
Abstract
Droughts are common in the US High Plains, causing declining water availability and lowering water allocations. The objective of this 4-year field study was to identify a period of water deficit least detrimental to potato production. Fully irrigated ‘Atlantic’ potato received 62–63 cm of applied water. Total applied water was reduced by 25 % in three seasonal periods, 50 % water from emergence to 8 weeks after emergence (WAE) (early stress), 50 % water from 8 to 13 WAE (late stress), and 50 % from 0 to 5 WAE and again from 10 to 13 WAE (outer stress). Main plots were irrigation regime and split with three N levels, 101, 168, and 235 kg/ha. Soil and petiole N were higher when applied water was reduced. Lower irrigation inhibited growth, i.e., canopy height (10–20 %), weight (20–30 %) and leaf area index (50–70 %). Yield decreased 25 % and 13 % with early and outer stress, respectively. Chip color was darkest with early stress compared to fully irrigated plots. Common scab occurrence was greater in early stress than with other regimes. N rate had no effect on canopy growth, yield, chip color, or common scab. If applied water is reduced 15 cm, it is best late in the season and worst between 5 and 8 WAE compared to fully irrigated plants
Resumen
Las sequías son comunes en las planicies altas de EUA, causando disminución en la disponibilidad de agua y bajando su ubicación. El objetivo de este estudio de campo de cuatro años fue identificar un período de déficit de agua que fuera lo menos perjudicial para la producción de papa. La papa Atlantic con riego completo recibió de 62 a 63 cm de agua. El total del agua aplicada se redujo en un 25 % en tres ciclos, 50 % de agua de la emergencia a 8 semanas después de la emergencia (WAE) (agobio temprano), 50 % de agua de 8 a13 WAE (agobio tardío), y 50 % de 0 a 5 WAE y otra vez de 10 a 13 WAE (agobio externo). Los lotes principales tuvieron regímenes de riego y fueron divididos con tres niveles de N, 101, 168, y 235 kg/ha. El N del suelo y del pecíolo fue más alto cuando se redujo el agua aplicada. Los riegos más bajos inhibieron crecimiento, i.e., altura del follaje (10–20 %), peso (20–30 %) y el índice de área foliar (50–70 %). El rendimiento disminuyó 25 % y 13 % con agobio temprano y externo, respectivamente. El color de la hojuela fue el más oscuro con agobio temprano comparado con los lotes de riego completo. La incidencia de la roña común fue mayor en agobio temprano que con los otros regímenes. El nivel de N no tuvo efecto en el crecimiento del follaje, rendimiento, color de la hojuela, o roña común. Si el agua aplicada se reduce 15 cm, es mejor tarde en el ciclo y peor entre 5 y 8 WAE en comparación con plantas de riego completo.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Droughts are common in the High Plains causing declining water availability for agriculture and decreasing irrigation allocations. Legislation to manage groundwater in Nebraska, Kansas and Colorado is decreasing water allocations available for irrigated agriculture. For growers in the Southwest and Panhandle of Nebraska, it simply means less water and greater risk for yield and market quality. Under limited irrigation, less water is applied than is required to meet full evapo-transpiration demand. As a result, the crop will be stressed.
In a project, conducted at North Platte, NE, three irrigation treatments (dryland, 15 cm of irrigation and non-evapotranspitation limiting) were applied to corn, soybean, and wheat (Hergert et al. 1993). Several cropping rotations were included. This study showed that proper limited irrigation management and crop rotations did not substantially reduce yield or income.
Potato is highly sensitive to soil moisture and nitrogen due to a shallow, fibrous root system (Alva 2008; Iwama 2008). Research has been reviewed on the effects of no-watering (drought) periods and irrigation scheduling on yield and canopy growth with the aim of optimizing yield (Levy et al. 2013; Shock et al. 2007; van Loon 1981; Vos and Haverkort 2007). The concept of deficit irrigation has been characterized as a way of optimizing irrigation during drought-sensitive stages in potato growth (Geerts and Raes 2009; Levy et al. 2013). Most studies were conducted by reducing water availability to differing degrees all season long (Alva 2008; Ferreira and Carr 2002; Fleisher et al. 2008; Kashyap and Panda 2003; Onder et al. 2005; Shock et al. 1998). There are some studies in which irrigation was completely withheld during select periods related to tuber growth (Dalla Costa et al. 1997; Karam et al. 2014; Levy 1985; Stark and McCann 1992) as well as gradual irrigation reduction during the growing season (Stark et al. 2013). There are few studies on reduced irrigation or mild stress for extended periods (Karam et al. 2014; Stark et al. 2013). Ojala et al. (1990) reported that increased soil N could increase yield under deficit irrigation. Therefore, nitrogen may mitigate the effects of limited irrigation on yield.
