
Abstract
The continuity of occupation at the settlement of Prague, Czech Republic, from the ninth to fourteenth centuries makes it a good case study for investigating the evolution of animal husbandry practices during the Early and High Middle Ages. Previous archaeozoological work shows that people’s dietary behaviour in Prague underwent a transformation at the beginning of the High Middle Ages (thirteenth century). This change has been attributed to large-scale socioeconomic transformations, which may have affected animal management practices. Stable carbon (δ13C) and nitrogen (δ15N) isotope analysis of the tooth dentine collagen of livestock (cattle, domestic pigs, sheep and goats) from three different areas in Prague (the Lesser Town, Prague Castle and the Old Town) presented in this study provides support for this hypothesis. The range of stable isotope values increased over time, which may, in addition to a farm-household model, indicate a more advanced system of livestock supply–for example, through urban markets–and may also be explained by more diversified breeding conditions in the town and its surroundings. The results show an increase in the δ15N values in pig diet over time and suggest that their management changed. This difference may have been caused by increased animal protein intake or higher δ15N value of the dietary sources. It has also been corroborated that the root dentine collagen of High Medieval cattle has slightly higher average δ15N values when compared to cattle from the earlier period, which could be related to gradual changes in landscape utilization and soil processing, such as fertilization.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Prague, Czech Republic, is among the most important archaeological sites in East-Central Europe. Its position has been brought about not only by a continuity of settlement that has lasted for more than a thousand years but mainly by its continuous role as a central meeting place–the Early Medieval seat of the Bohemian dukes and later Kings (Frolík and Smetánka 1997). Ever-increasing archaeological and historical research has gradually revealed how Prague started as an early urban settlement that grew into a High Medieval town characterized by new legal and territorial principles (Klápště 2016).
At the end of the Early Middle Ages in Bohemia (end of the twelfth century), the original system of country administration collapsed, changes took place in society and new rules for the promotion of power were established. Subsequently, during the High Middle Ages, there was more intense density of settlement, related to the founding of towns and villages as well as the restructuring of older settlement units. The original, traditional relations to allotments, which were now delimited, and which gave their owners discretions and duties, fundamentally changed. In this way, at the beginning of the High Middle Ages, in the thirteenth century, Bohemia underwent a significant transformation, the so-called Medieval transformation, consisting of gradual changes in cultural, political and economic aspects of society (Klápště 2012). The Medieval transformation took place in different parts of Bohemia asynchronously and with varying degrees of intensity and is very poorly documented by written sources (for details, see Žemlička 1997, 2002). In Prague, however, this transformation is well documented. As a result of multiple written and archaeological sources, it is possible to investigate the changes to everyday life, craft production and presence of ‘ethnic group’ settlements in Prague on both a chronological and a spatial scale.
The manifestations of the Medieval transformation included changes such as tool improvement, varied crop production methods (the three-field system), the adoption of soil management strategies (e.g. soil fertilization/manuring) and deforestation (Petráň and Buchvaldek 1985; Sádlo et al. 2008; Klápště 2012; Kuneš and Abraham 2017). Archaeological analyses suggest the use of stables and cowsheds at farmsteads from the High Middle Ages. Greater use of long, two-handed scythes as well as meadows is evidenced by historical, archaeological and pollen analyses (Klápště 2012).
The Medieval transformation in Central Europe, and in particular its impact on the natural environment, can be studied not only through archaeological and historical lenses but also using archaeobotanical techniques (macro- and microscopic). For example, results of these studies indicate the increasing eutrophication of the environment in High Medieval Prague, as well as a decline in plant species variability (Kozáková 2008). Some results document the change in the consumption of animal products that occurred at the beginning of the thirteenth century as well (Kovačiková et al. 2019).
Faunal material provides an opportunity to assess the change in the economic development of Medieval Prague. These investigations have shown that livestock breeding formed one of the essential components of the Medieval economy of Bohemia (Peške 1985). Most archaeozoological research into assemblages from Medieval Prague has focused on the basic characteristics of the osteological material obtained from particular sites. Studies that evaluate the system of livestock management are rarer; so too are studies that apply a diachronic approach that would allow the determination of long-term trends in breeding strategies (Kyselý 2015; Sůvová et al. 2018).
This study aims to investigate the variability of breeding strategies and animal husbandry management employed in Medieval Prague by combining the results of stable isotope analyses and the results of archaeozoological studies.
Appropriate consideration shall, therefore, be given to the following objectives:
-
(1)
Reconstruction of the dietary and feeding regimes of livestock in Early and High Medieval Prague, based on the stable carbon and nitrogen isotope values preserved in samples from selected sites
-
(2)
Identification of changes in livestock breeding strategies in relation to broader changes in the landscape associated with the Medieval transformation
The archaeological context of Medieval Prague
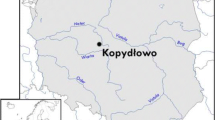
In the late eighth/early ninth century, a fortified settlement was built on the left bank of the Vltava River in the area of today’s Lesser Town (Fig. 1). The fortification enclosed a small area (of about seven hectares) along the riverbank, which probably controlled local fords (Čiháková 2018).

Location of Medieval sites from the Czech Republic mentioned in the study–(1) Malostranské Square No. 2/III (Lesser Town, ninth to eleventh centuries); (2) Prague Castle, Northern Promontory (Prague Castle, twelfth to thirteenth centuries); (3) Křižovnická St. No. 71/I (Old Town, thirteenth to fourteenth centuries): (4) Pařížská ul. No. 205/V (Old Town, thirteenth century)
By the mid-ninth century, the settlement structure had changed significantly, and the Hradčany (Hradschin) promontory came to the fore when the ruling family of the Přemyslids chose it as their principal residence. The first fortification was situated at the location where the Prague Castle (Fig. 1) was later built and dates to the second half of the ninth century. The ducal residence was first fortified with an earth and wood rampart, probably at the end of the reign of Duke Spytihnev I (†915 AD), while the original fortified area in the Lesser Town was significantly enlarged around 900 AD. The new precinct, together with the fortified Prague Castle, formed the seat of the Přemyslid dynasty. In the tenth century, the Early Medieval town of Prague became an important crossroads of European trade, and between 1135 and 1185 AD, the original stronghold was transformed into a Romanesque castle with stone walls (Boháčová and Herichová 2009).
After the eleventh century, a permanent settlement began to grow on the Old Town riverbank, with a predominance of craft and business activities. Expansion of the Old Town residential complex (Fig. 1) began once the central marketplace shifted to the area of today’s Old Town Square, probably at the beginning of the twelfth century (Hrdlička 2000). The absence of Romanesque churches and the preliminary results of archaeological research in the northern part of the Old Town suggest that the local Jewish community inhabited this area (where the High Medieval Jewish Quarter was located), probably from as early as the beginning of the twelfth century. The sprawling development of the Old Town settlement agglomeration culminated in the emergence of the High Medieval town in the mid-thirteenth century during the reign of Wenceslas I (1230–1253 AD).
The archaeozoological background of Medieval Prague
The taxonomic data and mortality profiles for the osteological assemblages used in this study have already been published (Kovačiková et al. 2019). The earlier phase of the Medieval settlement of Prague represented by the Lesser Town (ninth to eleventh centuries) and Prague Castle (twelfth to thirteenth centuries) is characterized by a high proportion of finds (NISP, Number of Identified Specimens) of domestic pig (Sus domesticus; Lesser Town: 63.7% of total NISP, Prague Castle: 49.0%; Fig. 2). The slaughter patterns for this species derived from both sites corroborate different trends in the meat production. While most animals in the earlier Lesser Town were slaughtered before the age of 18 months (53.6% of teeth; Fig. 3a), up in the later Prague Castle, the slaughter age varied between 18 and 24 months (52.6%; Fig. 3a). The representation of individuals in different age categories (suckling/weaning pigs, market pigs providing fresh/high yield meat and older animals culled from the breeding stock; Fig. 3a) seems to indicate that some animals were raised within the stronghold and were slaughtered for local consumption. At the same time, the marked predominance of male individuals attests to a consumer economy. Based on the character of both sites, it may be assumed that the strategies were combined.

Faunal spectra for different Medieval sites in Prague. The Number of Identified Specimens (NISP) totals are stated in parentheses

The survival curves based on dentition for pigs (a), cattle (b) and caprines (c) in the Medieval assemblages from Prague. As age classes feature different time spans, the relative frequencies established have been corrected according to the size of the time span of each age class. Numbers in parentheses indicate the number of teeth
The second most abundant species in both assemblages is cattle (Bos taurus; Lesser Town: 14.4% NISP, Prague Castle: 20.0%; Fig. 2). The kill-off pattern of cattle in the Lesser Town is indicative of secondary production, as more than half of the individuals lived for more than 6 years (70.0% of total teeth; Fig. 3b). The number of bones from sheep/goats (Ovis/Capra; Lesser Town: 9.8% of NISP, Prague Castle: 9.7%) is low in both assemblages (Fig. 2).
