
Abstract
The natural processes of mudflat accretion, vegetation succession, and plant invasion and associated anthropogenic impacts make the Yangtze Estuary an ideal area for the study of salt marsh dynamics. In order to understand the biotic and abiotic factors regulating the spatiotemporal dynamics of salt marsh vegetation in relation to the expansion of the exotic plant (Spartina alterniflora) and the consequent responses of native species (Phragmites australis and Scirpus mariqueter), a grid-based model was developed incorporating the key ecological processes at Chongming Dongtan and Jiuduansha wetlands. During the simulation period of 2000–2008, the area of S. mariqueter decreased significantly at both wetlands, due to the rapid expansion of S. alterniflora. The expansion rate of P. australis was slow. When compared with observations, the simulated spatiotemporal dynamics of salt marsh vegetation showed a percentage match of 73–91 %. The accuracy of the distribution area for each salt marsh species was even higher (89–97 %). Both simulation and observation revealed that the period of early salt marsh succession was crucial for colonization and establishment of S. alterniflora after its introduction. Projections for the period 2008–2015 indicated that the expansion rate of S. alterniflora would slow down. In contrast, the model predicted that the distribution of S. mariqueter and P. australis would increase at a steady pace. The probable reason behind this is that the accretion of habitat above the elevation threshold at the current rate of sedimentation is marginal compared to the previously rapid rate of habitat colonization by S. alterniflora over the past decade. This study indicates the model’s potential for simulating and predicting the dynamics of salt marsh vegetation in the Yangtze Estuary and demonstrates that, when appropriately parameterized, the model could be successfully applied elsewhere. Finally, the limitations of the model and its potential for monitoring and controlling invasive species in coastal wetlands are briefly discussed.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Salt marshes are vulnerable ecosystems threatened both by human activity and biological invasions (Gedan et al. 2009). Increasing human mobility has resulted in species introductions to previously unconnected areas and the introduced species may spread exponentially and have a major impact on biodiversity and ecosystem functioning (Levin et al. 2006; Wang et al. 2010; Weidenhamer and Callaway 2010).
During the past decade, the spatiotemporal dynamics of vegetation dispersal and establishment have been successfully incorporated into modeling simulations (Grevstad 2005; Huang et al. 2008; Fennell et al. 2012). Recently, increasing attention has been paid to mechanistic models, which can be used to investigate the environmental factors and habitat features controlling the range expansion of invasive species through an environment and to predict their potential expansion and impact on plant community dynamics (Sebert-Cuvillier et al. 2010; Coutts et al. 2011; Fennell et al. 2012). Furthermore, mechanistic models are increasingly important as a tool to assist management agencies in the design of efficient and effective control measures for reducing invasive spread (Coutts et al. 2011; Emry et al. 2011).
In China, the introduction of Spartina alterniflora from North America to coastal wetlands is a prime example of a spatially structured invasion (Huang et al. 2008; Zhu et al. 2012). This exotic species was introduced to the Yangtze Estuary in the 1990s and has expanded rapidly thereafter and the mono-dominant community of S. alterniflora now accounts for almost 25 % of the total intertidal salt marsh vegetation (Li et al. 2006; Huang et al. 2008). The invasion of S. alterniflora has had significant competitive effects on the native plant communities of Scirpus mariqueter and Phragmites australis (Huang et al. 2008), has strongly influenced the composition of the local macrobenthic communities (Wang et al. 2010), and has had important impacts on the stopover habitats for migratory shorebirds.
A number of studies on the underlying mechanisms and impacts of biotic and abiotic factors on the range expansion of salt marshes in the Yangtze Estuary have recently been conducted. Huang et al. (2008) tested the hypothesis of habitat pre-emption by the exotic at the expense of the native salt marsh species. The seed production, soil seed bank, and germination of S. alterniflora were studied along the intertidal gradient (Xiao et al. 2009). The range expansion patterns and the roles which sexual reproduction and asexual propagation played were investigated for S. mariqueter and S. alterniflora (Xiao et al. 2010a, b). The impact of bio-physical processes, such as recruitment, establishment, growth and reproduction of plants, mudflat dynamics and hydrodynamic conditions, on range expansion has also been studied along the typical fronts of salt marshes (Schwarz et al. 2011; Zhu et al. 2012).
Based on the reults of this previous research, a grid-based model was developed for the simulation of the spatiotemporal dynamics of salt marsh vegetation in the Yangtze Estuary. In the model, the multiple-resource parameters of biotic factors (including the seed bank and germination, duration of growing period, reproduction and establishment, clonal integration, and interspecies competition) and abiotic processes (including the tidal current for dispersal, the sedimentary regime, and inundation stress) for each species were incorporated for simulation. The performance of the model was evaluated on two landscape-scale coastal wetlands (Chongming Dongtan and Jiuduansha) in the Yangtze Estuary, and checked against existing salt marsh vegetation maps derived from remote sensing data for the period 2000–2008. Also, a model projection was conducted up to 2015. The main objective of this study was to evaluate the model that incorporated key ecological processes for simulating the spatiotemporal dynamics of vegetation at the two sites in the Yangtze Estuary and to test whether this model is applicable elsewhere, beyond the calibration site. The secondary goal was to predict the future patterns of salt marsh vegetation given the current sedimentation rate in the Yangtze Estuary. In addition, limitations of the model and its utilization potential for invasive species controlling are briefly discussed.
