 1980; Allison and Hughes 1983; Rozanski et al. 1993; Hartman and Danin 2010). This suggests that they consumed fresh vegetation throughout the whole year, likely by grazing within the lowlands of the Sparta Basin. Both Middle Neolithic goats and the third Middle Neolithic sheep, on the other hand, exhibit flattened δ13C sequences. This indicates that they did not eat fresh local vegetation during all seasons of their first year of life. Modern sheep that have been fed on fodder in Mongolia exhibit similarly dampened intra-tooth δ13C-value sequences (Makarewicz 2014), and it is likely that the three Kouphovouno individuals with dampened δ13C value sequences spent part of the year eating vegetation that was collected during another season and stored for lean-season foddering. Because obligate drinkers like sheep and goats obtain most of their body oxygen from ingested water, and because the consumption of dry fodder necessitates the ingestion of additional drinking water, the δ18O sequences of these animals were not affected by their consumption of fodder. Their δ18O sequences fluctuate predictably given the changing temperatures between the wet and the dry seasons. For this reason, it is argued herein that all of the Middle Neolithic ovicaprids stayed within the lowland region during their first year of life.
During the Late Neolithic, part of the picture changes, due to limited preservation, only one sheep from the Late Neolithic was available for analysis. This individual (KFO212) has the widest range of δ13C values and the lowest range of δ18O values of all the animals analyzed in this study. The dampened δ18O sequence suggests that the animal moved to a different altitude for part of the year, where it gained access to vegetation with more 13C-enriched values (which was not available to the animals that stayed in the Sparta Basin). It may have spent part of the year at a location either closer to the coast or in the Taygetos mountains. The seasonal dietary patterns of the goats did not change in the Late Neolithic. Their flat δ13C-value sequences indicate that the animals continued to be foddered during the latter part of the site’s occupation.
Overall, the findings show that while goats were managed the same way (in terms of isotopic composition of their diets) during both periods of occupation (they were kept close to the site and foddered presumably during the dry season), sheep management was more variable. In the Middle Neolithic, some sheep were herded in the surrounding Sparta Basin, while one individual was foddered. In the Late Neolithic, the one sheep available for analysis moved to a different location for part of the year.
Balasse and Ambrose (2005) argue that because it is so costly and labor-intensive, farmers only resort to foddering if it is necessary or facilitated. The geomorphological reconstruction of the landscape around Kouphovouno shows that water was abundant in the Sparta Basin during the Neolithic period (Fouache et al. 2007). The fertile soils in this region would have thus offered plentiful vegetation for livestock grazing, making foddering not necessary. If the soils were less productive (N-poor) than the environmental model suggests, pasture areas would have been limited, and this would be consistent with the observation that crops in the Late Neolithic grew in poorer soils, given δ15N values below 0‰. However, goats—being browsers—are able to subsist on woody vegetation and survive in dry and marginal areas without grass cover. Sheep, on the other hand, are grazers and need more nutritious grasses to survive. So if fresh browse was limited, the expectation would be that goats were allowed to graze on woody vegetation in the surrounding areas. As this is not the case, the hydrologically active environmental model cannot be disputed.
In the absence of an environmental explanation, the reason why the goats (and some sheep) were foddered may thus have been socio-economic: for the procurement of milk. Foddering may have been facilitated by intentional cultivation of barley and/or einkorn as fodder crops. The animals that were kept close to the site may have consumed these crops directly from the fields during the growing seasons, or as harvested fodder during the off-season.
The high-resolution stable isotopic results suggest that the differential management of sheep and goats was likely not dictated by possible shortages in fresh graze in the local landscape. Instead, it was suited to the procurement of milk from goats (which were kept close to the site all year round) and procurement of meat from sheep (which were taken to local and more distant pasture for fattening). The indication that sheep grazed in different parts of the landscape (including more distant parts either closer to the coast or at higher altitudes) can explain the small differences in bulk collagen δ15N values of Late Neolithic ovicaprids. The results of the isotopic analyses support the zooarchaeological interpretation that farmers at Kouphovouno practiced a mixed primary and secondary product exploitation strategy throughout the Neolithic occupation of the site.
Conclusions
The findings from this study show that farmers at Neolithic Kouphovouno developed agropastoral management strategies that were not uniform or haphazard. Instead, they were the result of intentional decisions attuned to culinary preferences, environmental opportunities and attempts to reduce risk of crop failure.
