
Abstract
Somatic embryogenesis was achieved from cell suspension cultures of niger (Guizotia abyssinica Cass.). Initially, friable embryogenic calluses were induced from cotyledonary leaves of niger on Murashige and Skoog (MS) agar medium containing 5 μM 2,4-Dichlorophenoxyacetic acid (2,4-D) and 0.5 μM kinetin (KIN). Cell suspension cultures were established by using embryogenic calluses in MS liquid medium containing 5 μM 2,4-D and 0.5 μM KIN. Initiation of somatic embryogenesis and development up to globular stage from embryogenic cell clumps occurred in the liquid medium itself. Thereafter embryogenic cell aggregates were transferred to MS agar medium supplemented with 3 μM KIN for embryo differentiation, whereas maturation of somatic embryos occurred in MS agar medium containing 10 μM abscisic acid.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Niger (Guizotia abyssinica Cass., Asteraceae) is most important oil crop of Ethiopia, providing 50–60% of the country’s indigenous edible oil. It is also minor oil crop in India, Kenya, Uganda, Sudan, Malawi and other African countries. Beside edible uses, niger seed oil can be utilized for the manufacture of soaps, paints and lubricants, and the protein rich meal obtained after oil extraction is used as feed or for manure (Getinet and Sharma 1996). Niger plant has an extremely low harvest index. The self incompatibility nature of niger causes serious difficulty for inbred line development and maintenance (Getinet and Sharma 1996). In vitro technology could serve as an alternative means for genetic upgrading and its application largely depends on the reliable plant regeneration system. There have been several reports concerning the regeneration of niger plants from tissue cultures, but these reports were confined to organogenesis and embryogenesis (Nikam and Shitole 1993, 1997; Sarvesh et al. 1994; Jadimath et al. 1998; Ashok Kumar et al. 2000). However, there is no report on regeneration of plants from suspension cultures via somatic embryogenesis. The establishment of embryogenic suspension cultures has great potential to aid crop improvement and is suitable for in vitro selection of variants specially selection of salt tolerance, disease/toxin resistance, cold tolerance lines in crop plants (Dixon and Gonzales 1994) and genetic transformation (Finer and McMullen 1991). The aim of the present work was to establish cell suspension culture and plant regeneration in niger (G. abyssinica Cass.).
Materials and methods
Plant material
Seeds of niger var. Ootacamund were surface sterilized using 0.1% (w/v) mercuric chloride for 15 min followed by repeated washing with sterile distilled water. The seeds were germinated on half strength Murashige and Skoog (MS 1962) basal medium taken in 60 mm × 95 mm culture jars. The medium was supplemented with 2% (w/v) sucrose and solidified using 0.8% agar (w/v) (type-1, Hi-media Co., Mumbai, India). The pH of the all media was adjusted to 5.8 prior to autoclaving at 1.4 kg cm−2 for 20 min. The cultures were incubated at 25 ± 2°C under the white fluorescent light (40 μmol m−2 s−1) of 16-h photoperiod.
Callus induction
Cotyledonary leaves were excised from 7-days old seedlings, cut in 0.5 cm2 segments and cultured on 15 ml MS medium taken in 25 × 150 mm culture tubes. Medium was supplemented with 2% (w/v) sucrose, 5 μM 2,4-dichlorophenoxyacetic acid (2,4-D) and 0.5 μM kinetin (KIN), 0.8% (w/v) agar, for embryogenic callus induction (Ashok Kumar et al. 2000). The cultures were incubated at 25 ± 2°C under the white fluorescent light (40 μmol m−2 s−1) of 16-h photoperiod.
Cell suspension culture and induction of somatic embryogenesis
Embryogenic callus obtained from cotyledonary leaves were used to initiate suspension cultures. Embryogenic calluses were transferred to 100 ml Erlenmeyer flasks containing 30 ml of MS liquid medium supplemented with 5 μM 2,4-D plus 0.5 μM KIN and 2% (w/v) sucrose. The embryogenic cell suspensions were maintained by adding 5 ml of the old suspension to 25 ml of fresh medium at 2-week intervals. Cultures were agitated at 100 rpm on a gyratory shaker under at 25 ± 2°C under the white fluorescent light (40 μmol m−2 s−1) of 16-h photoperiod. After 2 weeks of culture, embryogenic cell clumps were filtered using a 200 μm stainless steel sieve to remove the large clumps and settled by centrifugation for 5 min at 1,000g. Two hundred milligram cells were transferred to MS liquid medium supplemented with 5 μM 2,4-D in combination with 0.5 μM or 2.0 μM KIN or 2 μM benzylaminopurine (BAP) and 2 % (w/v) sucrose in 100 ml Erlenmeyer flasks containing 30 ml MS medium. The growth rate of cells (cell density) was determined at weekly intervals for 8 weeks by measuring fresh weight after centrifuging the suspension at 1,000g. In the subsequent step cells clusters were sub-cultured to fresh MS liquid medium containing 5 μM 2,4-D and 0.5 μM KIN. After 2 weeks, the number of globular somatic embryos per cell clusters (aggregates) was determined.