Materials and Methods
Study Area
The Yangtze Estuary is a typical medium-sized tidal estuary with multi-order bifurcations, shoals and sand bars (Yang 1999). The region of mouth bars in the estuary and the submerged delta nearby are major locations of sedimentation resulting from the large amount of silt brought down by the Yangtze River, which has created extensive areas of shoals and tidal flats that have been colonized by various types of salt marsh vegetation (Huang and Zhang 2007).
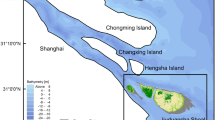
Chongming Dongtan wetland (hereafter CDW) is located on the eastern fringe of Chongming Island, between 31°25′ and 31°38′ N and 121°50′ and 122°05′ E and with an area of 242 km2 above the 0 m isobaths (Fig. 1) after the last reclamation. Jiuduansha wetland (hereafter JW) is an isolated shoal, between 31°03′ and 31°17′N, 121°46′ and 122°15′E, and covers 127 km2 above the 0 m isobaths (Fig. 1), which has received minimal human impact and has remained in a near-natural condition since it was formed in 1920s (Huang et al. 2007). The highest elevation of CDW and JW is around 4.0–4.2 m. These two wetlands are prime examples of where spatially structured invasion of S. alterniflora in the Yangtze Estuary has occurred, for which model performance can be evaluated against existing empirical observations of salt marshes dynamics.

Location of the Chongming Dongtan (CDW) and Jiuduansha (JW) wetlands in the Yangtze Estuary, with digital elevation maps of 2000 and sedimentary regime (1997–2010 investigation documents by the Survey Bureau of Hydrology and Water Resources of the Changjiang River Estuary)
Model Design
To simulate the spatiotemporal dynamics of salt marshes (including both the exotic and native species) on the CDW and JW wetlands, the model was developed on the Matlab® matrix platform. The model consisted of a spatial lattice matrix of interconnected cells (resolution 1 × 1 m), which were connected to all neighboring cells, with a distribution probability of vegetation spread associated with each connection. In the cell matrix, the native species (P. australis and S. mariqueter) and the exotic species (S. alterniflora) were coded with different numbers for identification and cells with null values indicated bare mudflat. Each cell evolves according to transition rules that depend on the state of its neighboring cells.
The spatial and temporal pattern of salt marsh vegetation is a product of synergetic functions based on biotic and abiotic processes, mainly including seed bank deposition and germination, seedling dispersal, individual establishment and survival, tolerance to inundation stress, clonal integration, and inter-specific competition (Fig. 2). Simulation of the salt marsh vegetation dynamics takes place in continuous time steps and a monthly time step was used to identify the growing (active period) and non-growing seasons (inactive period).

Simplified schematic for model building and testing
S. alterniflora was introduced in the Yangtze Estuary around 1997 (Li et al. 2006; Huang et al. 2008). After the initial colonization stage, the year 2000 marked the onset of rapid population growth and range expansion. Therefore, the observed salt marsh habitat maps, including vegetation area, bare mudflat above 0 m, and seawater area at −2 to 0 m for the year 2000 in both the study areas were used to produce the initialized matrix in the model (Huang et al. 2008; Wang et al. 2010), which were generated as the cell-matrix maps for simulation.
The Sedimentary Regime in Relation to Mudflat Elevation Changes
The simulation of vegetation expansion and establishment depends on mudflat accretion (see “The growing period and simulation of reproduction” section). Therefore, the input dataset for the model included the initial distribution maps of salt marsh vegetation, digital elevation maps (−2 to 4.2 m) and sedimentation maps of CDW and JW (Fig. 1). The spatial depositing regime presents the annual mean sediment rates for different parts of the Yangtze Estuary during 1997–2010, which were measured and documented by the Changjiang Water Resources Commission (CWRC 2012).
The model simulated mudflat accretion by determining the amount of sediment deposited and calculating the increase in elevation during each period of one year, based on the seasonal variations (flood and dry seasons) observed on monitoring strips established previously at CDW (Zhu et al. 2012). Using the data from these strips, the monthly percentage of sedimentary rate for elevation accretion was estimated according to the distribution shown in Table 1. In order to estimate spatially the vertical elevation changes at each cell, the model divides the geographic matrix (S) with the annual mean sedimentary rate into 12 month-specific components (S i , i = 1–12 months) to derive the mudflat elevation (E) on the elevation matrix and the subsequent increase in lateral mudflat area (when E is above 0 m) of CDW and JW on the habitat matrix at each time-step (t).
The growing period and simulation of reproduction
In the model, the period March–October was defined as the growing season, including the processes of seed germination, seedling dispersal and establishment, vegetative growth and clonal integration and seed bank deposition within the salt marsh vegetation (Zhang and Yong 1992). The residual months (November–February) corresponded to the inactive period for plants.
Empirical measurements for the seasonal seed bank dynamics and germination rate (listed in Table 2) recorded from March to October within the growing season during 2007–2010 were used to drive the model. Although the highest density of seeds in the soil seed bank were recorded in autumn and winter (Xiao et al. 2009), most of seeds (c.96–98 %) are lost before the next growing season, due to fungal attack, decay, and tidal overwash (Xiao et al. 2009; Zhu et al. 2012). Furthermore, low temperature during winter season limited germination for the seeds (Xiao et al. 2009; Zhu et al. 2011).
Dispersal and Establishment
In the Yangtze Estuary (Xiao et al. 2010a, b) and Willapa Bay, USA (Davis et al. 2004), it was shown that the salt marsh vegetation will spread to new habitats by seeds and seedlings that float on the tide. For P. australis and S. alterniflora, the model divided the growing season of vegetation into two periods of expansion, including a period of “long-distance” seedling dispersal by tidal currents (through growing season, Fig. 3a) followed by a period of vegetative growth and clonal integration (May–October, Fig. 3b). The pioneer native plant S. mariqueter was assumed to spread mainly through its corms and rhizomes, since few seeds are found in the seed banks on the bare mudflats (Zhang and Yong 1992).

