
Abstract
Potato plants are often exposed to soil drought, which results in crop losses, because potato root system is shallow. The aim of our study was to examine the relationship between root system architecture and drought tolerance. We measured rooting depth, total length, surface area, average diameter, and dry weight of the root system every 20 cm of soil depth. We examined four potato cultivars, two that were more tolerant to drought (Gwiazda and Tajfun), and two that were more sensitive (Oberon and Cekin). Our results showed different responses by the tested cultivars to drought. The root dry mass decreased in response to drought and this decrease was smaller in the more drought-tolerant cultivars. In response to drought, the more drought-tolerant cultivars developed elongated roots, but the roots of the less-tolerant cultivars remained the same length. The decrease in root diameter was also smaller in the more drought-tolerant cultivars.
Resumen
Las plantas de papa a menudo están expuestas a sequía, lo que resulta en pérdidas del cultivo, porque el sistema radical de la papa es superficial. La meta de nuestro estudio fue examinar la relación entre la arquitectura del sistema radical y la tolerancia a la sequía. Medimos la profundidad de la raíz, la longitud total, el área de la superficie, el promedio del diámetro y el peso seco del sistema radical a cada 20 cm de profundidad del suelo. Examinamos cuatro variedades de papa, dos que fueran mas tolerantes a la sequía (Gwiazda y Tajfun), y dos que fueran mas sensibles (Oberon y Cekin). Nuestros resultados mostraron diferentes respuestas de las variedades probadas a la sequía. La materia seca de la raíz disminuyó en respuesta a la sequía, y esta disminución fue menor en las variedades mas tolerantes. En respuesta a la sequía, las variedades mas tolerantes desarrollaron raíces alargadas, pero las raíces de las menos tolerantes permanecieron de la misma longitud. La disminución en el diámetro de la raíz también fue menor en las variedades mas tolerantes a la sequía.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Under global climate change, drought tolerance will likely become increasingly important for producing stable yields in all crops (Comas et al. 2013; Hijmans 2003). The root system plays a major role in drought tolerance, but roots are studied less than above-ground plant parts, due to difficulty of observing them (Dathe et al. 2014, Lynch 1995). Drought can cause substantial yield losses (Sołtys-Kalina et al. 2016), particularly for crops with shallow root systems such as potato (Solanum tuberosum). The potato root system is shallow, with weak soil penetration (Joshi et al. 2016, Stalhman et al. 2007), and is considered inefficient compared to those of other crops. However, potato roots can reach depths of about 100 cm and total root lengths throughout the soil profile of about 10–20 km, yet much of potato root system is concentrated up to 30 cm in soil depth. However, many studies have found large differences in the size of the root system between cultivars and in response to different environmental factors (Głuska 2004; Iwama 2008; Joshi et al. 2016; Rykaczewska 2015; Zarzyńska et al. 2017). Under the same environmental conditions and with similar agronomic factors, the differences in the root system mainly involve the maturity of the plants. Cultivars with long-lived vegetation continue to grow roots longer, generally reaching greater weight and deeper rooting than the earlier varieties (Kashiwagi et al. 2000).
Differences in root mass also exist amongst genotypes of the same maturity class (Opena and Porter 1999, Rykaczewska 2015, Rykaczewska et al. 2018, Stalhman and Allen 2001, Zarzyńska et al. 2017). Breeding new cultivars with improved root characteristics that can absorb water from deeper in the soil and under conditions of lower soil water potential will increase the usage of soil water (stored or irrigated), and thus contribute to more efficient utilization of water for potato production. Much research has attempted to find a correlation between the size of the root system, tolerance to water shortage, and tuber yield (Comas et al. 2013, Iwama et al. 1999; Khan et al. 2016, Lahlou and Ledent 2005, Villordon et al. 2014); however, the correlations were not always positive. In the current study, we evaluated the changes in root system size and architecture in response to soil drought in multiple potato cultivars that are known to differ in their reactions to drought.