Development and maturation of somatic embryos
Embryogenic suspension cell clusters (1–2 mm in size) were transferred to agar medium for further proliferation. The cell clusters were incubated onto MS medium taken in 25 × 150 mm culture tubes. The medium was supplemented 1, 3 and 5 μM of KIN or BAP, 2% (w/v) sucrose and 0.8% (w/v) agar. Torpedo-shaped embryos were subsequently transferred to MS agar medium containing 5, 10, 20 μM abscisic acid (ABA), 2% (w/v) sucrose and 0.8 % (w/v) agar. The cultures were incubated at 25 ± 2°C under the white fluorescent light (40 μmol m−2 s−1) of 16-h photoperiod.
Germination and plant regeneration
Cotyledonary-stage embryos were transferred to MS agar medium without growth regulators taken 250 ml Erlenmeyer flasks containing 50 ml of medium. Plants of about 7–10 cm in height were transferred to pots with a mixture of soil, sand and farmyard manure (1:2:2 v/v). Plants were reared in growth chamber conditions (25 ± 2°C, 16-h photoperiod, 80% relative humidity and irradiance of 50 μmol m−2 s−1) and after 2 weeks the plants were reared under green house.
Microscopic observations
Cell clusters were squashed by using 2% aceto-carmine stain for microscopic observation of embryo development. Slides were observed using Leica DMRB microscope.
Statistical analysis
For callus induction, at least 50 explants were used and each experiment was repeated three times. Samples of suspension cultures were taken randomly at the end of each subculture, and the number of embryos was counted using a microscope. Counts were made from 12 independent samples, and the percentage of embryos was calculated on the basis of the total number of cell clusters present in the field. A completely randomized design was used in all experiments and analysis of variance and mean separation were carried out using Duncan’s multiple range tests at P ≤ 0.05 levels.
Results
Callus induction and establishment of suspension cultures
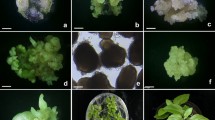
Cotyledonary leaf explants produced creamish-white callus (Fig. 1a) on 5 μM 2,4-D and 0.5 μM KIN supplemented medium within 6 weeks of culture. Friable calluses were initially transferred to MS liquid medium supplemented with 5 μM 2,4-D plus 0.5 μM KIN for cell dispersal. Subsequently, cell suspension cultures were established by transferring cells (200 mg) to MS liquid medium containing 5 μM 2,4-D plus 0.5-μM KIN; 5 μM 2,4-D plus 2.0 μM KIN and 5 μM 2,4-D plus 2.0 μM BAP. Suspended cells grew and multiply faster and reached their optimum concentration at the end of 4 weeks and become cell clusters (Fig. 2). Medium supplemented with 5 μM 2,4-D plus 0.5 μM KIN was found suitable for the accumulation of biomass.

Somatic embryogenesis from cell suspension cultures of niger. a Embryogenic callus (bar 12.7 mm), b globular-staged embryos (bar 2.05 mm), c globular-staged embryos (bar 1.5 mm), d heart-staged embryo (bar 1.5 mm), e torpedo-staged embryo (bar 1.5 mm), f cotyledonary-staged embryo (bar 1.5 mm), g plantlet regenerated from somatic embryo (bar 5.0 mm), h transplanted plant (bar 16.25 mm)

Growth curves of suspension cultured cells of niger (Guizotia abyssinica Cass.) cultured at different hormone combinations (5 μM 2,4-D in combination with 0.5 or 2.0 μM KIN or 2 μM benzylaminopurine (BAP) and 2 % (w/v) sucrose) in 100 ml Erlenmeyer flasks containing 30 ml MS liquid medium. Mean ± SE, n = 3
Development of somatic embryos
Embryogenic cells of the suspension culture underwent division and formed cell cluster. These cell clusters subsequently developed globular embryos upon subculture to the same media in another 2 weeks (Fig. 1b). Medium supplemented with 5 μM 2,4-D plus 0.5-μM KIN found superior for induction embryogenesis from the cell clusters (Fig. 3). Microscopic observation of suspension cluster revealed that initial spherical cells were embryogenic, containing viable dense cytoplasm, large nucleus with small vacuoles and deeply stained nucleus. These spherical cells were embryogenic and divided transversely resulting into two, four and subsequently to a group of cells, that was considered as proembryo. The proembryo further divided and formed globular embryos (Fig. 1c).

