
Abstract
Diminished ANK3 contributes to cell survival by inhibiting detachment-induced apoptosis. TP53BP1 that interacts with p53 and MFN1 that encodes a mitochondrial membrane protein are considered to have tumor suppressor gene (TSG) functions. HACD4 involving fatty acid synthesis and TCPL10 with transcription regulation functions are considered TSGs. Many genes involved in DNA methylations such as LCMT2, RNMT, TRMT6, METTL8 and METTL16 are often perturbed in cancer. The aim of our study was to find whether these genes were mutated in colorectal cancer (CRC). In a genome database, we observed that each of these genes harbored mononucleotide repeats in the coding sequences, which could be mutated in cancers with high microsatellite instability (MSI-H). For this, we studied 124 CRCs for the frameshift mutations of these genes and their intratumoral heterogeneity (ITH). ANK3, HACD4, TCP10L, TP53BP1, MFN1, LCMT2, RNMT, TRMT6, METTL8 and METTL16 harbored 11 (13.9%), 3 (3.8%), 0 (0%), 5 (6.3%), 1 (1.3%), 2 (2.5%), 4 (5.1%), 3 (3.8%), 2 (2.5%) and 2 (2.5%) of 79 CRCs with MSI-H, respectively. However, we found no such mutations in microsatellite stable (MSS) cancers in the nucleotide repeats. There were ITH of the frameshift mutations of ANK3, MFN1 and TP53BP1 in 1 (6.3%), 1 (6.3%) and 1 (6.3%) cases, respectively. Our data exhibit that cancer-related genes ANK3, HACD4, TP53BP1, MFN1, LCMT2, RNMT, TRMT6, METTL8 and METTL16 harbor mutational ITH as well as the frameshift mutations in CRC with MSI-H. Also, the results suggest that frameshift mutations of these genes might play a role in tumorigenesis through their inactivation in CRC.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Ankyrins are a family of proteins that link the integral membrane proteins to the underlying spectrin-actin cytoskeleton and play key roles in cell motility, activation, proliferation, contact, and maintenance of membrane integrity [1, 2]. In cancers, somatic mutations across its entire coding region have been described as driver mutations in breast carcinoma, chronic lymphocytic leukemia, cutaneous melanoma and uterine carcinoma. 1.75% of colorectal cancers (CRCs) carry non-driver mutations of ANK3 (https://www.intogen.org). Epithelial-mesenchymal transition (EMT) downregulates ANK3, a cytoskeletal protein, subsequently represses the tumor suppressor p14ARF and induces apoptosis resistance [3]. In addition, downregulation of ANK3 correlates with poor prognosis in diverse human cancers [4].
TP53BP1 binds with damaged chromatin and carries out multiple roles such as recruitment of DNA double strand break (DSB)-responsive proteins [5]. Apart from the DNA repair, TP53BP1 interacts with p53 and contributes to tumor suppressor function of p53 [6]. In colorectal cancer (CRC), decreased TP53BP1 expression is associated with poor survival of the patients [7]. Mitofusin 1 (MFN1) encodes a mitochondrial membrane protein that plays roles in mitochondria fusion and apoptosis [8]. ERK, which is constitutively upregulated in cancers, promotes phosphorylation of MFN1 protein and induces apoptosis [8].
HACD4 belongs to a family of enzymes that catalyze the dehydration step of very long chain fatty acid synthesis [9]. Knockdown of HACD4 increases STAT3 phosphorylation and cell proliferation [10]. TCPL10 encoding T-complex protein 10-like, a nuclear protein with transcription regulation functions, suppresses colony formation, cell cycle progression through G0/G1 phase and cell growth in vivo in liver [11]. Global and gene-specific DNA methylations are often perturbed in cancer. For example, a DNA methyltransferase-encoding gene DNMT3A is frequently mutated in acute myelogenous leukemia [12]. However, alterations of many other methyltransferase genes, such as carboxyl methyltransferase 2 (LCMT2), RNA guanine 7-methyltransferase (RNMT), methyltransferase like protein 16 (METTL16), methyltransferase like protein 8 (METTL8) and tRNA methyltransferase 6 (TRMT6) remain unclarified in cancer development. Together, these data suggest that genes listed above may possess tumor suppressor gene (TSG) activities.