The two different expansion periods: a seedling dispersal and b clonal integration of the salt marsh vegetation for model simulation
In the matrix of seedling dispersal, cell values are considered as occupied (1) by the seedlings or not (0) and are dispersed from meadows (occupied cells) to the bare mudflats (cells without plants) in the corresponding matrix. The number of living seedlings (N) produced for dispersal was regarded as equal to the seedlings that germinated in each month during a growing season and was determined by a sum function of vegetation distribution area on salt marsh (A), density of seed bank (D seed) and germination rate (R g) for each species in the corresponding row (month) (dataset in Table 2).
The seedlings were distributed via a random allocation process and allowed for multiple seedlings of each species in each cell.
During the growing season, the model assumed that the seedlings of P. australis and S. alterniflora could be transported in any of eight directions by the tidal currents as a stochastic process. The seedling amount distributed from the original cells (V i.j) within eight directions around each cell was based on a separable space-time Poisson distribution for determining number of dispersal events in the region of each direction (Janine et al. 2008):

where λ is the arrival rate, t is the time step, x is a random direction, V is the spatial matrix for seedling dispersal, and f(x) represents the spatial probability density function of these stochastic events.
The seedling dispersal event was assigned a dispersal distance derived from the previous field measurements for the different species. The mean transportation distance of seedlings from the vegetation meadow over one growing season was 60–100 m for S. alterniflora and around 10 m for P. australis (Li et al. 2006; Huang and Zhang 2007; Xiao et al. 2010a, b; Zhu et al. 2012).
Once seedlings become established, they form tussocks quickly by vegetative tillering and growth of rhizomes, finally merging into extensive meadows. The clonal integration by tillers and rhizomes was modeled as a neighboring spreading of plant cells, based on the expansion mode of “Moore neighborhood” comprising the eight cells surrounding a central cell (Wolfram 1986). The growth and horizontal spread of rhizomes can reach around 1 m per month during the period May–September (Xiao et al. 2010a, b; Zhu et al. 2012). Accordingly, the monthly (one time step) rate of spread was incorporated into the model by defining the radius of the “Moore neighborhood” as 1 for the salt marsh vegetation.
Environmental Stress and Tolerance
The probability of survival of seedlings surviving on a patch depends on the suitability of the habitat and the environmental stress. Empirically, the establishment probability of seedlings was dependent on the hydrodynamic conditions of the mudflat front, i.e. the closer patch is to the vegetation meadow with a higher elevation and weak tidal flow intensity, the higher the number and density of seedlings established. On the contrary, the nearer a patch is to the sea with a lower elevation and stronger tidal flow intensity, the fewer the number and density of seedlings established (Xiao et al. 2009, 2010a, b; Schwarz et al. 2011; Zhu et al. 2012). As described in Fig. 4a, the probability (P) of seedling establishment is estimated by an asymptotic diminishing function of dispersal distance (d, distance from meadow front seaward to dispersal boundary).

Mean daily inundation duration (curve) in relation to intertidal elevation (using the local Wushong bathymetric benchmark) in the Yangtze Estuary. The gray belt indicates the survival front of P. australis, S. alterniflora and S. mariqueter observed in the field
Inundation duration is also a significant stress for salt marsh plants. Once again, from previous field surveys and measurements, differences were found in the tolerance of seedlings or adults to submergence (Zhang and Yong 1992; Li et al. 2006; Huang et al. 2008; Zhu et al. 2012), based on the mean duration of daily tidal inundation in relation to mudflat elevation above the local Wushong bathymetric benchmark (Fig. 4b).
In order to set the spreading and survival limits for the seedlings of salt mash plants in the model, the mudflat elevation of the study area was assumed to reflect the mean daily inundation duration. The elevation matrix map was then overlain on top of the matrix of plant distribution to determine the habitable cells for plant survival and establishment. A discriminant function was employed to assign a value of 1 (true) or 0 (false) to decide whether the grid could be occupied by each of the salt mash plants, taking the elevation (E) into account.
Interspecific Competition
Chen et al. (2004) reported that S. alterniflora was a strong competitor with S. mariqueter. The introduction of S. alterniflora to a S. mariqueter-dominated community resulted in a significant decrease in the abundance, coverage, seed production, and fresh corm output of S. mariqueter during the growing season. Data obtained through remote sensing also showed that S. mariqueter was almost completely replaced by S. alterniflora at the monitoring sites in the Yangtze Estuary (Huang and Zhang 2007; Li et al. 2009). In contrast, P. australis and S. alterniflora generally showed non-aggression to each other and share similar niches in the intertidal zones (Wang et al. 2006).
In the model, the interspecific competitive balance between the salt marsh vegetation species for habitats (space pre-emption, succession, and inhibition) was incorporated into the simulation procedures. As presented in Fig. 5, the cells representing bare mudflat with null value could be colonized by all of the three salt marsh species, and then, the outcome of interspecific competition was used to regulate the spatial pattern of habitat occupancy and species replacement.

Simplified interpretation of interspecies competition from time step of t to the next one t + 1. Abbreviation: SA—S. alterniflora, PA—P. australis, SM—S. mariqueter, blank—bare mudflat
Data Sources for Model Calibration
A number of other research studies conducted on the salt marshes of the Yangtze Estuary lie behind the parameterization and calibration of the current version of the model and further details are presented in Table 3. The model parameters (or running rules) have either been measured or estimated in previous research performed in relation to the growing periods and reproduction strategies of the species, their seed banks and germination characteristics, seed/seedling dispersal, seedling survival and establishment, clonal integration and expansion range, environmental stress, tolerance, and interspecific competition.