Studies on the effects of drought periods on tuber quality such as shape and cracks have been reported primarily on Russet Burbank (Painter and Augustin 1976). More recently, some work on white-skinned cultivars has been reported (Walworth and Carling 2002). Studies on chipping cultivars such as Atlantic, a standard in the industry, are lacking. Research outside of North America has focused on drought-sensitive morphological parameters such as plant stature and leaf size (Deblonde and Ledent 2000; Kashyap and Panda 2003; Lahlou et al. 2003; Tourneux et al. 2003). A study in Turkey (Onder et al. 2005) reported that more than a 33 % reduction in irrigation could significantly reduce yield of an early maturing potato under their semi-arid conditions. These trials compared irrigated plants versus extended water-limited periods.
Western Nebraska is well-known for its low rainfall, and periodic, long and severe droughts causing declining water availability for agriculture (Basara et al. 2013). Irrigation allocations are decreasing, and has or planned to be decreased by as much as 15 cm (Bleed and Babbitt 2015). The objective of this study was to answer the question: Where irrigation allocations are reduced by 15 cm, when should the irrigation be partially withheld with the least amount of loss?
Materials & Methods
Field Trial Conditions
Between 2009 and 2012, four field trials were conducted on potato (Solanum tuberosum) cultivar Atlantic at the University of Nebraska’s Panhandle Research & Extension Center, Scottsbluff (lat. 41.9 N, long. 103.7 W, elevation 1126 m). Soil was a Tripp fine sandy loam at pH 7.8–8.1 and organic matter content of 0.7–1.0 %. The crops previous to potato were sunflower (Helianthus annus), soybean (Glycine max), camelina (Camelina sativa), and canola (Brassica napus), respectively. Seed tubers were cut, and dusted with thiophanate-methyl with mancozeb plus imidacloprid (Tops MZ Gaucho™) by Western Potatoes Inc. (Alliance, NE) and planted in mid-May (Table 1). Emergence (50 %) was about three weeks later. A mix of metribuzin and metolaclor, at mid-label rate, was applied pre-emergence for weed control. No other pesticides were applied during the season. Other than the various rates of urea-ammonium nitrate (UAN), no other fertilizer was added since soil tests showed the soil to be very high in both phosphorus and potassium. Plots consisted of four rows spaced 91 cm apart and 11.6 m long. Plants were spaced 24 cm apart within the rows resulting in a population of 45,000 plants per ha. Stand counts taken between 18 and 21 days after emergence showed an average of 80 % across years. In September, 10 m of the center two rows were harvested 91 to 101 days between emergence and vine kill. Vine kill was accomplished by flailing and plots were harvested within a week thereafter (Table 1). Harvesting was conducted with a 2-row Farmhand harvester.
Irrigation Regimes
Irrigation was applied through an overhead, linear-move system. A 0.32-ha area was divided into 16 equal parts arranged in a 4 by 4 matrix for a Latin Square of main plots. Four irrigation regimes were applied in four replications in a Latin Square; these comprised the main plots. The check regime (full irrigation, FI) was the application of water to supplement rainfall with the aim to achieve 61 cm of applied water from emergence to harvest following a set irrigation schedule (Table 2). Irrigation was scheduled and rainfall monitored semi-weekly. Irrigation was modified based on rainfall the previous 3 to 4 days. Irrigation was reduced on ‘Atlantic’ potatoes by 50 % during three irrigation timing regimes and compared to fully irrigated plants (62–63 cm water available). Plants exposed to reduced irrigation all received 46 cm of water (irrigation + rainfall), a 25 % reduction in water availability but the distribution followed the three regimes. The three reduced irrigation regimes (Table 2) were: 1. outer stress (OS) where applied water was 50 % for the first five weeks and for the last three weeks, i.e., 0 to 5 WAE and 10 to 13 WAE, 2. early stress (ES) where applied water was 50 % during the first 8 weeks, i.e., 0 to 8 WAE, and 3. late stress (LS) where applied water was reduced by 50 % during the last five weeks, i.e., 8 to 13 WAE. In all three limited-irrigation regimes, the goal was to reduce water availability by 15.3 cm compared to fully irrigated (FI) but in different periods. The actual total water availability for FI were 64, 62, 62, and 63 cm for 2009, 2010, 2011, and 2012, respectively, with a four-year mean of 62.6 cm (Table 2). Rainfall and temperature were monitored by the High Plains Regional Climate Center (Changnon et al. 1990) from a weather station located next to the field. The seasonal water use from emergence to vine desiccation for potato was 60 to 61 cm based on evapo-transpiration, and the water holding capacity of the soil was 6 cm in the top 30 cm.
Soil water was monitored using the gravimetric method. Soil cores were taken at 0 (base of hill) to 20 cm below the seed-piece at the start of irrigation and at 5, 8, 10, and 13 WAE for each plot. The soil samples were weighed the same day, dried in an oven set at 107 °C for 22–24 h and re-weighed. Soil moisture as percent was calculated using the standard formula of (soil fresh weight - soil dry weight) divided by soil dry weight and converted to percent. Additionally in 2010, 2011 and 2012, soil moisture was monitored using the Watermark system (Irrometer Co., Inc., Riverside, CA). A pre-soaked sensor was placed in each main plot (irrigation treatments) at a depth of about 20 cm. Water deficit was measured hourly, averaged daily and calculated weekly. Watermark data were compared to gravimetric data.