Assemblages from the Old Town (thirteenth century) are of a different character from the assemblages from the Lesser Town and Prague Castle, probably due to the greater wealth of the inhabitants of the High Medieval town (Klápště 2016). The Old Town assemblages have in common a higher proportion of bones from cattle (Křižovnická St.: 39.0% of NISP; Pařížská St.: 49.4% Fig. 2), which were slaughtered predominantly after 3 years of age at the latest (Křižovnická ul.: 56.2% of NISP, Pařížská St.: 69.2%; Fig. 3b). Pigs were slaughtered between 1.5 and 2 years of age (Křižovnická St.: 60.0% of teeth; similar results were obtained for the latter group originating from Prague Castle; Fig. 3a). In some assemblages, pig bones are less numerous (Křižovnická ul.: 27.1% of NISP, Pařížská St.: 4.2%; Fig. 2), and caprine finds are more common (Křižovnická St.: 23% of NISP, Pařížská St.: 4.2%; Fig. 2). The slaughter age of sheep and goats in the Old Town (Křižovnická St.; Fig. 3c) indicates that they were used in several ways, with a stronger emphasis on secondary production. These caprines were slaughtered at between 6 months and 2 years (30.2%) and over 4 years of age (41.5%).
Principles of stable isotope analysis
The stable carbon and nitrogen isotope composition of the bone and tooth dentine collagen provides information about past animal diets and landscape use (Fisher and Thomas 2012; Hamilton and Thomas 2012; Hammond and O’Connor 2013; Millard et al. 2013; Alexander et al. 2019). δ13C values are primarily determined by the photosynthetic pathway that plants–which were consumed by the animals–use to fix CO2 (DeNiro and Epstein 1981; Tieszen et al. 1983). δ15N values are indicative of the trophic level of individuals on the food chain (Schoeninger and DeNiro 1984). The mean value for C3 plants is − 27‰ (Smith and Epstein 1971), with a range from − 20 in dry and open habitats to − 37‰ in shaded forest habitats affected by the canopy effect (Drucker et al. 2008; Kohn 2010). Bone collagen δ13C values of herbivores grazing C3 plants in north-western Europe are around − 21.5‰ (Millard et al. 2013). The non-indigenous crops and weeds that employ the C4 photosynthetic mechanism, such as common millet (Panicum miliaceum), foxtail millet (Setaria italica) and barnyard grass (Echinochloa crus-galli), represent a smaller part of the Medieval archaeobotanical material in the Czech Republic (e.g. < 10% in Žatec; Kočár et al. 2010). The remains of common millet as a staple are an integral part of Medieval crop plant assemblages from Prague, while foxtail millet is less common (Opravil 1994; Čulíková 2010). The average δ13C values for C4 plants may reach − 12.5‰ and range from − 15 to − 9‰ (O'Leary 1988). The δ13C values in plants can also be affected by temperature, humidity and irradiance (Tieszen 1991; Heaton 1999).
Animals cannot synthesize all the amino acids they need to build proteins and so must obtain essential amino acids from external plant-based sources (Galili et al. 2016). While herbivores include them mainly by the transamination of keto acids derived from carbohydrates in their diet, omnivores include proteins in their diet and eventually involve carbohydrates in their protein synthesis. Thus, δ13C values of the collagen of omnivores may not reflect their total diet but only the animal protein portion there (Krueger and Sullivan 1984). The δ13C value increases by up to 2‰ with each trophic level (Bocherens and Drucker 2003) and the δ15N value by up to + 3–6‰ (DeNiro and Epstein 1981; O'Connell et al. 2012). The δ15N values in plants are closely linked to the amount of mineralizable N in soils and are influenced by environmental factors such as soil type, age and depth, moisture or climatic conditions (Ambrose 1991; Pardo and Nadelhoffer 2010; Szpak 2014). The nitrogen-15 abundance in large herbivore collagen increases with rising temperatures, which lead to higher soil activity (Drucker et al. 2003). The δ15N values of adult animal tissues can be affected by seasonal changes in dietary composition and its quality (Sponheimer et al. 2003). Some agroecological measures may also be reflected in the isotopic record, such as the fertilization of agricultural crops, which increases the δ15N values in plants (Bogaard et al. 2007; Fraser et al. 2011).
Materials and methods
Teeth extracted from the mandibular and maxillary bones of cattle (Bos taurus), domestic pigs (Sus domesticus), sheep (Ovis aries) and goats (Capra hircus) were individually selected from excavation contexts originally belonging to four Medieval assemblages from the historical centre of Prague (Table 1, Fig. 1): the Lesser Town, (1) Malostranské Square No. 2/III; Prague Castle, (2) Prague Castle, Northern Promontory; the Old Town, (3) Křižovnická Street No. 71/I; and (4) Pařížská Street No. 205/V. The Lesser Town is represented by the settlement on Malostranské Square No. 2/III, which in the Early Middle Ages (ninth to eleventh centuries) was within the southern fortified outer bailey of Prague Castle (Čiháková 2009, 2018). The site of Prague Castle, Northern Promontory (Frolík 1997; Hurajčíková 2014) formed part of the fortified western bailey of Prague Castle in the twelfth to thirteenth centuries. The area of the Old Town is represented in this study by an assemblage originating from Křižovnická St. No. 71/I. During the High Middle Ages (thirteenth to fourteenth centuries), this lay outside the fortified area of the Medieval town and close to the Old Town walls (Starec 2017a). The assemblage from Pařížská St. No. 205/V, dating to the thirteenth century (Starec 2017b), represents the area of the former Jewish Quarter in the Old Town. The dates are based on the chronology established through Medieval Prague ceramics sherds.
Teeth were selected for analysis based on age assignments, and only adult individuals were included in the study.
The estimation of age at death for all individuals listed in Table 2 was based on the stages of tooth eruption, replacement and wear. For cattle, dental wear stage ages for the incisors (I1–I2), premolars (P2–P4) and molars (M2–M3) were estimated according to the stages of tooth eruption and replacement provided by Higham (1967) and observation of the occlusal surface (Legge 1992; Komárek 1993). The age at death assessments for pigs are based on the recording of eruption and wear stages on the premolar (P4) and molars (M2–M3; Grant 1982; Horard-Herbin 1997). For the premolar (P4) and molars (M2–M3) of sheep and goats, the age estimation methods follow Helmer (1995), see Helmer and Vigne 2004) and Payne (1973). The teeth of sheep were distinguished from those of goats, and the teeth of domestic pigs from those of wild boar, on the basis of tooth morphology or dimensions (Helmer 2000; Zeder and Pilaar 2010; Evin et al. 2014).
The selection of animal teeth (N = 120; Tables 1 and 2) for the purpose of stable isotope analysis (each sample corresponding to one individual) was limited by the size of archaeological contexts. The Prague Castle assemblage did not contain enough teeth of cattle, sheep and goats; the material originating from the Lesser Town lacks the teeth of sheep and goats; and the Pařížská St. assemblage lacks the teeth of pigs (cf. Table 1). No traces of diagenetic or pathological changes were observed on the teeth, and the state of preservation of all finds is comparable. Stable carbon and nitrogen isotope values measured in the samples of the root dentine collagen retain the stable isotope composition from the period of the root formation (Hillson 2005). Root dentine can give insights into the animal diet over the time span of root growth, and it is not remodelled, unlike bone collagen (Drucker et al. 2001). When root formation is completed and the tooth is abraded, dentine formation continues slowly in the root canal (Hillson 2005). The continuously deposited secondary dentine may develop until the root canal is nearly filled in (Gustafson 1950; Brown et al. 1960). Most of the roots of younger individuals develop over a time span of 6–7 months (Fig. 4); they can form at different times in their lives, and the stable isotope values can reflect seasonal variation in birth and sources of food. On the other hand, the isotope measurements obtained for older individuals represent a longer time span because of the deposition of secondary dentine in the roots (Gustafson 1950; Hillson 2005). The modalities of dentine formation can have an influence on the increase of interindividual variability. The time span of the root formation of the incisors (I1–I2), premolars (P2–P4) and molars (M2–M3) of cattle (Brown et al. 1960), domestic pigs (Tonge and McCance 1973) and caprines (Hillson 2005) included in this study are summarized in Fig. 4. M1 teeth were not sampled due to potential 15N enrichment from breast milk.