Materials and Methods
Plant Material and Climatic Conditions
The study was carried out on four potato cultivars during two vegetation seasons between 2015 and 2016 in the Potato Agronomy Department at the Plant Breeding and Acclimatization Institute-National Research Institute. The Gwiazda and Oberon cultivars were provided by the Zamarte Breeding Station, and the Tajfun and Cekin cultivars were provided by the Pomeranian-Mazurian Breeding Station. Gwiazda is an early genotype and Oberon, Tajfun, and Cekin are middle-early genotypes. Potato cultivars were selected based on their levels of drought tolerance based on maintenance of yield shown in field trials (Nowacki 2012) and pot experiments (Boguszewska-Mańkowska et al. 2018, Pieczynski et al. 2018). These experiments showed that Gwiazda and Tajfun are drought tolerant, since they maintain high yields during soil drought, whereas Oberon and Cekin are drought susceptible cultivars.
Pot Experiment (Experiment I)
The pot experiment was conducted in a vegetation hall from 2015 to 2016 on four potato cultivars (Gwaizda Oberon, Cekin and Tajfun). Plants were grown in 14-L pots filled with a thin layer of gravel in the bottom and 12 L of the universal vegetable soil substrate ‘Hollas’ produced from peat with the addition of chalk at a pH range of 5.5–6.5. For improved soil aeration, a gum pipe was installed in each pot. Six pots were placed on mobile platforms (1.85 × 0.80 m) outdoors in an open space (4 plants per m2) and were rearranged daily to avoid border effects. In the case of rain, the mobile platforms were sheltered under a glass roof to keep the soil and plants dry. During the growing season, plants were fertilized once with (MIS-3) Intermag with a dose of 10 g per plant. Pest and disease control was carried out 3 times against Colorado beetles and 4 times against Phytophthora infestans. To obtain optimal water condition, that is 70% of water capacity, plants were watered daily by drip irrigation. The water field capacity was measured by soil moisture tester (PAT.P. Nieuwkoop B.V. Aalsmer Holland). Two weeks after the initiation of the tuberisation drought stress condition was applied: drought stress without any irrigation for two weeks and the control with optimal irrigation (70% of water capacity). Six pots of plants were used for each treatment (control condition and drought condition) in each growing seasons (n = 12). After the dry period, the plants were watered, and the optimal water supply (70% of water capacity) was reinstated until the end of the experiment. After the end of the growing season, tuber yield (kg) was measured.
Root Experiment (Experiment II)
The study was conducted in 2015–2016 in specially constructed cylindrical pots with open bottoms (Fig. 1) on four potato cultivars (Gwaizda Oberon, Cekin and Tajfun). Pots were made from sheets of galvanized steel with a thickness of 0.8 mm and were 1 m tall and 40 cm wide to allow for proper development of the above-ground and below-ground parts of the plants. The pots could be opened along the seam of the cylinder, allowing extraction of the entire root system without damage (Głuska 1996, with modifications).

Cylindrical pots used for root experiments (left) and potato plants growing under drought conditions in the specially constructed shelter (right)
Pots were filled with a light, loamy soil brought in from a field where potatoes were grown and mixed with sand in a 1:1 ratio. A metal net with a mesh size of approx. 0.5 cm was placed every 20 cm, dividing the pot into 5 horizontal layers, each 20-cm thick. Seed potato tubers were placed approximately 5 cm below the surface of the soil. The mesh discs do not impede the growth of roots, and allowed the roots from growing plants to pass into the soil profile. During the growing season, plants were fertilized twice with Yara Mila Viking NPK 14–14-21 with a dose of N = 2.1, P = 1.0, K = 2.4, and Mg = 0.4 g per plant. Pest and disease control was carried out 3 times against Colorado beetles and 4 times against Phytophthora infestans.