About one third of CRCs are classified as high microsatellite instability (MSI-H) cancers [13]. Many TSGs harbor frameshift mutations at monocleotide repeats in MSI-H cancers [13]. In the human genome database, we observed that ANK3, HACD4, TCP10L, TP53BP1, MFN1, LCMT2, RNMT, TRMT6, METTL8 and METTL16 genes possess nucleotide repeats in coding sequences that might be mutated in MSI-H cancers. However, frameshift mutations of the repeats in these genes in CRC have not been reported. Intratumoral heterogeneity (ITH) is a common phenomenon in cancers, which may result in cancer evolution and influence on clinical outcomes [14]. Thus, identification of genetic ITH is important in understanding biological and clinicopathologic features of the cancers. The current study aimed to find whether ANK3, HACD4, TCP10L, TP53BP1, MFN1, LCMT2, RNMT, TRMT6, METTL8 and METTL16 frameshift mutations are common and harbor ITH in MSI-H CRC.
Materials and Methods
Tissue Samples and Microdissection
In this study, we used 124 CRCs that consisted of 45 CRCs with microsatellite stable (MSS) and 79 CRCs with MSI-H. These samples overrepresent MSI-H cases due to our collection method (MSI-H and MSS cases were separately collected in different times). We adopted an MSI evaluation system using five mononucleotide repeats [15]. For 16 MSI-H CRCs, we picked 4–7 tumor areas and one normal area per CRC for the ITH analysis. Each ITH area was histologically confirmed under light microscope. These ITH areas were studied for detecting mutational ITH of ANK3, HACD4, TCP10L, TP53BP1, MFN1, LCMT2, RNMT, TRMT6, METTL8 and METTL16. Pathologic features of the cancers were evaluated in all blocks of all cases by a pathologist and are summarized in Table 1. Tumor and normal cells were microdissected as described previously [16]. Approval of this study was obtained from the institutional review board of the Catholic University of Korea.
Single Strand Conformation Polymorphism (SSCP) Analysis
We analyzed DNA sequences in ANK3 (one A8 and two A7), HACD4 (A8), TCP10L (G7), TP53BP1 (A7), MFN1 (A7), LCMT2 (T7), RNMT (A7), TRMT6 (A8), METTL8 (A7) and METTL16 (A7 and A6). Genomic DNA was amplified using polymerase chain reaction (PCR). [32P]dCTP was incorporated to the PCR products for visualization in autoradiogram. We determined aberrant gel motility in the SSCP (FMC Mutation Detection Enhancement system; Intermountain Scientific, Kaysville, UT, USA) using visual inspection, which subsequently sequenced by Sanger DNA sequencing (3730 DNA Analyzer, Applied Biosystem, Carlsbad, CA, USA). Other procedures in detail were described in our earlier studies [17, 18].
Results
Mutational Analysis
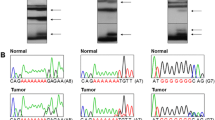
Genomic DNAs of the 124 CRCs (79 with MSI-H and 45 with MSS) were studied to detect frameshift mutations in the mononucleotide repeats by the PCR-SSCP analyses. On SSCP and Sanger sequencing, we observed that of ANK3, HACD4, TCP10L, TP53BP1, MFN1, LCMT2, RNMT, TRMT6, METTL8 and METTL16 harbored 11 (13.9%), 3 (3.8%), 0 (0%), 5 (6.3%), 1 (1.3%), 2 (2.5%), 4 (5.1%), 3 (3.8%), 2 (2.5%) and 2 (2.5%) of 79 CRCs with MSI-H, respectively (Table 2). Normal tissues of the corresponding patients from the slides were microdissected and analyzed, but there was no evidence of the mutations on SSCP and Sanger sequencing, which indicated that the mutations had risen somatically (Fig. 1). All detected mutations exhibited both wild-type and mutant signals in SSCP and Sanger sequencing, indicating that they were heterozygous mutations (Fig. 1).

