
Abstract
Whole common Kilka fish was hydrolyzed separately using four commercial enzymes, Alcalase, Neutrase, Protamex at 50 °C and Pepsin at 37 °C for 30, 60 and 90 min. Degree of hydrolysis, angiotensin-I-converting enzyme (ACE) inhibitory activity and antimicrobial activity of each hydrolysate against Gram-negative (Escherichia coli, Salmonella enteritidis) and Gram-positive (Staphylococcus aureus, Listeria innocua) bacteria were studied. Results showed that the degree of hydrolysis for all enzymes was in the range of 2.63–3.36%. Electrophoresis profiles of the Kilka protein hydrolysates showed that most of produced peptides were in the range of 30 D but Alcalase and Neutrase had a better performance in the production of low molecular weight peptides in the range of 10 D. This led to increase the antimicrobial activity against the examined bacteria at the concentration of 200 µg/mL peptide solution. The Neutrase enzyme produced hydrolysate with the highest ACE inhibitory activity (53% ± 1.8 at 500 µg/mL). Antimicrobial activity of Kilka protein hydrolysates using Protamex and Pepsin was lower than the others due to lack of considerable amount of small peptides. The current research has demonstrated that the peptides derived from the enzymatic hydrolysis of Kilka fish protein in optimum conditions are capable of being converted to antimicrobial and antihypertensive agents to be used in functional foods.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Fish and fishery products represent a valuable source of nutrients with vital role for diversified and healthy diets. Apart from being highly nutritious, underutilized fish proteins and also the waste generated from fish processing operations can be transformed into a valuable food products through enzymatic hydrolysis. Fish protein hydrolysate (FPH) can be used as food ingredients to improve the nutritional properties of formulated food products or as a source of bioactive peptides to confer a health benefit on the host [1]. FPH generated through enzymatic hydrolysis consists of peptide mixtures with various molecular weight. It is well known that peptides molecular weight are closely related to their functional properties [2]. They have a positive impact on the functioning or conditions of human beings, thereby improving their health [3]. Bioactive peptides exhibit various biological activities such as antioxidant, antihypertensive, antimicrobial, antiproliferative, anticoagulant, antiobesity, antidiabetic and calcium-binding activities [4, 5]. Such diverse functionality make them highly desirable in novel functional food applications [6]. Researchers have reported that small peptides (1–5 D) from fish protein hydrolysates have strong influence on angiotensin I-converting enzyme (ACE) inhibition [2]. ACE plays an important role in the regulation of blood pressure and hypertension [7] and the inhibition of ACE activity (> 50%) is a good target for antihypertension treatment. Thus, the separation of this fraction from the protein hydrolysate mixtures becomes a priority [2].
The increase in antibiotic-resistant pathogenic bacteria has stimulated the search for alternative antibacterial agents from natural sources. Based on these facts, Antimicrobial peptides are being developed to be used as a novel class of antimicrobial agents, which could be extended to various food, biomedical and health applications [8]. Antimicrobial peptides (AMPs) (1–50 amino acids) with cationic and hydrophobic properties are known to be potent defenses of the host organism, providing activity against a wide variety of pathogenic microorganisms such as Gram-negative and Gram-positive bacteria, fungi, viruses, and parasites [9]. The marine AMPs were found to be structurally different from their counterparts produced by terrestrial species [10]. They are structurally diverse, and they display a wide spectrum of anti-infective activities, a low bio-deposition rate in body tissues, and are highly specific to targets [11].
