
Abstract
Seeds of an exquisite orchid, Rhynchostylis retusa, germinated in vitro on ½ Murashige and Skoog (MS) medium supplemented with different concentrations of coconut milk (CM). Of the different concentrations of CM employed for seed germination, 15% gave optimum response. On this medium a maximum of 93% cultures produced seedlings 90 days after inoculation. Individual seedlings with a length of about 0.5 cm were subcultured on MS medium supplemented with various concentrations of 6-benzylaminopurine (BA) and α-naphthalene acetic acid (NAA), with or without activated charcoal (AC), for further growth. Seedling growth was maximum on MS medium supplemented with 6 μM BA, 0.2 μM NAA, and 1 g L−1 AC. Here a maximum seedling length of 2.3 cm was observed after 1 month of culture. The seedlings were subcultured on MS medium supplemented with kinetin (Kn) or thidiazuron (TDZ), in the presence or absence of AC, for multiple shoot induction. A maximum multiple shoot number of 8.2 was observed on MS medium supplemented with 2 μM TDZ in the presence of AC. The shoots were rooted on ½ MS medium supplemented with 2 μM indole-3-butyric acid (IBA) and successfully transplanted to soil. Of the 45 plantlets transferred to soil 40 survived. The reproducible protocol standardized here will enable rapid propagation and conservation of this precious orchid.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Orchidaceae represents approximately 10% of angiosperms. It is one of the largest, most diverse, and most important categories of botanically and commercially significant flowering plants with 20,000–30,000 species (Cronquist 1981; Dressler 1981). Nearly 1,300 species are estimated to occur in India. The floral characteristics of orchids cover an exceptionally wide range of different shape, form, size, and colouration, surpassing flowers of all the other angiosperms. As a result, they are exceedingly valued in flori-trade as species or hybrids for pot plant or cut flowers, and hold enormous promise.
Orchids are well known for their beauty and use (Griesbach 2003). Many orchids can survive only in a specific undisturbed microclimate and are botanically very interesting for their floral complexity, free gene flow across specific barriers, minute seeds with undifferentiated embryos, suppressed endosperm formation, long-lived pollen tubes, and dependence on a suitable mycorrhizal association for germination of seeds.
Rhynchostylis retusa Blume. is an epiphytic herbaceous orchid commonly seen in the forests of Western Ghats. Owing to the great specialization in structure, colour, and longevity of their flowers, most of the orchids have been over-exploited by horticulturists and traders. R. retusa has graceful, pendulous racemes as long as leaves, and medium sized flowers pale pink in colour. Normally R. retusa reproduces vegetatively by pseudobulbs. But this natural way of propagation in this plant is generally hampered by very slow division of the pseudobulbils, so the rate of production of young plants is time-consuming. Hence for conservation and commercial cultivation of this species, an efficient in-vitro propagation procedure is necessary. Orchid seeds are minute and have no endosperm, and in nature they must be symbiotic with some kinds of fungi in order to germinate. The technique of asymbiotic seed germination by in-vitro culture was first introduced by Knudson (1946). Since then, in-vitro seed germination protocols have been established for many orchid species, and a number of media, salts, and plant-growth regulators have been employed for several orchid species (Arditti 1977; reviewed by Arditti 1992). Malmgren (1996) suggested use of simple basal medium supplemented with complex organic materials such as coconut milk (CM), pineapple juice, boiled potato, or other similar substances for better seed germination.
Formation of orchid seedlings from the seed involves three successive phases—germination, formation of protocorm like bodies (PLBs), and seedling development. Germination of orchid seed is begun by passage of water through the testa of the seed. Early germination is recognised by observing the colour and shape of the seed. The first visible signs of germination are swelling of the embryo followed by turning green and emergence of the breasted seed coats. This is called the spherical stage, and subsequently develops into the protocorm stage and later into the swelling stage. The formation of rhizoids in the protocorm is a unique feature of Orchidaceae. Subsequent cell division occurs in the apical and basal regions of the protocorm and in leaf primordia; morphogenesis is started from the shoot meristem. Later, the first roots are formed endogenously (Mitra et al. 1976).