Representative SSCP and DNA sequencings of repeats in ANK3 in colon cancers. SSCP (a) and DNA sequencing analyses (b) of ANK3 from tumor (Lane T) and normal tissues (Lane N). a. SSCPs of the PCR products from A7 repeat in exon 1 (left), A7 repeat in exon 37 (middle) and A8 repeat in exon 37 (right) show aberrant bands (arrows in lane T) as compared to SSCP from normal tissues (N). b. Direct DNA sequencing analyses (b) show heterozygous A deletions in exon 1 (left), A7 repeat in exon 37 (middle) and A8 repeat in exon 37 (right) in tumor tissues as compare to normal tissues
The frameshift mutations detected in this study were found in MSI-H CRCs, but there was none in MSS CRCs (Table 2) (Fisher’s exact test, p < 0.001). There was no significant association of these mutations with the grade, differentiation and tumor stage. In those with MSI-H, no correlation was observed between histological features (medullary pattern, mucinous histology and tumor-infiltrating lymphocytes) and the mutations.
Intratumoral Heterogeneity of the Frameshift Mutations
Ninety-six areas from 16 CRCs with MSI-H were analyzed to find ITH in the frameshift mutations described above. We observed that the frameshift mutations of ANK3, MFN1 and TP53BP1 showed regional ITH in 1 (6.3%), 1 (6.3%) and 1 (6.3%) cases, respectively (Table 3 and Fig. 2). There was no significant histological difference between mutation (+) and mutation (−) ITH areas. HACD4, TCP10L, LCMT2, RNMT, TRMT6, METTL8 and METTL16 frameshift mutations did not harbor the ITH in these CRCs.

Intratumoral heterogeneity of the ANK3 frameshift mutation in a colon cancer. A: Direct DNA sequencings show ANK3 c.9349delA mutation (MT) in two regional biopsies (53–1 and 53–6) and wild-type (WT) ANK3 in the other five regional biopsies (53–2, 53–3, 53–4, 53–5 and 53–7)
Discussion
In EMT, one of the most crucial phenotypes is alteration of cytoskeletal proteins, including loss of E-cadherin and cytokeratin expression [19]. Based on earlier reports that showed inactivation of ANK3 was related to EMT and to apoptosis resistance [3], we attempted to disclose whether somatic frameshift mutations of ANK3 gene was present in CRC. In the present study, we found ANK3 frameshift mutations in 13.9% of CRCs with MSI-H. TSG or anti-oncogene is a gene that protects a cell from one step on the path to cancer [20]. When a TSG is mutated, the cells can progress to cancer in combination with other genetic or epigenetic changes. Earlier studies identified that HACD4, TP53BP1 and MFN1 possessed TSG activities in cells [5,6,7,8,9,10,11]. Loss of cell polarity is a typical hallmark of cancer development and progression in epithelial cells [5]. The present study here identified that MSI-H CRC exhibited frameshift mutations within the repeats of HACD4, TP53BP1 and MFN1 in CRC. These results indicate that the functions of these genes are altered by frameshift mutations identified in this study and suggest that these inactivating mutations might inhibit their TSG functions in MSI-H CRC as well. These frameshift mutations might play a role in the pathogenesis of MSI-H CRC by inhibiting their TSG activities. Also, the inactivating mutations of methyltransferase genes MFN1, LCMT2, RNMT, TRMT6, METTL8 and METTL16 might possibly play a role in tumorigenesis of CRC. However, consequences of the inactivating mutation of methyltransferase genes in MSI-H cancers remain unclear, because their normal biological functions are not known. Of importance, there was a significant difference in mutation frequencies between MSI-H and MSS cancers, indicating that these frameshift mutations with MSI-H were specific
All of the mutations detected in the present study would result in premature stops of amino acid synthesis in the proteins and hence resemble a typical loss-of-function mutation. It is also possible that both of these would be expected to lead to premature stop codons in the mRNAs but not necessarily the production of a truncated protein as these transcripts are more likely to be degraded by nonsense mediated decay [13]. Since cancers with MSI show extremely severe mutation phenotype with frameshift mutation in microsatellite sequences, the frameshift mutations in this study could possibly be passenger mutations. By contrast, many TSGs have been found to harbor mutations at nucleotide repeats in the coding sequences in the cancers with MSI (BAX, type II transforming growth factor β receptor, caspase-5, hMSH3 and TCF4) [21]. We were not able to find any significant difference in clinocopathologic parameters between those with and without the mutations probably due to the small number of mutated cases. Analyses of a larger cohort with the mutations are required in future studies.