Kilka fish is the Russian name to cultriventris fish native to the Black, Azov and Caspian Seas. It is a member of the Herring family (Clupeidae) and consists of two species of Anchovy Kilka, C. engrauliformis (Borodin, 1904) and Bigeye Kilka, C. grimmi (Kessler, 1877) and subspecies. Common Kilka is C. cultriventris caspi (Svetovidov, 1941). Common Kilka inhabits shallow areas close to the Caspian Sea coast. Changing the catch has led to an increase in the harvest of this type of fish compared to previous years [12]. This species accounts for 97% of all Kilka fish in the southern coast of the Caspian Sea and is recognized as the predominant species of catch [13]. These fish are considered as the main components of the diet for sturgeon and salmon in the Caspian Sea [14]. Kilka fish has dark and sensitive muscles, a small size and high fat content. Due to these inappropriate features, consumers do not exhibit their highly interest to use it directly in their diet. This is why more than 95% of Kilka fish in Iran have been converted in to animal feed as a source of protein due to having lower price compared to other fish [15]. Hence, the utilization of Kilka fish to produce value added products for human diet is gaining importance [14]. An interesting alternative is to transform the whole fish proteins into biologically active peptides by protease treatments. To the best of our knowledge, no studies have been performed on the antibacterial activity and ACE inhibitory of Kilka fish protein hydrolysates. In this study, the antibacterial activity and ACE inhibitory of Kilka fish protein hydrolysates obtained by treatment with four commercial enzymes were evaluated. The molecular weights of peptides produced were also determined.
Materials and methods
Materials
Common Kilka was caught in spring 2016 along the Caspian Sea coast and was brought to the laboratory in iced condition. Once received in the laboratory, the whole fish was minced twice in a kitchen blender at medium speed (5 mm plate size), then pooled, divided into plastic containers and kept frozen at − 20° C until the day of experiment.
ACE (EC 3.4.15.1) from rabbit lung and hippuryl–histidyl–leucine (HHL) as a substrate of ACE were purchased from Sigma Chemical Co. (St. Louis, MO). Commercial proteases (Alcalase, Neutrase, Pepsin and Protamex) were obtained from Novozymes (Bagsvaerd, Denmark). Details of the proteolytic enzymes used in this study are given in Table 1.
Ultrafiltration membrane (centriprep-10) was purchased from Amicon (Amicon Inc. Beverly, MA USA). All other reagents used in this study were analytical grade chemicals.
Four bacterial species were used to examine the antibacterial activity: Escherichia coli (ATCC 25922), Salmonella enteritidis (ATCC 13076), Listeria innocua (ATCC 33090) and Staphylococcus aureus (ATCC 25923), were obtained from American type culture collection.
Preparation of Kilka fish hydrolysates
Kilka fish hydrolysates were prepared according to the method described by Ovissipour et al. [14]. Briefly, for each batch, 50 g of minced fish was placed in a 250 mL Erlenmeyer flask, mixed with deionized water (1:2 w/v) and homogenized in a kitchen blender for about 1 min and the homogenate was used as substrate. Each enzyme was added separately to the homogenate at a level of 1% based on substrate protein concentration and mixed well. The optimum conditions used for hydrolysate preparation are presented in Table 1. All treatments were kept in a shaker incubator (Sartorius System, Germany) with constant agitation (150 rpm) for the period of 30, 60, and 90 min. The reaction was terminated by heating the solution to 95° C for 15 min to ensure enzyme inactivation. The hydrolysates were then cooled on ice and centrifuged at 8000×g for 20 min in refrigerated centrifuge (Sigma, USA) to collect the supernatant. The pH of the mixtures before and after hydrolysis was adjusted by adding 2 N NaOH and HCl. The FPH soluble fraction supernatant was freeze-dried for further analysis.
Determination of protein content
Total crude protein (N × 6.25) in the raw material and FPHs was determined using the Kjeldahl method [16].
Degree of hydrolysis (DH)
Degree of hydrolysis was estimated according to the method of Hoyle and Merritt [17]. Aliquots (10 mL) of enzyme digests of Kilka protein, obtained after 30, 60 and 90 min of hydrolysis, were fully mixed with the same volumes of 20% (w/v) TCA solution. The mixtures were placed at room temperature for 15 min, and then centrifuged at 7000×g for 10 min. The nitrogen contents of the supernatants were determined by conventional Kjeldahl method [16]. Soluble nitrogen in hydrolysates was determined using Bradford method [18]. The degree of hydrolysis was then computed as:
Analyses were performed in triplicate.