The three main cereal crops cultivated at Kouphovouno – free-threshing wheat, hulled barley and einkorn – were grown under distinct cultivation treatments, and the choice of cultivation treatments was attuned to the crops’ ecological adaptations. Free-threshing wheat has higher soil nutrient and water requirements than barley and einkorn. The Neolithic farmers thus managed this crop at a higher intensity (in terms of labor input per square area) than barley and einkorn. Intensive cultivation was likely carried out within 1 km of the settlement and increased the 15N enrichment of the soils. Einkorn was grown in the most 15N-depleted soils, suggesting that the farmers did not make any effort to increase the crop yields of this cereal, and may have cultivated it as a risk buffer. This species-specific crop management system was thus built around notions of higher value ascribed to free-threshing wheat (grown exclusively for human consumption) and lower value assigned to the other cereals (possibly grown as a fodder crop).
Results of sequential tooth enamel carbonate δ13C and δ18O values provide direct evidence that foddering was indeed an integral component of the pastoral management strategy. While goats were foddered during both phases of occupation, the management of sheep was more variable. One sheep from the Middle Neolithic was foddered, two individuals from the MN grazed on fresh vegetation in the surrounding valley and one individual from the Late Neolithic partook in trans-altitudinal mobility in search of fresh vegetation during the dry season. The sample sizes of these groups are extremely limited, so caution must be exercised with interpreting any chronological change in sheep management. Nevertheless, the conclusion can be drawn that the management of sheep and goats was distinct. The difference in early life seasonal management explains the subtle differences in the bulk bone collagen δ15N values of ovicaprids (Vaiglova et al. 2014a) and provides support for the explanation that the differences in the multi-annual diets during the Late Neolithic were the result of distinct management strategies rather than varying dietary adaptations.
In addition to providing nuanced information about the dietary behavior of these two animals, the analysis presented herein furthers our understanding of the roles of the two animals in the Neolithic subsistence strategy. Analysis of animal mortality profiles led to inconclusive results, suggesting that sheep and goat exploitation strategies may have changed between the MN and the LN, or that they were mixed in both phases of occupation. The findings from this study – which show that goats were confined to the settlement likely for supplying milk, while sheep were herded on fresh vegetation for fattening prior to slaughter for meat – support the latter scenario. Overall, the integrated stable isotopic analyses of the archaeological plant and animal remains from Neolithic Kouphovouno indicate not only that agropastoral management was flexible and diverse, but that the diversity in management was driven by cultural choices favoring consumption of free-threshing wheat and goat milk.
References
Allison GB, Hughes MW (1983) The use of natural tracers as indicators of soil-water movement in a temperate semi-arid region. J Hydrol 60:157–173
Ambrose SH (1991) Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs. J Archaeol Sci 18:293–317
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In: Lambert JB, Grupe G (eds) Prehistoric human bone: archaeology at the molecular level. Springer-Verlag, Berlin, pp 1–37
Araus J, Febrero A, Buxó R et al (1997a) Identification of ancient irrigation practices based on the carbon isotope discrimination of plant seeds: a case study from the south-East Iberian Peninsula. J Archaeol Sci 24:729–740
Araus JL, Febrero A, Buxó R et al (1997b) Changes in carbon isotope discrimination in grain cereals from the North-Western Mediterranean Basin during the past seven millennia. Palaeoenvironmental evidence of a differential change in aridity during the late Holocene. Glob Chang Biol 3:107–118