In R. retusa in-vitro propagation through leaf and root explants has been reported (Vij et al. 1984, 1987). However in-vitro propagation through asymbiotic seed germination has been considered as an alternate method of rapid propagation of R. retusa. Hence this study was carried out with the objective of standardizing the concentration of plant-growth regulators in culture medium for asymbiotic development of seedlings, multiple shoot induction from seedlings, rooting, field transfer, and acclimatization of R. retusa plantlets to greenhouse conditions. In this study immature seeds were used for germination and subsequent proliferation. The advantage of using immature seeds is that they can be harvested before dormancy has become established, and there is no question of treating the seeds for dormancy release as the embryo continues to grow during seed culture in vitro. However immature seeds need careful and timely harvesting and immediate culture of these seeds in a suitable medium (Steele 1996).
Materials and methods
Healthy plants of R. retusa were planted in pots and preserved for further experiments. Several flowers were pollinated and a pair of bipartite pollinia was picked and deposited on the stigma of a flower of a different plant during pollination. The pollinated plants were maintained in the botanical garden of St Thomas College, Pala, India.
Green fertilized and well-developed capsules containing the seeds were collected from the healthy plants for seed germination experiments. Seeds of immature capsules were cultured immediately after collection. The immature capsules were collected 50 days after hand pollination and soaked in an aqueous solution of commercial detergent (Savlone, Qualigens, India) for 30 min. Solid dirt particles adhering to the surface of capsules were removed using a fine brush followed by rinsing with sterile distilled water. The capsules were surface-disinfected in 70% ethanol for 30 s, followed by treatment with 0.5% sodium hypochlorite (Hi Media, India) for 20 min and rinsed three times with sterile distilled water before air-drying in a laminar airflow cabinet for 5 min.
Green capsules were dissected longitudinally with a sterile surgical blade. The immature seeds were collected from the capsule with the help of a sterilized spatula and small a mass of the aggregated seeds was sown on culture medium in 22 × 120 mm glass test tubes containing half-strength Murashige and Skoog (1962; MS) basal medium supplemented with four different concentrations of CM (Sigma–Aldrich, USA) for in-vitro asymbiotic seed germination. The cultures were incubated for 3 months at 25 ± 1°C under cool white fluorescent tubes at irradiance of 38 μM m−2 s−1 with a 14 h photoperiod. The percentage of seed germination was scored after 90 days of culture. The 90-day-old seedlings were subcultured individually on MS medium supplemented with 2–6 μM 6-benzylaminopurine (BA) and 0.2–1 μM naphthalene acetic acid (NAA) alone or in combination with 1 g L−1 activated charcoal (AC; Sigma–Aldrich). For multiple shoot induction the seedlings were cultured on MS medium supplemented with various concentrations of Kn (0.5–6 μM) or TDZ (0.5–6 μM) with or without the presence of AC. The shoots were cultured on ½ MS medium supplemented with 0.5–6 μM NAA or 0.5–6 μM indole-3-butyric acid (IBA) for root induction.
Plantlets with more than two roots were removed from the culture tubes, washed with tap water to remove agar, and transplanted into plastic cups containing soil and wood charcoal (1:3). The young plants were kept under 30–40% natural light, sprayed with water twice a day, and fertilized with full-strength macroelements of the medium used in this study every 5–7 days. The acclimatized plants were eventually transferred to their natural habitat.
The basal medium used in this study was MS medium, 30 g L−1 sucrose, and 8 g L−1 agar. The pH of all media was adjusted to 5.8 prior to autoclaving at 121°C for 20 min. All the cultures were grown at 25 ± 2°C under a 16 h photoperiod supplied by two Philips TL 40 W fluorescent tubes. At least twenty-four cultures were raised for each treatment and all experiments were repeated three times. Analysis of variance and Duncan’s multiple range test was used for comparison among treatment means.