Laboratory sample preparation for the stable isotope analysis
One root was extracted from each tooth, and its cementum layer was mechanically cleaned using a drill bit. The root was then cleaned in an ultrasonic bath, dried and crushed. The weight of the samples ranged from 100 to 300 mg depending on the size and preservation of the dental root. Tooth dentine collagen was extracted using the standard procedures of Longin (1971), modified by Bocherens (1992) at the Anthropological Department of the National Museum in Prague. Each sample was demineralized in 1 M HCl for 20 min to remove exchangeable carbonates. Subsequently, the samples were purified in 0.125 M NaOH for 20 h to remove soil humic substances, followed by a plunge of the residue into 0.01 M HCl for 17 h, to gelatinize the collagen. Subsequently, the samples were filtered and lyophilized in a freeze dryer. Stable carbon and nitrogen isotope compositions were determined using a Sercon 20-20 CF-IRMS coupled to an elemental analyser at the Iso-Analytical Ltd. laboratory in Crewe, UK.
Stable carbon and nitrogen isotope measurements were calibrated relative to the V-PDB and AIR scales using IA-R068 (soy protein). Measurement uncertainty was monitored using collagen standards with well-characterized isotopic compositions: IA-R038 (L-alanine), IA-R069 (tuna protein) and a mixture of IAEA-C7 (oxalic acid) and IA-R046 (ammonium sulphate). Following Szpak et al. (2017, Appendix F-G), repeated measurements of the calibration standard, check standards and sample replicates were used to determine a random error (precision) of ± 0.07‰ for δ13C and ± 0.06‰ for δ15N. Systematic measurement error (accuracy) was determined to be ± 0.11‰ for δ13C and ± 0.16‰ for δ15N, and the total analytical uncertainty was estimated to be ± 0.13‰ for δ13C and ± 0.17‰ for δ15N. Additional details (calibration and check standards and sample replicates) are provided in Supplementary Material 1.
Statistical differences between groups of data were tested in the Statistica 12 program using t test when the condition of normality data was met (Lilliefors test), the Mann-Whitney test when the condition of normality data was not met, histograms and interpolation of distribution functions. Significance was tested at α = 0.05 level and also at α = 0.017 level when the Bonferroni correction was used. The aim of this approach is to reduce the chances of false-positive results when multiple pairwise tests are performed on a single set of data.
Results
The δ13C and δ15N results are summarized in Tables 2 and 3. The carbon content in the animal dentine collagen was between 33.0 and 47.2%, and the nitrogen content was between 12.0 and 17.6%. The C/N ratios of the faunal samples fit into the acceptable range of 3.0 and 3.6, and all samples are deemed of acceptable quality (Ambrose 1990; van Klinken 1999). The results contain no extreme values, and only δ13C values in samples SV19 (domestic pig; − 19.0‰) and PAR3 (cattle; − 18.6‰) and δ15N in sample KR20 (domestic pig; + 11.2‰) may be considered outliers (all three samples are marked with an * in Table 2). All samples were processed in large batches and following a same protocol. The deviation of samples is not due to a processing error but may reflect particular life stories.
The δ13C values of cattle from Medieval Prague range from − 21.8 to − 18.5‰ (Lesser Town: − 20.7 ± 0.5‰, Křižovnická St.: − 20.9 ± 0.6‰, Pařížská St.: − 20.1 ± 0.8‰), and δ15N values vary from + 4.4 to + 9.3‰ (Lesser Town: + 6.7 ± 1.0‰, Křižovnická St.: + 7.4 ± 1.1‰, Pařížská St.: + 7.6 ± 1.6‰). The δ13C values of pigs range from − 22.4 to − 19.0‰ (Lesser Town: − 21.1 ± 0.6‰, Prague Castle: − 21.0 ± 0.7‰, Křižovnická St.: − 20.9 ± 0.6‰) and δ15N values from + 4.5 to + 11.2‰ (Lesser Town: + 7.0 ± 1.1‰, Prague Castle: + 7.8 ± 1.4‰, Křižovnická St.: + 8.0 ± 1.3‰). The stable isotope values for sheep and goats were grouped due to the small sample size. Their δ13C values vary from − 22.2 to − 20.4‰ (Křižovnická St.: − 21.2 ± 0.4‰, Pařížská St.: − 20.7 ± 0.2‰) and δ15N values between + 5.9 and + 9.8‰ (Křižovnická St.: + 7.9 ± 1.2‰, Pařížská St.: + 8.8 ± 0.6‰).
Statistical comparison of the two High Medieval sites in the Old Town (Pařížská St. and Křižovnická St.), taking into account Bonferroni correction, shows a significant difference in neither the δ13C values of cattle (t-value = 2.238, df = 17, p = 0.039) nor the δ15N values (t-value = 0.257, df = 17, p = 0.800). By contrast, the results for δ13C obtained for sheep and goats were close to the level of significance (δ13C: t-value = 2.749, df = 13, p = 0.017; δ15N: t-value = 1.580, p = 0.138). There was no statistically significant difference between the mean δ13C and δ15N values of cattle and caprines from the Old Town for each site separately (Pařížská St.: δ13C: t-value = 1.361, df = 10, p = 0.203, δ15N: t-value = − 1.566, p = 0.149; Křižovnická St.: δ13C: t-value = 1.639, df = 20, p = 0.117, δ15N: t-value = − 0.956, p = 0.350).
Cattle and pig dentine collagen from the earlier period is characterized by consistently low δ15N values and higher δ15N values in the later period (Fig. 5). The results of the stable isotope measurements of all farm animals clearly show that their diet was 15N enriched in the High Middle Ages (Fig. 5), and the values are statistically conclusive (t-value = − 4.292, df = 118, p < 0.0001). There was no similar shift for δ13C values (t-value = − 0.134, df = 118, p = 0.893). In addition to the increase in δ15N in the tooth dentine collagen of cattle and domestic pigs, its variability was also increasing.

Mean δ13C and δ15N values measured in the tooth dentine collagen of animals for the different areas of Prague. Error bars represent ± 1σ.
Cattle
The measured samples of cattle come from the Early Medieval Lesser Town and the High Medieval Old Town (Křižovnická and Pařížská St.; Table 1). The δ13C values of pairs of sites–Lesser Town and Křižovnická St. (t-value = − 0.967, df = 30, δ13C: p = 0.341) and Lesser Town and Pařížská St. (t-value = 2.177, df = 25, δ13C: p = 0.039)–were compared. The difference in the δ13C mean values among sites taking into account Bonferroni correction is not statistically significant. The δ15N mean values of cattle from the Lesser and Old Towns increased over time from + 6.7 to + 7.5‰, but this cannot be conclusively corroborated by the data including Bonferroni correction (t-value = 2.165, df = 37, p = 0.037). The coefficient of variation of δ15N values for the Early Medieval site is lower (15.0) than for the High Medieval Old Town (17.2).
Comparison of the δ13C and δ15N values in cattle from the Lesser Town (N = 20) slaughtered at the ages of up to and above 6.5 years (Table 4) indicates no statistically significant differences between the age groups (δ13C: t-value = − 0.671, df = 18, p = 0.510; δ15N: t-values = − 0.761, df = 18, p = 0.456). Analysis of samples of the same age groups of cattle from Křižovnická St. (N = 12; Table 4) yielded similar results (δ13C: t-value = 0.228, df = 10, p = 0.824; δ15 N: t-value = − 0.896, df = 10, p = 0.391). The stable isotope values of cattle from Pařížská St. were not statistically tested because of the small sample size.
The distributions of δ13C values of cattle in the Early and High Middle Ages are close to the normal (Gaussian) distributions (Fig. 6a). In contrast to the Early Middle Ages, the distribution curve of the δ15N values for samples from the High Middle Ages shifts towards higher values, and the platykurtic shape of the distribution curve in its higher value section also seems to corroborate to varying degrees a relative 15N enrichment of the diet (Fig. 6b).

Distribution of stable isotope values of cattle (Bos taurus) from Prague over time. δ13C values (a) and δ15N values (b)
Domestic pigs
Samples of domestic pig were obtained from the Lesser and the Old Towns (only from Křižovnická St.) and from Prague Castle (Table 2). The coefficient of variation of δ13C for the Old Town assemblage (2.7) is lower than that of the Lesser Town (3.0) and Prague Castle (3.4; Table 3) assemblages. As the δ13C values do not show a normal distribution (Lilliefors test, p < 0.01), the Mann-Whitney test was used to compare them. The results obtained indicate that δ13C values do not differ in any of the pairs of sites–the Lesser Town and Prague Castle (Z = − 0.257, p = 0.797), the Lesser Town and the Old Town (Z = − 1.032, p = 0.302) or the Old Town and Prague Castle (Z = 0.783, p = 0.433). The coefficient of variation of δ15N reaches the lowest values in the earliest set from the Lesser Town (16.3) and increases in the later periods from the Old Town (16.8) and Prague Castle (17.3; Table 3). In the case of δ15N, no difference from a normal distribution can be attested. The differences resulting from the comparison of δ15N in the tooth dentine collagen of pigs in the assemblages from Prague Castle and the Lesser Town, taking into account Bonferroni correction, are not statistically significant (t-value = 2.163, df = 41, p = 0.036). The same is true of the mean δ15N values for Prague Castle and the Old Town (t-value = − 0.273, df = 40, p = 0.786). The mean δ15N values of pigs significantly differs between Lesser Town and the Old Town (t-value = − 2.584, df = 45, p = 0.013).