Balasse M (2002) Reconstructing dietary and environmental history from enamel isotopic analysis: time resolution of intra-tooth sequential sampling. Int J Osteoarchaeol 12:155–165
Balasse M, Ambrose SH (2005) Distinguishing sheep and goats using dental morphology and stable carbon isotopes in C4 grassland environments. J Archaeol Sci 32:691–702
Balasse M, Bocherens H, Mariotti A (1999) Intra-bone variability of collagen and apatite isotopic composition used as evidence of a change of diet. J Archaeol Sci 26:593–598
Balasse M, Bocherens H, Mariotti A, Ambrose SH (2001) Detection of dietary changes by intra-tooth carbon and nitrogen isotopic analysis: an experimental study of dentine collagen of cattle (Bos taurus). J Archaeol Sci 28:235–245
Balasse M, Ambrose SH, Smith AB, Price TD (2002) The seasonal mobility model for prehistoric herders in the south-western cape of South Africa assessed by isotopic analysis of sheep tooth enamel. J Archaeol Sci 29:917–932
Balasse M, Boury L, Ughetto-Monfrin J, Tresset A (2012) Stable isotope insights (δ18O, δ13C) into cattle and sheep husbandry at Bercy (Paris, France, 4th millennium BC): birth seasonality and winter leaf foddering. Environ Archaeol 17:29–44
Balasse M, Bălăşescu A, Janzen A et al (2013) Early herding at Măgura-Boldul lui Moş Ivănuş (early sixth millennium BC, Romania): environments and seasonality from stable isotope analysis. Eur J Archaeol 16:221–246
Bateman AS, Kelly SD, Jickells TD (2005) Nitrogen isotope relationships between crops and fertilizer: implications for using nitrogen isotope analysis as an indicator of agricultural regime. J Agric Food Chem 53:5760–5765
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Bocherens H, Mashkour M, Billiou D et al (2001) A new approach for studying prehistoric herd management in arid areas: intra-tooth isotopic analyses of archaeological caprine from Iran. Earth Planet Sci 332:67–74
Bogaard A (2004) The nature of early farming in central and south-East Europe. Doc Praehist 31:49–58
Bogaard A (2005) ‘Garden agriculture’ and the nature of early farming in Europe and the near east. World Archaeol 37:177–196
Bogaard A, Heaton THE, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J Archaeol Sci 34:335–343
Bogaard A, Fraser R, Heaton THE, Wallace M, Vaiglova P, Charles M, Jones G, Evershed RP, Styring AK, Andersen NH, Arbogast RM, Bartosiewicz L, Gardeisen A, Kanstrup M, Maier U, Marinova E, Ninov L, Schäfer M, Stephan E (2013) Crop manuring and intensive land management by Europe’s first farmers. Proc Natl Acad Sci 110:12589–12594. https://doi.org/10.1073/pnas.1305918110
Bogaard A, Styring A, Ater M, et al (2016) From Traditional Farming in Morocco to Early Urban Agroecology in Northern Mesopotamia: Combining Present-day Arable Weed Surveys and Crop Isotope Analysis to Reconstruct Past Agrosystems in (Semi-) arid Regions From Traditional Farming in Morocco to Earl. Environ Archaeol 1–20. https://doi.org/10.1080/14614103.2016.1261217
Bowen GJ, Wilkinson B (2002) Spatial distribution of δ18O in meteoric precipitation. Geology 30:315–318
Britton K, Grimes V, Dau J, Richards MP (2009) Reconstructing faunal migrations using intra-tooth sampling and strontium and oxygen isotope analyses: a case study of modern caribou (Rangifer tarandus granti). J Archaeol Sci 36:1163–1172
Brown WAB, Christofferson PV, Massler M, Weiss MB (1960) Postnatal tooth development in cattle. Am J Vet Res 21:7–34
Bryant JD, Froelich PN, Showers WJ, Genna BJ (1996) Biologic and climatic signals in the oxygen isotopic composition of Eocene-Oligocene equid enamel phosphate. Palaeogeogr Palaeoclimatol Palaeoecol 126:75–89
Buckley M, Whitcher Kansa S, Howard S et al (2010) Distinguishing between archaeological sheep and goat bones using a single collagen peptide. J Archaeol Sci 37:13–20
Cantuel J, Gardeisen A, Renard J (2008) L’exploitation de la faune durant le Néolithique dans le bassin Égéen. In: Vila E, Gourichon L, Choyke AM, Buitenhuis H (eds) Archaeozoology of the near east VIII. Maison de l’Orient et de la Méditerranée, Lyon, pp 279–298
Cavanagh W, Renard J (2014) Neolithic figurines from Kouphovouno. J Prehist Relig 24:19–35
Cavanagh W, Mee C, Renard J (2004) Sparta before Sparta: report on the Intensive Survey at Kouphovouno 1999--2000. Annu Br Sch Athens 99:49–128