In this study, we identified mutational ITH of ANK3, TP53BP1 and MFN1 in CRCs, which is consistent with previous studies that had reported frequent mutational ITH in CRCs with MSI-H [22]. Genomic instability in a cancer may result in an elevated level of somatic mutations and contribute to ITH development by providing a pool of mutations upon which selection can act in a given microenvironment [14, 23]. The cancer ITH of driver mutations could result in poor clinical outcomes in patients as well. For instance, single or a group of mutations with a metastasis potential might redirect clinical outcomes since such rare clones may accomplish dominance during tumor progression [14, 23]. It is reasonable to conceive that other regions of these genes as well as other genes besides ANK3, TP53BP1 and MFN1 could harbor mutational ITH in these CRCs. Based on the cancer-related properties of the genes, our results suggest that loss of their properties may have regional heterogeneity in CRC that could be further selected and influence the clinical outcomes. However, presence of ITH of the frameshift mutations might suggest a possibility that there could be a mixed or ameliorated effect of the frameshift mutations in MSI-H cancers.
References
Bennett V, Baines AJ (2011) Spectin and ankyrin-based pathways: metazoan inventions for integrating cells into tissues. Physiol Rev 81:1353–1392
Hryniewicz-Jankowska A, Czogalla A, Bok E, Sikorsk AF (2002) Ankyrins, multifunctional proteins involved in many cellular pathways. Folia Histochem Cytobiol 40:239–249
Kumar S, Park SH, Cieply B, Schupp J, Killiam E, Zhang F, Rimm DL, Frisch SM (2011) A pathway for the control of anoikis sensitivity by E-cadherin and epithelial-to-mesenchymal transition. Mol Cell Biol 31:4036–4051
Glinsky GV, Berezovska O, Glinskii AB (2005) Microarray analysis identifies a death-from-cancer signature predicting therapy failure in patients with multiple types of cancer. J Clin Invest 115:1503–1521
Panier S, Boulton SJ (2014) Double-strand break repair: 53BP1 comes into focus. Nat Rev Mol Cell Biol 15:7–18
Cuella-Martin R, Oliveira C, Lockstone HE, Snellenberg S, Grolmusova N, Chapman JR (2016) 53BP1 integrates DNA repair and p53-dependent cell fate decisions via distinct mechanisms. Mol Cell 64:51–64
Bi J, Huang A, Liu T, Zhang T, Ma H (2015) Expression of DNA damage checkpoint 53BP1 is correlated with prognosis, cell proliferation and apoptosis in colorectal cancer. Int J Clin Exp Pathol 8:6070–6082
Pyakurel A, Savoia C, Hess D, Scorrano L (2015) Extracellular regulated kinase phosphorylates mitofusin 1 to control mitochondrial morphology and apoptosis. Mol Cell 58:244–254
Ikeda M, Kanao Y, Yamanaka M, Sakuraba H, Mizutani Y, Igarashi Y, Kihara A (2008) Characterization of four mammalian 3-hydroxyacyl-CoA dehydratases involved in very long-chain fatty acid synthesis. FEBS Lett 582:2435–2440
Zhu S, Wang Z, Zhang Z, Wang J, Li Y, Yao L, Mei Q, Zhang W (2014) PTPLAD2 is a tumor suppressor in esophageal squamous cell carcinogenesis. FEBS Lett 588:981–989