To obtain optimal water condition, that is 70% of water capacity, plants were watered daily by drip irrigation. Two weeks after the initiation of the tuberisation drought stress condition was applied: drought stress without any irrigation for two weeks and the control with optimal irrigation (70% of water capacity). Three pots of plants were used for each treatment (control condition and drought condition) in each growing seasons (n = 6). After the dry period finished that is in full vegetation (blooming) selected root system parameters were measured. It is when potato plants display the largest differences in the size of their root systems (Iwama 2008).
Root Sampling and Measurements (Experiment II)
The root system was removed from the pots by opening the cylinders. The roots were cut across the net separating the individual layers and then washed away with a strong stream of water to produce five root sections from each plant for measurements.
For all cultivars (Gwiazda, Oberon, Tajfun, and Cekin) the total dry weight of the root system (g) and dry weight of roots in the individual layers (g) were also assessed. With this, the ratio of shoot to root dry mass (g), and the proportion of roots in total plant biomass (%) was calculated. Total plant biomass includes the whole plant (shoots and roots dry mass). Roots and shoots were dried in two stages: 24 h drying at 75 °C and then at 105 °C until they reached a constant weight.
For the Gwiazda and Oberon cultivars, extra measurements were made: rooting depth (cm), total root length (m), root surface area (m2), root average diameter (mm), root length in the individual layers (m), root surface area in the individual layers (m2), and root diameter in the individual layers (mm). The length, diameter, and surface area of the roots were measured using an Epson Expression scanner 10000XL (Seiko Epson Corp., Japan) and the WinRhizo software (Regent Instruments, Canada).
Relative Water Content
First and second mature and fully expanded leaves next to terminal leaflet from the third level of compound leaf counting from the top of the plant, comparable in size, were sampled to assess relative water content (RWC). Leaves were cut from the plant, weighed immediately (fresh weight, FW), floated in dark for 24 h to achieve turgidity (saturated weight, SW), then oven-dried (105 °C) for 24 h and weighed again (dry weight, DW). RWC of leaves was calculated according to the formula: [(fresh weight − dry weight)/(saturated weight − dry weight)] × 100% (Boguszewska et al. 2010).
Statistical analyses of the results were performed with an analysis of variance using Statistica software (StatSoft, Poland). The significance of the sources of variation was tested with a Fisher-Snedecor test, and the significance of differences was assessed by Tukey’s test.
Results
Water Availability Had a Significant Effect on Plant Parameters
An analysis of variance showed different responses by the tested cultivars to drought, as pointed by variations in multiple biometric characteristics of plant growth. Different water availabilities significantly affected all parameters too. The layer of soil that was sampled had a significant influence on all tested parameters. Years of the experiments significantly influenced only tuber yield. Significant interactions between cultivar and treatment (control and drought conditions) were observed for yield, RWC, shoot and root dry mass (Table 1).
Tuber Yield in Relation to Water Availability
The tuber yield depended on cultivar and treatment, and a significant interaction between cultivar and treatment was shown (Table 1). The average yield in the control treatment amounted to 1347 g per plant while plants from drought conditions had yields of 994 g per plant. All investigated cultivars displayed significant decrease of tuber yield under drought stress. The highest yield in the drought treatment (averaged over two growing seasons) was observed in Gwiazda (1248 g) and Tajfun (1075 g) cultivars, and the lowest was observed in Cekin (788 g) and Oberon (866 g) cultivars. (Fig. 2). In the year 2016 the average yield was significantly higher than in 2015.

Tuber yield (in g per plant) under optimal water availability (control) and two-week soil drought
The Changes of Shoot Mass in Response to Drought
In control conditions the water content (RWC) in leaves of investigated potato cultivars was similar. In cultivars which were stressed by withholding water for 21 days RWC diminished considerably to the level of about 41–55%. The lowest decrease was observed in cultivars Tajfun and Gwiazda and the highest in Oberon and Cekin (Fig. 3).