The analysed data regarding domestic pigs from the Lesser Town (N = 24) and the Old Town (N = 23) can be divided into two age groups based on the slaughter age: up to 2 years and over 2 years (Table 4). The sample ratios of younger and older pigs are 2:1 (Lesser Town) and 1.9:1 (Old Town). Comparison of the δ13C and δ15N mean values (Mann-Whitney test) in both age groups has not shown statistically significant differences between the Early Medieval Lesser Town (δ13C: Z = 0.827, p = 0.408, δ15N: Z = − 0.214, p = 0.830) and the High Medieval Old Town (δ13C: Z = 0.355, p = 0.723, δ15N: Z = 0.097, p = 0.923). The data for domestic pigs from Prague Castle are missing because of the significant imbalance in samples from both age groups (younger, N = 14; older, N = 4; Table 2).
The δ13C distribution curve for the Lesser Town shows a bimodal distribution, while the curve for the High Medieval Old Town is unimodal and symmetrical (Fig. 7a). The distribution curve of δ15N values for the High Middle Ages covers a broader range of values compared to the Early Medieval Lesser Town (Fig. 7b), which corresponds to the comparison of coefficients of variation (Table 3).

Distribution of stable isotope values of domestic pig (Sus domesticus) from Prague over time. δ13C values (a) and δ15N values (b)
Sheep and goats
The δ13C and δ15N values from caprines which were kept for more than 2 years (Table 2) were collected only for High Medieval assemblages from the Old Town–Pařížská St. and Křižovnická St. (Table 1). The sample ratio of sheep and goats was 1:2 (Pařížská St.) and 1:1 (Křižovnická St.). Because of the limited number of specimens identified as either sheep or goat (Table 2), the data for these ruminants were grouped. The coefficient of variation of δ13C values is higher in caprines from Křižovnická St. (2.0) than from Pařížská St. (1.1; Table 3). The coefficient of variation of δ15N values is also more pronounced in Křižovnická St. (15.3) than in Pařížská St. (7.0; Table 3). The δ15N values of the tooth dentine collagen of caprines do not show statistically significant differences depending on the area of the Old Town, unlike δ13C values (see above).
Discussion
The δ13C values of cattle from the Lesser Town (ninth to eleventh centuries) point to grazing in an open habitat and not in a closed canopy environment (Drucker et al. 2008; Kohn 2010). The obtained δ13C values (Table 3) are comparable to the δ13C values measured in Neolithic cattle at Černý Vůl (Prague-West district) preferentially grazing on patches of steppe grasslands and open-canopy forests or forest boundaries (− 20.6 + 0.4‰; Berthon et al. 2018). As evidenced by pollen and plant macro-remains analyses, the Lesser Town’s surroundings likely provided enough grazing opportunities for livestock (Kozáková et al. 2009; Čulíková 2010). The archaeozoological results do not support self-sufficiency in cattle breeding, but it is presumed that some animals were kept on site (more than half of the cattle in the Lesser Town were older than 6 years of age; Fig. 3b) to provide secondary products, such as traction or milk, and these animals needed grazing space.
The δ13C values of domestic pigs (Table 3) indicate primary consumption of C3 plants (Tieszen 1991; Millard et al. 2013) in an open grazing environment (Drucker et al. 2008; Kohn 2010). Results of isotopic measurements for the Lesser Town indicate that the dietary behaviour of domestic pigs was similar to that of the Early Medieval wild boars and domestic pigs from the Great Moravian centre at Mikulčice, situated in the valley floodplain of the Morava River (eighth to ninth centuries; Kovačiková et al. 2020), as well as domestic pigs from Prague Castle (2nd courtyard) and the Levý Hradec stronghold, dating to the ninth to eleventh centuries (Kaupová et al. 2019). For further details, see Table 5. The diet of extensively herded pigs, like that of wild boar, comprises primarily plant matter (80–90% of total food mass) and small vertebrates and invertebrates (Holá et al. 2015). A suitable precondition for extensively managed pigs is the existence of forest areas rich in acorns, roots and other food components (Albarella et al. 2007). The pollen spectra from the Lesser Town show a relatively low abundance of oaks in the tenth century (Kozáková and Pokorný 2007). By contrast, archaeobotanical data from Nebovidy, which is close to the Lesser Town (eleventh to twelfth/thirteenth centuries), gives evidence of a higher diversity of habitats, including forest vegetation with dominant oak, subdominant pine and other broadleaved species (Kočár and Kočárová 2013). Mixed oak forests can provide good foraging for pigs, but more open stands are generally better, as they produce larger quantities of seeds every year (Leuschner and Ellenberg 2017). Distortion of the distribution curve (the local extreme) plotted for the Lesser Town pigs (Fig. 7a) in the section of lower δ13C values could indicate that these pigs consumed plants from more shaded or wetter habitats (Tieszen 1991; Hamilton and Thomas 2012). In the ninth century, the western boundary of the fortified Lesser Town was formed by the old river channel of the Vltava River (Čiháková 1999), running past the marshes and floodplain vegetation under the slope of Prague Castle (Čulíková 2010). Plant taxa that benefit from flood sediments and higher nutrient concentrations have been identified in the pollen spectra from Valdštejnská St. in the Lesser Town (Kozáková 2008).
A small number of pigs and cattle from the Lesser Town with higher δ15N values can be considered to have been fattened by different methods, based on the partial intake of animal proteins (pigs, range 4.3‰; Frémondeau et al. 2017) or 15N-enriched plants (cattle, range 4.2‰; Commiso and Nelson 2006, 2007; Bogaard et al. 2007). A greater variability in δ15N values may provide evidence of grazing on vegetation in different microclimates or microhabitats (Stevens et al. 2006; Madgwick et al. 2012) or of grazing close to settlements where human and animals waste had accumulated (Alexander et al. 2019). Swine herding associated with woodland pasture, grazing on grass, roots and crop stubble or consuming such leftovers as household food waste (Albarella 2006; Jørgensen 2013) with variable animal control (Price 2016) was the traditional extensive method typical of less urbanized areas (Price 2016; Leuschner and Ellenberg 2017).
The results of the stable isotope analysis and the archaeozoology suggest that the inhabitants of the Lesser Town used several breeding strategies within the same area or that this particular part of the town was partially supplied by animals from another environment. Different strategies seem to be corroborated by the bimodal distribution of the δ13C curve plotted for pigs (Fig. 7ab). The household-based agricultural system in the Early Middle Ages, where household was a basic unit of residence, production and consumption (Halstead 1989), still followed the farming practices used earlier (Kuneš and Abraham 2017). The household herd of livestock produced milk, meat and wool, supplied manure for the fields and sometimes provided a saleable surplus above domestic requirements (Halstead 1996). Such herd management is well documented throughout Europe in written sources from as early as the beginning of the Middle Ages (Petráňová and Petráň 2000). In the Early Middle Ages, the Lesser Town formed part of the fortified Přemyslid’s seat of power and was located on the southern outskirts of the area. This was described by the Arabic intellectual of Jewish origin, Ibrahim Ibn-Yakub, who visited Prague in the tenth century, as a town and a centre of long-distance trade (Čiháková 2018). The archaeozoological data and the results of stable isotope analyses do not exclude the possibility that from the ninth to eleventh centuries, household farming could have been combined with a specific dependence on the hinterland’s production and market.
The δ13C values of domestic pigs from Prague Castle (twelfth/thirteenth centuries) indicate consumption of C3 plants (Tieszen 1991) and grazing in predominantly open areas (Drucker et al. 2008; Kohn 2010). This conclusion is in line with the pollen diagrams from Prague Castle during the Early Middle Ages (ninth to tenth centuries), which show low numbers of pollen grains from tree species such as oak and shrub communities and high ratios of pollen from xerophytes, indicating a pasture that probably grew on the southern slope below Prague Castle (Kozáková 2008; Kozáková and Boháčová 2008). Although the mean δ13C values of pigs from Prague Castle revealed no statistically significant differences from the Lesser Town, their range is wider (Table 3). A similar inference can be made from the δ15N values. The animals likely came from comparable pasture habitats, as seen on the δ13C value distribution curves for both sites (Fig. 7a); the δ15N value distribution curves, however, changed over time (Fig. 7b).
The pigs from Prague Castle show both an increasing variability in their δ15N values and a higher δ15N mean value when compared to the pigs from the Lesser Town (range of δ15N values of 5‰ and 4.3‰, respectively; Table 3). The proportion of pigs with a 15N-enriched diet (δ15N > 8‰), whether indicative of a higher intake of animal protein or 15N-enriched plant fodder, is higher at Prague Castle, totalling nine specimens (43% of the assemblage).