Results and discussion
The hand-pollinated orchid flowers were observed regularly and after 50 days the green, well-developed capsules were isolated from the field-grown plants. The seeds present in the fruits remained light green in colour at this time. The seeds were immediately inoculated on ½ MS medium supplemented with 5, 10, 15, and 20% CM. Visible sign of seed germination was observed after culture for two weeks. The germinating seeds increased in size, swelled, and later turned green. The seeds germinated and reached a length of about 0.5 cm after 90 days of culture (Fig. 1a). Even though there are a number of reports on orchid germination, scientists believe that orchid seed germination is a difficult job since it needs specific nutritional and environmental conditions (Arditti et al. 1990; Rasmussen 1995). Of the four different concentrations of CM used, 15% gave optimum response (Table 1). On this medium 93% cultures responded and the seedlings showed comparatively faster growth. It took only 25 days to develop the seed in to a green globular mass and 50 days to develop the first leaf on 15% CM-containing medium. CM-induced seed germination and protocorm and callus regeneration has been reported in several orchid species (Kitsaki et al. 2004; Roy and Banerjee 2003; Seeni and Latha 2000). The advantage of using seeds from immature capsules is that embryos become viable and easy to surface-sterilize for in-vitro seed germination (Van Waes and Debergh 1986; Yam and Weatherhead 1988; Mitchell 1989). The orchid seeds are very small and have a poor level of differentiation. Endosperm development does not occur and the embryo development stops at the globular stage. Since the endosperm is absent the seeds have limited food reserves, for example lipid droplets, and small amounts of proteins. Irrespective of these limitations the orchid seeds can germinate in vitro (Knudson 1946; Arditii 1967).

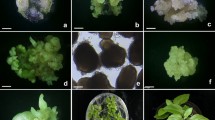
Asymbiotic seed germination and establishment of Rhynchostylis retusa plantlets in soil. (a) Asymbiotic germination of seeds from capsule after 90 days of culture on ½ MS medium supplemented with 15% coconut milk. (b) A seedling 1 month after growth on MS medium supplemented with BA (6 μM), NAA (0.2 μM), and activated charcoal (1 g L−1). The seedling has developed to a length of about 2.4 cm. (c) Multiple shoot formation from seedling on MS medium supplemented with TDZ (2 μM) without activated charcoal. Comparatively the shoot number and length is low. (d) Effect of activated charcoal on multiple shoot formation on MS medium supplemented with TDZ (4 μM). Here the number of multiple shoots has increased significantly. (e) A rooted seedling taken out of ½ MS medium supplemented with 2 μM IBA. Several roots have emerged from the basal region of the shoot. (f) Plants approximately 4 cm long 1 week after transfer to soil and wood charcoal (1:1). Scale bars represent 1 cm (a), 1.5 cm (b), 1 cm (c), 1 cm (d), 1 cm (e) and 2.5 cm (f)
Although seed germination takes place on ½ MS medium supplemented with various concentrations of CM, growth was slow. Hence to enhance further growth, the 90-day-old seedlings measuring a size of about 0.5 cm were transferred to MS medium supplemented with different concentrations of BA (2–6 μM) and NAA (0.2–1 μM) alone or in combination with AC (1 g L−1). The seedlings showed faster growth on MS medium supplemented with various concentrations of BA alone or in combination with NAA with or without the presence of AC. The synergistic effect of AC and phytohormones for asymbiotic germination of orchid seeds has been reported (Ernst 1974; Yam and Weatherhead 1988). However in Paphiopedilum ciliolare, AC is not necessary during the early stages of seedling growth but had a critical role in further development of the seedlings (Pierik et al. 1988).
The presence of AC in the medium significantly enhanced the pace of seedling growth, irrespective of BA concentrations used. However a combination of 6 μM BA and 0.2 μM NAA had a more critical role in seedling growth than all other treatments. On this medium the seedlings attained a maximum length of 2.4 cm in 1 month (Table 2; Fig. 1b). Usually an exogenous supply of growth regulators is necessary for in-vitro seed germination in some orchids (Kishor et al. 2006; Stewart and Kane 2006; Shimura and Koda 2004), because in some cases this is essential for lipid mobilization during germination of orchid seeds (Lo et al. 2004).