Zuo J, Cai H, Wu Y, Ma H, Jiang W, Liu C, Han D, Ji G, Yu L (2014) TCP10L acts as a tumor suppressor by inhibiting cell proliferation in hepatocellular carcinoma. Biochem Biophys Res Commun 446:61–67
Yan XJ, Xu J, Gu ZH, Pan CM, Lu G, Shen Y, Shi JY, Zhu YM, Tang L, Zhang XW, Liang WX, Mi JQ, Song HD, Li KQ, Chen Z, Chen SJ (2011) Exome sequencing identifies somatic mutations of DNA methyltransferase gene DNMT3A in acute monocytic leukemia. Nat Genet 43:309–315
Imai K, Yamamoto H (2008) Carcinogenesis and microsatellite instability: the interrelationship between genetics and epigenetics. Carcinogenesis 29:673–680
Marusyk A, Almendro V, Polyak K (2012) Intra-tumour heterogeneity: a looking glass for cancer? Nat Rev Cancer 12:323–334
Murphy K, Zhang S, Geiger T, Hafez MJ, Bacher J, Berg KD, Eshleman JR (2006) Comparison of the microsatellite instability analysis system and the Bethesda panel for the determination of microsatellite instability in colorectal cancers. J Mol Diagn 8:305–311
Yoo NJ, Kim HR, Kim YR, An CH, Lee SH (2012) Somatic mutations of the KEAP1 gene in common solid cancers. Histopathology 60:943–952
Je EM, Kim MR, Min KO, Yoo NJ, Lee SH (2012) Mutational analysis of MED12 exon 2 in uterine leiomyoma and other common tumors. Int J Cancer 131:E1044–E1047
Jo YS, Choi MR, Song SY, Kim MS, Yoo NJ, Lee SH (2016) Frameshift mutations of HSPA4 and MED13 in gastric and colorectal cancers. Pathol Oncol Res 22:769–772
Thiery JP, Sleeman JP (2006) Complex networks orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell Biol 7:131–142
Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Nature Reviews Cell 144:646–674
Calin GA, Gafà R, Tibiletti MG, Herlea V, Becheanu G, Cavazzini L, Barbanti-Brodano G, Nenci I, Negrini M, Lanza G (2000) Genetic progression in microsatellite instability high (MSI-H) colon cancers correlates with clinico-pathological parameters: a study of the TGRbetaRII, BAX, hMSH3, hMSH6, IGFIIR and BLM genes. Int J Cancer 89:230–235
Choi YJ, Kim MS, An CH, Yoo NJ, Lee SH (2014) Regional bias of intratumoral genetic heterogeneity of nucleotide repeats in colon cancers with microsatellite instability. Pathol Oncol Res 20:965–971
Choi YJ, Rhee JK, Hur SY, Kim MS, Lee SH, Chung YJ, Kim TM, Lee SH (2017) Intraindividual genomic heterogeneity of high-grade serous carcinoma of the ovary and clinical utility of ascitic cancer cells for mutation profiling. J Pathol 241:57–66
Acknowledgements
This study was supported by grants from Korea Research Foundation (2012R1A5A2047939 and 2017R1A2B2002314).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of Interest
None
Rights and permissions
About this article

Cite this article
Yeon, S.Y., Jo, Y.S., Choi, E.J. et al. Frameshift Mutations in Repeat Sequences of ANK3, HACD4, TCP10L, TP53BP1, MFN1, LCMT2, RNMT, TRMT6, METTL8 and METTL16 Genes in Colon Cancers. Pathol. Oncol. Res. 24, 617–622 (2018). https://doi.org/10.1007/s12253-017-0287-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12253-017-0287-2