
Abstract
Aluminum exposure can mediate either acute toxicity or chronic toxicity. Aluminum exerts toxic effects on the cardiovascular system, but there are few studies on its related mechanisms. In this study, we investigated the molecular mechanism of aluminum-induced oxidative damage and apoptosis in rat cardiomyocytes. Thirty-two male Wistar rats were randomly divided into four groups, including the control group (GC), low-dose group of aluminum exposure (GL), medium-dose group (GM), and high-dose group (GH), with eight rats in each group. The GL, GM, and GH groups were given 5, 10, and 20 mg/(kg·d) of AlCl3 solution by intraperitoneal injection, and the GC group received intraperitoneal injection of the same volume of normal saline (2 ml/rat/day), 5 times a week for 28 days. At the end of the experiment, the levels of aluminum, malondialdehyde (MDA), plasma lactate dehydrogenase (LDH), creatine kinase (CK), creatine kinase isoenzyme (CKMB), and alpha-hydroxybutyrate dehydrogenase (HBDH) were measured. The pathological changes of myocardium were observed by H&E staining. The apoptosis of cardiomyocytes was detected by TUNEL staining, and the expression of apoptosis-related proteins was determined by western blot. The results showed that the levels of CKMB and HBDH in the GM and GH groups were significantly higher than those in the GC group (P < 0.05). The content of aluminum in the myocardium and serum of the aluminum exposure groups was significantly higher than that of the GC group (P < 0.05). The level of MDA in the GM and GH groups was significantly higher than that in the GC group (P < 0.05). The pathological results showed that vacuolated and hypertrophied cardiomyocytes were found in aluminum exposure groups, especially in the GM and GH groups. The TUNEL staining showed that the apoptosis rate of the aluminum exposure groups was considerably higher than that of the GC group (P < 0.05). Western blot showed that the expression of Bcl-2, an anti-apoptotic protein, in cardiomyocytes of aluminum exposure groups was lower than that of the GC group (P < 0.05), while the levels of Bax and caspase-3 in the cardiomyocytes of the GM and GH groups were higher than those of the GC group (P < 0.05). The experimental results showed that aluminum could accumulate in myocardial tissues and cause damage to cardiomyocytes. It could induce oxidative stress damage by increasing the content of MDA in cardiomyocytes and trigger cardiomyocyte apoptosis by activating the pro-apoptotic proteins caspase-3 and Bax and reducing the anti-apoptotic protein Bcl-2.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Aluminum is the most abundant metallic element in the Earth’s crust, which is widely used in our daily life and industry due to its good physical and chemical properties. It has been shown that aluminum is a non-essential element in organisms, which can mediate acute toxicity or chronic toxicity [1]. Among various toxic effects caused by aluminum, neurotoxicity is undoubtedly the most prominent. Studies have confirmed that aluminum toxicity is involved in neurodegenerative diseases such as Alzheimer’s disease and Parkinson’s disease [2, 3]. In addition, it can also cause liver, blood, immune, kidney, reproduction, and bone tissue damage [4,5,6].
With the development of research, the cardiovascular toxicity of aluminum and its compounds has attracted wide attention [7]. Epidemiological studies have found that aluminum has toxic effects on the cardiovascular system. With the increase of aluminum exposure, the risk of hypertension, coronary heart disease, atherosclerosis, and other cardiovascular diseases increases, and mortality of cardiovascular diseases also increases [8,9,10]. A previous study has found that aluminum can accumulate in the myocardium, leading to myocardial injury and affecting the electrical activity and systolic function of the heart [11]. However, the mechanism of aluminum cardiotoxicity is still unclear and urgent to be clarified.
It has been verified that oxidative stress is the main pathway of metal toxicity [12]. Apoptosis induced by mitochondrial damage is one of the main mechanisms of oxidative stress injury in cardiomyocytes [13]. The Bcl-2 family participates in the mitochondrial pathway of apoptosis and induces apoptosis through a series of processes, ultimately through the activation of caspase-3 [14]. This study aimed to investigate the molecular mechanism of aluminum-induced oxidative damage and apoptosis in rat cardiomyocytes, thereby providing a basis for the risk assessment of cardiovascular toxicity induced by aluminum exposure.
Materials and Methods
Reagent
Al chloride (Howei Chemical Co., Ltd., Guangzhou, China) was diluted with distilled water prior to administration.