Seedlings measuring about 2.4 cm were used for inducing multiple shoots. The seedlings were cultured on MS medium supplemented with various concentrations of Kn (0.5–6 μM) or TDZ (0.5–6 μM) with or without the presence of AC. Here also the presence of AC in the medium resulted in a better response than medium containing plant-growth regulators alone (Table 3). In treatments without AC a low shoot number and slow growth were observed (Fig. 1c). Comparatively, TDZ gave superior response to Kn in terms of multiple shoot formation. In terms of percentage response 2 μM TDZ gave maximum response (88%); 4 μM gave optimum response in terms of number of shoots per explant (2.6 shoots per explant), however.
In AC-containing medium the seedlings showed more multiple shoot induction than in medium without AC. The maximum response was observed on MS medium supplemented with 4 μM TDZ. Here 100% cultures responded with an average number of 8.4 shoots per explant (Fig. 1d). The induced multiple shoots had an average length of 3.0 cm. In 2 μM TDZ-containing medium also 100% cultures responded with an average number of 7.6 shoots per explant and an average length of 3.4 cm per shoot. Promotion of orchid seedling growth by TDZ has been reported in Haemaria discolor (Shiau et al. 2005). In this report the H. discolor seedlings had greater height on MS medium supplemented with a combination of TDZ and NAA than all other treatments. Recently TDZ has been used in orchid tissue culture for various purposes due to its remarkable ability to induce callus or organogenesis. TDZ-induced callus formation from various explants occurred when TDZ was used along with either NAA or 2,4-D (Huan and Tanaka 2004; Teixeira da Silva et al. 2006). TDZ-induced organogenesis and somatic embryogenesis has been reported in several orchid species (Ernst 1994; Chen and Piluek 1995; Nayak et al. 1997; Chen and Chang 2001).
AC often promotes orchid shoot proliferation. AC has been reported to promote multiple shoot formation in nodal segments of A. formosanus (Ket et al. 2004). AC has beneficial and harmful effects in culture medium, depending upon the medium, explant, and plant-growth regulators used. The beneficial effects of AC on tissue responses in vitro could be attributed to:
-
1.
providing a dark environment by darkening the medium (Dumas and Monteuuis 1995);
-
2.
adsorption of harmful substances produced by either the media or explant (Fridborg et al. 1987; Fridborg and Eriksson 1975);
-
3.
adsorption of plant-growth regulators and other organic compounds (Constantin et al. 1977; Nissen and Sutter 1990; Weatherhead et al. 1978); or
-
4.
release of substances naturally present in or adsorbed by AC (Ernst 1974; 1975; Johansson et al. 1990).
The multiple shoots were separated and shoots containing velamen roots were directly transferred to the field. Several velamen roots were induced from the basal region of seedlings (Fig. 1e). The roots remained yellowish and healthy. However, shoots which did not have roots were transferred to ½ MS medium supplemented with NAA (0.5–6.0 μM) or IBA (0.5–6.0 μM) and produced velamen roots. IBA was comparatively better than NAA both in terms of percentage response and average number of roots per shoot (Table 4). Maximum response was observed on ½ MS medium supplemented with 2 μM IBA. On this medium 100% of shoots produced roots, with an average number of 4.5 roots per shoots (Fig. 1e). The roots remained yellowish and healthy.
The rooted shoots were taken out from the medium and hardened in a greenhouse. Ninenty five percent of the rooted plantlets grew well after they were acclimatized and transferred to plastic cups containing garden soil and wood charcoal (Fig. 1f). The acclimatized plants showed morphological similarity to the parental plants. The technique standardized here could be useful in re-establishment of these plants in their natural habitat and cultivation for other useful purposes.