Cavanagh W, Mee C, Renard J (2007) Excavations at Kouphovouno. Annu Br Sch Athens 102:11–101
Cerling T, Harris J (1999) Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and palaecological studies. Oecologia 120:347–363
Cerling TE, Harris JM, MacFadden BJ et al (1997) Global vegetation change through the Miocene/Pliocene boundary. Nature 389:153–158
Cleveland WS (1979) Robust locally weighted regression and smoothing scatterplots. J Am Stat Assoc 74:829–836
Collins MJ, Nielsen-Marsh CM, Hiller J et al (2002) The survival of organic matter in bone: a review. Archaeometry 44:383–394
Craine JM, Elmore AJ, Aidar MPM, Bustamante M, Dawson TE, Hobbie EA, Kahmen A, Mack MC, McLauchlan K, Michelsen A, Nardoto GB, Pardo LH, Peñuelas J, Reich PB, Schuur EA, Stock WD, Templer PH, Virginia RA, Welker JM, Wright IJ (2009) Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytol 183:980–992
Dansgaard W (1964) Stable isotopes in precipitation. Tellus 16:436–468
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta 42:495–506
Ehleringer JR, Sage R, Flanagan LB, Pearcy RW (1991) Climate change and the evolution of C4 photosynthesis. Trends Ecol Evol 6:95–99
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annu Rev Plant Physiol Plant Mol Biol 40:503–437
Ferrio JP, Araus JL, Buxó R et al (2005) Water management practices and climate in ancient agriculture: inferences from the stable isotope composition of archaeobotanical remains. Veg Hist Archaeobotany 14:510–517
Fouache E, Cosandey C, Renard J et al (2007) Les ressources en eau dans le bassin de Sparte au Néolithique: les contextes géomorphologique et hydrologique du site archéologique de Kouphovouno. Environnements et Sociétés 1–6
Fraser R, Bogaard A, Heaton T et al (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38:2790–2804
Fraser R, Bogaard A, Charles M et al (2013) Assessing natural variation and the effects of charring, burial and pre-treatment on the stable carbon and nitrogen isotope values of archaeobotanical cereals and pulses. J Archaeol Sci 40:4754–4766
Gat JR (1980) The isotopes of hydrogen and oxygen in precipiation. In: Fritz P, Fontes JC (eds) Handbook of environmental isotope geochemistry. Elsevier, Amsterdam, pp 21–42
Halstead P (1981) Counting sheep in Neolithic and bronze age Greece. In: Pattern of the past: studies in honour of David Clarke. Cambridge University Press, Cambridge, pp 307–339
Halstead P (2011) Farming, material culture and ideology: repackaging the Neolithic of Greece (and Europe). In: Hadjikoumis A, Viner S (eds) Dynamics of Neolithisation in Europe: studies in honour of Andrew Sherratt. Oxbow Books, Oxford, pp 131–151
Hare PE (1980) Organic geochemistry of bones, and its relation to the survival of bone in the natural environment. In: Fossils in the making. The University of Chicago Press, Chicago, pp 208–219
Hartman G, Danin A (2010) Isotopic values of plants in relation to water availability in the eastern Mediterranean region. Oecologia 162:837–852
Heaton THE, Vogel JC, von la Chevallerie G, Collett G (1986) Climatic influence on the isotopic composition of bone nitrogen. Nature 322:822–823
Hedges REM, Reynard LM (2007) Nitrogen isotopes and the trophic level of humans in archaeology. J Archaeol Sci 34:1240–1251
Henton E, Meier-Augenstein W, Kemp HF (2010) The use of oxygen isotopes in sheep molars to investigate past herding practices at the Neolithic settlement of Çatalhöyük, Central Anatolia. Archaeometry 3:429–449
Hillson S (2005) Teeth, 2nd edn. Cambridge University Press, Cambridge
Hobbie EA, Högberg P (2012) Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen dynamics. New Phytol 196:367–382
Hoefs J (2009) Stable isotope geochemistry. Springer, New York
Högberg P (1997) 15N natural abundance in soil-plant systems. New Phytol 137:179–203
Indermühle A, Stocker TF, Joos F et al (1999) Holocene carbon-cycle dynamics based on CO2 trapped in ice at Taylor dome, Antarctica. Nature 398:121–126
Kirsanow K, Makarewicz C, Tuross N (2008) Stable oxygen (δ18O) and hydrogen (δD) isotopes in ovicaprid dentinal collagen record seasonal variation. J Archaeol Sci 35:3159–3167