SDS-PAGE
Analysis of the molecular weight distribution of the different hydrolysates was carried out by tricine sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), according to Schagger and Von Jagow [19], using a 4% stacking gel and a 15% resolving gel. The freeze-dried hydrolysates were dissolved at a concentration of 20 mg/mL in the loading buffer (50 mM Tris–HCl, 4% SDS, 12% glycerol, 2% mercaptoethanol and 0.01% SERVA Blue G), heat-denatured at 90 °C for 5 min and then loaded. Protein bands were stained with Coomassie Brilliant Blue R250. The approximate molecular weight (MW) of the hydrolysates was determined using a pre-stained protein marker consisting of proteins between 3 and 100 D (Sigma Aldrich).
Ultrafiltration
The hydrolysate solutions were passed through ultrafiltration membrane with molecular weight cut-off (MWCO) of 10–30 D (Amicon Inc. Beverly, MA). The hydrolysate permeate fractions (10–30 D) were stored at − 20 °C until further use.
Bacteria and antibacterial assays
Four bacterial species were used as test microorganisms for determination of antibacterial activity: E. coli (ATCC 25922), S. enteritidis (ATCC 13076), L. innocua (ATCC 33090) and S. aureus (ATCC 25923). These bacterial species were maintained at − 24 °C in nutrient broth (NB) containing glycerol and were then transferred on nutrient agar medium at 37 °C for 24 h for reactivation. A pure colony was inoculated in NB and incubated at 37 °C and the density of the organism suspension was adjusted to equal 0.5 McFarland standard by adding sterile distilled water. Suspensions should contain between 107and 108 cfu/mL.
Minimum inhibitory concentration (MIC) and inhibitory concentration 50% (IC50)
The antimicrobial activity of the hydrolysates was determined using a liquid growth inhibition assay according to the procedures of NCCLS [20]. Briefly, a two-fold serial dilution of each hydrolysate solution in nutrient broth was made in flat-bottomed 96-well polystyrene microplates (Sorfa, China). Wells thus each contained 200 µL of dilution, were inoculated with 10 µL of overnight culture of the target bacteria diluted to a concentration of approximately 1.5 × 105 CFU/mL. Plates were incubated at 37 °C for 24 h and absorbance was measured at 630 nm using a microtiter plate reader (Stat fax, USA). MIC was calculated from the highest dilution showing complete inhibition of the tested strain. The experiment was repeated three times. For the blank sample, 100 µL of hydrolysate solution was mixed with 100 µL nutrient broth; for the positive control, 100 µL nutrient broth was used; and for the negative control, 100 µL of nutrient broth was mixed with 10 µL of bacterial suspension. Inhibitory concentration 50% was calculated using the following equation:
Determination of ACE inhibitory activity
The ACE inhibitory assay of Cushman and Cheung [21] was used with the slight modification. 50 µL of sample solution and 50 µL of ACE (1 U/mL) was pre-incubated for 10 min at 37 °C. The mixture was then mixed with 150 µL of substrate (8.3 mM HHL in 50 mM sodium borate buffer containing 0.3 M NaCl at pH 8.3) and incubated at 37 °C for 30 min. The reaction was stopped by adding 250 µL 1 N HCl. The hippuric acid (HA) was extracted with 500 µL of ethylacetate. After centrifugation (800xg, 15 min), 200 µL of the upper layer was removed by evaporation in a dry-oven at 80 °C. The residue was dissolved in 1 mL of distilled water, and the absorbance was measured at 228 nm. ACE-inhibitory activity was calculated using the equation:
where A is the absorbance of HA generated in the presence of ACE inhibitor and B is the absorbance of HA generated without ACE inhibitor.
Statistical analysis
The ANOVA and Duncan test at 5% probability were used to determine differences between the means of the degree of hydrolysis as well as the contents of peptides and free amino acids in the Kilka fish hydrolysates using PASW Statistics 16.0 software (SPSS, Chicago, IL, USA).