Nitrogen Application
Each of the 16 irrigation main-plots was sub-divided into three N treatments. These split-plots comprised individual plots for measurements and analysis. Nitrogen was applied three times with three rates of liquid UAN (urea + ammonium nitrate). The three application timings were the day before or just prior to planting, 4 to 8 days before 50 % emergence, and 20 to 25 days after the second application before tuber initiation (Table 1). Nitrogen rates per application were 33.6, 56.0 and 78.5 kg ⋅ ha−1 to achieve 100, 168 and 235 kg ⋅ ha−1 total N, respectively. UAN was applied to the ground using a CO2 plot sprayer set at 2.8 kg ⋅ cm−2 using 9501E nozzles. UAN was diluted with water and sprayed at 196 L ⋅ ha−1. Residual soil N was determined in April, pre-planting, in the top 40 cm, and was 72, 58, 43, and 55 kg ⋅ ha−1 for year 2009, 2010, 2011, and 2012, respectively, with a year average of 57 kg ⋅ ha−1. About 40 % was in the top 20 cm.
Data Collection
Canopy height, width, and weight were measured at the start of irrigation and at each change of irrigation regime, i.e., 5, 8, 10, and 13 WAE (Table 1). Canopy height and width was measured with a meter-stick in three places in the center rows of each plot and averaged at each reading. Canopy weight was determined by cutting off two plants in each plot at the base and weighing the canopy the next day to obtain the fresh weight. After which, the canopy was placed in an aluminum tray and into a drying oven set at 50 °C for 7 to 10 days then re-weighed to obtain its dry weight. The two plants' weights were averaged per plot. In 2010, 2011 and 2012, leaf area index of each plot was measured at the 8 WAE regime change and at harvest, 13 WAE, using an LAI 2200 (Li-Cor Bioscience, Lincoln, NE). Soil N was measured by collecting soil samples at depths of 0 (base of hill or seed piece location) to 20 cm below and from 20 to 40 cm below for each plot at pre-irrigation, 5, 8, 10, and 13 WAE. Soil samples were dried in an oven at 107 °C for 22–24 h then sent to Ward Laboratory, Kearney, NE, for N analysis. At 5, 8, 10, and 13 WAE, the petiole of the fourth or youngest fully-expanded leaf of 20 to 25 plants per plot were removed, pooled and shipped to Ward Laboratory for N content determination. Percent flowering was monitored weekly until after full bloom. Plant senescence was determined visually at 13 WAE by the percent of brown (dying or dead) foliage.
Harvested tubers were placed on a grader fitted with chains to separate tubers by diameters since the cultivar Atlantic produces round white tubers used for chipping and are commonly size-graded. Yield and quality were determined on tubers with diameters between 5.7 to 10.8 cm (A). Tubers with diameters less than 5.7 cm (small) or greater than 10.8 cm (oversize) were discarded. Specific gravity was determined on 3.6 kg samples of 5.7 to 8.3 cm tubers from each plot using a Snack Food Association hydrometer (Gould and Plimpton 1985; Pavlista 1997). Harvested tubers were stored for one month at about 15 °C to allow for healing after which samples were sliced and fried using standard practice (Gould and Plimpton 1985). After frying, slices were evaluated as to lightness using the “Fry Color Standards for Potatoes for Chipping” available through the Snack Food Association. Tuber defects were quantified on 20 tubers, sized between 5.7 and 8.3 cm diameter, per plot. External defects were mis-shaping, common scab caused by Streptomyces scabies, and black scurf caused by Rhizoctonia solani. Internal defects were hollow heart and internal discolorations.
Data were analyzed as factorial comparisons using Proc ANOVA in SAS with means separated using least significant differences and inferences based on a 5 % significance level (SAS Institute 2003) when p < 0.05. Possible interactions between irrigation and nitrogen were also determined.
Results
Soil Water
Differential irrigation began between 1 and 3 WAE depending on previous rainfall that year (Table 1). Following the onset of irrigation, soil water in the 40 cm below the seed piece was measured using gravimetrics at 5, 8, 10 and 13 WAE corresponding with changes in the period of reduced (50 %) water availability (Fig. 1). Following the expected pattern, at 5 WAE, reducing irrigation resulted in less soil water for the ES and OS regimes compared to FI and LS both being fully irrigated at this point. At 5 WAE, OS began to receive the full compliment of irrigation and the soil water level increased to that of FI (Fig. 1). At 8 WAE, ES began being fully irrigated and LS began exposure to 50 % less water availability. At 10 WAE, soil moisture was lower for LS compared to FI (Fig. 1). After 10 WAE, irrigation of OS was again reduced. So, at 13 WAE, soil moisture for both LS and OS were lower than for FI and ES (Fig. 1).