Krueger HW, Sullivan CH (1984) Models for carbon isotope fractionation between diet and bone. In: Turnlund JR, Johnson PE (eds) Stable isotopes in nutrition. American Chemical Society, Washington, DC, pp 205–222
Land LS, Lundelius EL, Valastro S (1980) Isotopic ecology of deer bones. Palaeogeogr Palaeoclimatol Palaeoecol 32:143–151
Lee-Thorp JA (2008) On isotopes and old bones. Archaeometry 50:925–950
Lee-Thorp JA, Sealy JC, van der Merwe NJ (1989) Stable carbon isotope ratio differences between bone collagen and bone apatite, and their relationship to diet. J Archaeol Sci 16:585–599
Leuenberger M, Siegenthaler U, Langway C (1992) Carbon isotope composition of atmospheric CO2 during the last ice age from an Antarctic ice core. Nature 357:488–490
Longinelli A (1984) Oxygen isotopes in mammal bone phosphate: a new tool for paleohydrological and paleoclimatological research? Geochim Cosmochim Acta 48:385–390
Longinelli A, Selmo E (2003) Isotopic composition of precipitation in Italy: a first overall map. J Hydrol 270:75–88
Luz B, Kolodny Y, Horowitz M (1984) Fractionation of oxygen isotopes between mammalian bone-phosphate and environmental drinking water. Geochim Cosmochim Acta 48:1689–1693
Makarewicz CA (2014) Winter pasturing practices and variable fodder provisioning detected in nitrogen (δ15N) and carbon (δ13C) isotopes in sheep dentinal collagen. J Archaeol Sci 41:502–510
Makarewicz CA, Arbuckle BS, Oztan A (2017) Vertical transhumance of sheep and goats identified by intra-tooth sequential carbon (δ13C) and oxygen (δ18O) isotopic analyses: evidence from chalcolithic Köşk Höyük, Central Turkey. J Archaeol Sci 86:68–80
Mee C, Cavanagh B, Renard J (2014) The middle–late Neolithic transition at Kouphovouno. Annu Br Sch Athens 1–31
Minagawa M, Wada E (1984) Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochim Cosmochim Acta 48:1135–1140
Nitsch EK, Charles M, Bogaard A (2015) Calculating a statistically robust δ13C and δ15N offset for charred cereal and pulse seeds. Sci Technol Archaeol Res 1:1–8
Nitsch E, Andreou S, Creuzieux A, et al (2017) A bottom-up view of food surplus: using stable carbon and nitrogen isotope analysis to investigate agricultural strategies and diet at Bronze Age Archontiko and Thessaloniki Toumba , northern Greece. World Archaeol 1–34
Nitsch EK, Jones G, Sarpaki A, et al (2019) Farming Practice and Land Management at Knossos, Crete: New Insights from d13C and d15N Analysis of Neolithic and Bronze Age Crop Remains. In: Garcia D, Orgeolet R, Pomadere M, Zurbach J (eds) Country in the City: Agricultural Functions in Protohistoric Urban Settlements (Aegean and Western Mediterranean). Archaeopress, Oxford, pp 152–168
O’Connell TC, Kneale CJ, Tasevska N, Kuhnle GGC (2012) The diet-body offset in human nitrogen isotopic values: a controlled dietary study. Am J Phys Anthropol 149:426–434
Payne S (1973) Kill-off patterns in sheep and goats: the mandibles from Asvan kale. Anatol Stud 23:281–303
Rivals F, Gardeisen A, Cantuel J (2011) Domestic and wild ungulate dietary traits at Kouphovouno (Sparta, Greece): implications for livestock management and paleoenvironment in the Neolithic. J Archaeol Sci 38:528–537
Rozanski K, Araguás-Araguás L, Gonfiantini R (1993) Isotopic patterns in modern global precipitation. In: Swart PK, Lohmann KC, McKenzie J, Savin S (eds) Climate change in continental isotopic records. American Geophysical Union, Washington, DC, pp 1–36
Sarpaki A (1995) Toumba Balomenou, Chaeronia: plant remains from the early and middle Neolithic levels. In: Kroll R, Pasternak R (eds) Res Archaeobotanicae, international workgroup for Palaeoethnobotany, proceedings of the 9th symposium. Oetker-Vosges, Kiel, pp 5–15
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639
Sharp Z (2007) Principles of stable isotope geochemistry. Pearson Education, Upper Saddle River
Snoeck C, Pellegrini M (2015) Comparing bioapatite carbonate pre-treatments for isotopic measurements: part 1–impact on structure and chemical composition. Chem Geol 417:349–403
Styring AK, Ater M, Hmimsa Y et al (2016) Disentangling the effect of farming practice from aridity on crop stable isotope values: a present-day model from Morocco and its application to early farming sites in the eastern Mediterranean. Anthropol Rev 3:2–22