Results and discussion
Degree of hydrolysis
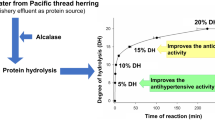
The degree of hydrolysis achieved by different enzyme treatments is shown in Fig. 1. After 90 min incubation, The DH was in a range of 2.5–3.5%, for whole Kilka fish individually hydrolyzed with Alcalase, Protamex, Neutrase and Pepsin. The effect of different enzymes on the hydrolysis process has been studied by many researchers. Similar levels of DH were previously reported for sardine (1.9–3.1%) and cuttlefish (3.2–7%) viscera hydrolysates [22]. As well as for Atlantic mackerel (Scomber scombrus) and its processing by-products [23]. However, several studies reported higher DH for their protein hydrolysates which reflect the complexity of this process. Significant differences (p < 0.05) was observed between the degree of hydrolysis of four examined enzymes. In the recent study on Kilka fish protein, the highest DHs of 3.36% in 30 min, 3.29%, 2.98 and 2.79 in 60 min of hydrolysis were obtained for Protamex, Pepsin, Alcalase and Neutrase respectively.

Degree of hydrolysis of Kilka fish protein hydrolysates prepared using different enzymes. Error bars represent standard deviations from triplicate determinations. Different letters on the error bar indicate that the results are significantly different (P < 0.05)
As expected, an analysis of Fig. 1 showed that the DH related to Alcalase and Pepsin hydrolysates increased with hydrolysis time but there was variation in DH with time for Neutrase and Protamex. Decrease of the DH was observed during the course of hydrolysis but in different period for each enzyme. Such behavior can be related to the reduction in peptide bonds capable of being cleaved, competition between the substrate and the hydrolysis products and enzyme denaturation that decreases its activity [24]. The DH is significantly dependent on the hydrolysis conditions such as incubation time, pH, temperature, as well as type and concentration of enzyme. Such variations were reported for fish proteins by many studies [23, 25, 26].
To explore the complexity of the effect of different type of enzyme on the degree of hydrolysis, the nature and behavior of two commercial enzymes applied in this study are discussed. Alcalase is an endopeptidase and has a range of specificity of peptide bonds for hydrolysis. On the other hand, Neutrase is a mixture of an endoprotease and exopeptidase, produced by Aspergillus oryzae, giving it a broader range of action and thus a higher DH is expected when compared with hydrolysis by Alcalase [27]. However, this trend was not seen in all reported studies. The nature of protein is also considered as a vital parameter. Vaštag et al. [28] reported that the limited hydrolysis (lower DH) and the presence of both high and low molecular weight bands in the electrophoretic profile could indicate that the effectiveness of Neutrase towards protein was due to exopeptidase activity.
SDS-PAGE analysis
To study the effect of different enzymes on protein breakdown in Kilka fish protein, SDS-PAGE was performed to obtain information on their molecular weight (Fig. 2). The results indicated that the protein bands of hydrolyzed samples have molecular weights ranging from ~ 10 to 30 D and above. However, most molecular weight of protein bands were observed in the range of ~ 30 D. The higher molecular weight of peptide bands belonged to Pepsin and Protamex hydrolysates were in the range of ~ 30 D. The molecular weight of Alcalase and Neutrase hydrolysates were in the range of ~ 10 D. The molecular size distribution of the Alcalase and Neutrase hydrolysates in our study was similar and different from Pepsin and Protamex hydrolysates.

SDS-PAGE profile of obtained Kilka fish protein hydrolysates with different enzymes. Lane 1, molecular weight markers; lane 2, for Neutrase at 60 min; lane 3, Neutrase at 30 min; lane 4, Alcalase at 60 min; lane 5, Alcalase at 30 min; lane 6, molecular weight markers; lane 7, Protamex at 90 min; lane 8, Pepsin at 90 min; lane 9, Neutrase at 90 min; lane 10, Alcalase at 90 min; lane 11, Protamex at 60 min; lane 12, Protamex at 30 min; lane 13, molecular weight markers; lane 14, Pepsin at 60 min; lane 15, Pepsin at 30 min
Alcalase and Neutrase are obviously the most efficient enzymes when it comes to degrading the larger proteins of the substrate mixture, as can be seen from the lack of strong bands in the high molecular weight area in lane 2, 3, 4, 9 and 10 in Fig. 2. The varying molecular weight (MW) profiles revealed considerable differences in the degree of protein breakdown depending on the enzyme used [29]. Based on the electrophoretic profile, the Alcalase and Neutrase hydrolysates showed higher levels of peptides with MW below 30 D, whereas in Pepsin and Protamex hydrolysates, peptides with MW above 30 D were predominant.