Grouping and Modeling of Animals
Thirty-two male Wistar rats of SPF grade, weighing 150–180 g, were provided by Changsha Tianqin Biotechnology Co., Ltd. (License No. SCXK (Xiang) 2019-0014). The rats were reared at 27 °C room temperature and 50% humidity. The rats were fed with standard commercial rodent diet (Changsha Tianqin Biotechnology Co., Ltd., Changsha, China) and drinking water ad libitum. After 1 week of adaptation, thirty-two rats were randomly divided into the blank control group (GC), low-dose group (GL), medium-dose group (GM), and high-dose group (GH), with eight rats in each group. According to the methods in references [15, 16], the rats from the GL, GM, and GH groups were intraperitoneally injected with 5, 10, and 20 mg/(kg·d) AlCl3 solution, which was prepared with distilled water and autoclaved. The rats from the control group were injected with the same volume of normal saline (2 ml/rat/day). The frequency of injection was once a day, 5 days a week, for a total of 28 days. The rats were observed daily for abnormalities in food intake, drinking, behavior, urine, and feces. All animal experiments and procedures used in this study were approved by the Ethics Committee of Animal Experiments of Youjiang Medical University for Nationalities (approval date, 20 July 2020), according to the recommendations of the Guide for the Care and Use of Laboratory Animals in our institutes. All surgery was performed under pentobarbital anesthesia with minimum suffering in the animals.
Sample Collection
After fasting for 12 h at the end of modeling, the rats were anesthetized by i.p. administration of pentobarbital (45 mg·kg−1·BW−1). After anesthesia, 6 to 8 ml of blood was drawn from the abdominal aorta to a common anticoagulant-free biochemical tube. After centrifugation for 5 min at a speed of 4000 rpm, one serum sample was taken to determine the myocardial enzyme and the other sample was utilized to detect the content of aluminum.
The bloodstains of the heart tissue were rinsed with normal saline. Some of them were fixed with formalin for 48 h for routine section and embedding. The sections were then stained with hematoxylin and eosin (H&E) and TUNEL by the Pathology Department technicians. The pathological changes and apoptosis of cardiomyocytes were observed under a light microscope, and the other part was stored at − 80 °C in a cryopreservation tube for subsequent biological tests.
Determination of the Myocardial Enzyme
Lactate dehydrogenase (LDH), creatine kinase (CK), creatine kinase isoenzyme (CKMB), and alpha-hydroxybutyrate dehydrogenase (HBDH) were measured using an automatic biochemical analyzer (BECKMAN COULTER LX20, USA).
The experimental detection was carried out according to the operation instructions of kits (lactate dehydrogenase activity assay kit, creatine kinase activity assay kit, creatine kinase isoenzyme activity assay kit, alpha-hydroxybutyrate dehydrogenase activity assay kit (Colorimetric) (Biovision, Wuhan, China)) and instruments. Only the separated serum needs to be put on the machine. All other operations including sample loading and separation, stirring, and incubation were performed by the automatic biochemical analyzer.
Determination of Aluminum Content
The ICP-OES method was used to identify the level of aluminum in blood and heart tissue. The method for preprocessing the sample was as follows: the weighed sample was put into a digestion tube of polytetrafluoroethylene and added with a mixed solution of 0.5 ml hydrofluoric acid and 7 ml nitric acid. The reaction system was placed in a digestion tank and digested for 8 h at 180 °C. After complete dissolution and cooling, the sample was transferred to a constant volume apparatus for testing.
Determination of Malondialdehyde
The extent of lipid peroxidation by measuring thiobarbituric acid reactive substances (TBARS) in terms of malondialdehyde (MDA) formation was measured according to the method of Draper and Hadley [17], and the content of MDA was determined according to the methods indicated in the kit instructions (Beyotime Bio, Beijing, China). Take 100 mg of myocardial tissue into a cryopreservation tube for use. Normal saline homogenate of myocardial tissue was prepared under the condition of 1 to 4 °C, and 0.5 ml of a heart extract supernatant was mixed with 1 ml of trichloroacetic acid solution and centrifuged at 2500g for 10 min. The resulting supernatant (0.5 ml) and 1 ml of a solution containing 0.67% thiobarbituric acid (TBA) were incubated for 15 min at 90 °C and cooled. The mixture was measured for absorbance at 532 nm using a spectrophotometer (Jenway UV-6305, Shimadzu, Japan). The malondialdehyde values were calculated using 1,1,3,3-tetraethoxypropane as a standard and expressed as nanomoles of malondialdehyde per milligram of protein.
Histopathological Examination
Rat myocardial tissue was fixed in formalin solution, followed by paraffin embedding, conventional 4-μm section, and H&E staining. The morphological changes of myocardial tissue were observed under an optical microscope (BX53, Olympus, Japan).