Relative water content (RWC in %) of the third mature leaves (costing from the plant top) from the control (optimal water availability) and drought potato plants
The cultivar significantly affected shoot dry mass. All investigated cultivars displayed a significant decrease in shoot dry mass in response to soil drought (Table 2).
The Dry Mass of Root System Decreased in Response to Drought
Both the cultivar and water availability significantly affected root dry mass. Root dry mass was also affected by the combination of these factors (Table 1). Among the four cultivars, Gwiazda had the lowest root mass and Tajfun had the highest. All tested cultivars displayed a decrease in dry root mass in response to drought treatment. Significant decrease in root dry mass was observed in drought susceptible cultivars to soil drought (Oberon and Cekin) (Table 2).
The ratio of shoot to root dry mass significantly differed between soil moisture conditions (Table 1). Under drought stress, this average ratio was lower (3.44) than in control conditions (4.57). In control conditions Gwiazda cultivar had significantly higher shoot to root ratio than rest of cultivars. Under drought conditions shoot to root dry mass was significant lower in all cultivars except from drought tolerant cultivar Tajfun (Table 2).
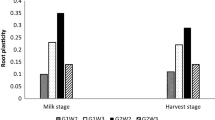
The opposite relationship was observed between the proportion of roots in total plant biomass. The lowest share of roots in total plant biomass had cultivar Gwiazda (16.5%) and the highest Tajfun (19.5%) cultivar. Under drought stress this parameter significantly increased in all tested cultivars. The highest share of roots in total biomass had susceptible cultivars Oberon (23.5%) and Cekin (23.9%) (Table 2).
We also observed differences in the root dry mass among the various soil profile layers. In both the control and drought stress conditions, root mass was largest in the first soil layer (0–20 cm), and gradually decreased with each successive soil layer. The root dry matter decreased in response to drought in each individual soil layer, yet the highest decrease was observed in the deepest soil layer (80–100 cm) (Table 3).
Changes in Length, Area, and Diameter of Roots in Response to Drought Stress
Additional measurements were made for the two cultivars Gwiazda and Oberon: total root length, root length in each soil layer, total area of the root system, surface area of the roots in each soil layer, average diameter of roots, and diameter of the roots in each individual soil layer. The cultivars differed significantly in all tested parameters (Table 1). The Gwiazda cultivar had a shorter root system than the Oberon cultivar, and each cultivar reacted to drought differently. Under drought stress, the Gwiazda cultivar reacted by increasing the length of its roots, whereas the Oberon cultivar maintained the same root length even under drought stress (Fig. 4a). Root surface area in the drought-resistant cultivar Gwiazda was not affected by drought conditions; however, this parameter decreased significantly in response to drought in the drought-susceptible cultivar, Oberon (Fig. 4b). Under drought conditions, both cultivars displayed a statistically significant decrease in average root diameter, but in the drought-susceptible cultivar Oberon, the decrease was twice as large as in the drought-tolerant cultivar Gwiazda (Fig. 4c).

Total root length (A), root area (B), and average root diameter (C) of Gwiazda (drought tolerant) and Oberon (drought-sensitive) cultivars under optimal water availability and two-week soil drought at tuberization phase. Values followed by the same letters (a, b, c) are not significantly different at p ≤ 0.05 (Tukey’s HSD test)
Changes in Root Parameters among Soil Layers
The root length was highest in the second and third soil layers (40–60 cm) and then decreased in each subsequent deeper soil layer in both the control and drought conditions. This decrease in root length was smaller in the Gwiazda cultivar than in the Oberon cultivar (Fig. 5). The decrease in root diameter was more dramatic in the Oberon cultivar than in the Gwiazda cultivar (Fig. 5).