Model Simulation and Evaluation
The initial salt marsh vegetation maps of 2000 and the digital elevation maps and annual sedimentary regime for the study areas of CDW and JW (Fig. 1) were entered to an 8-year simulation. The simulations for each year were repeated ten times to test the effects of stochastic elements. The published spatiotemporal dynamics of salt marsh vegetation for 2004, 2006, and 2008 (Li et al. 2006; Huang and Zhang 2007; Huang et al. 2008; Wang et al. 2010) were then used to evaluate the performance of model. The assessment was based on a comparison between the simulated (M odeled) and the observed vegetation distribution matrix (M observed) in the corresponding year. Calculation of the percentage difference (%DIFF) between the simulated and observed salt marsh vegetation maps was implemented in the Matlab® platform by overlaying the classified salt marsh vegetation maps and the simulation images in the corresponding years.
Furthermore, the simulations were run until 2015 to predict the potential future spatiotemporal dynamics of the salt marsh vegetation at CDW and JW, using the same empirically derived drivers and parameters.
Results
The Spatiotemporal Dynamics of Salt Marsh Vegetation
With the continuous monthly time-step, the model was able to separate the simulation period into two stages of salt marsh vegetation dynamics: firstly, over the past decade and secondly, into the future using both the growing (March–October) and non-growing seasons (November–February) (Fig. 6). With ten repeated simulations, the deviations of vegetation distribution area ranged from 0.9 to 4.8 % for S. mariqueter, from 1.1 to 6.2 % for S. alterniflora and from 0.7 to 4.5 % for P. australis regardless of study sites. Visual interpretation of the maps for each of the 10 simulations showed little variation in the spatial distribution of salt marsh vegetation on CDW and JW at each simulation period (maps not presented). Therefore, for each year of simulations, one of the 10 matrix outputs was selected as a representative example, as shown in Fig. 7. The model produced a similar distribution pattern for the period of 2000–2008 against the observed vegetation maps of 2004, 2006, and 2008. At the study site of CDW, the introduced species S. alterniflora invaded rapidly eastward from a northern initial location. Both the simulated and observed salt marsh vegetation maps showed that this rapid range expansion of S. alterniflora occupied most of the habitats of S. mariqueter and bare mudflat in the middle-eastern parts of CDW by the end of simulation period.

Modeled monthly range expansion of salt marsh vegetation at (a) CDW and (b) JW over the period of 2000–2015. The bars represent the variation over 10 repeated simulations. The upper annotation indicates the high-speed and low-speed invasion periods of S. alterniflora

Observed and modeled spatiotemporal dynamics of salt marsh vegetation at CDW (left panel) and JW (right panel) after plant invasion between 2000 and 2008
At the study site of JW, S. alterniflora spread in all directions from the initial location. By 2008, most of the habitat of S. mariqueter was displaced by S. alterniflora, with the exception of the western isolated shoal without initial S. alterniflora. The native plant P. australis expanded at a much slower rate than S. alterniflora.
Model Evaluation by Comparison of Simulated and Observed Salt Marsh Vegetation Dynamics
Besides the visual comparison with the salt marsh vegetation maps, the performance of the model was evaluated by comparing the simulated and observed distribution area for each species in the corresponding year of 2000–2008 (Fig. 8). On average through the 10 simulations, S. alterniflora expanded quickly from 33 ha (CDW) and 94 ha (JW) in 2000, to 916 ± 131 and 2,304 ± 244 ha in 2008. However, the distribution area of S. mariqueter decreased sharply from 1,660 (CDW) and 3,602 (JW) ha in 2000 to 963 ± 137 (on average by −41 %) and 1,263 ± 196 ha (on average by −65 %) in 2008. The increases in occupancy area of P. australis over the simulation period were 206 ± 25 and 277 ± 28 ha at CDW and JW, respectively.

Modeled range expansion of each salt marsh vegetation at CDW (a, c, e) and JW (b, d, f) over the period 2000–2008, plotted against the observed values. The bars represent the variation over 10 repeated simulations. The linear functions fitted to time series are shown for P. australis and S. mariqueter, and sigmoid functions for S. alterniflora with SPSS software (version 16.0, SPSS Ltd., USA)
The correlations between the modeled area of S. alterniflora against the corresponding measured values in 2004, 2006, and 2008 (Fig. 8) were high and statistically significant (R 2 = 0.97 for CDW and 0.95 for JW, p < 0.01). The simulated area of S. mariqueter explained the variation in measured area well (R 2 = 0.94 for CDW and 0.89 for JW, p < 0.01). As a comparison, the modeled area of P. australis had a similar extent to the measured area in 2004, 2006, and 2008 (R 2 = 0.92 for CDW and 0.93 for JW, p < 0.01).
The percentage difference in spatiotemporal dynamics listed in Table 4 reflected the comparison of the actual versus the modeled presence/absence of species on the distribution maps. The model underestimated the distributional area of S. mariqueter, with mean %DIFF figures of −18.9, −19.6, and −20.7 % for 2004, 2006, and 2008, respectively at CDW. The accuracy of modeling output was relatively higher at JW, with lower %DIFF figures (−14.3 to −18.2 %) at JW. The values for %DIFF of modeled range expansion of S. alterniflora at CDW were +9.5, +11.1, and +14.2 % for 2004, 2006, and 2008, respectively. However, the values of %DIFF were higher (+18.7 to +26.9 %) at JW, probably due to overestimation of expansion. For the distribution pattern of P. australis during the period 2000–2008, the mean values of %DIFF ranged from −12.6 to −20.1 % at CDW and −13.2 to −23.6 % at JW, respectively.