The increasing range of stable isotope values (both δ13C and δ15N; Table 3) can be interpreted not only as a reflection of landscape changes (e.g. declining woodland coverage, eutrophication) or another foddering regime but also as a manifestation of the social transformation concerning the High Medieval castle. Prague Castle served as an administrative and economic centre, the seat of the monarchs, and fulfilled representative functions. Moreover, it was gradually rebuilt into a Romanesque and later Early Medieval castle during the twelfth century (Frolík and Smetánka 1997). Although the results of analyses of the archaeozoological data from the later period corroborate the hypothesis of partial self-sufficiency in pig production, it can be assumed that the castle was supplied from more varied sources, either through the market or from animals supplied to the royal court, as indicated by, for example, the predominance of male pigs in the assemblage. Compared to the Early Medieval Lesser Town, the higher isotopic variation of samples from Prague Castle in terms of nitrogen (on average 0.8‰; Table 3) might also be interpreted as a shift from local pork to pork from elsewhere (Millard et al. 2013).
Due to human impact and land use, the high level of diversity of the Early Medieval landscape around Prague decreased after the end of twelfth century (Kozáková et al. 2009; Pokorná et al. 2014). Increasing ruderalisation and the intensification of human impact were manifested by indicators of trampled vegetation and nitrophilous plant species (Pokorná et al. 2014).
Climate data support the prevalence of warm conditions across the greater part of Europe at the end of the twelfth and the beginning of the thirteenth centuries (Brázdil et al. 2005; Poschlod 2015). Such favourable climate significantly affected the availability of biomass, supporting population growth and the expansion of human settlement into former pastures (Brázdil et al. 2005). The development and economy of newly founded Medieval towns depended on their connection with their agriculture environs and the production of the countryside (Klápště 2012). Therefore, a presence of non-local animals in Medieval towns is likely.
Although the δ13C values in samples of the tooth dentine collagen of the majority of cattle from Křižovnická and Pařížská St. in the Old Town (thirteenth to fourteenth centuries) indicate the consumption of C3 plants (Tieszen 1991) and open grazing habitats (Drucker et al. 2008; Kohn 2010), the dataset from Pařížská St. (Table 3) is significantly influenced by one sample (PAR3). As bone collagen stable isotope composition primarily reflects the protein component of diet (Jim et al. 2004), this sample, which has higher δ13C values (− 18.6‰), may point to individuals with a different dietary history. The presence of C4 plants such as common millet or foxtail millet in the cattle diet may explain such an occurrence (Fuller et al. 2012). Both C4 grains were present in the archaeobotanical evidence from Medieval Prague, although always in relatively minor quantities when compared to other cereals (Opravil 1994; Čulíková 2010). Additionally, the influence of environmental factors, such as increasing light intensity, declining water availability and higher nutrient content in the natural areas (Tieszen 1991), suggests that a different origin for this individual cannot be excluded.
The urbanization process in the thirteenth century was related to the development of numerous crafts, the butchery profession included. The number of butchers increased in Prague in the second half of the thirteenth century (Tomas 1983). There were 241 butcher stalls in Prague before the beginning of the fifteenth century (an estimated one butcher stall per 187 inhabitants; Hoffmann 2010). The proportion of animal bones with recorded cut marks found in Prague increases from the thirteenth century (Sůvová et al. 2018), probably as a result of changes in cutting techniques and the distribution of carcass portions suitable for cuisine. The butchers bought livestock in the countryside near Prague or in the livestock market directly in the town. Moreover, inhabitants of the town also acquired their own animals (Pokorná et al. 2019). The archaeobotanical data supports the use of meadow plants like hay, straw as litter for animals and manure in household stables and provides evidence of the agricultural character of the Old Town (Havrda et al. 2017).
Written texts from the fifteenth century reveal that urban livestock grazed on defined lands outside the walls and explicitly mention stubble, meadows and forest (CIM IV/2, n. 389; Šedivý 2015). As suggested by isotopic analyses, cattle was put out to municipal pastures near the town, and hay or kernel fodder could be obtained at the municipal market (Pokorná et al. 2019). The latter types of feed, however, are not directly reflected in the isotope record unless the fodder contained a mixture of plants with C3 and C4 photosynthesis cycles (Fuller et al. 2012). The archaeozoological data indicate that nearly 60% of the cattle were slaughtered before they reached 3 years of age. Isotopic data of individuals slaughtered at younger and older ages show no significant differences, and it can therefore be concluded that their future use (for meat or dairy production or as draught animals) did not influence the fattening method during the period when their teeth roots were formed.
With the development of the beef market, the territory from which cattle were delivered to Prague expanded. The higher variability and distribution of the cattle δ15N values (range 4.8‰; Table 3, Fig. 6b) may reflect the varying pastures grazed by these animals. Although cattle from the Old Town stayed in a similar pasture habitat as cattle from the Lesser Town, the mean of nitrogen isotopic values of the Old Town cattle is slightly raised by 0.7–0.9‰ (Table 3). This increase can be considered a consequence of the intake of fodder containing 15N-enriched components or of higher nitrogen supply to the biosphere, e.g. by fertilization (Bogaard et al. 2007; Fraser et al. 2011). Stable nitrogen isotope analysis of soil samples from the Medieval village of Roudnička (40 km south-west of Prague), for example, suggests the application of organic fertilizers in Bohemia in the thirteenth century (Součková et al. 2013). Moreover, archaeological research into a loose distribution of ceramic sherds has indicated the beginnings of widespread fertilization during the High Middle Ages (Klápště 2012).
Establishing the precise dietary preferences of caprines is difficult because of the distinction between species. According to the slaughter age data, there was a higher proportion of adult sheep and goats (above 4 years of age) of breeding age at the Old Town sites (Křižovnická and Pařížská St.; Fig. 3c). Even though their δ13C values (on average − 21.0‰) corroborate consumption of C3 plants and the open grazing environments (Tieszen 1991; Drucker et al. 2008; Kohn 2010), they do differ. The δ13C values from Pařížská St. are higher, as are those of cattle from this site. It could be that the inhabitants of this part of town acquired livestock with a different dietary history or from different environments. The Jewish society from the Old Town was supplied by its own butchers, who offered the meat exclusively inside this community (Valášková 2014). The isotopic data could indicate that these butchers obtained the animals by different supply chain. Compared to cattle, the mean δ15N values of caprines shifted by 0.5–1.2‰ (although not statistically significant; Table 3), which could indicate that these ungulates, unlike cattle, were raised on plots adjacent to houses, which were more likely to be fertilized with animal dung than more distant fields that were fertilized less often (Jones et al. 1999), or that they stayed in a relatively small space where animal dung was concentrated (Makarewicz 2014).
The relative proportion of determined pig bone remains decreased in the High Medieval Old Town (Fig. 2). Most of the animals were slaughtered before 2 years of age (Fig. 3a), and the carbon and nitrogen isotopic composition of their samples does not suggest a foddering strategy that differed from that for the less numerous pigs slaughtered at a later age (> 2 years). The absence of pig finds in Pařížská St. could be explained by the fact that this part of the Old Town was inhabited by a Jewish community, which forbade the consumption of pork (Valenzuela-Lamas et al. 2014). The mean δ13C values of pig samples from Křižovnická St. and the other sites did not show statistical differences; the course of the δ13C distribution curve (Fig. 7a), however, indicates a higher uniformity of the plant component in the diet of pigs at the Old Town. Moreover, there is a broader range of δ15N values (Table 3) and an increase in the proportion of individuals with δ15N values above 8‰ (52%); five of these even exceed 9‰, which is probably related to more significant variability of feed intake. The continuously increasing range of δ15N values can be explained as a result of differing pig husbandry strategies (Buonincontri et al. 2017). Such a trend can be explained by pigs being more frequently fed by household, agricultural and street leftovers containing more δ15N. In this context, the written sources attest the presence of pigs in towns and state that pigs were allowed to move freely but not to damage the property of other citizens (CIM IV/2 – Codex iuris municipalis Regni Bohemiae IV/2 (1453-1500) 1960, n. 38). Although backyard pigs in the urban space are well known from historical sources (Šedivý 2015), it must be emphasized that non-local animals can be presumed to have been present in High Medieval towns (Alexander et al. 2019). These pigs could have been kept in villages or courts under various husbandry regimes.
Conclusion
The stable isotope data (δ13C and δ15N values) obtained from three areas within Medieval Prague show that different animal husbandry strategies were applied and point to a more complex acquisition of resources by the inhabitants of the High Medieval town. Compared to the previous period (ninth to eleventh centuries), the range of δ13C and δ15N values in dental collagen samples of the livestock from the thirteenth and fourteenth centuries may indicate that the farm-household model was used by the inhabitants of Prague but that they also obtained meat more often from elsewhere, e.g. through the market.