TUNEL Staining
Myocardial tissue was embedded in paraffin and sectioned routinely. After dewaxing, specimens were treated with protease K to digest membrane protein and nuclear protein and then incubated with the TUNEL reaction solution at 37 °C for 1 h. The anti-fluorescein antibody was incubated and then visualized with DAB. The section was dehydrated, transparent, and mounted after nucleus counterstaining. The cells whose nuclei were stained brown-yellow were positive apoptotic cells. At high magnification, three fields were randomly selected from each section to calculate the apoptosis rate of cardiomyocytes in each field. The apoptosis rate (%) was equal to the number of apoptotic cells divided by the total number of cells.
Western Blot Analysis of Bcl-2, Bax, and Caspase-3
The heart tissue was ground with a liquid nitrogen grinder, and then, 100 μl of RIPA lysis buffer (Beyotime Bio, Beijing, China) per 10 mg of tissues was added. After shaking and mixing, the lysate was placed on ice for 5 min and centrifuged at 4 °C for 10 min (13,000 rpm). The supernatant was collected and the total myocardial protein was extracted. The BCA method was used to determine the protein concentration. After the loading concentration was adjusted, the protein was denatured and stored at −20 °C. Next, 5% stacking gel and 12% separation gel were prepared to load protein samples. GAPDH (Abbkine, Wuhan, China) was used as the internal reference. After loading the samples, a constant voltage of 80 V was first applied to electrophoresis until the bromphenol blue indicator runs to the junction of the two gels in a line. Then, change to a constant voltage of 120 V until the bromphenol blue runs to the bottom of the gel. The gel was transferred to the filter paper wet in transfer buffer solution at a constant current of 300 mA. After blocking, the membrane was placed in a shaker and incubated with rabbit polyclonal antibody (1:3000, Proteintech, Wuhan, China) of Bcl-2, Bax, and caspase-3 at 4 °C overnight. After rinsing the membrane thoroughly, the secondary antibody was hereby added and incubated at room temperature for 1.5 h. The film was exposed in the dark room after rinsing again. The film was then scanned after drying. The gray values were analyzed with Image-Pro Plus 6.0 (Media Cybemetics, USA).
Statistical Analysis
Statistical analyses were carried out using SPSS 20.0. The quantitative data were expressed as mean ± standard deviation (\( \overline{x}\pm s \)). One-way analysis of variance (ANOVA) was used for comparison among groups and the LSD test was used for comparison between any two of these groups. The non-parametric test (Kruskal-Wallis H test) was used when the variance was not uniform. The difference was statistically significant when P < 0.05. Data analysis and graph construction were performed with the aid of GraphPad Prism 5 (GraphPad Software Inc., La Jolla, CA, USA).
Results
Myocardial Enzyme Levels
The myocardial enzyme levels of rats in this study were determined to understand the pathogenesis of cardiac toxicity and the effect of aluminum, and the results are listed in Table 1. With the increase of aluminum exposure concentration, the levels of myocardial enzymes tended to increase. Compared with that of the GC group, the serum LDH level of the GH group was significantly increased (P < 0.05); compared with that of the GC group, the serum CK level of the GM group was significantly increased (P < 0.05); the serum CKMB and HBDH levels of the GM and GH groups were significantly higher than those of the GC group (P < 0.05) (Table 1).
Levels of Aluminum in Serum and Tissues
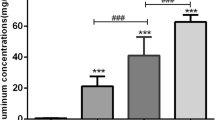
After 28 days of aluminum exposure, the aluminum content in the myocardium and serum of rats was detected. Our results showed that the concentration of aluminum in the myocardium and serum showed an upward trend with the increase of aluminum exposure concentration. Compared with that of the GC group, the content of aluminum in the myocardium of the GL, GM, and GH groups was significantly increased (P < 0.05), showing a significant dose-response relationship in the myocardium (Fig. 1).

Aluminum concentrations in the serum (a) and myocardium (b). GC, GL, GM, and GH refer to 0, 5, 10, and 20 mg/(kg·d) of AlCl3 solution, respectively. Results are expressed as mean ± SEM. Statistical significance was determined by one-way analysis of variance (ANOVA) (a) and Kruskal-Wallis H test (b) (n = 8, aP < 0.05 vs GC; bP < 0.05 vs GL; cP < 0.05 vs GM)
The Level of MDA in Myocardium
To investigate the oxidative stress level, MDA was used as an indicator of lipid peroxidation in the myocardium. As shown in Fig. 2, with the increase of aluminum exposure concentration, the concentration of MDA showed an upward trend; compared with that of the GC group, the myocardial aluminum content of the GM and GH groups was increased significantly (P < 0.05) (Fig. 2).