The Future Dynamics of the Salt Marsh Vegetation
In terms of predicted future changes within the spatiotemporal sedimentary regime, the model raises the mudflat elevation every year (Fig. 9a and b). The area of mudflat above 2.75 m increases by 106 ha at CDW and 144 ha at JW during the period 2000–2015. For mudflats above 2.9 m, the area increases by 94 ha at CDW and 152 ha at JW. The available habitat (above the elevation threshold) for S. alterniflora decreased sharply from 981 to 61 ha at CDW (Fig. 9c) and from 2,620 to 381 ha at JW (Fig. 9d) during the period 2000–2015. In comparison, the area of available habitat for P. australis did not decrease to the same extent.

Calculated increase in the area of mudflat habitat above 2.75 and 2.9 m based on the modeled spatiotemporal sedimentation regime (data refer to Fig. 1) at CDW (a) and JW (b), and available habitat for S. alterniflora and P. australis above the elevation threshold at CDW (c) and JW (d) during 2000–2015
As shown in Figs. 6 and 10, the model predicted the spatiotemporal dynamics of salt marsh vegetation in the Yangtze Estuary up to 2015. The results showed that the range expansion speed of S. alterniflora at the study sites slowed down, when compared with the period 2008–2009 with the mean distribution area of 965 ± 92 ha at CDW, representing an increase of only around 5 % during the period of 2008–2015. The expansion area of S. alterniflora at JW was predicted to increase by around 7 % (2,670 ± 228 ha) during the period of 2009–2015. Due to the decrease in the invasion rate of the exotic species, the distribution area of S. mariqueter is predicted to be relatively constant for the period 2008–2009 up to 2015 at both wetlands. The average distribution area of P. australis is increasing to 374 ± 43 and 1,051 ± 114 ha at CDW and JW in 2015, respectively.

The predicted spatial distribution of the saltmarsh vegetation at CDW (a) and JW (b) in 2015
Discussion and Conclusions
Outline of Model Output
The aim of this study was to use a grid-based modeling approach to recapture and predict the spatiotemporal dynamics of salt marsh vegetation after plant invasion on the coastal wetland habitats in the Yangtze Estuary. Based on long-term empirical data and observations on the salt marsh dynamics and field studies on the functional and bio-physical factors of the dominant species, the key ecological processes including the seasonal seed bank, the tidal current for seedling dispersal, establishment and reproduction, interspecific competition, sedimentary regime and inundation stress were incorporated into the model. Two important wetland locations of CDW and JW in the Yangtze Estuary were used to evaluate the model performance.
The simulated distribution matrix showed a good outcome, with a matching percentage of 73–91 % (refer to %DIFF) against the observed spatiotemporal dynamics of salt marsh vegetation at CDW and JW. Additionally, the accuracy of the distribution area for each salt marsh species was high (89–97 %), when compared to the observed values based on satellite images. Both simulation and observation suggested that the period of early salt marsh succession was crucial for the colonization and establishment of S. alterniflora after its introduction. However, the speed of expansion of the exotic species slowed down after 2006. This acceptable performance of the model was attributed to careful parameterization based on previous empirical field measurements of biotic and abiotic processes at the salt marsh (see Table 3). As presented in the results, the model is also capable of reproducing the general spatial dynamics of salt marsh vegetation at JW and CDW, and furthermore could be used elsewhere along the eastern seaboard of China where invasion of S. alterniflora is occurring.
The Importance of Species-Specific Parameterization
One of the advantages of the model is that the different species of salt marsh have specific parameters for simulation at a monthly time scale for successive annual years. The model assumed that the flood and ebb tide would transport the seedlings from an an initial colonizing cell to the neighboring area in all of eight directions in the cell matrix, which was in line with similar models applied elsewhere (Grevstad 2005). Over the whole time period of the simulation, the model identified a ‘long-distance’ seedling-dispersal pattern (for P. australis and S. alterniflora) during the early growing season due to tidal transportation and a clonal-integration pattern during the later growing season. The probability of seedling establishment on the new habitat, as a function of transportation distance seaward, was incorporated into the model. Moreover, the variations of dispersal distance of seedlings for the different species were parameterized in the model, based on continuous field measurements since 2007 (Xiao et al. 2009, 2010a, b; Zhu et al. 2012).
As a result, both the observed and simulated data indicated that the success of S. alterniflora could be largely attributed to the longer distance of its seedling dispersal and the higher survival rate of its propagules. In contrast, the soil seed banks of P. australis were small, compared to S. alterniflora, and few seeds of S. mariqueter were observed for all seasons on the seaward frontier mudflat where there are strong tidal currents (Xiao et al. 2009; Zhu et al. 2011). A review by DiTomaso and Healy (2006) reported that P. australis reproduces mainly through stolons and rhizomes in the field, and that persistent seed banks could not be produced under tidal conditions. Therefore, the transportation of living seedlings to new habitat by tidal current during the early growing season plays the most important role in population dispersal on the mudflats.