Soil moisture as affected by irrigation schedule was measured from the seed piece level to 40 cm below using gravimetric methodology. Measurements were taken of each plot at each irrigation regime change (5, 8, 10, and 13 weeks after emergence, WAE). Each bar represents the mean of four years (2009, 2010, 2011, and 2012). Bars at each WAE with a different letter above them are significantly different at p < 0.05 using least square separation of the means
Watermark water sensors were installed at 20 cm below seed pieces a few days prior to emergence and soil water was measured from emergence to harvest. There was a steep decline in soil water measured with Watermark sensors after emergence to 4 WAE for FI, LS and OS while for ES that decline continued to 6 WAE (Fig. 2). With FI and LS, soil water rebounded and returned to normal by 7 WAE; OS which was fully irrigated between 5 and 8 WAE, returned by 8 WAE while ES which began full irrigation at 8 WAE did not return to normal until 10 WAE (Fig. 2). After 8 WAE, LS which now had reduced irrigation showed soil water level declining again; the same occurred with OS after 10 WAE (Fig. 2). These data showed that the periods of reduced irrigation had a corresponding decrease in soil moisture content indicating that the irrigation parameters were effective on water availability. The loss of soil moisture early in the season may be explained by rapid root growth from pre-emergence to 3–4 WAE. This water would be needed to support rapid vine growth during the first six weeks following emergence.

Deficit moisture at 18 to 20 cm below the seed piece of potato cv. Atlantic as affected by irrigation schedule as measured using Watermark sensors. Hourly measurements were averaged on a per week basis starting from the day of emergence (0 WAE) to vine kill (13 WAE)
Soil Nitrate-N
The third and last application of N was prior to the onset of irrigation except in 2012 when the drought forced an earlier than planned irrigation start. Soil nitrate-N was measured below the seed piece at the times of irrigation regime change (5, 8, 10, and 13 WAE). Soil nitrate-N levels directly corresponded with applied N levels and decreased over time to 10 WAE (Table 3). Soil nitrate-N was significantly affected by irrigation level. At 5 WAE, soil N for ES and OS under reduced irrigation was significantly lower than FI and LS (Table 3). After 5 WAE, applied water to OS was increased (Table 2). The soil N level declined for FI and LS at 5 WAE as well. The soil nitrate-N level for ES remained statistically level to 10 WAE and was significantly greater than the other irrigation regimes at both 8 and 10 WAE (Table 3). Between 10 and 13 WAE, the soil N for ES was declined while the other irrigation regimes remained constant (Table 3).
Petiole Nitrate-N
At the same time that soil nitrate-N was measured, petioles were collected and sent for nitrate-N analysis. Unlike with soil nitrate-N, at 5 WAE, ES and OS had higher petiole nitrate-N than FI and LS (Table 4). Petiole nitrate-N declined under all regimes; however the regimes differed in the rate of decline. Between 5 and 8 WAE, petiole nitrate-N for FI and LS, both receiving full irrigation, decreased by 60 % and for OS, which had received reduced irrigation before 5 WAE and then returned to full irrigation, the decline was 50 %, and for ES, which received 50 % of the applied water as FI and LS throughout the 8 weeks, petiole nitrate-N decreased by only 30 % (Table 4). At 8 WAE, FI and LS had significantly less petiole nitrate-N than OS or ES but OS was now significantly greater than ES which still had reduced irrigation (Table 4). At 10 WAE, the petiole nitrate-N of FI, ES and OS, which were receiving full irrigation, declined 30 % of their 8 WAE level. On the other hand, LS which during these two weeks was receiving reduced irrigation remained statistically the same in petiole nitrate-N (Table 4). Between 10 and 13 WAE, except for LS, petiole nitrate-N decreased further with FI having the lowst petiole nitrate-N level and ES the highest (Table 4). Petiole nitrate-N was the lowest in plants receiving the lowest applied N which was below that recommended for ‘Atlantic’ (Table 4). There was no difference between the recommended N and the greater amount at 5 and 8 WAE but they differed thereafter with plants receiving the higher N having greater petiole nitrate-N. There was a steady decline in petiole nitrate-N over time for the two lower N applications, but for the highest N application, the decline stopped at 10 WAE (Table 4).
Canopy Height and Weight
Canopy growth was followed by measuring height, dry and fresh weights, and leaf area index (LAI). Applied N level did not affect height or weight (data not shown). Height of ES and OS were lowest at 5 and 8 WAE while the fully irrigated plots, FI and LS, were taller. Dry weight was likewise the lowest for ES and OS at 5 WAE. At 8 WAE, the dry weight of OS, which had been fully irrigated for three weeks, was statistically the same as FI and LS but remained shorter, similar to ES (Table 5). At 10 WAE, ES still had the lowest height and weight compared to the other three regimes. At 13 WAE, all canopies were about 27 cm high. However, ES remained with the lowest dry weight (Table 5). Fresh weight of the canopy was similar and therefore not presented.
Leaf Area Index
Although the effects of reduced irrigation on canopy height and weight showed a similar trend, years differed.
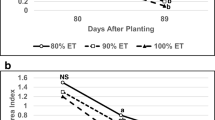
The mean LAI was highest in 2011 and lowest in 2012 (Table 6). In 2012, the canopies were visually shorter, smaller and showed less rapid growth (data not presented). The effect of reduced water application was as might be expected from the height and weight data at 8 WAE (Table 5 compared to Table 6). At 8 WAE, ES which had received 50 % applied water had the lowest LAI; OS, which received less water up to 5 WAE and then returned to full irrigation, was intermediate, and FI and LS, which were fully irrigated during the eight weeks, had the largest LAI (Table 6). At 13 WAE, LAI was reduced below 2 due to leaf senescence and there was no difference between irrigation regimes (data not shown). The level of N applied had no effect on LAI in any year (Table 6).