Antimicrobial assays
The results of antimicrobial activity of FPHs prepared by different enzymes are presented in Tables 2, 3, 4 and 5. The hydrolysates were ultra-filtered using 10–30 D MWCO membranes and were evaluated for their inhibitory activity against different bacterial strains with 200 µg/mL. The results showed that only Alcalase hydrolysate at 90 min (200 µg/mL) had complete growth inhibition against four examined bacterial strains E. coli, S. aureus, S. enteritidis and L. innocua. Neutrase hydrolysate at 90 min exhibited the highest growth inhibition against S. aureus. Partial antibacterial activity was detected with Pepsin and Protamex hydrolysates obtained in 30, 60 and 90 min. S. aureus ATCC25923 was the most sensitive strain and was more inhibited with hydrolysate samples at 200 µg/mL.
Alcalase and Neutrase hydrolysates composed of low molecular weight (~ 10 D) exhibited higher growth inhibition against E. coli, S. aureus, S. enteritidis than Pepsin and Protamex hydrolysates. The low inhibitory activity of protamox and Pepsin hydrolysates against examined bacteria might be due to the insufficient amount of low molecular weight peptides generated by such enzymes in the defined conditions.
It is well known that the presence of small peptides is responsible for high inhibitory activity in fish protein hydrolysates [8, 23, 30]. Peptides from fish hydrolysates having low molecular weights were shown to possess many biological activities [31]. When the antimicrobial activity of the peptide fractions is concerned, several factors such as amino acid composition, sequence, molecular weight, structural features like peptide helicity, hydrophobicity, hydrophobic moment, charge and the type of bacteria need to be taken into account [32, 33]. The mode of action and the specificity of the antibacterial compounds are closely related to the differences existing in the membrane composition of the target microorganism [34]. Moreover, the electrostatic interaction between the peptide and positively charged and anionic lipids on the surface of the microorganism, as well as the hydrophobicity and flexibility of the peptides are essential for peptides to exert their antimicrobial activity [35].
In this study, peptide fractions obtained from hydrolysates were effective against Gram-positive (S. aureus) and Gram-negative (E. coli and S. enteritidis) strains. The proteases used for hydrolysis determine the size and sequence of the resulting peptide and can therefore affect their activity. Our results suggest that the hydrolysates from whole Kilka fish protein might be used as potential source of novel antimicrobial agents for food and health applications. Further characterization of bioactive peptides reported in this study should be performed.
ACE inhibitory activity of Kilka fish protein hydrolysates
The ACE has been associated with the renin-angiotensin system, which regulates peripheral blood pressure. The hydrolysates obtained after enzyme hydrolysis were tested for ACE inhibitory activity (Table 6). The Alcalase and Neutrase hydrolysates exhibited higher ACE inhibition (with different DH values) in comparison with Pepsin and Protamex. The ACE inhibitory activities of the soluble peptide fractions produced by Alcalase, Neutrase, Pepsin and Protamex varied in terms of the molecular weight. This can be attributed to the impact of enzyme specificity, which is a key factor influencing both the characteristics of hydrolysates and thus the nature and composition of the peptides produced [36].
Neutrase hydrolysates (at 30 min) composed of low molecular weight peptides (~ 10 D) showed the highest ACE inhibitory activity among the tested hydrolysates at the concentration of 500 µg/mL. After 90 min and by increasing the degree of hydrolysis, the ACE inhibitory activity of the peptide solution (30–10 D) from the Neutrase hydrolysates decreased, which might due to the further hydrolysis of responsible peptides of the ACE inhibitory and production of peptides with less or no activity. Theodore and Kristinsson [37] found that the connection between ACE inactivation and degree of hydrolysis of catfish protein isolate may not be a simple and straightforward connection with peptide molecular size. Furthermore, these results suggest that only a short duration of hydrolysis was enough to get very high ACE inactivation.