Agreement between the archaeozoological and isotope data seems to corroborate increased diversity in pig feeding, although mainly under extensive conditions, in the Early Medieval site of the Lesser Town (ninth to eleventh centuries), where pig bones predominated in the local osteological assemblage. The diet of pigs consists mainly of plants from open habitats, and more than half of them were slaughtered before the age of 18 months. The grazing environment, which had to be sufficiently nutritious due to the shorter pig fattening period, must have included several types of habitats, including areas close to human settlements. The majority of pigs from later assemblages, which were obtained from Prague Castle (twelfth to thirteenth centuries) and the Old Town (thirteenth to fourteenth centuries), were slaughtered at 18–24 months; their population was characterized by increasing variability in nitrogen isotope values and a higher intake of animal protein-rich diet. Long-term pig breeding, probably influenced by a demand for more meat and fat, led to a broader range of fattening methods.
The slaughter age of Early Medieval cattle at the Lesser Town seems to corroborate their use for both meat and milk; any long-term sustainability of farming such animals would have relied on sufficient grazing opportunities nearby. In contrast, the development of the High Medieval Old Town significantly increased the demand for beef from younger animals, and it is evident that the cattle samples originating from this part of Prague are more heterogeneous in δ13C values than the assemblage from the Lesser Town, which may imply a mixing of animals coming from several types of environment.
In the High Middle Ages, most of the sheep and goats clearly lived for more than 4 years, as the inhabitants preferred to use them in various ways rather than simply consume them.
The values of δ13C in sheep and goat samples differ in two sites within the High Medieval Old Town (the samples from Pařížská St. are significantly enriched in 13C compared to Křižovnická St.). Likewise, the higher mean δ13C values in cattle from Pařížská St. support the dissimilarity between settlements. This can be explained by the fact that different factors, such as social milieu or ethnic background, could have affected the feeding regime of the livestock in a single area of Prague. At the same time, it is clear that expanding the existing database of isotope measurements by representative sets from other sites in Prague would allow a better assessment and understanding of these influences in the future.
References
Albarella U (2006) Pig husbandry and pork consumption in Medieval England. In: Woolgar CM, Serjeantson D, Waldron T (eds) Food in Medieval England: Diet and Nutrition. Oxford University Press, Oxford, pp 72–87
Albarella U, Manconi F, Vigne JD, Rowley-Conwy P (2007) Ethnoarchaeology of pig husbandry in Sardinia and Corsica. In: Albarella U, Dobney K, Ervynck A, Rowley-Conwy P (eds) Pigs and Humans: 10,000 Years of Interaction. Oxford University Press, Oxford, pp 285–307
Alexander MM, Gutiérrez A, Millard AR, Richards MP, Gerrard CM (2019) Economic and socio-cultural consequences of changing political rule on human and faunal diets in medieval Valencia (c. fifth–fifteenth century AD) as evidenced by stable isotopes. Archaeol Anthropol Sci 11:3875–3893. https://doi.org/10.1007/s12520-019-00810-x
Ambrose SH (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. J Archaeol Sci 17:431–451. https://doi.org/10.1016/0305-4403(90)90007-R
Ambrose SH (1991) Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs. J Archaeol Sci 18(3):293–317. https://doi.org/10.1016/0305-4403(91)90067-Y
Berthon R, Kovačiková L, Tresset A, Balasse M (2018) Integration of Linearbandkeramik cattle husbandry in the forested landscape of the mid-Holocene climate optimum: seasonal-scale investigations in Bohemia. Archaeol Anthropol Sci 51:16–27. https://doi.org/10.1016/j.jaa.2018.05.002
Bocherens H (1992) Biogéochimie isotopique (13C, 15N, 18O) et paléontologie des vertébrés: applications à l'étude des réseaux trophiques révolus et des paléoenvironnements. PhD. thesis. Université Paris
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13(1-2):46–53. https://doi.org/10.1002/oa.662
Bogaard A, Heaton THE, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J Archaeol Sci 34(3):335–343. https://doi.org/10.1016/j.jas.2006.04.009
Boháčová I, Herichová I (2009) Raně středověký sídelní areál v západní části hradčanského ostrohu. Archeologica Pragensia 19:257–308
Brázdil R, Pfister C, Wanner H, Von Storch H, Luterbacher J (2005) Historical climatology in Europe–the state of the art. Clim Chang 70(3):363–430
Brown WAB, Christofferson PV, Massler M, Weiss MB (1960) Postnatal tooth development in cattle. Am J Vet Res 21:7–34
Buonincontri MP, Pecci A, Pasquale GD, Ricci P, Lubritto C (2017) Multiproxy approach to the study of medieval food habits in Tuscany (central Italy). Archaeol Anthropol Sci 9:653–671. https://doi.org/10.1007/s12520-016-0428-7
Čiháková J (1999) Malá Strana od pravěku do vrcholného středověku. In: Vlček P (ed) Umělecké památky Prahy. Malá Strana, Academia, Praha, pp 11–27
Čiháková J (2009) Archeologický výzkum v rotundě sv. Václava na Malostranském náměstí. Časopis Společnosti Přátel Starožitností 117:14–30
Čiháková J (2018) The Lesser Town of Prague in the Tenth and Eleventh Centuries. In: Kouřil P, Procházka R (eds) Moravian and Silesian strongholds of the tenth and eleventh centuries in the context of Central Europe, Spisy Archeologického ústavu AV ČR Brno 57. Archeologický ústav AV ČR, Brno, pp 283–303
CIM IV/2 – Codex iuris municipalis Regni Bohemiae IV/2 (1453-1500) (1960) In: Haas A (ed) Nakladatelství československé akademie věd, Praha
Commiso RG, Nelson DE (2006) Modern plant δ15N values reflect ancient human activity. J Archaeol Sci 33(8):1167–1176. https://doi.org/10.1016/j.jas.2005.12.005
Commiso RG, Nelson DE (2007) Patterns of plant δ15N values on a Greenland Norse farm. J Archaeol Sci 34(3):440–450. https://doi.org/10.1016/j.jas.2006.06.012
Čulíková V (2010) Medieval floodplain of the river Vltava in the neighbourhood of the Lesser Town in Prague (Valdštejnská St. No. 154/III, Kolovratský Palace). Archeologické Rozhledy 62:72–116
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta 42(5):495–506. https://doi.org/10.1016/0016-7037(78)90199-0
Drucker D, Bocherens H, Pike-Tay A, Mariotti A (2001) Isotopic tracking of seasonal dietary change in dentine collagen: preliminary data from modern caribou. C R Acad Sci Ser IIA Earth Planet Sci 333(5):303–309. https://doi.org/10.1016/S1251-8050(01)01640-8
Drucker DG, Bocherens H, Billiou D (2003) Evidence for shifting environmental conditions in Southwestern France from 33 000 to 15 000 years ago derived from carbon-13 and nitrogen-15 natural abundances in collagen of large herbivores. Earth Planet Sc Lett 216(1-2):163–173. https://doi.org/10.1016/S0012-821X(03)00514-4
Drucker DG, Bridault A, Hobson KA, Szuma E, Bocherens H (2008) Can carbon-13 in large herbivores reflect the canopy effect in temperate and boreal ecosystems? Evidence from modern and ancient ungulates. Palaeogeogr Palaeoclimatol Palaeoecol 266(1-2):69–82. https://doi.org/10.1016/j.palaeo.2008.03.020
Evin A, Cucchi T, Escarguel G, Owen J, Larson G, Vidarsdottir US, Dobney K (2014) Using traditional biometrical data to distinguish West Palearctic wild boar and domestic pigs in the archaeological record: new methods and standards. J Archaeol Sci 43:1–8. https://doi.org/10.1016/j.jas.2013.11.033
Fisher A, Thomas R (2012) Isotopic and zooarchaeological investigation of later medieval and post-medieval cattle husbandry at Dudley Castle, West Midlands. Environ Archaeol 17(2):151–167. https://doi.org/10.1179/1461410312Z.00000000013
Fraser RA, Bogaard A, Heaton T, Charles M, Jones G, Christensen BT, Halstead P, Merbach I, Poulton PR, Sparkes D, Styring AK (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38(10):2790–2804. https://doi.org/10.1016/j.jas.2011.06.024