The previous reports from both the Chinese and American coasts indicated that P. australis and S. alterniflora could share similar niches (Chambers et al. 1998; Huang et al. 2008), and these two species may not replace each other once either of P. australis and S. alterniflora has colonized a new habitat. The experimental results of Wang et al. (2006) showed that the competitive balance between S. alterniflora and P. australis varied depending on the conditions of salinity and immersion, but also that these two species generally co-existed in the intertidal zones under typical conditions (oligohaline and half-immersion). However, S. alterniflora had strong competitive effects on S. mariqueter in the Yangtze Estuary (Li et al. 2009). S. mariqueter grows on the mudflats with elevation range of 1.5–3.5 m, and S. alterniflora can also grow well in the elevation range (Chen et al. 2004; Zhu et al. 2012). The survey on the Spartina–Scirpus neighboring transect showed that S. alterniflora was locally out-competing and displacing S. mariqueter on the tidelands (Chen et al. 2004). These species interactions have been successfully tested by setting P. australis and S. alterniflora as obstacles to each other while both being capable of invading the cells containing S. mariqueter and bare mudflat in the model.
In order to link potential and realized distributions of the salt marsh vegetation, species-specific tolerance to environmental stress and interspecific competition were programmed when developing the model. Generally, the duration of daily inundation is a limiting stress for the survival of salt marsh plants in the coastal habitat (Thompson 1991).
Deficiencies of Model Building
Although the model has performed well overall, there are still some weak points in terms of model structure and calibration, resulting in uncertainty of prediction and potential problems of practical application. The model overestimated (+9.5 to +14.2%DIFF) the spatial invasion ranges at CDW, as well as a further overestimation at JW. A probable explanation was that the data for model development and calibration were derived from measurements on sampling sites/strips located in the north-eastern part of CDW, where the expansion speed of S. alterniflora was relatively higher. When simulated at the landscape-scale, the cumulative effect appeared. Another limitation of the model was that, although this version of model set an elevation threshold for vegetation establishment, the hydrological factors of tidal flow energy and storm events were also important controlling those variables causing attrition of the seed banks and limiting plant survival (Schwarz et al. 2011; Zhu et al. 2012), and these factors were not included in the model.
The current model was calibrated using previous field observations and data from the analysis of satellite images (Chen et al. 2004; Wang et al. 2006; Huang and Zhang 2007; Li et al. 2009), assuming ‘complete’ replacement when one more competitive species encountered another. In the real world, however, the mechanisms of interspecific competition for habitat vary in relation to environmental stress, geomorphological conditions, resource availability, and species tolerances.
Another consideration is that few data are available on seed/seedling bank dynamics and germination rate for salt marsh species in different regions of the world with varying coastal hydrodynamic regimes. Therefore, more mechanistic relationships between hydrological conditions and vegetation establishment, as well as interspecies competition, need to be investigated for model in order to refine the model further. In tidal salt marshes, the feedback between vegetation establishment and hydrodynamic regimes and sedimentation is extremely important. In the Yangtze Delta, Li and Yang (2009) studied the trapping ability of salt marsh vegetation on suspended sediment, showing that salt marsh vegetation exhibited a positive feedback loop with sedimentation. Lambrinos and Bando (2008) also reported that once established, S. alterniflora attenuated hydrological energy significantly and enhanced sediment accretion, further resulting in higher seedling recruitment and wider range expansion. Although point-based mechanistic processes are understood, the model development approach at the landscape scale is currently not practical. Therefore, more available data regarding spatial heterogeneity of vegetation cover and density, quantitative differences in seedling dispersal rates with sediment accretion, and the effects of sediment deposition on plant growth are needed for model update.
Future Patterns and Conclusions
The continuous simulation within this model makes it possible to predict the future dynamics of salt marsh vegetation. The predictive simulation showed that the range expansion speed of S. alterniflora declined to a low-level during 2008–2015 at CDW and 2009–2015 at JW. In the Yangtze Estuary, S. mariqueter is the pioneer vegetation which creates new niches for S. alterniflora and P. australis to occupy. All range expansions of these species depend on the rate of accretion in the dynamic intertidal zone to provide the new niches. However, the current sedimentation rate may not continue the high-speed expansion of S. alterniflora that has occurred in the past decade, resulting in a slowing in the rate of expansion, due to less and less availability of habitat for establishment. The native S. mariqueter, which is more tolerant of inundation than other species, could continuously expand seaward. For another native species, the portion of P. australis settling in the southern part of CDW and the isolated shoal of JW far from S. alterniflora also could expand continuously on the high-elevation habitat.
Mechanistic models, similar to the one described here, that simulate how biotic and abiotic processes regulate the spatiotemporal dynamics of vegetation are increasingly being used for environment management (Grevstad 2005; Coutts et al. 2011; Emry et al. 2011; Fennell et al. 2012). As predicted by the model, the expansion speed of the exotic plant can be slowed down, due to feedback processes causing a reduction in the available habitat and limitations on inundation. This should be an opportune moment to introduce effective management protocols for the control and even the elimination of the exotic invasive species. Research into the developmental stages of S. alterniflora in relation to environmental stress on survival and inter-species competition has demonstrated that targeted measures could be introduced, for instance (1) removing or disabling the seeds/seedlings during the early growing season, (2) breaking and waterlogging of rhizomes at the period of tussock development, (3) cutting and replacing by native P. australis.