Flowering
In 2010 and 2011, flowering peaked 5 WAE and showed no difference between FI, OS and ES (data not shown). At 8 WAE, flowering declined and ES had significantly fewer flowers than FI and OS (data not shown). In 2012, at 5 WAE, flowering had not peaked, and by 8 WAE, FI and OS reached peak flowering and had greater flowering than ES (data not shown). Compared to the earlier years, 2012 had a severe drought.
Canopy Senescence
Senescence at 13 WAE was erratic comparing the influence of irrigation regimes and nitrogen application across years; different irrigation treatments showed greater senescence in different years. Averaged across years, there was no effect by either irrigation or nitrogen on senescence (data not shown). Years were significantly different with senescence (% leaf death) being 59 A, 33 B, 63 A, and 44 AB for 2009, 2010, 2011, and 2012, respectively. Since no foliar fungicide was applied, senescence may have been influenced by early blight caused by Alternaria solani.
Tuber Yield and Quality
Yield of tubers, 5.7–10.8 cm diameter, was greatest with full irrigation during the first 8 WAE, i.e., FI and LS (Table 7). The lowest yield was obtained with irrigation by half during this period, i.e., ES. Reducing irrigation between 2 and 5 WAE, returning to full irrigation and then reducing it again for the last three weeks (OS) was not significantly different from FI. Nitrogen level did not affect yield. Yields were similar in 2009, 2010 and 2011, but significantly lower in 2012 (Table 7), and may be explained by the drought conditions of that year as will be discussed. Specific gravity was not affected by irrigation regime or nitrogen applied, but in 2012, as with yield, it was significantly less and below the normal for this cultivar in Nebraska (Table 7). Potato chips from ES were significantly the darkest (Table 7), and chips from OS were significantly darker than FI. Applied N had no effect on chip color (Table 7). The incidence of common scab was significantly greater also with ES compared to the other irrigation regimes while OS and LS showed no difference compared to FI (Table 7). As with yield, specific gravity and chip color, the percent of tubers with common scab was not affected by applied N. Years were significantly different with regard to common scab occurrence; 2010 and 2011 were the highest while 2009 and 2012 had 5 % or less (Table 7). The incidence of black scurf was less than 10 % in all years and near zero in 2010; therefore the data presented are only from 2009 when its incidence was the highest (Table 7). The difference between irrigation regimes was marginal due to limited data but tubers with black scurf may be greater with LS than all the other regimes.
Discussion
Nitrogen in Soil and Plant
Initially, at 5 WAE, soil nitrate-N is lower in ES and OS (Table 3). Between 5 and 8 WAE, ES is the only regime not receiving full irrigation and soil nitrate-N does not decline as it does with the other regimes. ES maintains the highest soil nitrate-N level at 10 WAE. Between 10 and 13 WAE, soil nitrate-N ceases to decline for FI, OS and LS while ES continued (Table 3). These observations imply that, when plants are exposed to reduced water application, plants take up less N from the soil. Dalla Costa et al. (1997) concluded that when season-long water reduction was lower than 80 % ET, N uptake was lowest, agreeing with these findings.
Petiole nitrate-N fluctuated according to level of applied water (Table 4). Although petiole nitrate-N declined over time with all regimes, the rate of decrease was slower when irrigation was reduced and may suggest that when plants are exposed to reduced water application that plants utilize less N in the plant tissue.
Canopy Growth and Development
Canopy growth as measured by height was lowered by less applied water during the first 8 WAE (Table 5). After 8 WAE, height no longer increased as plants began to spread outwardly. Season-long irrigation reduction has been reported to result in shorter potato plants (Fleisher et al. 2008; Sharma et al. 2011; Trebejo and Midmore 1990). Tourneux et al. (2003) reported no significant affect by irrigation reduction on plant height. However, in their study, irrigation reduction did not start until 54 days after plantlets, cultivated in vitro, were transplanted at which time tubers were formed and irrigation was reduced gradually.
Vine dry weight showed a similar pattern in that plants with reduced irrigation weighed less at 5 WAE, but when full irrigation returned to OS, dry weight at 8 WAE was statistically the same between OS and FI (Table 5). From 8 to 13 WAE, OS, LS and FI did not differ. ES, which had reduced irrigation for the first eight weeks, had the lowest weight from 5 to 13 WAE. During the period between 5 and 8 WAE, vines are still rapidly growing, and tubers were already initiated but their growth is slow, often referred to as early bulking. At 8 WAE, vine growth plateaus and tubers start enlarging rapidly (Ojala et al. 1990; Pavlista 1995). This shift in growth focus may explain why the vine was no longer affected by the level of applied water as can be observed with LS’s vine weight at 10 and 13 WAE, and OS’s vine weight at 13 WAE (Table 5). Kashyap and Panda (2003) also reported vine weight loss associated with water deficit however their water regimes involved a season-long reduction in water supply so it is not possible to relate their observations to a specific growth stage.