Abbreviations
- AC:
-
Activated charcoal
- BA:
-
6-Benzylaminopurine
- CM:
-
Coconut milk
- IBA:
-
Indole-3-butyric acid
- MS:
-
Murashige and Skoog
- NAA:
-
Naphthalene acetic acid
- TDZ:
-
Thidiazuron
References
Arditii J (1967) Factors affecting the germination of orchid seeds. Bot Rev 33:1–97
Arditti J (1977) Clonal propagation of orchids by means of tissue culture-a manual. In: Arditti J (ed) Orchid biology, vol. I, Cornell University Press, New York, pp 203–293
Arditti J (1992) Fundamental of orchids biology. Wiley, New York
Arditti J, Ernst R, Yam TW, Glabe C (1990) The contribution to orchid mycorrhizal fungi to seed germination: a speculative review. Lindleyana 5:249–255
Chen YQ, Piluek C (1995) Effects of thidiazuron and N6-benzylaminopurine on shoot regeneration of Phalaenopsis. Plant Growth Regul 16:99–101
Chen JT, Chang WC (2001) Effects of auxins and cytokinins on direct somatic embryogenesis on leaf explants of Oncidium ‘Gower Ramsey’. Plant Growth Regul 34:229–232
Constantin MJ, Henke RR, Mansur MA (1977) Effect of activated charcoal on callus growth and shoot organogenesis in tobacco. Vitro Cell Dev Biol Plant 13:293–287
Cronquist A (1981) An integrated system of classification of flowering plants. Columbia University Press, Ithaca
Dressler RL (1981) The orchids: natural history and classification. Harvard University Press, Cambridge
Dumas E, Monteuuis O (1995) In vitro rooting of micropropagated shoots from juvenile and mature Pinus pinaster explants—influence of activated charcoal. Plant Cell Tissue Organ Cult 40:231–235
Ernst R (1974) The use of activated charcoal in asymbiotic seedling culture of Paphiopedilum. Am Orchid Soc Bull 43:35–38
Ernst R (1975) Studies in asymbiotic culture of orchids. Am Orchid Soc Bull 44:12–18
Ernst R (1994) Effects of thidiazuron on in vitro propagation of Phalaenopsis and Doritaenopsis (Orchidaceae). Plant Cell Tissue Organ Cult 39:273–275
Fridborg G, Pedersen ML, Eriksson T (1987) The effect of activated charcoal on tissue cultures: adsorption of metabolites inhibiting morphogenesis. Physiol Plant 43:104–106
Fridborg G, Eriksson T (1975) Effects of activated charcoal on morphogenesis in plant tissue cultures. Physiol Plant 34:306–308
Griesbach RJ (2003) Orchids emerge as major world floral crop. Chron Horticult 43:6–12
Huan LVT, Tanaka M (2004) Callus induction from protocorm- like body segments and plant regeneration in Cymbidium (Orchidaceae). J Hortic Sci Biotechnol 79:406–410
Johansson L, Galleberg E, Gedin A (1990) Correlation between activated charcoal, Fe EDTA and other organic media ingredients in cultures of anthers of Anemone canadensis. Physiol Plant 80:243–249
Ket NV, Hahn EJ, Park SY, Chakrabarty D, Peak KY (2004) Micropropagation of an endangered orchid Anoectochilus formosanus Biol Plant 48:339–344
Kishor R, Sha Valli Khan P S, Sharma G J (2006) Hybridization and in vitro culture of an orchid hybrid Ascocenda ‘Kangla’. Sci Hortic 108:66–73
Kitsaki CK, Zygouraki S, Ziobora M, Kintzios S (2004) In vitro germination, protocorm formation and plantlet development of mature versus immature seeds from several Ophrys species (Orchidaceae). Plant Cell Rep 23:284–290
Knudson L (1946) A nutrient for germination of orchid seeds. Am Orchid Soc Bull 15:214–217
Lo SF, Nalawade SM, Kuo CL, Chen CL, Tsay HS (2004) Asymbiotic germination of immature seeds, plantlet development and ex vitro establishment of plants of Dendrobium tosaense Makino: a medicinally important orchid. In Vitro Cell Dev Biol Plant 40:528–535
Malmgren S (1996) Orchid propagation. Theory and practice. In: In: Allen C (eds) North American native terrestrial orchids. Propagation and production. North Am Native Terrestrial Orchid Conf Proc. C. Allen, Germantown, pp 63–71