Increasingly, landscape-scale ecosystem models are becoming useful tools for scientific research, allowing future projections linked to environmental and climate change (Stralberg et al. 2011; Fagherazzi et al. 2012). Of the various models that have been developed to date, Sea Level Affecting Marshes Model (SLAMM) is a useful spatial model that simulates the dominant processes involved in wetland conversions and shoreline modifications during periods of long-term sea level rise (Church et al. 2001; Glick et al. 2013). The objectives of SLAMM focus on the impacts of environmental stresses, including inundation, erosion, overwash, salinity, and soil saturation under varying scenarios of sea level rises. Until now, similar types of models have not modeled critical local processes, such as lateral expansion of vegetation with horizontal habitat accretion and elevation change realistically (Stralberg et al. 2011). The combination of mechanistic models and spatial models will be needed to estimate feedback impacts of salt marsh dynamics on future environmental change more effectively.
In conclusion, the model developed in this study was able to reproduce realistically the spatiotemporal dynamics of salt marsh vegetation at both monthly and annual scales on the coastal habitats of the Yangtze Estuary after plant invasion. The long-term simulations of the model on these two different periods indicated the potential utility of the model, when appropriately parameterized, to the provision of information on the changes in the range pattern and expansion potential of invasive species and the consequent responses of native species. The model can be used by decision-makers with responsibility for coastal management and natural ecosystem maintenance to plan the strategic control of invasions at intermediate spatial and temporal scales
References
Chambers, R.M., T.J. Mozdzer, and J.C. Ambrose. 1998. Effects of salinity and sulfide on the distribution of Phragmites australis and Spartina alterniflora in a tidal salt marsh. Aquat Bot 62: 161–169.
Changjiang Water Resources Commission (CWRC), 2012. Changjiang Sediment Bulletin. Hubei: Changjiang Press, Wuhan, 40pp (in Chinese).
Chen, Zy., B. Li, Y. Zhong, and J.K. Chen. 2004. Local competitive effects of introduced Spartina alterniflora on Scirpus mariqueter at Dongtan of Chongming Island, the Yangtze River estuary and their potential ecological consequences. Hydrobiologia 528: 99–106. 2004.
Church, J.A., Gregory, J.M., Huybrechts, P., Kuhn, M., Lambeck, K., Nhuan, M.T., Qin, D., and Woodworth, P.L., 2001 Changes in sea level. In: Climate Change 2001: The Scientific Basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. (Eds.) Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M., van der Linden, P.J., Dai, X., Maskell, K., and Johnson, C.A.: Cambridge University Press, 881 pp.
Coutts, S.R., R.D. van Klinken, H. Yokomizo, and Y.M. Buckley. 2011. What are the key drivers of spread in invasive plants: dispersal, demography or landscape: and how can we use this knowledge to aid management? Biol Invasions 13: 1649–1661.
Davis, H.G., C.M. Taylor, and J.C. Civille. 2004. An Allee effect at the front of a plant invasion: Spartina in a Pacific estuary. J Ecol 92: 321–327.
DiTomaso, J.M., and Healy, E.A., 2006. Weeds of California and other Western States. Davis, CA: University of California Division of Agriculture and Natural Resources, Publication 3488.
Emry, D.J., H.M. Alexander, and M.K. Tourtellot. 2011. Modelling the local spread of invasive plants: importance of including spatial distribution and detectability in management plans. J Appl Ecol 48: 1391–1400.
Fagherazzi, S., M.L. Kirwan, S.M. Mudd, G.R. Guntenspergen, S. Temmerman, A. D’Alpaos, J. Koppel, J.M. Rybczyk, E. Reyes, and C. Craft. 2012. Numerical models of salt marsh evolution: ecological, geomorphic, and climatic factors. Rev Geophys 50, RG1002. doi:10.1029/2011RG000359.
Fennell, M., J.E. Murphy, C. Armstrong, T. Gallagher, and B. Osborne. 2012. Plant spread simulator: a model for simulating large-scale directed dispersal processes across heterogeneous environments. Ecol Model 230: 1–10.
Gedan, K.B., B.R. Silliman, and M.D. Bertness. 2009. Centuries of human-driven change in salt marsh ecosystems. Annual Review of Marine Science 1: 117–141.
Glick, P., J. Clough, A. Polaczyk, B. Couvillion, and B. Nunley. 2013. Potential effects of sea-level rise on coastal wetlands in southeastern Louisiana. J Coast Res 63: 211–233.
Grevstad, F.S. 2005. Simulating control strategies for a spatially structured weed invasion: Spartina alterniflora (Loisel) in Pacific Coast estuaries. Biol Invasions 7: 665–677.
Huang, H.M., and L.Q. Zhang. 2007. A study on the population dynamics of Spartina alterniflora at Jiuduansha Shoals Shanghai. Ecol Eng 29: 164–172.
Huang, H.M., L.Q. Zhang, and L. Yuan. 2007. The spatio-temporal dynamics of salt marsh vegetation for Chongming Dongtan National Nature Reserve, Shanghai. Acta Ecol Sin 27: 4166–4172 (in Chinese with English abstract).
Huang, H.M., L.Q. Zhang, Y.J. Guan, and D.H. Wang. 2008. A cellular automata model for population expansion of Spartina alterniflora at Jiuduansha Shoals, Shanghai, China. Estuar Coast Shelf Sci 77: 47–55.
Janine, I., A. Penttinen, H. Stoyan, and D. Stoyan. 2008. Statistical analysis and modelling of spatial point patterns. The Atrium, Southern Gate, Chichester: John Wiley & Sons Ltd.
Levin, L.A., C. Neira, and E.D. Grosholz. 2006. Invasive cordgrass modifies wetland trophic function. Ecology 87: 419–432.
Lambrinos, J.G., and K.J. Bando. 2008. Habitat modification inhibits conspecific seedling recruitment in populations of an invasive ecosystem engineer. Biol Invasions 10: 729–741.