Effect of different hormone combination on formation of globular embryos in suspension culture of niger (Guizotia abyssinica Cass.). Those marked with the same letters within the same parameter are not significantly different at P < 0.05 according to Duncan’s multiple range test, n = 12
Development and maturation of somatic embryos
Globular embryos upon subculture to MS agar medium containing BAP or KIN developed further into heart-shaped embryos (Fig. 1d) showing bipolar structure within 2 weeks. Among the varied concentrations of cytokinins tested for embryo differentiation medium amended with 3 μM KIN found superior (Fig. 4). Heart shaped embryos differentiated into torpedo stage (Fig. 1e) with in another week. Cell clusters possessed all the stages of somatic embryos (globular, heart-shaped and torpedo-staged). Further developments of embryos was not achieved, however, cell clusters possessing torpedo shaped embryos, were developed into cotyledonary embryos within a week upon sub-culturing to MS agar medium containing ABA (Fig. 1f). MS medium supplemented 10 μM ABA was optimal for embryo maturation (Fig. 5) among the varied concentrations tested.

Influence of different cytokinin concentrations to niger (Guizotia abyssinica Cass.) on embryo differentiation on MS agar medium. Those marked with the same letters within the same parameter are not significantly different at P < 0.05 according to Duncan’s multiple range test, n = 12

Effect of ABA concentrations to niger (Guizotia abyssinica Cass.) on maturation of embryos on MS agar medium. Those marked with the same letters within the same parameter are not significantly different at P < 0.05 according to Duncan’s multiple range test, n = 12
Germination of embryos and plant regeneration
The cotyledonary staged embryos developed on ABA-amended medium, converted into plants within a week after transfer to MS agar medium without growth regulators (Fig. 1g). Converted plants from somatic embryos were hardened (Fig. 1h) and transplanted to soil. An average 80% survival was found after transplanting.
Discussion
The establishment of embryogenic suspension cultures for the regeneration of plants is ideal tool for the efficient in vitro selection and production of transgenic plants (Finer and McMullen 1991). Embryogenic suspension cultures have been established in only few crops, including sweetpotato (Liu et al. 2001), cowpea (Prem Anand et al. 2000), horsegram (Varisai Mohamed et al. 2004).
In the present study, embryogenic suspension cultures for regeneration of niger plant have been established for the first time. Ashok Kumar et al. (2000) reported development of somatic embryos from leaf explants of niger in MS medium containing 2,4-D in combination with BAP or KIN. Similar results were reported in the present study where globular embryos developed in MS liquid medium containing 2,4-D in combination with BAP or KIN, however, 5 μM 2,4-D plus 0.5-μM KIN was found best for the embryo induction from the suspended cell. Ontogeny of somatic embryo development has been studied in few crop plants. In Vigna species, Eapen and George (1990) traced the development of pro-embryos from two distinct cells (round/oval and elongated) forming filamentous structures, which in turn produced single or multiple embryos. The present study reveals the development of globular embryos through a division of spherical cells. In many plants including Cajanus cajan (Anbazhagan and Ganapathi 1999), Vigna unguiculata (Prem Anand et al. 2000) reported development (globular stage) and differentiation (heart and torpedo stages) of somatic embryos in MS liquid medium containing 2,4-D. Whereas, in the present studies globular embryos which were induced from the cell clusters on MS medium containing 2,4-D and KIN arrested at the same stage, they differentiated into heart and torpedo embryos transferring upon to medium supplemented with BAP or KIN. Similar to the present results cytokinins have been used to support the growth of somatic embryos in Acanthopanx koreanum (Choi et al. 1997).
Although somatic embryogenesis has been reported for several crop species, the quality of somatic embryos with regard to their germinability or conversion into plants has been generally very poor. This is because the apparently normal looking somatic embryos are actually incomplete in development (Bhojwani and Razdan 1996). ABA which prevents precocious germination and promotes normal development of embryos by suppression of secondary embryogenesis and pluricotyledony is reported (Ammirato 1983). In the present study, the torpedo-staged embryos matured into cotyledonary embryos on MS medium containing ABA. Our present results further indicated that ABA at 10 μM was optimal for the embryo maturation.