Mitchell RB (1989) Growing hardy orchids from seeds at Kew. Plantsman 11:152–169
Mitra GC, Prasad RN, Choudhury AR (1976) Inorganic salts and differentiation of protocorm in seed callus of an orchid and correlated changes in its free amino acid content. Indian J Exp Biol 14:350–351
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15:473–497
Nayak NR, Patnaik S, Rath SP (1997) Direct shoot regeneration from foliar explants of an epiphytic orchid Acampe praemorsa (Roxb.) Blatter and McCann. Plant Cell Rep 16:583–586
Nissen SJ, Sutter EG (1990) Stability of IAA and IBA in nutrient medium to several tissue culture procedures. Hortic Sci 25:800–802
Pierik RLM, Sprenkels PA, Van der Harst B, Van der Meys QG (1988) Seed germination and further development of Paphiopedilum ciliolare Pfitz in vitro. Sci Hortic 34:139–153
Rasmussen HN (1995) Terrestrial orchids from seeds to mycotropic plants. Cambridge University Press, Cambridge
Roy J, Banerjee N (2003) Induction of callus and plant regeneration from shoot tip explants of Dendrobium fimbriatum Lindl. var. oculatum Hk.f. Sci Hortic 97:333–340
Seeni S, Latha PG (2000) In vitro multiplication and ecorehabilitation of the endangered Blue Vanda. Plant Cell Tissue Organ Cult 61:1–8
Shiau YJ, Nalawade SM, Hsai CN, Tsay HS (2005) Propagation of Haemaria discolor via in vitro seed germination. Biol Plant 49:341–346
Shimura H, Koda Y (2004) Micropropagation of Cypripedium macranthos var. rebunense through protocorm like bodies derived from mature seeds. Plant Cell Tissue organ Cult 78:273–276
Steele WK (1996) Large-scale seedling production of North American native terrestrial orchids. In: Allen C (eds) North American native terrestrial orchids. Propagation and production. North Am Native Terrestrial Orchid Conf Proc. C. Allen, Germantown, pp 11–26
Stewart and Kane (2006) Asymbiotic seed germination and in vitro seedling development of Habenaria macroceratitis (Orchidaceae), a rare Florida terrestrial orchid. Plant Cell Tissue Organ Cult 86:147–158
Teixeira da Silva JA, Singh N, Tanaka M (2006) Priming biotic factors for optimal protocorm-like body and callus induction in hybrid Cymbidium (Orchidaceae), and assessment of cytogenetic stability in regenerated plantlets. Plant Cell Tissue Organ Cult 84:135–144
Van Waes JM, Debergh PC (1986) In vitro germination of some Western European orchids. Physiol Plant 67:253–261
Vij SP, Sood A, Plaha KK (1984) Propagation of Rhynchostylis retusa BL. (Orchidaceae) by direct organogenesis from leaf segment cultures. Bot Gaz 145:210–214
Vij SP, Pathak P, Sharma M (1987) On the regeneration potential of Rhynchostylis retusa root segments: a study in vitro. J Orchid Soc Ind 1:71–74
Weatherhead MA, Burdon J, Henshaw GG (1978) Some effects of activated charcoal as an additive to plant tissue culture media. Z Pflanzenphysiol 89:141–147
Yam TW, Weatherhead MA (1988) Germination and seedling development of some Hong Kong orchids. Lindleyana 3:156–160
Acknowledgments
We thank the Principal, St Thomas College, Pala, India, for providing with necessary laboratory facilities. TDT acknowledges financial assistance from Department of Science and Technology (DST), Government of India in the form of a young scientist project (Project No. SR/FTP/LSA-06/2002).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Thomas, T.D., Michael, A. High-frequency plantlet regeneration and multiple shoot induction from cultured immature seeds of Rhynchostylis retusa Blume., an exquisite orchid. Plant Biotechnol Rep 1, 243–249 (2007). https://doi.org/10.1007/s11816-007-0038-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11816-007-0038-z