Concentration of MDA. Results are expressed as mean ± SEM. GC, GL, GM, and GH refer to 0, 5, 10, and 20 mg/(kg·d) of AlCl3 solution, respectively. Statistical significance was determined by one-way analysis of variance (ANOVA) (n = 8, aP < 0.05 vs GC; bP < 0.05 vs GL; cP < 0.05 vs GM)
H&E Staining of Myocardial Tissue
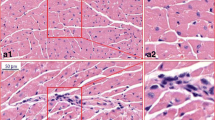
The effect of aluminum exposure on myocardial morphological changes was observed by H&E staining. The cardiomyocytes of the GC rats were arranged closely with clear boundary, uniform staining, and abundant cytoplasm; no obvious thickening was seen in the pericardium, and no obvious inflammation was found in the tissues. A few hypertrophic cardiomyocytes were scattered in the GL and GM groups (black arrow). As for the GH group, a small number of cardiomyocytes underwent vacuolar degeneration, which was manifested by small round vacuoles in the cytoplasm (blue arrow); a few hypertrophic cardiomyocytes were scattered, and some cardiac muscle fibers broken (Fig. 3).

Pathological changes of myocardium tissues of rats in each group (H&E ×400). GC, GL, GM, and GH refer to 0, 5, 10, and 20 mg/(kg·d) of AlCl3 solution, respectively. The scale bar represents 50 μm. Hypertrophic cardiomyocyte cells (black arrow) and vacuolar degeneration (blue arrow)
TUNEL Staining
The apoptosis of cardiomyocytes was detected by TUNEL staining. As shown by the TUNEL staining in Fig. 4, the apoptosis rate of cardiomyocytes showed an upward trend with the increase of aluminum exposure concentration; the apoptosis rate of cells in the GL, GM, and GH groups was significantly higher than that in the GC group, and the difference was statistically significant (P < 0.05) (Fig. 4).

The TUNEL-positive renal cells for four groups of rats (TUNEL staining, ×200) and TUNEL-positive cell count in rat myocardium. GC, GL, GM, and GH refer to 0, 5, 10, and 20 mg/(kg·d) of AlCl3 solution, respectively. Results are expressed as mean ± SEM. Statistical significance was determined by one-way analysis of variance (ANOVA) (n = 3, aP < 0.05 vs GC; bP < 0.05 vs GL; cP < 0.05 vs GM)
Comparison of the Expression of Bcl-2, Bax, and Caspase-3
To investigate the effects of aluminum exposure on the levels of Bcl-2, Bax, and caspase-3, we detected the levels of these proteins using western blot. Our results showed that the level of Bcl-2 in the myocardium of the GL, GM, and GH groups was significantly decreased compared with that of the GC group (P < 0.05). With the increase of aluminum exposure concentration, the levels of Bax and caspase-3 in cardiomyocytes showed an upward trend; compared with that of the GC group, the level of Bax in the myocardial tissue was significantly increased in each dose of the aluminum exposure groups (P < 0.05); the level of caspase-3 in the myocardial tissue of the GM and GH groups was higher than that of the GC group (P < 0.05) (Fig. 5). Taken together, these results suggested that aluminum exposure could activate apoptosis by increasing the pro-apoptotic proteins Bax and caspase-3, and inhibiting anti-apoptotic protein Bcl-2.

The protein expression of Bax, Bcl-2, and caspase-3 in the myocardium tissues of each group. GC, GL, GM, and GH refer to 0, 5, 10, and 20 mg/(kg·d) of AlCl3 solution, respectively. Statistical significance was determined by one-way analysis of variance (ANOVA) (n = 8, aP < 0.05 vs GC; bP < 0.05 vs GL; cP < 0.05 vs GM)
Discussion
Aluminum in drinking water, food, aluminum cookers, tableware, and drugs can enter the body through the skin, respiratory tract, and especially the digestive tract [18]. The majority of aluminum ions absorbed into the blood is combined with plasma proteins, and a small part forms free aluminum [19]. The combined aluminum and free aluminum can not only be eliminated by kidney but also be deposited in various tissues and organs with blood flow through the capillary wall. With the increase of aluminum intake, the amount of aluminum deposition is also accumulating, which aggravates the aluminum load in tissues and serum and eventually causes damage to tissues and organs [20, 21]. The blood aluminum level directly reflects the degree of aluminum exposure, while the tissue aluminum level can reflect the degree of aluminum accumulation and the possible damage to the tissues. This study demonstrated that the rats exposed to aluminum at different doses showed increased blood aluminum level compared with the control rats; the aluminum accumulation in the myocardium was enhanced with the increase of aluminum exposure dose, showing an obvious dose dependence, which was consistent with the study of Novaes RD et al. [11]. It was suggested that the heart was also one of the target organs of aluminum toxicity.