The production of low molecular weight peptides in the Alcalase hydrolysates increased ACE inhibitory activity with increasing the degree of hydrolysis.
The Pepsin and Protamex hydrolysates with high molecular weight peptide (≥ 30 D) had no ACE inhibitory activity. The ability of low MW peptides to inhibit ACE activity has also been reported for peptides derived from a variety of fish species such as tilapia [7], yellowfin sole [38], Alaska pollock [39] and sea bream [40].
Although the structure–activity relationship of food-derived ACE inhibitory peptides has not yet been fully established, correlation among different peptide inhibitors of ACE indicated that binding to this enzyme is strongly influenced by the C-terminal tripeptide sequence of the substrate or inhibitor. ACE appears to prefer substrates or competitive inhibitors that mainly have hydrophobic amino acid residues at the three C-terminal positions [41]. Other studies demonstrated that Proline at the antepenultimate position in the peptide sequence also enhanced binding to the ACE enzyme [42]. The high ACE-inhibitory activity showed by Alcalase and Neutrase peptide fractions may be related to the high content of hydrophobic amino acids in their composition, as well as to a high content of proline.
Conclusions
Fish sources show promise as functional foods ingredients as they are valuable source of bioactive peptides encrypted within primary amino acid sequences and released upon enzymatic hydrolysis. In this study, we evaluated potentially bioactive protein hydrolysates with antibacterial and anti-ACE properties from whole Kilka fish protein by enzymatic hydrolysis using several commercially available enzymes. The results of this study demonstrated that the degree of hydrolysis and resulting peptide properties were affected by both digestion time and the type of commercial enzyme. Alcalase and Neutrase hydrolysates with low molecular weight (~ 10 D) have shown potent inhibitory activity against different bacterial strains and a potent ACE inhibitory activity agent.
Our results suggest that the hydrolysates from whole Kilka fish may be a potential source of antimicrobial and antihypertensive agents for food and health applications. Further research needs to be carried out to isolate, purify and characterize the peptides responsible for such functionalities of protein hydrolysates.
References
S. Saidi, A. Deratani, R.B. Amar, M.P. Belleville, Fractionation of a tuna dark muscle hydrolysate by a two-step membrane process. Sep. Purif. Technol. 108, 28–36 (2013)
J. Roslan, S.M.M. Kamal, K.F.M. Yunos, N. Abdullah, Assessment on multilayer ultrafiltration membrane for fractionation of tilapia by-product protein hydrolysate with angiotensin I-converting enzyme (ACE) inhibitory activity. Sep. Purif. Technol. 173, 250–257 (2017)
H. Korhonen, A. Pihlanto, Bioactive peptides: production and functionality. Int. Dairy J. 16(9), 945–960 (2006)
P.A. Harnedy, R.J. FitzGerald, Bioactive peptides from marine processing waste and shellfish: a review. J. Funct. Foods 4(1), 6–24 (2012)
D.H. Ngo, I. Wijesekara, T.S. Vo, Q. Van Ta, S.K. Kim, Marine food-derived functional ingredients as potential antioxidants in the food industry: an overview. Food Res. Int. 44(2), 523–529 (2011)
T. Rustad, M. Hayes, Marine bioactive peptides and protein hydrolysates: generation, isolation procedures, and biological and chemical characterizations, in Marine Bioactive Compounds, ed. by M. Hayes (Springer, Boston, 2012), pp. 99–113
S. Raghavan, H.G. Kristinsson, ACE-inhibitory activity of tilapia protein hydrolysates. Food Chem. 117(4), 582–588 (2009)