Leaf growth was estimated using LAI which, in essence, determines the amount of light penetration through the plant across the row. LAI showed the inhibitory effect on canopy growth resulting from reduced applied water. At 8 WAE every year, FI and LS had higher LAI than OS which in turn had a higher LAI than ES (Table 6). The severe drought conditions of 2012 (Table 8) reduced growth of all plants but the pattern of the effect of irrigation regimes was consistent each year for LAI as it was for canopy height and weight. Changes in LAI follows plant growth during the season and rises during tuber bulking followed by a decrease during vine senescence (Wright and Stark 1990). In studies where reduced irrigation was season-long, lower irrigation levels resulted in lower leaf area index (Kashyap and Panda 2003; Trebejo and Midmore 1990; Vos and Haverkort 2007), leaf area and rate of expansion (Fleisher et al. 2008), and leaf number (Vos and Haverkort 2007). Applying water stress to plantlets from 54 days after transplanting also lowered LAI but partial recovery was possible when water was re-applied later (Tourneux et al. 2003).
Tuber Yield and Quality
Yield was most severely reduced when water availability was reduced by 50 % up to 8 WAE (ES) (Table 7). Limiting applied water to 5 WAE and then limiting it between 10 and 13 WAE (OS) also reduced yield compared to FI, the unreduced control (Table 7). Yield for limiting water from 8 to 13 WAE (LS) did not show a significant yield reduction. This indicates that having sufficient water for yield is most critical between 2 and 8 WAE between tuber initiation to mid-tuber bulking. Yield reduction due to reducing applied water early in the season, from tuber initiation to early tuber growth, are greater than when applied water is reduced during tuber bulking have been reported for several potato cultivars (Dalla Costa et al. 1997; Deblonde et al. 1999; Levy 1985). In contrast, some studies on Russet Burbank indicated that reducing water any time during tuber bulking reduced yield (Ojala et al. 1990; Stark and McCann 1992). Stark et al. (2013), when comparing several cultivars, concluded that the least yield reduction was when plants were fully irrigated up to mid-bulking followed by a gradual reduction in water supply. Also, there have been reports that yield reduction was greatest when water stress was applied toward the end of the season or during tuber ripening (Fabeiro et al. 2001; Karam et al. 2014). Deblonde and Ledent (2000) related water stress-induced yield reduction with lower leaf area index and smaller canopy. Comparing Tables 6 and 8, this relation was present also with the cultivar Atlantic. In the study reported here, N application at 101, 168 and 235 kg ⋅ ha−1 did not affect yield.
Specific gravity of ‘Atlantic’ was not significantly affected by water stress nor nitrogen (Table 7). Irregular water stress during tuber bulking has been reported in reviews to lower specific gravity (Wright and Stark 1990; Shock et al. 2007). High N is reported also to lower specific gravity of frying cultivars (Painter and Augustin 1976) but the amounts of N added here were within recommended parameters for the Atlantic cultivar.
Chip color was significantly darker than the 1.5 acceptability threshold when water was limited during the first eight WAE whereas chip color for both OS and LS were not (Table 7). This suggests that the critical moisture period to maintain light chip color is between 5 and 8 WAE, during early tuber bulking. With French frying cultivars, water stress during early tuber bulking was associated with poor distribution of sugars in the tuber resulting in ‘sugar-ends’ as reviewed by Shock et al. (2007).
Irregular water stress during tuber bulking of fresh market and French fry cultivars has been associated with various tuber mis-shaping and growth cracking (Painter and Augustin 1976; Robins and Domingo 1956) but most studies have been on the cultivar Russet Burbank, a French frying and baking cultivar (Singh 1969). Atlantic, a chip cultivar, did not show any promotion of mis-shaping by water stress at any period nor by N in this study (data not shown).
The incidence of common scab was greatest across years with ES while there was no difference between OS and LS compared to FI. This strongly indicates that common scab susceptibility of tubers is between 5 and 8 WAE, early tuber bulking. This timing corresponds to the development of young tubers and the formation of lenticels which are the infection point of Streptomyces scabies, the pathogen causing common scab (Lapwood and Hering 1970). Maintaining adequate water supply during early tuber growth lowers the incidence of common scab (Lapwood et al. 1973).
Because the data on black scurf are limited to one year, little can be concluded but the data from 2009 does suggest that the critical period for tuber susceptibility is between 8 and 10 WAE when tubers are in mid-bulking.