The early reaction of the heart caused by the short-term effect of cardiovascular toxicants is biochemical alternation, such as the changes of myocardial enzyme activity, energy metabolism, and ion homeostasis [22]. A series of cellular and molecular events of cardiomyocytes can be triggered by the continuous action of toxicants. Myocardial hypertrophy can be induced by the activation of hypertrophy genes and upregulation of transcription factors [23]. Myocardial hypertrophy in a non-physiological state at the initial stage belongs to the compensatory response of the heart to the changes of cardiac function caused by toxic effects, and such myocardial injury is reversible [24]. If the toxic substances still cannot be removed, a series of changes in the shape and function of the heart will appear, eventually leading to cardiomyocyte death characterized by cardiomyocyte apoptosis and necrosis [25].
Myocardial enzymes are important markers for the diagnosis of myocardial injury. Cardiomyocyte injury leads to the leakage of enzymes in cardiomyocytes to the outside of cells, so the release of these enzymes can be used as an indicator of the degree of cell injury [26]. This study showed that the levels of CKMB and HBDH in the middle- and high-dose groups of rats were significantly higher than those in the control rats. The LDH level in the high-dose group of rats was higher than that in the control rats, which indicated that aluminum load could induce cardiomyocyte injury. Additionally, H&E staining revealed that the cardiomyocytes of aluminum-exposed rats showed pathological damages, mainly manifested as vacuolar degeneration and myocardial hypertrophy. Cardiac hypertrophy attenuates the ability of cells to transport ions, including the relative deficiency of Ca2+ influx, which reduces the function of cardiomyocytes [27]. Moreover, the biological oxidation in hypertrophic myocardium is relatively weakened, resulting in insufficient myocardial energy generation and decreased cardiac function [28]. The pathological results further demonstrated the direct toxic effect of aluminum on rat myocardium.
Oxidative damage plays a vital role in the mechanism of aluminum toxicity. Aluminum is a positive trivalent element with relatively active properties, which can generate a large amount of ROS by interfering with the mitochondrial electron transport chain [29]. When the production of ROS exceeds the clearance rate of the body, the oxidation system and antioxidant system are out of balance; the excessive ROS participates in the intracellular reaction, causes lipid peroxidation, and finally generates MDA [30]. MDA can damage the biomembrane and cause cross-linking of proteins, nucleic acids, and other macromolecules, leading to cell metabolism dysfunction and even death. Due to the short half-life of ROS and the high determination requirement, this study adopted MDA to measure the degree of oxidation reaction. The results showed that the MDA level in the serum of rats exposed to aluminum was significantly higher than that of the control rats, which indicated that aluminum exposure triggered the oxidative stress reaction and activated the process of lipid peroxidation in vivo, thus increasing the production of MDA, the end product of lipid peroxidation. The result of this study was consistent with the finding of Ghorbel I. et al. [17], suggesting that aluminum caused myocardial injury through oxidative stress.
Oxidative stress is considered a critical regulatory signal of apoptosis [31]. The heart is composed primarily of long-lived and post-mitotic cells, which prefer fatty acids as substrate for energy production, so it becomes more susceptible to oxidative damage than other tissues. Oxidative damage leads to Ca2+ overload, MDA content increase, mitochondrial matrix over-swelling, permeability increase, and cytochrome C release. The free cytochrome C in the cytoplasm can activate caspase-related proteins to initiate the intracellular apoptosis signaling pathway, thus causing irreversible cardiomyocyte injury and apoptosis [32, 33]. Due to the nonrenewable nature of cardiomyocytes, apoptosis may lead to the permanent loss of some cardiac functions. The caspase family is one of the core proteins in apoptosis regulation. Emerging evidence has found that the caspase family proteins involved in cardiomyocyte apoptosis are mainly caspase-8 and caspase-3. Caspase-8 is located upstream of the apoptotic pathway, which can stimulate the downstream caspase-3 to cause apoptosis [34]. In this study, western blot results revealed that the myocardial tissues of rats exposed to aluminum showed increased levels of pro-apoptotic proteins (Bax and caspase-3) and decreased level of anti-apoptotic protein (Bcl-2), suggesting that Bcl-2, Bax, and caspase-3 might be implicated in cardiomyocyte apoptosis induced by aluminum. In the apoptosis signal transduction pathway, Bcl-2 is an anti-apoptotic protein and related to the stability of mitochondria, which can protect the stability of the cell membrane and prevent the transduction of apoptosis signal. The increase of Bax can form a heterodimer with Bcl-2 protein, which inactivates Bcl-2 and triggers the cascade reaction of caspase [35]. Caspase-3, the key executive molecule of apoptosis in the caspase family, is activated at the early stage of apoptosis and induces apoptosis [36]. A previous study has demonstrated that the number of early and late apoptotic cells is increased significantly after ALP treatment through flow cytometry assay, and apoptosis is mainly induced via the mitochondrial pathway [37], which is consistent with the conclusion of this study. TUNEL staining also showed that the apoptosis rate of cardiomyocytes in rats exposed to aluminum was significantly increased, which confirmed the toxicity of aluminum load on the myocardial injury.