L. Beaulieu, J. Thibodeau, M. Desbiens, R. Saint-Louis, C. Zatylny-Gaudin, S. Thibault, Evidence of antibacterial activities in peptide fractions originating from snow crab (Chionoecetes opilio) by-products. Probiotics Antimicrob. Proteins 2(3), 197–209 (2010)
R.E. Hancock, H.G. Sahl, Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 24(12), 1551–1557 (2006)
M. Charlet, S. Chernysh, H. Philippe, C. Hetru, J.A. Hoffmann, P. Bulet, Innate immunity isolation of several cysteine-rich antimicrobial peptides from the blood of a mollusc, Mytilus edulis. J. Biol. Chem. 271(36), 21808–21813 (1996)
H.K. Kang, C.H. Seo, Y. Park, Marine peptides and their anti-infective activities. Mar. Drugs 13(1), 618–654 (2015)
M.R. Soleimani, S.F. Hosseini, M. Nikkhah, Evaluation of antioxidant activity of protein hydrolysate from common kilka (Clupeonella cultriventris caspia). J. Fish. Sci. Technol. 5(3), 95–108 (2016)
H. Fazli, Some environmental factors effects on species composition, catch and CPUE of kilkas in the Caspian Sea. ECOPERSIA 15(2), 157–164 (2011)
M. Ovissipour, B. Rasco, S.G. Shiroodi, M. Modanlow, S. Gholami, M. Nemati, Antioxidant activity of protein hydrolysates from whole anchovy sprat (Clupeonella engrauliformis) prepared using endogenous enzymes and commercial proteases. J. Sci. Food Agric. 93(7), 1718–1726 (2013)
B. Shabanpour, A. Shabani, S. Moini, M. Hamedi, M. Poorkabireh, The effect of different washing methods on chemical and gel forming properties of Kilka surimi. Pajouhesh and Sazandegi 81(4), 84–92 (2007). (in Persian)
AOAC International, W. Horwitz, G.W. Latimer, Official Methods of Analysis of AOAC International, 18th edn. (Mary land, Gaithersburg, 2005)
N.T. Hoyle, J.H. Merritt, Quality of fish protein hydrolysate from Herring (Clupea harengus). J. Food Sci. 59, 76–79 (1994)
M. Bradford, A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 72, 248–254 (1976)
H. Schägger, G. Von Jagow, Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 166(2), 368–379 (1987)
NCCLS. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically: approved standard, 5th edn., NCCLS document M7-A5, ISBN 1-56238-394-9. NCCLS, Pennsylvania, USA, 2000
D. Cushman, H. Cheung, Spectrophotometric assay and properties of the angiotensin converting enzyme of rabbit lung. Biochem. Pharmacol. 20(7), 1637–1648 (1971)
E.S. Kechaou, J. Dumay, C. Donnay-Moreno, P. Jaouen, J.P. Gouygou, J.P. Bergé, R.B. Amar, Enzymatic hydrolysis of cuttlefish (Sepia officinalis) and sardine (Sardina pilchardus) viscera using commercial proteases: effects on lipid distribution and amino acid composition. J. Biosci. Bioeng. 107(2), 158–164 (2009)
N. Ennaas, R. Hammami, L. Beaulieu, I. Fliss, Production of antibacterial fraction from Atlantic mackerel (Scomber scombrus) and its processing by-products using commercial enzymes. Food Bioprod. Process. 96, 145–153 (2015)
F. Guerard, L. Guimas, A. Binet, Production of tuna waste hydrolysates by a commercial neutral protease preparation. J. Mol. Catal. B 19, 489–498 (2002)
N. Bhaskar, T. Benila, C. Radha, R.G. Lalitha, Optimization of enzymatic hydrolysis of visceral waste proteins of Catla (Catla catla) for preparing protein hydrolysate using a commercial protease. Bioresour. Technol. 99(2), 335–343 (2008)
J. Dumay, C. Donnay-Moreno, G. Barnathan, P. Jaouen, J.P. Berge, Improvement of lipid and phospholipid recoveries from sardine (Sardina pilchardus) viscera using industrial proteases. Process Biochem. 41(11), 2327–2332 (2006)
J. Pedroche, M.M. Yust, J. Girón-Calle, M. Alaiz, F. Millán, J. Vioque, Utilisation of chickpea protein isolate for production of peptides with angiotensin I-converting enzyme (ACE)-inhibitory activity. J. Sci. Food Agric. 82(9), 960–965 (2002)