Changes in the root length (A), root surface area (B), and root diameter (C) in individual soil layers of Gwiazda (drought tolerant) and Oberon (drought-sensitive) cultivars under optimal water availability and two-week soil drought at tuberization phase. Mean values and standard errors are presented
Discussion
Observation of root systems under field conditions is tedious and requires a lot of time and labor which is often emphasized in the literature (Smith et al. 1994; Tracy et al. 2011; Silberbush 2013; Khan et al. 2016). Due to the detail of measurements, our research was carried out in controlled conditions. Relationships obtained in pot experiments are most often confirmed under field conditions and can be a model for finding general dependencies (Wishard et al. 2013).
Root length and root mass are the most commonly examined root characteristics in agronomic and physiological studies. Some studies on potatoes reported that tuber production is related to root mass, rather than root length (Gregory et al. 1997, Lahlou and Ledent 2005). However, other studies show that water uptake by the roots is related to root length, rather than root mass (Blum 2005; MacKerron 2007). In this study, we investigated the relationship between drought stress, and root size and architecture, as well as above-ground plant mass. Among four cultivars, Gwiazda and Tajfun are recognized as more tolerant to drought, while Cekin and Oberon are more sensitive to drought. We found that all four cultivars reacted to drought by decreasing their root masses, but this decrease was more pronounced in the drought-sensitive cultivars. Similar results have been previously reported: Gregory et al. (1997), and Lahlou and Ledent (2005) both show a reduction in root dry mass in response to drought. We also found changes in the abundance of root dry matter in individual soil layers in response to drought treatment. Interestingly, the highest differences in the abundance of root dry matter were observed in the deepest soil layers (80–100 cm). Several studies have shown that potato cultivars differ in the size of the entire root system, and in distribution of the roots among the soil profile (Leszczynski and Tanner 1976; Opena and Porter 1999; Parker et al. 1989; Stalhman and Allen 2001; Vos and Groenwold 1986; Zarzyńska et al. 2017). Across all four cultivars and conditions (soil drought or optimal water), we observed an average root dry mass of 5.99 g per plant. Opena and Porter (1999) observed root dry mass of 4.77 and 5.38 g per plant at 63 and 77 days after planting, respectively. Sharifi et al. (2005) obtained maximum root dry mass of 18.5 and 22.0 g per plant at 76 days after planting.
Many studies found a positive relationship between the size of the root system and the amount of above-ground biomass (Gregory et al. 1997; Blum 2005; Iwama 2008; Puértolas et al. 2014, Rykaczewska 2015, Rykaczewska et al. 2018). In our experiment only cultivar Gwiazda had similar relationship.
The tested cultivars reacted in the decrease of shoot mass under the drought. It should be noted that the cultivars reacted with high decrease of the above-ground part also had high decrease in root mass. This observation can be an indicator of the response of cultivars on root system changes to water scarcity. This requires however confirmation in further studies.
In our previous work (Zarzyńska et al. 2017) we mainly evaluated cultivar differences in the size of the root system and we tried to find dependence between its size and drought tolerance. In our current research, we focused on the changes of the above and under-ground part of plant under soil drought conditions. Our research showed that the higher variation concerned the above-ground parts of plants rather than roots. The decrease in shoot mass under the drought amounted to 34.4% and the root mass decreased only by 14.3%. As a result of such changes the ratio of shoot mass to root mass was higher in control conditions than in drought. The reverse situation was related to the share of roots in the total plant biomass. Such dependence was confirmed by work of Dathe et al. (2014) which showed that the root-to-shoot ratio increases as irrigation decreases. In drought conditions, the ratio of above-ground dry mass to root dry mass decreased in all cultivars that we tested.
Despite the fact that all cultivars reacted with a decrease in the shoot to root index and the increase in the share of roots in the total plant biomass, in each case, smaller changes concerned more resistant cultivars.