In conclusion, the experimental results showed that aluminum could accumulate in myocardial tissues and cause damage to cardiomyocytes. It could induce oxidative stress damage by increasing the content of MDA in cardiomyocytes and trigger cardiomyocyte apoptosis by activating the pro-apoptotic proteins caspase-3 and Bax and reducing the anti-apoptotic protein Bcl-2.
References
Shaw CA, Tomljenovic L (2013) Aluminum in the central nervous system (CNS): toxicity in humans and animals, vaccine adjuvants, and autoimmunity. Immunol Res 56(2-3):304–316. https://doi.org/10.1007/s12026-013-8403-1
Nie J (2018) Exposure to aluminum in daily life and Alzheimer’s disease. Adv Exp Med Biol 1091:99–111. https://doi.org/10.1007/978-981-13-1370-7_6
Laabbar W, Elgot A, Elhiba O, Gamrani H (2019) Curcumin prevents the midbrain dopaminergic innervations and locomotor performance deficiencies resulting from chronic aluminum exposure in rat. J Chem Neuroanat 100:101654. https://doi.org/10.1016/j.jchemneu.2019.101654
Mouro VGS, Menezes TP, Lima GDA, Domingues RR, Souza ACF, Oliveira JA, Matta SLP, Machado-Neves M (2018) How bad is aluminum exposure to reproductive parameters in rats? Biological trace element research 183(2):314–324. https://doi.org/10.1007/s12011-017-1139-3
Sun X, Liu J, Zhuang C, Yang X, Han Y, Shao B, Song M, Li Y, Zhu Y (2016) Aluminum trichloride induces bone impairment through TGF-β1/Smad signaling pathway. Toxicology 371:49–57. https://doi.org/10.1016/j.tox.2016.10.002
Balgoon MJ (2019) Assessment of the protective effect of lepidium sativum against aluminum-induced liver and kidney effects in albino rat. Biomed Res Int 2019:4516730–4516739. https://doi.org/10.1155/2019/4516730
Yuan Y, Xiao Y, Feng W, Liu Y, Yu Y, Zhou L, Qiu G, Wang H, Liu B, Liu K, Yang H, Li X, Min X, Zhang C, Xu C, Zhang X, He M, Hu FB (2017) Plasma metal concentrations and incident coronary heart disease in Chinese adults: the Dongfeng-Tongji cohort. Environmental health perspectives 125(10):107007. https://doi.org/10.1289/EHP1521
Schmidt PM, Escobar AG, Torres JG, Martinez CS, Rizzetti DA, Kunz SN, Vassallo DV, Alonso MJ, Peçanha FM, Wiggers GA (2016) Aluminum exposure for one hour decreases vascular reactivity in conductance and resistance arteries in rats. Toxicol Appl Pharmacol 313:109–118. https://doi.org/10.1016/j.taap.2016.10.023
Martinez CS, Piagette JT, Escobar AG, Martín Á, Palacios R, Peçanha FM, Vassallo DV, Exley C, Alonso MJ, Miguel M, Salaices M, Wiggers GA (2017) Aluminum exposure at human dietary levels promotes vascular dysfunction and increases blood pressure in rats: a concerted action of NAD(P)H oxidase and COX-2. Toxicology 390:10–21. https://doi.org/10.1016/j.tox.2017.08.004
Liu Z, He C, Chen M, Yang S, Li J, Lin Y, Deng Y, Li N, Guo Y, Yu P, Li X (2018) The effects of lead and aluminum exposure on congenital heart disease and the mechanism of oxidative stress. Reprod Toxicol 81:93–98. https://doi.org/10.1016/j.reprotox.2018.07.081
Novaes RD, Mouro VGS, Gonçalves RV, Mendonça AAS, Santos EC, Fialho MCQ, Machado-Neves M (2018) Aluminum: a potentially toxic metal with dose-dependent effects on cardiac bioaccumulation, mineral distribution, DNA oxidation and microstructural remodeling. Environ Pollut 242:814–826. https://doi.org/10.1016/j.envpol.2018.07.034
Yu H, Zhang J, Ji Q, Yu K, Wang P, Song M, Cao Z, Zhang X, Li Y (2019) Melatonin alleviates aluminium chloride-induced immunotoxicity by inhibiting oxidative stress and apoptosis associated with the activation of Nrf2 signaling pathway. Ecotoxicol Environ Saf 173:131–141. https://doi.org/10.1016/j.ecoenv.2019.01.095
Xu J, Lin C, Wang T, Zhang P, Liu Z, Lu C (2018) Ergosterol attenuates LPS-induced myocardial injury by modulating oxidative stress and apoptosis in rats. Cell Physiol Biochem 48(2):583–592. https://doi.org/10.1159/000491887