Frémondeau D, De Cupere B, Evin A, Van Neer W (2017) Diversity in pig husbandry from the Classical-Hellenistic to the Byzantine periods: an integrated dental analysis of Düzen Tepe and Sagalassos assemblages (Turkey). J Archaeol Sci Rep 11:38–52. https://doi.org/10.1016/j.jasrep.2016.11.030
Frolík J (1997) Archeologický výzkum v tzv. Severním výběžku na Pražském hradě. Archaeol Pragensia 13:75–92
Frolík J, Smetánka J (1997) Archeologie na Pražském hradě. Paseka, Praha a Litomyšl
Fuller BT, De Cupere B, Marinova E, Van Neer W, Waelkens M, Richards MP (2012) Isotopic reconstruction of human diet and animal husbandry practices during the Classical-Hellenistic, imperial, and Byzantine periods at Sagalassos, Turkey. Am J Phys Anthropol 149(2):157–171. https://doi.org/10.1002/ajpa.22100
Galili G, Amir R, Fernie AR (2016) The regulation of essential amino acid synthesis and accumulation in plants. Annu Rev Plant Biol 67:153–178. https://doi.org/10.1146/annurev-arplant-043015-112213
Grant A (1982) The use of tooth wear as a guide of the age of domestic ungulates. In: Wilson B, Grigson C, Payne S (eds) Ageing and sexing animal bones from archaeological sites, British Archaeological Reports 109. B.A.R. Publishing, Oxford, pp 91–108
Gustafson G (1950) Age determinations on teeth. J Am Dent Assoc 41:45–54. https://doi.org/10.14219/jada.archive.1950.0132
Halstead P (1989) The economy has a normal surplus. In: Halstead P, O'Shea J (eds) Bad Year Economies: Cultural Responses to Risk and Uncertainty. Cambridge University Press, Cambridge, pp 68–80
Halstead P (1996) Pastoralism or household herding? Problems of scale and specialization in early Greek animal husbandry. World Archaeol 28(1):20–42. https://doi.org/10.1080/00438243.1996.9980329
Hamilton J, Thomas R (2012) Pannage, pulses and pigs: isotopic and zooarchaeological evidence for changing pig management practices in later Medieval England. Mediev Archaeol 56(1):234–259. https://doi.org/10.1179/0076609712Z.0000000008
Hammond C, O’Connor T (2013) Pig diet in medieval York: carbon and nitrogen stable isotopes. Archaeol Anthropol Sci 5:123–127. https://doi.org/10.1007/s12520-013-0123-x
Havrda J, Kočár P, Kočárová R, Kozáková R, Podliska J, Sůvová Z (2017) K vývoji historického nadloží Staroměstského náměstí v Praze. Příspěvek k mezioborovému výzkumu veřejných prostranství. Forum Urbes Medii aevii 10(1-2):160–191
Heaton TH (1999) Spatial, species, and temporal variations in the 13C/12C ratios of C3 plants: implications for Palaeodiet studies. J Archaeol Sci 26(6):637–649. https://doi.org/10.1006/jasc.1998.0381
Helmer D (1995) Biometria i arqueozoologia a partir d’alguns exemples del Proxim Orient. Cota Zero 11:51–60
Helmer D (2000) Discrimination des genres Ovis et Capra à l’aide des prémolaires inférieures 3 et 4 et interprétation des âges d’abattage: l’exemple de Dikili Tash (Greece). Anthropozoologica 31:29–38
Helmer D, Vigne JD (2004) La gestion des cheptels de caprinés au Néolithique dans le midi de la France. In: Bodu P, Konstantin C (eds) Approches fonctionnelles en Préhistoire, Actes du XXVe Congrès Préhistorique de France Nanterre, 24–26 Novembre 2000. Société Préhistorique Française Édition, Paris, pp 397–407
Higham CF (1967) Stock rearing as a cultural factor in prehistoric Europe. Proc Prehist Soc 33:84–106. https://doi.org/10.1017/S0079497X00014067
Hillson S (2005) Teeth. Cambridge University Press, Cambridge
Hoffmann F (2010) Řemeslník a kupec. In: Nodl M, Šmahel F (eds) Člověk českého středověku. Argo, Praha, pp 364–391
Holá M, Ježek M, Kušta T, Košatová M (2015) Trophic discrimination factors of stable carbon and nitrogen isotopes in hair of corn fed wild boar. PLoS One 10(4):e0125042. https://doi.org/10.1371/journal.pone.0125042
Horard-Herbin MP (1997) Le village celtique des Arènes à Levroux. L’élevage et les productions animales dans l’économie de la fin du second âge du Fer – Levroux 4. Fédération pour l'édition de la Revue archéologique du Centre de la France, Tours
Hrdlička L (2000) Centrum raně středověké Prahy. In: Piekalski J, Wachowski K (eds) Średniowieczny Śląsk i Czechy. Centrum średniowiecznego miasta. Wrocław a Europa środkowa. Uniwersytet Wrocławski, Wrocław, pp 191–214
Hurajčíková V (2014) Příspěvek k počátkům Pražského hradu (doklady a proměny osídlení západního předhradí na místě dnešního tzv. Severního výběžku). Master's thesis, Charles University
Jim S, Ambrose SH, Evershed RP (2004) Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen and apatite: implications for their use in palaeodietary reconstruction. Geochim Cosmochim Acta 68(1):61–72. https://doi.org/10.1016/S0016-7037(03)00216-3
Jones G, Bogaard A, Halstead P, Charles M, Smith H (1999) Identifying the intensity of crop husbandry practices on the basis of weed floras. Annu Br Sch at Athens 94:167–189
Jørgensen D (2013) Pigs and pollards: medieval insights for UK wood pasture restoration. Sustainability 5(2):387–399. https://doi.org/10.3390/su5020387
Kaupová S, Velemínský P, Stránská P, Bravermanová M, Frolíková D, Tomková K, Frolík J (2019) Dukes, elites, and commoners: dietary reconstruction of the early medieval population of Bohemia (9th–11th Century AD, Czech Republic). Archaeol Anthropol Sci 11:1887–1909. https://doi.org/10.1007/s12520-018-0640-8
Klápště J (2012) The Czech lands in medieval transformation. Brill, Leiden and Boston
Klápště J (2016) The archaeology of Prague and the Medieval Czech lands, 1100-1600. Equinox, Sheffield
Kočár P, Kočárová R (2013) Flóra a vegetace Nebovid. In: Havrda J, Tryml M (eds) Nebovidy. Středověká osada v pražském podhradí. Archeologické prameny k dějinám Prahy. Národní památkový ústav v hlavním městě Praze, Praha, pp 223-241
Kočár P, Čech P, Kozáková R, Kočárová R (2010) Environment and economy of the early medieval settlement in Žatec. Interdiscip Archaeol 1:45–60
Kohn MJ (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo) ecology and (paleo)climate. Proc Natl Acad Sci 107:19691–19695. https://doi.org/10.1073/pnas.1004933107
Komárek V (1993) Odhad věku domácích přežvýkavců. Institut výchovy a vzdělávání ministerstva zemědělství ČR v Praze, Praha
Kovačiková L, Trojánková O, Meduna P, Starec P, Burian M, Čiháková J, Frolík J (2019) Trendy v konzumaci masa a dalších živočišných produktů ve středověké Praze. Archeologické Rozhledy 71:529–552
Kovačiková L, Drtikolová Kaupová S, Poláček L, Velemínský P, Limburský P, Brůžek J (2020) Pig-breeding management in the early medieval stronghold at Mikulčice (eighth–ninth centuries, Czech Republic). Environ Archaeol. https://doi.org/10.1080/14614103.2020.1782583
Kozáková R (2008) Pollen analysis from Medieval flood loams and cultural layers deposited within an old channel of the River Vltava in Prague. In: Beneš J, Pokorný P (eds) Bioarchaeology in the Czech Republic. České Budějovice, Praha, pp 383–420
Kozáková R, Boháčová I (2008) Přírodní prostředí Pražského hradu a jeho zázemí v raném středověku – výpověď pylové analýzy sedimentů ze III. nádvoří. Archeologické Rozhledy 60:547–564
Kozáková R, Pokorný P (2007) Dynamics of the biotopes at the edge of a medieval town: pollen analysis of Vltava River sediments in Prague, Czech Republic. Preslia 79:259–281
Kozáková R, Pokorný P, Havrda J, Jankovská V (2009) The potential of pollen analyses from urban deposits: multivariate statistical analysis of a data set from the medieval city of Prague, Czech Republic. Veget Hist Archaeobot 18:477–488. https://doi.org/10.1007/s00334-009-0217-7
Krueger HW, Sullivan CH (1984) Models for carbon isotope fractionation between diet and bone. In: Turnlund JR, Johnson PE (eds) Stable Isotopes in Nutrition, ACS Symposium series 258. American Chemical Society, Washington, pp 205–220
Kuneš P, Abraham V (2017) History of Czech vegetation since the Late Pleistocene. In: Chytrý M, Danihelka J, Kaplan Z, Pyšek P (eds) Flora and Vegetation of the Czech Republic. Springer International Publishing, Cham, pp 193–227
Kyselý R (2015) Archeozoologická analýza raně středověkých kostí. In: Moucha V, Nechvátal B, Varadzin L et al. (eds.) Vyšehrad. Knížecí a královská akropole. Svědectví archeologie. Archeologický ústav AV ČR, Praha, pp 421-528