Significant attempts have been made to identify a correlation between the size of a root system and the size of the crop, however, a simple relationship remains elusive. Ahmadi et al. (2017) and Iwama (2008) show that the root mass is generally positively correlated with tuber yield. Lahlou and Ledent (2005) found that fresh tuber yield is significantly correlated with root dry mass in field conditions. However, we were unable to confirm such a simple relationship.
The main purpose of this study was to assess the impact of water shortages on root systems in various potato cultivars. We found that both drought-tolerant cultivars, Gwiazda and Tajfun, displayed a smaller decrease in root mass under drought conditions than the drought sensitive cultivars, Cekin and Oberon. Some authors identified simple relationships between root size and drought resistance. Rykaczewska (2015) found that the drought-tolerant Tetyda cultivar has the largest root system mass of the 17 tested cultivars under aeroponic conditions. Also, Lahlou and Ledent (2005) showed that the drought tolerance index is significantly associated with root depth, but only when the plant is under stress, and Zarzyńska et al. (2017) showed a similar relationship.
On the basis of current research, however, it can not be unambiguously confirmed. Variety, which deviates from this principle is Gwiazda which has a small root system but belongs to resistant cultivars. This cultivar used different strategies, concerning physiological traits, to cope with soil drought condition comparing to drought resistance Tajfun cultivar (Boguszewska-Mańkowska et al. 2018). It should be noted, however, that the changes of shoot as well as roots mass under the drought were lower in both drought resistance cultivars.
In this study, we also observed an interesting relationship between the length and surface area of roots. The drought-sensitive Oberon cultivar had longer roots than the drought-tolerant Gwiazda. However, other research has found that increased root production in potatoes is associated with improved drought tolerance (Ahmadi et al. 2011, MacKerron 2007, Puértolas et al. 2014). Interestingly, the Gwiazda cultivar reacted to the stress by elongating its roots, yet the Oberon cultivar roots remained the same length.
The changes in root diameter of these cultivars in response to drought was also different. Although both cultivars reacted to drought stress by decreasing the diameter of their roots, this decrease was more pronounced in the Oberon cultivar than in the Gwiazda cultivar. This trend was also observed in individual soil layers. Wishart et al. (2014) demonstrated that root parameters that are associated with drought tolerance in potato, such as those identified in our study, can be used to select genotypes with resilience to drought.
The reactions of root systems to drought stress are complex. According to our results, the dry mass of potato roots decreased in response to drought, and this decrease was smaller in more tolerant cultivars. More resistant cultivars reacted to drought by elongating their roots, while the roots of less tolerant cultivars remained the same length, and decreases in root diameter were smaller in the more drought resistant cultivar.
On the basis on our experiment we can also conclude that the changes in plant morphology under drought stress are lower in tolerant cultivars than in sensitive. We can also say that smaller changes of above-ground mass means smaller changes in root mass. These information could be important for breeders.
References
Ahmadi, S.H., F. Plauborg, M.N. Andersen, A.R. Sepaskhah, C.R. Jensen, and S. Hansen. 2011. Effects of irrigation strategies and soils on field grown potatoes: Root distribution. Agricultural Water Management 98: 1280–1290.
Ahmadi, S.H., M. Agharezaee, A. Kamgar-Haghighi, and Sepeskhah. 2017. Compatibility of root growth and tuber production of potato cultivars with dynamic and static water saving irrigation managements. Soil Use and Manag. 33: 106–119.
Blum, A. 2005. Drought resistance, water-use efficiency, and yield potential—Are they compatible, dissonant, or mutually exclusive? Australian J Agricult. Res. 56: 1159–1168.
Boguszewska, D., M. Grudkowska, and B. Zagdanska. 2010. Drought responsive antioxidant enzymes in potato (Solanum tuberosum L.). Pot. Res. 53: 373–382.
Boguszewska-Mańkowska, D., M. Pieczyński, A. Wyrzykowska, H.M. Kalaji, L. Sieczko, Z. Szweykowska-Kulińska, B. Zagdańska. 2018. Divergent strategies displayed by potato (Solanum tuberosum L.) cultivars to cope with soil drought. J Agron. And crop Sci. 204:13–30.