Sergio L, Thomé AMC, Trajano L, Mencalha AL (2018) Photobiomodulation prevents DNA fragmentation of alveolar epithelial cells and alters the mRNA levels of caspase 3 and Bcl-2 genes in acute lung injury. Photochem Photobiol Sci 17(7):975–983. https://doi.org/10.1039/c8pp00109j
Li Z, Zhao G, Qian S, Yang Z, Chen X, Chen J, Cai C, Liang X, Guo J (2012) Cerebrovascular protection of β-asarone in Alzheimer’s disease rats: a behavioral, cerebral blood flow, biochemical and genic study. J Ethnopharmacol 144(2):305–312. https://doi.org/10.1016/j.jep.2012.09.013
Cheraghi EC, Golkar A, Roshanaei K, Alani B (2017) Aluminium-induced oxidative stress, apoptosis and alterations in testicular tissue and sperm quality in Wistar rats: ameliorative effects of curcumin. Int J Fertil Steril 11(3):166–175. https://doi.org/10.22074/ijfs.2017.4859
Ghorbel I, Khemakhem M, Boudawara O, Marrekchi R, Jamoussi K, Ben Amar R, Boudawara T, Zeghal N, Grati Kamoun N (2015) Effects of dietary extra virgin olive oil and its fractions on antioxidant status and DNA damage in the heart of rats co-exposed to aluminum and acrylamide. Food Funct 6(9):3098–3108. https://doi.org/10.1039/c5fo00342c
Jeong CH, Kwon HC, Kim DH, Cheng WN, Kang S, Shin DM, Yune JH, Yoon JE, Chang YH, Sohn H, Han SG (2020) Effects of aluminum on the integrity of the intestinal epithelium: an in vitro and in vivo study. Environ Health Perspect 128(1):17013. https://doi.org/10.1289/EHP5701
Oztürk B, Ozdemir S (2015) Effects of aluminum chloride on some trace elements and erythrocyte osmotic fragility in rats. Toxicol Ind Health 31(12):1069–1077. https://doi.org/10.1177/0748233713486956
Willhite CC, Ball GL, McLellan CJ (2012) Total allowable concentrations of monomeric inorganic aluminum and hydrated aluminum silicates in drinking water. Crit Rev Toxicol 42(5):358–442. https://doi.org/10.3109/10408444.2012.674101
Galal SM, Hasan HF, Abdel-Rafei MK, El Kiki SM (2019) Synergistic effect of cranberry extract and losartan against aluminium chloride-induced hepatorenal damage associated cardiomyopathy in rats. Arch Physiol Biochem 125(4):357–366. https://doi.org/10.1080/13813455.2018.1465437
Sun X, Sun H, Yu K, Wang Z, Liu Y, Liu K, Zhu Y, Li Y (2018) Aluminum chloride causes the dysfunction of testes through inhibiting the ATPase enzyme activities and gonadotropin receptor expression in rats. Biol Trace Elem Res 183(2):296–304. https://doi.org/10.1007/s12011-017-1120-1
Martínez-Martínez S, Lozano-Vidal N, López-Maderuelo MD, Jiménez-Borreguero LJ, Armesilla ÁL, Redondo JM (2019) Cardiomyocyte calcineurin is required for the onset and progression of cardiac hypertrophy and fibrosis in adult mice. Febs j 286(1):46–65. https://doi.org/10.1111/febs.14718
Saleem N, Prasad A, Goswami SK (2018) Apocynin prevents isoproterenol-induced cardiac hypertrophy in rat. Mol Cell Biochem 445(1-2):79–88. https://doi.org/10.1007/s11010-017-3253-0
Ge J, Yu H, Li J, Lian Z, Zhang H, Fang H, Qian L (2017) Assessment of aflatoxin B1 myocardial toxicity in rats: mitochondrial damage and cellular apoptosis in cardiomyocytes induced by aflatoxin B1. J Int Med Res 45(3):1015–1023. https://doi.org/10.1177/0300060517706579
Lu Y, Feng Y, Liu D, Zhang Z, Gao K, Zhang W, Tang H (2018) Thymoquinone attenuates myocardial ischemia/reperfusion injury through activation of SIRT1 signaling. Cell Physiol Biochem 47(3):1193–1206. https://doi.org/10.1159/000490216
Kumar S, Wang G, Liu W, Ding W, Dong M, Zheng N, Ye H, Liu J (2018) Hypoxia-induced mitogenic factor promotes cardiac hypertrophy via calcium-dependent and hypoxia-inducible factor-1α mechanisms. Hypertension 72(2):331–342. https://doi.org/10.1161/HYPERTENSIONAHA.118.10845