The embryogenesis protocol developed earlier by Ashok Kumar et al. (2000) in niger was on agar medium and it is having limitations for large scale culture. The results of present study reveals that once the embryogenic cell clusters was established in liquid medium, proliferation of embryogenic cell clumps actively occurred without loss of embryogenic competency through consecutive subcultures. In conclusion, a protocol for somatic embryogenesis was found to be reproducible from embryogenic suspension cultures of niger. This protocol has great potential for improvement of niger by biotechnological approaches such as in vitro selection, clonal propagation, genetic transformation and cell-induced mutation in niger.
Abbreviations
- ABA:
-
Abscisic acid
- BAP:
-
Benzylaminopurine
- KIN:
-
Kinetin
- MS:
-
Murashige and Skoog
- 2,4-D:
-
2,4-Dichlorophenoxyacetic acid
References
Ammirato PV (1983) The regulation of somatic embryo development in plant cell cultures: suspension culture techniques and hormone requirements. Biotechnology 1:68–74
Anbazhagan VR, Ganapathi A (1999) Somatic embryogenesis in cell suspensions of pigeonpea (Cajanus cajan L.). Plant Cell Tissue Organ Cult 56:179–184
Ashok Kumar HG, Murthy HN, Jadimath VG, Sheelavantmath SS, Pyati AN, Ravishankar BV (2000) Direct somatic embryogenesis and plantlet regeneration from leaf explants of niger, Guizotia abyssinica (L. f.) Cass. Indian J Exp Biol 38:1073–1075
Bhojwani SS, Razdan MK (1996) Plant tissue culture: theory and practical, a revised edition. Elsevier science, Netherlands, pp 148–150
Choi YE, Kim JW, Soh WY (1997) Somatic embryogenesis and plant regeneration from suspension cultures of Acanthopanx koreanum Nakai. Plant Cell Rep 17:84–88
Dixon RA, Gonzales RA (1994) Plant cell culture: a practical approach, 2nd edn. Oxford University Press, New York, pp 87–97
Eapen S, George L (1990) Ontogeny of somatic embryos of Vigna aconitifolia, Vigna mungo and Vigna radiata. Ann Bot 66:219–226
Finer JJ, McMullen D (1991) Transformation of soybean via particle bombardment of embryogenic suspension culture tissue. In Vitro Cell Dev Biol Plant 27:175–182
Getinet A, Sharma SM (1996) Niger—Guizotia abyssinica (L.f.) Cass. Promoting the conservation and use of underutilized and neglected crops. Institute of Plant Genetics and Crop Plant Research, Gatersleben/International Plant Genetic Resources Institute, Rome, pp 1–59
Jadimath VG, Murthy HN, Pyati AN, Ashok Kumar HG, Ravishankar BV (1998) Plant regeneration from leaf cultures of Guizotia abyssinica (niger) and Guizotia scabra. Phytomorphology 48:131–134
Liu QC, Zhai H, Wang Y, Zhang DP (2001) Efficient plant regeneration from embryogenic suspension cultures of sweet potato. In Vitro Cell Dev Biol Plant 37:564–567
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Nikam TD, Shitole MG (1993) Regeneration of niger (Guizotia abyssinica Cass.) cv. Sahyadri from seedling explants. Plant Cell Tissue Organ Cult 32:345–349
Nikam TD, Shitole MG (1997) In vitro plant regeneration from callus of niger (Guizotia abyssinica Cass.) cv. Sahyadri. Plant Cell Rep 17:155–158
Prem Anand R, Ganapathi A, Anbazhagan VR, Vengadesan G, Selvaraj N (2000) High frequency plant regeneration via somatic embryogenesis in cell suspension cultures of cowpea. Vigna unguiculata (L.) Walp. In Vitro Cell Dev Biol Plant 36:475–480
Sarvesh A, Reddy TP, Kavi Kishor PB (1994) Somatic embryogenesis and organogenesis in Guizotia abyssinica. In Vitro Cell Dev Biol Plant 30:104–107
Varisai Mohamed S, Wang CS, Thiruvengadam M, Jayabalan N (2004) In vitro plant regeneration via somatic embryogenesis through cell suspension cultures of horsegram (Macrotyloma uniflorum (Lam.) Verdc.). In Vitro Cell Dev Biol Plant 40:284–289
Acknowledgments
The senior author H.N. Murthy is grateful to Council of Scientific and Industrial Research [No. 38(1098)/04/EMR-II] and University Grants Commission [(F. No. 33-188/2007 (SR)], New Delhi, India for financial support in the form of research projects.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Werbrouck.
Rights and permissions
About this article
Cite this article
Naik, P.M., Murthy, H.N. Somatic embryogenesis and plant regeneration from cell suspension culture of niger (Guizotia abyssinica Cass.). Acta Physiol Plant 32, 75–79 (2010). https://doi.org/10.1007/s11738-009-0380-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-009-0380-6