Comas, L.H., V. Becker, P.F. Byrne, and D.A. Dierig. 2013. Root trials contributing to plant productivity under drought. Frontiers in Plant Science 4: 442.
Dathe, A., D.H. Fleisher, D.J. Timlin, J.K. Fisher, and V.R. Reddy. 2014. Modeling potato root growth and water uptake under water stress conditions. Agricult. and Forest Meteorol. 194: 37–49.
Głuska, A. 1996. Konstrukcja dużego wazonu cylindrycznego do badań systemu korzeniowego ziemniaka. Acta Agrobotanica 54: 161–170.
Głuska, A. 2004. Differentiation of root system size in potato cultivars. Biul. IHAR. 232: 37–46.
Gregory, P.J., J.A. Palta and G.R. Batts. 1997. Root systems and root mass ratio - carbon allocation under current and projected atmospheric conditions in arable crops. Plant and Soil. 187.
Hijmans, R.J. 2003. The effect of climate change on global potato production. Am J Pot. Res 80: 271–280.
Iwama, K. 2008. Physiology of the potato. New insights into root systems and repercussions for crop managements. Pot. Res. 51: 333–353.
Iwama, K., T. Hasegawa and K. Nakaseko. 1999. New potato lines with high productivity and drought tolerance. Proc.Intern. Sym. W.F.S.Kyoto 189-193.
Joshi, M., E. Fogelman, E. Belausov, and I. Ginzberg. 2016. Potato root system development and factors that determine its architecture. Journal of Plant Physiology 205: 113–123.
Kashiwagi, J., K. Iwama, T. Hasegawa, T. Nagata. 2000. The relationship between the root mass in seedlings and tuber generation in potato. Abstr. Of 12 th Symp. Of ISTRC. Tsukuba, Japan 446-450.
Khan, M.A., D.C. Gemenet, and A. Villordon. 2016. Root system architecture and abiotic stress tolerance: Current knowledge in root and tuber crops. Front. in Plant Sci. 7: 1584.
Lahlou, O., and J.F. Ledent. 2005. Root mass and depth, stolons and root formed on stolons in four cultivars of potato under water stress. Europ. J.Agron. 22: 159–173.
Leszczynski, D.B., and C.B. Tanner. 1976. Seasonal variation of root distribution of irrigated, field-grown russet Burbank potato. Am. Potato J: 69–78.
Lynch, J. 1995. Root architecture and plant productivity. Plant Physiology 109: 7–13.
MacKerron, D.K. 2007. Mathematical models of plant growth and development. In Potato biology and biotechnology: Advances and perspectives (eds D. Vreugdenhil, et al.), 753–776. Oxford, UK: Elsevier Science.
Nowacki, W. (Ed.). 2012. Characteristics of national register of potato cultivars (in Polish). ISBN: 83-891172-55-0.
Opena, G.B., and G.A. Porter. 1999. Soil management and supplemental irrigation effect on potato: II root growth. Agronomy Journal 91: 426–431.
Parker, C.J., M.K.V. Carr, N.J. Jarris, M.T.B. Evans, and V.H. Lee. 1989. Effect of subsoil loosening and irrigation on soil physical properties, root distribution and water uptake of potatoes (Solanum tuberosum). Soil and Tillage Research 13: 267–285.
Pieczynski, M., A. Wyrzykowska, K. Milanowska, D. Boguszewska-Mańkowska, B. Zagdańska, W. Karlowski, A. Jarmolowski, and Z. Szweykowska-Kulińska. 2018. Genome-wide identification of genes involved in the potato response to drought indicates functional evolutionary conservation with Arabidopsis plants. Plant Biotech. J. 16: 603–614.