Li, B., C.Z. Liao, X.D. Zhang, H.L. Chen, Q. Wang, Z.Y. Chen, X.J. Gan, J.H. Wu, B. Zhao, Z.J. Ma, X.L. Cheng, L.F. Jiang, and J.K. Chen. 2009. Spartina alterniflora invasions in the Yangtze River estuary, China: an overview of current status and ecosystem effects. Ecol Eng 35: 511–520.
Li, H., and S.L. Yang. 2009. Trapping effect of tidal marsh vegetation on suspended sediment, Yangtze Delta. J Coast Res 25: 915–924.
Li, H.P., L.Q. Zhang, and D.H. Wang. 2006. A study on the distribution of an exotic plant Spartina alterniflora in Shanghai. Biodivers Sci 14: 114–120 (in Chinese with English abstract).
Schwarz, C., T. Ysebaert, Z.C. Zhu, L.Q. Zhang, T.J. Bouma, and P.M.J. Herman. 2011. Abiotics governing the establishment and expansion of two contrasting salt marsh species in the Yangtze estuary, China. Wetlands 31: 1011–1021.
Sebert-Cuvillier, E., M. Simonet, V. Simon-Goyheneche, F. Paccaut, O. Goubet, and G. Decocq. 2010. PRUNUS: a spatially explicit demographic model to study plant invasions in stochastic, heterogeneous environments. Biol Invasions 12: 1183–1206.
Shi, B., J.Y. Ma, and K.Y. Wang. 2010. Effects of atmospheric elevated temperature on the growth, reproduction and biomass allocation of reclamation Phragmites australis in East Beach of Chongming Island. Resources and Environment in the Yangtze basin 19: 383–388 (in Chinese with English abstract).
Stralberg, D., M. Brennan, J.C. Callaway, J.K. Wood, L.M. Schile, D. Jongsomjit, M. Kelly, V.T. Parker, and S. Crooks. 2011. Evaluating tidal marsh sustainability in the face of sea-level rise: a hybrid modeling approach applied to San Francisco Bay. PloS one 6: e27388.
Thompson, J.D. 1991. The biology of an invasive plant—what makes Spartina anglica so successful? Bioscience 41: 393–401.
Wang, Q., C.H. Wang, B. Zhao, Z.J. Ma, Y.Q. Luo, J.K. Chen, and B. Li. 2006. Effects of growing conditions on the growth and interactions between salt marsh plants: implications for invasibility of habitats. Biol Invasions 8: 1547–1560.
Wang, R.Z., L. Yuan, and L.Q. Zhang. 2010. Impacts of Spartina alterniflora invasion on the benthic communities of salt marshes in the Yangtze Estuary, China. Ecol Eng 36: 799–806.
Weidenhamer, J.D., and R.M. Callaway. 2010. Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J Chem Ecol 36: 59–69.
Wolfram, S. 1986. Theory and application of cellular automata. Singapore: World Scientific.
Xiao, D.R., L.Q. Zhang, and Z.C. Zhu. 2009. A study on seed characteristics and seed bank of Spartina alterniflora at salt marshes in the Yangtze Estuary, China. Estuar Coast Shelf Sci 83: 105–110.
Xiao, D.R., L.Q. Zhang, and Z.C. Zhu. 2010a. The range expansion patterns of Spartina alterniflora on salt marshes in the Yangtze Estuary, China. Estuar Coast Shelf Sci 88: 99–104.
Xiao, Y., J.B. Tang, H. Qing, L. Gan, B.B. Tang, C.F. Zhou, and S.Q. An. 2010b. Effects of substrate, storage way and salinity on the seed germination of three coastal plants. Acta Ecol Sin 31: 1675–1683 (in Chinese with English abstract).
Yang, S.L. 1999. Sedimentation on a growing intertidal island in the Yangtze River mouth. Estuar Coast Shelf Sci 49: 401–410.
Zhang, L.Q., and X.K. Yong. 1992. Studies on phenology and spatial pattern of Scirpus mariqueter population. Acta Phytoecologica Sinica 16: 43–51 (in Chinese with English abstract).
Zhu, Z.C., L.Q. Zhang, N. Wang, C. Schwarz, and T. Ysebaert. 2012. Interactions between range expansion of salt marsh vegetation and hydrodynamic regimes in the Yangtze Estuary, China. Estuar Coast Shelf Sci 96: 273–279.
Zhu, Z.C., L.Q. Zhang, and D.R. Xiao. 2011. Seed production of Spartina alterniflora and its response of germination to temperature at Chongming Dongtan, Shanghai. Acta Ecol Sin 31: 1574–1581 (in Chinese with English abstract).
Acknowledgements
The authors would like to thank members of the Ecological Section of the State Key Laboratory of Estuarine and Coastal Research, East China Normal University, for their assistance with the collection of field data. We thank Professor Martin Kent (Plymouth University, UK) for kindly checking the English in the revision. This work was funded through the Natural Science Foundation of China (41201091), the Shanghai Pujiang Program (13PJ1402200) and Social-development project (12230707500) of Shanghai Science & Technology Committee, the Global Change Scientific Research Program of China (2010CB951204), and the Project-sponsored by SRF for ROCS, SEM.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Linda K. Blum
Rights and permissions
About this article

Cite this article
Ge, ZM., Zhang, LQ. & Yuan, L. Spatiotemporal Dynamics of Salt Marsh Vegetation regulated by Plant Invasion and Abiotic Processes in the Yangtze Estuary: Observations with a Modeling Approach. Estuaries and Coasts 38, 310–324 (2015). https://doi.org/10.1007/s12237-014-9804-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-014-9804-7