Ž. Vaštag, L. Popović, S. Popović, V. Krimer, D. Peričin, Production of enzymatic hydrolysates with antioxidant and angiotensin-I converting enzyme inhibitory activity from pumpkin oil cake protein isolate. Food Chem. 124(4), 1316–1321 (2011)
A. Alemán, E. Pérez-Santín, S. Bordenave-Juchereau, I. Arnaudin, M.C. Gómez-Guillén, P. Montero, Squid gelatin hydrolysates with antihypertensive, anticancer and antioxidant activity. Food Res. Int. 44(4), 1044–1051 (2011)
J.X. Huo, Z.H.A.O. Zheng, Study on enzymatic hydrolysis of Gadus morrhua skin collagen and molecular weight distribution of hydrolysates. Agric. Sci. China 8(6), 723–729. (2009)
S. He, C. Franco, W. Zhang, Functions, applications and production of protein hydrolysates from fish processing co-products (FPCP). Food Res. Int. 50(1), 28–297 (2013)
M. Dathe, T. Wieprecht, Structural features of helical antimicrobial peptides: their potential to modulate activity on model membranes and biological cells. Biochim. Biophys. Acta (BBA)-Biomembr. 1462(1), 71–87 (1999)
R. Di Bernardini, P. Harnedy, D. Bolton, J. Kerry, E. O’Neill, A.M. Mullen, M. Hayes, Antioxidant and antimicrobial peptidic hydrolysates from muscle protein sources and by-products. Food Chem. 124(4), 1296–1307 (2011)
R.E.W. Hancock, A. Patrzykat, Clinical development of cationic antimicrobial peptides: from natural to novel antibiotics. Curr. Drug Targets-Infect. Disord. 2(1), 79–83 (2002)
H. Jenssen, P. Hamill, R.E. Hancock, Peptide antimicrobial agents. Clin. Microbiol. Rev. 19(3), 491–511 (2006)
R. Balti, N. Nedjar-Arroume, A. Bougatef, D. Guillochon, M. Nasri, Three novel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttlefish (Sepia officinalis) using digestive proteases. Food Res. Int. 43(4), 1136–1143 (2010)
A.E. Theodore, H.G. Kristinsson, Angiotensin converting enzyme inhibition of fish protein hydrolysates prepared from alkaline-aided channel catfish protein isolate. J. Sci. Food Agric. 87(12), 2353–2357 (2007)
W.K. Jung, E. Mendis, J.Y. Je, P.J. Park, B.W. Son, H.C. Kim, Y.K. Choi, S.K. Kim, Angiotensin I-converting enzyme inhibitory peptide from yellowfin sole (Limanda aspera) frame protein and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 94(1), 26–32 (2006)
J.Y. Je, P.J. Park, J.Y. Kwon, S.K. Kim, A novel angiotensin I converting enzyme inhibitory peptide from Alaska pollack (Theragra chalcogramma) frame protein hydrolysate. J. Agric. Food Chem. 52(26), 7842–7845 (2004)
A. Fahmi, S. Morimura, H.C. Guo, T. Shigematsu, K. Kida, Y. Uemura, Production of angiotensin I converting enzyme inhibitory peptides from sea bream scales. Process Biochem. 39(10), 1195–1200 (2004)
B.A. Murray, R.J. FitzGerald, Angiotensin converting enzyme inhibitory peptides derived from food proteins: biochemistry, bioactivity and production. Current pharmaceutical design 13(8), 773–791 (2007)
M. del Mar Contreras, R. Carrón, M.J. Montero, M. Ramos, I. Recio, Novel casein-derived peptides with antihypertensive activity. Int. Dairy J. 19(10), 566–573 (2009)
Acknowledgements
This work was financially supported by Ferdowsi University of Mashhad-Research Affairs (contract No. 40890).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Rights and permissions
About this article

Cite this article
Qara, S., Habibi Najafi, M.B. Bioactive properties of Kilka (Clupeonella cultriventris caspi) fish protein hydrolysates. Food Measure 12, 2263–2270 (2018). https://doi.org/10.1007/s11694-018-9843-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11694-018-9843-z