Puértolas, J., C. Ballester, E.D. Elphinstone, and I.C. Dodd. 2014. Two potato (Solanum tuberosum) varieties differ in drought tolerance due to differences in root growth at depth. Functional Plant Biology 41: 1107–1118.
Rykaczewska, K. 2015. Morphology and anatomy of the root system of new potato cultivars. Part I. Morphology of the root system. Biul. IHAR 275: 99–109.
Rykaczewska, K., K. Zarzyńska, and D. Boguszewska-Mańkowska. 2018. Architecture of the root system of potato cultivars grown in aeroponics. Electr. J Polish Agricult. Univers. 21 (1).
Sharifi, M., B.J. Zebarth, M.A. Hajabbasi, and M. Kalbasi. 2005. Dry matter and nitrogen accumulation and root morphological characteristic of two clonal selection of russet Norkotah potato as affected by nitrogen fertilization. Journal of Plant Nutrition 28: 2243–2253.
Silberbush, M. 2013. Root study: Why is it behind other plant studies? Am. J. Plant Science 4: 198–203.
Smith, A.L., J. Groenwold, and J. Vos. 1994. The Wageningen rhizolab – A facility to study soil-root-shoot-atmosphere interactions in crops. II. Methods of root observations. Plant and Soil 161: 289–298.
Sołtys-Kalina, D., J. Plich, D. Strzelczyk-Zyta, J. Śliwka, and W. Marczewski. 2016. The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of ‘Katahdin’-derived potato cultivars. Breeding Science 66: 1–4.
Stalhman, M.A., and E.J. Allen. 2001. Effect of variety, irrigation regime and planting date on depth, rate duration and density of root growth in the potato (Solanum tuberosum) crop. J.Agric. Sci 137: 251–270.
Stalhman, M.A., E.J. Allen, A.B. Rosenfeld, and F.X. Herry. 2007. Effect of soil compaction in potato (Solanum tuberosum L). The Journal of Agricultural Science 145: 295–312.
Tracy, S.R., C.R. Black, J.A. Roberts, and S.J. Mooney. 2011. Soil compaction: A review of past and present techniques for in vestigating effects on root growth. Journal of the Science of Food and Agriculture. 91: 1528–1537.
Villordon, A.Q., J. Giuzberg, and N. Firon. 2014. Root architecture and root tuber crop productivity. Trends in Plant Sci. 19: 419–425.
Vos, J., and J. Groenwold. 1986. Root growth of potato crops on a marine –clay soil. Plant and Soil 161: 289–298.
Wishard, J., T.S. George, L.K. Brown, G. Ramsay, J.E. Bradshaw, P.J. White, and P.J. Gregory. 2013. Measuring variation in potato roots in both field and glasshouse: The research for useful yield predictors and simple screen for roots trials. Plant and Soil 368: 231–249.
Wishart, J., T.S. George, L.K. Brown, P.J. White, G. Ramsay, H. Jones, and P.J. Gregory. 2014. Filed phenotyping of potato to assess root and shoot characteristics associated with drought tolerance. Plant and Soil 378: 531–363.
Zarzyńska, K., D. Boguszewska-Mańkowska, and A. Nosalewicz. 2017. Differences in size and architecture of the potato cultivars root system and their tolerance to drought stress. Plant Soil and Envin. 63: 159–164.
Acknowledgments
This study was supported by grants awarded by the Ministry of Agriculture, Poland (MRiRW: 59:4-3-00-3-02), and Dzialalnosc Statutowa (1-3-00-2-01) of the Potato Agronomy Department, Plant Breeding and Acclimatization Institute-NRI, Division Jadwisin, Poland.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Boguszewska-Mańkowska, D., Zarzyńska, K. & Nosalewicz, A. Drought Differentially Affects Root System Size and Architecture of Potato Cultivars with Differing Drought Tolerance. Am. J. Potato Res. 97, 54–62 (2020). https://doi.org/10.1007/s12230-019-09755-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12230-019-09755-2