Szpak P (2014) Complexities of nitrogen isotope biogeochemistry in plant-soil systems: implications for the study of ancient agricultural and animal management practices. Front Plant Sci 5:1–19
Szpak P, Metcalfe JZ, Macdonald RA (2017) Best practices for calibrating and reporting stable isotope measurements in archaeology. J Archaeol Sci Rep 13:609–616
Tiedje JM, Sexstone AJ, Myrold DD, Robinson JA (1982) Denitrification: ecological niches, competition and survival. Antonie Van Leeuwenhoek 48:569–583
Tieszen LL (1991) Natural variations in the carbon isotope values of plants: implications for archaeology, ecology and palaeoecology. J Archaeol Sci 18:227–248
Tieszen LL, Fagre T (1993) Effect of diet quality and composition on the isotopic composition of respiratory CO2, bone collagen, bioapatite, and soft tissues. In: Lambert JB, Grupe G (eds) Prehistoric human bone: archaeology at the molecular level. Springer-Verlag, Berlin, pp 121–155
Towers J, Gledhill A, Bond J, Montgomery J (2014) An investigation of cattle birth seasonality using δ13C and δ18O profiles within first molar enamel. Archaeometry 56:208–236
Vaiglova P, Bogaard A, Collins M et al (2014a) An integrated stable isotope study of plants and animals from Kouphovouno, southern Greece: a new look at Neolithic farming. J Archaeol Sci 42:201–215
Vaiglova P, Snoeck C, Nitsch E, Bogaard A, Lee-Thorp J (2014b) Impact of contamination and pre-treatment on stable carbon and nitrogen isotopic composition of charred plant remains. Rapid Commun Mass Spectrom 28:2497–2510
Valamoti SM, Jones G (2003) Plant diversity and storage at Mandalo, Macedonia, Greece: Archaeobotanical evidence from the final Neolithic and early bronze age. Annu Br Sch Athens 98:1–35
Valamoti SM, Kotsakis K (2007) Transitions to agriculture in the Aegean: the archaeobotanical evidence. In: Colledge S, Connoly J (eds) Origins and spread of domestic plants in Southwest Asia and Europe. Left Coast Press, Walnut Creek, pp 75–91
van der Merwe NJ, Vogel JC (1978) 13C content of human collagen as a measure of prehistoric diet in woodland North America. Nature 276:815–816
Vogel JC, van der Merwe NJ (1977) Isotopic evidence for early maize cultivation in New York state. Am Antiq 42:238–242
Weinreb MM, Sharav Y (1964) Tooth development in sheep. Am J Vet Res 25:891–908
Zohary D, Hopf M, Weiss E (2012) Domestication of plants in the Old World: the origin and spread of domesticated plants in south-West Asia, Europe, and the Mediterranean Basin, 4th edn. Oxford University Press, Oxford
Acknowledgements
We wish to thank the British School at Athens and the École française d’Athènes for their support of the excavations carried out at Kouphovouno, the Centre national de la recherche scientifique for supporting the faunal analysis, Maria Theodosi from the Ephorate of Antiquities of Laconia for helping with the identification and sampling of teeth and Erika Nitsch for writing the R normalization script. We are grateful to Peter Ditchfield and Andy Gledhil for assistance with the isotopic analyses. The research was generously supported by the Oxford University Press, the American School of Classical Studies at Athens and the European Research Council. This paper is dedicated to Chris Mee, one of the directors of the Kouphovouno project, who was always very supportive and excited about the isotopic analyses.
Funding
This research was made possible by the financial support of the Oxford University Press (Clarendon award for doctoral study, awarded to P. Vaiglova), the American School of Classical Studies at Athens (Malcolm H. Wiener Laboratory Predoctoral Fellowship awarded to P. Vaiglova) and the European Research Council starting grant (awarded to A. Bogaard, grant number 312785).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Vaiglova, P., Gardeisen, A., Buckley, M. et al. Further insight into Neolithic agricultural management at Kouphovouno, southern Greece: expanding the isotopic approach. Archaeol Anthropol Sci 12, 43 (2020). https://doi.org/10.1007/s12520-019-00960-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-019-00960-y