Weather Comparison Across Years
Plant growth and yield results were similar for 2009, 2010 and 2011 but significantly reduced in 2012. The relative effect of irrigation was not substantially altered between years. The rainfall and temperature conditions for these four years are compared in Table 8. From planting to emergence, rain was +/− 30 % of the 30-year average for the first three years but less than half in 2012. Between emergence and 2 WAE, in 2009, rain was twice that of the average resulting in a delay in irrigation to fit the schedule (Tables 1 and 2) while rain in 2010 and 2011 was below but near the average. However, rain in 2012 was about a third of the average (Table 8) causing an earlier than planned start of irrigation. This year is noted for its extreme drought in the High Plains. However, although rain was highly variable, the water scheduling was constant between the years by changing the irrigation scheduling to compliment rain within the regime periods. Therefore, water availability alone cannot explain the year discrepancy. Besides low rainfall, the season of 2012 was noted for its high temperature. During the first two WAE, the temperature was nearly 4 °C above the 30-year average (Table 8) and this continued throughout the season (data not shown). Although temperature could explain some of the year differences, the combination of low rain and high temperature result in low relative humidity. Observations were made that in 2012 while collecting data, there was practically no dew on the leaves. Dew can be a major source of water uptake to plants in addition to roots. Could the lack of dew account for the low performance of potato plants in 2012?
Conclusion
The primary objective of this study was to identify when reducing water availability by half, i.e., 15 cm reduction, would be least deleterious to the chipping potato cultivar Atlantic, to maintain plant health, yield and marketable tuber quality. The answer appears to be that it is best to withhold the 15 cm water toward the end of the growing season, i.e., the last 5 weeks before harvest (8 to 13 WAE). The poorest plant growth and lowest yield occurred when applied water was reduced by half at tuber initiation through to early tuber bulking period (early log phase of tuber growth), from 2 to 8 WAE. However, when tuber quality, i.e., chip color and common scab incidence, and plant health, i.e., LAI, weight and height, are considered as well then the worst time to reduce water is during the early tuber bulking period between 5 and 8 WAE. Reducing or increasing the application of N, by 40 % of the recommended rate, early in the season compared to the recommended amount for cv. Atlantic, had no mitigating effect on tuber yield or quality.
References
Alva A.K. 2008. Setpoints for potato irrigation in sandy soils using real-time, continuous monitoring of soil-water content in soil profile. Journal of Crop Improvement 21: 117–137. doi:10.1080/15427520701885311.
Basara, J.B., Maybourn, J.N., Peirano, C.M., Tate, J.E., Brown. P.J., Hoey, J.D., Smith, B.R. 2013. Drought and associated impacts in the great plains of the United States - a review. International Journal of Geosciences 4:72–81.
Bleed, J.B., Babbitt, C.H. 2015. Nebraska’s Natural Resources Districts: An assessment of a large-scale locally controlled water governance framework. Policy Report 1, Robert B. Daugherty Water for Food Inst., Univ. of Nebraska-Lincoln, 155 pp.
Changnon S.A., P. Lamb, and K.G. Hubbard. 1990. Regional climate centers: new institutions for climate services and climate-impact research. Bulletin of the American Meteorological Society 71: 527–537.
Dalla Costa L., G. Delle Vedove, G. Gianquinto, R. Giovanardi, and A. Peressotti. 1997. Yield, water use efficiency and nitrogen uptake in potato: influence of drought stress. Potato Research 40: 19–34.
Deblonde P., and J.F. Ledent. 2000. Effects of moderate drought conditions on crop growth parameters and earliness of six potato cultivars under field conditions. Agronomie 20: 595–608.
Deblonde P.M.K., A.J. Haverkort, and J.F. Ledent. 1999. Responses of early and late potato cultivars to moderate drought conditions: agronomic parameters and carbon isotope discrimination. European Journal of Agronomy 11: 91–105.
Fabeiro C., R. Martin de Santa Olalla, and J.A. de Juan. 2001. Yield and size of deficit irrigated potatoes. Agricultural Water Management 48: 255–266.
Ferreira T.C., and M.K.V. Carr. 2002. Responses of potatoes (Solanum tuberosum L.) to irrigation and nitrogen in a hot, dry climate. Field Crops Research 78: 51–64.
Fleisher D.H., D.J. Timlin, and V.R. Reddy. 2008. Interactive effects of carbon dioxide and water stress on potato canopy growth and development. Agronomy Journal 100: 711–719.
Geerts S., and D. Raes. 2009. Deficit irrigation as an on-farm strategy to maximize crop water productivity in dry areas. Agricultural Water Management 96: 1275–1284.
Gould, W.A. and S. Plimpton. 1985. Quality evaluation of potato cultivars for processing. North Central Regional Research Pulication # 305.
Hergert G.W., N.L. Klocke, J.L. Petersen, P.T. Nordquist, R.T. Clark, and G.A. Wicks. 1993. Cropping systems for stretching limited irrigation supplies. Journal of Production Agriculture 6: 520–529.
Iwama K. 2008. Physiology of the potato: new insights into root system and reprecussions for crop management. Potato Research 51: 333–353. doi:10.1007/s11540-008-9120-3.
Karam, F., N. Amacha, S. Fahed, T. El Asmar, and A. Dominguez. 2014. Response of potato to full and deficit irrigation under semiarid climate: Agronomic and economic implications. Agricultural Water Management 142:144–151. doi.org/10.1016/j.agwat.2014.05.007
Kashyap P.S., and R.K. Panda. 2003. Effect of irrigation scheduling on potato crop parameters under water stressed conditions. Agricultural Water Management 59: 49–66.