Brasil GA, Silva-Cutini MA, Moraes FSA, Pereira TMC, Vasquez EC, Lenz D, Bissoli NS, Endringer DC, de Lima EM, Biancardi VC, Maia JF, de Andrade TU (2018) The benefits of soluble non-bacterial fraction of kefir on blood pressure and cardiac hypertrophy in hypertensive rats are mediated by an increase in baroreflex sensitivity and decrease in angiotensin-converting enzyme activity. Nutrition 51-52:66–72. https://doi.org/51-52:66-72. https://doi.org/10.1016/j.nut.2017.12.007
Haghi Aminjan H, Abtahi SR, Hazrati E, Chamanara M, Jalili M, Paknejad B (2019) Targeting of oxidative stress and inflammation through ROS/NF-kappaB pathway in phosphine-induced hepatotoxicity mitigation. Life Sci 232:116607. https://doi.org/10.1016/j.lfs.2019.116607
Ghorbel I, Maktouf S, Kallel C, Ellouze Chaabouni S, Boudawara T, Zeghal N (2015) Disruption of erythrocyte antioxidant defense system, hematological parameters, induction of pro-inflammatory cytokines and DNA damage in liver of co-exposed rats to aluminium and acrylamide. Chem Biol Interact 236:31–40. https://doi.org/10.1016/j.cbi.2015.04.020
Zhang M, Zhu J, Qin X, Zhou M, Zhang X, Gao Y, Zhang T, Xiao D, Cui W, Cai X (2019) Cardioprotection of tetrahedral DNA nanostructures in myocardial ischemia-reperfusion injury. ACS Appl Mater Interfaces 11(34):30631–30639. https://doi.org/10.1021/acsami.9b10645
Onaolapo AY, Ayeni OJ, Ogundeji MO, Ajao A, Onaolapo OJ, Owolabi AR (2019) Subchronic ketamine alters behaviour, metabolic indices and brain morphology in adolescent rats: involvement of oxidative stress, glutamate toxicity and caspase-3-mediated apoptosis. J Chem Neuroanat 96:22–33. https://doi.org/10.1016/j.jchemneu.2018.12.002
Wallach D, Kang TB, Dillon CP, Green DR (2016) Programmed necrosis in inflammation: toward identification of the effector molecules. Science 352(6281):aaf2154. https://doi.org/10.1126/science.aaf2154
Martínez MA, Ares I, Rodríguez JL, Martínez M, Roura-Martínez D, Castellano V, Lopez-Torres B, Martínez-Larrañaga MR, Anadón A (2018) Pyrethroid insecticide lambda-cyhalothrin induces hepatic cytochrome P450 enzymes, oxidative stress and apoptosis in rats. Sci Total Environ 631-632:1371–1382. https://doi.org/10.1016/j.scitotenv.2018.03.030
Wang GX, Tu HC, Dong Y, Skanderup AJ, Wang Y, Takeda S, Ganesan YT, Han S, Liu H, Hsieh JJ, Cheng EH (2017) ΔNp63 inhibits oxidative stress-induced cell death, including ferroptosis, and cooperates with the BCL-2 family to promote clonogenic survival. Cell Rep 21(10):2926–2939. https://doi.org/10.1016/j.celrep.2017.11.030
Choudhary GS, Al-Harbi S, Almasan A (2015) Caspase-3 activation is a critical determinant of genotoxic stress-induced apoptosis. Methods Mol Biol 1219:1–9. https://doi.org/10.1007/978-1-4939-1661-0_1
Baghaei A, Solgi R, Jafari A, Abdolghaffari AH, Golaghaei A, Asghari MH, Baeeri M, Ostad SN, Sharifzadeh M, Abdollahi M (2016) Molecular and biochemical evidence on the protection of cardiomyocytes from phosphine-induced oxidative stress, mitochondrial dysfunction and apoptosis by acetyl-L-carnitine. Environ Toxicol Pharmacol 42:30–37. https://doi.org/10.1016/j.etap.2015.12.019
Funding
This work was co-financed by the Guangxi Science Natural Foundation for Young Scientists (No. 2019JJB140036), the Middle-aged and Young Teachers’ Basic Ability Promotion Project of Guangxi (No. 2019KY0556), and the First Batch of High-level Talent Scientific Research Projects of the Affiliated Hospital of Youjiang Medical University for Nationalities in 2019 (No. R20196314).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Zhou, L., He, M., Li, X. et al. Molecular Mechanism of Aluminum-Induced Oxidative Damage and Apoptosis in Rat Cardiomyocytes. Biol Trace Elem Res 200, 308–317 (2022). https://doi.org/10.1007/s12011-021-02646-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12011-021-02646-w