Legge AJ (1992) Excavations at Grimes Graves, Norfolk, 1972–1976: Animals, Environment and the Bronze Age Economy. British Museum Press, London
Leuschner Ch, Ellenberg H (2017) Ecology of Central European Forests. Vegetation Ecology of Central Europe, Volume I, Springer
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature 230:241–242. https://doi.org/10.1038/230241a0
Madgwick R, Mulville J, Stevens RE (2012) Diversity in foddering strategy and herd management in late Bronze Age Britain: an isotope investigation of pigs and other fauna from two midden sites. Environ Archaeol 17(2):126–140. https://doi.org/10.1179/1461410312Z.00000000011
Makarewicz CA (2014) Winter pasturing practices and variable fodder provisioning detected in nitrogen (δ15N) and carbon (δ13C) isotopes in sheep dentinal collagen. J Archaeol Sci 41:502–510. https://doi.org/10.1016/j.jas.2013.09.016
Millard AR, Jimenez-Cano NG, Lebrasseur O, Sakai Y (2013) Isotopic investigation of animal husbandry in the Welsh and English Periods at Dryslwyn Castle, Carmarthenshire, Wales. Int J Osteoarchaeol 23:640–650. https://doi.org/10.1002/oa.1292
O'Connell TC, Kneale CJ, Tasevska N, Kuhnle GG (2012) The diet-body offset in human nitrogen isotopic values: a controlled dietary study. Am J Phys Anthropol 149:426–434. https://doi.org/10.1002/ajpa.22140
O'Leary MH (1988) Carbon isotopes in photosynthesis. BioScience 38(5):328–336. https://doi.org/10.2307/1310735
Opravil E (1994) Příspěvek k poznání rostlinných makrozbytků ze staré Prahy. Archeologické rozhledy 46:105–114
Pardo LH, Nadelhoffer KJ (2010) Using nitrogen isotope ratios to assess terrestrial ecosystems at regional and global scales. In: West JB, Bowen GJ, Dawson TE, Tu KP (eds) Isoscapes. Understanding movements, patterns and process on earth through stable isotope mapping. Springer, New York, pp 221–249
Payne S (1973) Kill-off patterns in sheep and goats: the mandibles from Aşvan Kale. Anatol Stud 23:281–303. https://doi.org/10.2307/3642547
Peške L (1985) Domácí a lovná zvířata podle nálezů na slovanských lokalitách v Čechách. Sborník Národního muzea v Praze Řada A – Historie 39:209–216
Petráň J, Buchvaldek M (1985) Dějiny hmotné kultury 1. Státní pedagogické nakladatelství, Praha
Petráňová L, Petráň J (2000) Rolník v tradiční evropské kultuře. Set Out, Praha
Pokorná A, Houfková P, Novák J, Bešta T, Kovačiková L, Nováková K, Zavřel J, Starec P (2014) The oldest Czech fishpond discovered? An interdisciplinary approach to reconstruction of local vegetation in Mediaeval Prague suburbs. Hydrobiologia 730(1):191–213. https://doi.org/10.1007/s10750-014-1837-1
Pokorná A, Pokorný P, Meduna P (2019) Pátrání po rubu středověkého města. Botanická exkurze do Prahy pod Prahou. In: Haluzík R (ed) Město naruby. Vágní terén, vnitřní periferie a místa mezi místy. Academia, Praha, pp 63–84
Poschlod P (2015) The origin and development of the central European man-made landscape, habitat and species diversity as affected by climate and its changes–a review. Interdiscip Archaeol 6:197–221
Price M (2016) Pigs and power: pig husbandry in Northern Mesopotamia during the emergence of social complexity (6500-2000 BC). PhD. thesis. Harvard University, Cambridge Massachusetts
Sádlo J, Pokorný P, Hájek P, Dreslerová D, Cílek V (2008) Krajina a revoluce: významné přelomy ve vývoji kulturní krajiny českých zemí. Malá Skála, Praha
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639. https://doi.org/10.1016/0016-7037(84)90091-7
Šedivý J (2015) Zvieratá v stredovekom meste na príklade Bratislavy. In: Dvořáková D (ed) Človek a svet zvierat v stredoveku. VEDA Vydavateľstvo Slovenskej akadémie vied, Bratislava, pp 466–489
Smith BN, Epstein S (1971) Two categories of 13C/12C ratios for higher plants. Plant Physiol 47:380–384. https://doi.org/10.1104/pp.47.3.380
Součková M, Hejcman M, Klír T (2013) Medieval farming practices in deserted villages can be determined based on the nitrogen isotopic signature in recent forest soils. Interdiscip Archaeol 4:63–71
Sponheimer S, Robinson T, Ayliffe L, Roeder B, Hammer J, Passey B, West A, Cerling T, Dearing D, Ehleringer J (2003) Nitrogen isotopes in mammalian herbivores: hair δ15N values from a controlled feeding study. Int J Osteoarchaeol 13:80–87. https://doi.org/10.1002/oa.655
Starec P (2017a) Praha 1 – Staré Město, Křižovnická ulice čp. 71/I. In: Dragoun Z (ed) Archeologický výzkum v Praze v letech 2015–2016, Pražský sborník historický 45. Archiv hlavního města Prahy, Praha, p 635
Starec P (2017b) Praha 1 – Josefov, Pařížská ulice čp. 205/V. In: Dragoun Z. (ed) Archeologický výzkum v Praze v letech 2015–2016. Pražský sborník historický 45. Archiv hlavního města Prahy, Praha, p 615
Stevens RE, Lister AM, Hedges RE (2006) Predicting diet, trophic level and palaeoecology from bone stable isotope analysis: a comparative study of five red deer populations. Oecologia 149(1):12–21. https://doi.org/10.1007/s00442-006-0416-1
Sůvová Z, Cymbalak T, Kapustka K (2018) Nálezy obratlovců v předlokačních až novověkých souborech z pražské Národní třídy a z kolínského Karlova náměstí. Archaeol Pragensia 24:479–492
Szpak P (2014) Complexities of nitrogen isotope biogeochemistry in plant-soil systems: implications for the study of ancient agricultural and animal management practices. Front Plant Sci 5:1–19. https://doi.org/10.3389/fpls.2014.00288
Szpak P, Metcalfe JZ, Macdonald RA (2017) Best practices for calibrating and reporting stable isotope measurements in archaeology. J Archaeol Sci Rep 13:609–616. https://doi.org/10.1016/j.jasrep.2017.05.007
Tieszen LL (1991) Natural variations in the carbon isotope values of plants: implications for archaeology, ecology, and paleoecology. J Archaeol Sci 18:227–248. https://doi.org/10.1016/0305-4403(91)90063-U
Tieszen LL, Boutton TW, Tesdahl KG, Slade NA (1983) Fractionation and turnover of stable carbon isotopes in animal tissues: implications for δ13 C analysis of diet. Oecologia 57(1-2):32–37
Tomas J (1983) Řemeslníci 11.–13. století v českých zemích v písemných pramenech. Archaeologia Historica 8:73–84
Tonge CH, McCance RA (1973) Normal development of the jaws and teeth in pigs, and the delay and malocclusion produced by calorie deficiencies. J Anat 115(1):1–22
Valášková T (2014) Postavení Židů v českých královských městech v pozdním středověku. Master's thesis. Technical University of Liberec.
Valenzuela-Lamas S, Valenzuela-Suau L, Saula O, Colet A, Mercadal O, Subiranas C, Nadal J (2014) Shechita and Kashrut: Identifying Jewish populations through zooarchaeology and taphonomy. Two examples from Medieval Catalonia (North-Eastern Spain). Quat Int 330:109–117. https://doi.org/10.1016/j.quaint.2013.12.035
van Klinken GJ (1999) Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J Archaeol Sci 26:687–695. https://doi.org/10.1006/jasc.1998.0385
Zeder MA, Pilaar SE (2010) Assessing the reliability of criteria used to identify mandibles and mandibular teeth in sheep, Ovis, and goats, Capra. J Archaeol Sci 37:225–242. https://doi.org/10.1016/j.jas.2009.10.002
Žemlička J (1997) Čechy v době knížecí. Nakladatelství Lidové noviny, Praha
Žemlička J (2002) Počátky Čech královských 1198-1253. Proměna státu a společnosti. Nakladatelství Lidové noviny, Praha
Acknowledgements
The authors would like to thank Tereza Šálková for botanical consultation and Jaromír Kovárník for preparation of Fig. 1 and Anton Baer, Allastair Millar and Petra Maříková Vlčková for linguistic advice. The authors thank both of the reviewers for their valuable comments leading to the improvement of this paper.
Funding
The research project was supported by the Czech Science Foundation (GA ČR) under Grant 18-10003S and the Grant Agency of Charles University (GAUK project No. 20817).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 39 kb)
Rights and permissions
About this article

Cite this article
Kovačiková, L., Trojánková, O., Starec, P. et al. Livestock as an indicator of socioeconomic changes in Medieval Prague (Czech Republic). Archaeol Anthropol Sci 12, 283 (2020). https://doi.org/10.1007/s12520-020-01229-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-020-01229-5