Lahlou O., S. Ouattar, and J.F. Ledent. 2003. The effect of drought and cultivar on growth parameters, yield and yield components of potato. Agronomie 23: 257–268.
Lapwood D.H., and T.F. Hering. 1970. Soil moisture and the infection of young potato tubers by Streptomyces scabies (common scab). Potato Research 13: 296–304.
Lapwood D.H., L.W. Welling, and J.H. Hawkins. 1973. Irrigation as a practical means to control potato common scab (Streptomyces scabies): final experiment and conclusions. Plant Pathology 22: 35–41.
Levy D. 1985. The response of potatoes to a single transient heat or drought stress imposed at different stages of tuber growth. Potato Research 28: 415–424.
Levy D., W.K. Coleman, and R.E. Veilleux. 2013. Adaptation of potato to water shortage: irrigation management and enhancement of tolerance to drought and salinity. American Journal of Potato Research 90: 186–206. doi:10.1007/s12230-012-9291-y.
Ojala J.C., J.C. Stark, and G.E. Kleinkopf. 1990. Influence of irrigation and nitrogen management on potato yield and quality. American Potato Journal 67: 29–43.
Onder S., M.E. Caliskan, D. Onder, and S. Caliskan. 2005. Different irrigation methods and water stress effects on potato yield and yield components. Agricultural Water Management 73: 73–86.
Painter C.G., and J. Augustin. 1976. The effect of soil moisture and nitrogen on yield and quality of the russet Burbank potato. American Potato Journal 53: 275–284.
Pavlista, A.D. 1995. Potato production stages: Scheduling key practices. Univ. Nebraska Ext. Circ. #1249. Potato Education Guide. http://cropwatch.unl.edu/web/potato/key_production_stages
Pavlista A.D. 1997. Potato types & characteristics: laboratory exercises. The Amer. Biology Teacher 59: 30–34.
Robins J.S., and C.E. Domingo. 1956. Potato yield and tuber shape as affected by severe soil-moisture deficits and plant spacing. Agronomy Journal 48: 488–492.
SAS Institute. 2003. SAS System for Windows. Release 9.1. ed. SAS Institute Inc., Cary, NC.
Sharma N., P. Kumar, M.S. Kadian, S.K. Pandey, S.V. Singh, and S.K. Luthra. 2011. Performance of potato (Solanum tuberosum) clones under water stress. Indian Journal Agricultural Sciences 81: 825–829.
Shock C.C., E.B.G. Feibert, and L.D. Saunders. 1998. Potato yield and quality response to deficit irrigation. Hortscience 33: 655–659.
Shock C.C., A.B. Pereira, and E.P. Eldredge. 2007. Irrigation best management practices for potato. American Journal of Potato Research 84: 29–37.
Singh G. 1969. A review of the soil-moisture relationship in potatoes. American Potato Journal 46: 398–403.
Stark J.C., and I.R. McCann. 1992. Optimal allocation of limited water supplies for russet Burbank potatoes. American Potato Journal 69: 413–421.
Stark J.C., S.L. Love, B.A. King, J.M. Marchall, W.H. Bohl, and T. Salaiz. 2013. Potato cultivar response to seasonal drought patterns. American Journal of Potato Research 90: 307–316.
Tourneux C., A. Devaux, M.R. Camacho, P. Mamant, and J.F. Ledent. 2003. Effects of water shortage on six potato genotypes in the highlands of Bolivia (I): morphological parameters, growth and yield. Agronomie 23: 169–179.
Trebejo I., and D.J. Midmore. 1990. Effect of water stress on potato growth, yield and water use in a hot and a cool tropical climate. Journal of Agriculture Science (Cambridge) 114: 321–334.
van Loon C.D. 1981. The effect of water stress on potato growth. Development, and yield. American Potato Journal 58: 51–69.
Vos, J. and A.J. Haverkort. 2007. Water availability and potato crop performance. In Potato Biology and Biotechnology Advances and Perspectives, eds. D. Vreugdenhil, J. Bradshaw, C. Gebhardt, F. Govers, M.A. Taylor, D.K.I. McKerron, and H.A. Ross. pub. Elsevier. Amsterdam, The Netherlands. pp. 333–351.
Walworth J.L., and D.E. Carling. 2002. Tuber initiation and development in irrigated and non-irrigated potatoes. American Journal of Potato Research 79: 387–395.
Wright J.L., and J.C. Stark. 1990. Potato. In irrigation of agricultural crops, eds, 859–887. Madison, WI, USA: B.A. Stewart and D.R. Nielsen. pub. ASA/CSSA/SSSA.
Acknowledgments
The author is grateful for the technical support of Les Kampbell, Allison Hazen and Todd Wilson, and the assistance of summer interns. I give special appreciation for the financial support of the USDA Specialty Crops Block Grant Program administered by the Nebraska Department of Agriculture.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Pavlista, A.D. Scheduling Reduced Irrigation on ‘Atlantic’ Potato for Minimal Effect. Am. J. Potato Res. 92, 673–683 (2015). https://doi.org/10.1007/s12230-015-9483-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12230-015-9483-3