
Abstract
Fluoride (F), anion of fluorine which is naturally present in soil and water, behaves as toxic inorganic pollutant even at lower concentration and needs immediate attention. Its interaction with flora, fauna and other forms of life, such as microbes, adversely affect various physiochemical parameters by interfering with several metabolic pathways. Conventional methods of F remediation are time-consuming, laborious and cost intensive, which renders them uneconomical for sustainable agriculture. The solution lies in cracking down this environmental contaminant by adopting economic, eco-friendly, cost-effective and modern technologies. Biological processes, viz. bioremediation involving the use of bacteria, fungi, algae and higher plants that holds promising alternative to manage F pollution, recover contaminated soil and improve vegetation. The efficiency of indigenous natural agents may be enhanced, improved and selected over the hazardous chemicals in sustainable agriculture. This review article emphasizes on various biological approaches for the remediation of F-contaminated environment, and exploring their potential applications in environmental clean-up. It further focuses on thorough systemic study of modern biotechnological approaches such as gene editing and gene manipulation techniques for enhancing the plant-microbe interactions for F degradation, drawing attention towards latest progresses in the field of microbial assisted treatment of F-contaminated ecosystems. Future research and understanding of the molecular mechanisms of F bioremediation would add on to the possibilities of the application of more competent strains showing striking results under diverse ecological conditions.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Fluoride (F) is a monoatomic anion of fluorine, which comes under halogen group of gases and accounts for 0.3 g kg−1 of the Earth’s crust. Being highly electronegative in nature, F exists only in combination with other elements to form compounds and minerals such as fluorspar, cryolite and fluorapatite (Ghosh et al. 2013). Fluoride contamination is a worldwide problem and is anthropogenically added into the environment through airborne as well as non-airborne sources. Airborne industrial sources include aluminium smelters, brick works and phosphate fertilizer factories, while non-airborne causes originates from spraying F-rich fertilizers in the fields and using contaminated water for irrigation (Ali et al. 2016). Fluoride has been reported to be endemic in 20 countries of the world (Messaitfa 2008). The Union Health and Family Welfare Ministry reported that 19 states of India have high F contamination in their ground water, amongst which Rajasthan occupied the topmost position (24 mg L−1) followed by Andhra Pradesh, Telangana, Uttar Pradesh, Karnataka, Madhya Pradesh, Haryana, Bihar, Chhattisgarh, Maharashtra, Assam, West Bengal and Gujarat (Ali et al. 2016).
Fluoride is considered to be an essential micronutrient for animals, plants and human beings; however, it has proved to exert deleterious effects if exists beyond the permitted values. The permissible amount of F in drinking water is set to 1.5 mg L−1 by the World Health Organization (Geneva, Switzerland), while 0.8–1.2 mg L−1 exerts the maximum benefit and minimum harmful effect (World Health Organisation 2008). Prolong exposure to F generates devastating effects in almost all live forms including humans. Cellular accumulation of F alters the sensitive biochemical processes by interfering with various enzymatic activities, directly or indirectly. It acts as a protoplasmic poison, which can cause various biochemical modifications including oxidative stress and DNA damage resulting into apoptosis and cell cycle alterations (He and Chen 2006; Chouhan et al. 2009). Fluoride may covalently bind to DNA, leading to DNA damage thereby initiating a series of events in the development of chemical carcinogenesis (Zhang et al. 2008). Excessive intake of F in human beings leads to serious health hazards like fluorosis, characterized by various deformities in teeth and skeleton such as dental mottling, osteoporosis and osteosclerosis (Davis 1980). Fluoride has also proved to be a phytotoxic element which interferes with various metabolic pathways in plants. It is absorbed through root system and transported via xylem to different parts, generating many adverse effects.
Fluoride uptake and transport in biological systems occur through the non-ionic diffusion of hydrogen fluoride (HF). The molecules of HF being highly permeant penetrate cell membranes much faster, resulting in a more pronounced intracellular intake and accumulation. The other routes of F intake in cells are considered to be the much demonstrated anion channels and co-transporters (Barbier et al. 2010). Fluoride also combines with other metals such as calcium, aluminium and beryllium to form inorganic complexes for easy penetration through cell membrane. However, the physiological and toxicological effects depend on extracellular metal concentration and the type of ion channels.
Fluoride at micromolar levels promotes cell proliferation, whereas at millimolar concentrations inhibit several enzymes, both in vivo and in vitro. The increasing load of F in the ecosystem produces significant amounts of toxicity which needs to be treated. The many approaches of bioremediation are gradually being accepted as the standard practice over conventional chemical and physical methods as they hold strong potential in removal of F even at low concentrations in a cost-effective and more efficient way.
Toxic effects of fluoride
In plants
Fluoride accumulation in plants is due to its absorption from the air, soil and water, which occurs in two ways: firstly, gaseous and particulate forms are actively absorbed by leaf stomata and to some extent by the cuticle of leaves and secondly, through passive absorption by roots (Baunthiyal and Ranghar 2013). Absorbed F can then be transported to the leaves by the symplastic or apoplastic pathways or via xylematic flow (Elloumi et al. 2005). The uptake is dependent upon several parameters such as soil pH, its activity and composition. A few plants are highly tolerant to F and hence can accumulate up to 4000 μg g−1 dry mass (DM) without showing any sign of toxicity. However, other plants show toxicity symptoms even at much lower concentration (< 20-μg g−1 DM). Normal concentration of F in plants, growing in non-polluted soil, is 0.1–15-mg kg−1 DM (Cooke et al. 1976). Plants like Avena sativa L., Acer pseudoplanatus L. and Camellia sinensis L. can accumulate more than the normal concentration of F. Table 1 displays the list of plants and their F accumulation capacity. Fluoride toxicity in plants occurs at three different levels: physiological, biochemical and molecular.
Physiological level
The initial symptoms of F injury are necrosis at the leaf tips, and margins which on prolonged exposure causes notched leaf and its falling-off. Chlorosis is also believed as a sensitive indicator of its toxicity. Severities of these symptoms are influenced by many environmental factors such as concentration of F, distance from source, length of exposure and meteorological conditions (Singh and Verma 2013). It is also reported to adversely affect plant growth and yield responses by causing reductions in growth parameters, i.e. germination percentage, lengths of root and shoot, number and size of leaves, number of flowers, fruit-set percentage and seed-set frequency (Singh et al. 2013).
Biochemical level
The high internal F concentration disturbs almost all the biochemical processes of plants. At micromolar concentration, it promotes cell proliferation, while at millimolar concentration disrupts, the activities of enzymes involved in several metabolic processes by binding to functional amino acid groups that surrounds their active centre and inhibition of protein secretion and synthesis.
Molecular level
Excess availability of F adversely affects to signalling pathways involved in cell proliferation and apoptosis, regulation of active oxygen species (AOS), patterns of gene expression, etc. (Barbier et al. 2010) (Fig. 1).

Molecular mechanisms of fluoride toxicity in plants
In humans and domestic animals
In humans, adequate amount of F is crucial for the integrity of bones and teeth. It improves bone density, enhances the remineralization of enamel and checks dental caries and osteoporosis (Ericsson 1970; Arlappa et al. 2013). On the contrary, excessive intake of F may cause dental fluorosis and other deformities which include long-term frequent joint pain, limited joint movement and limb motor dysfunction. It becomes more challenging for the people suffering from severe health problems like paralysis (Choubisa and Sompura 1996). Fluoride may extensively affect mammalian nervous system because it can penetrate the blood-brain barrier, and excessive ingestion of it can lead to brain-specific metabolic disorders due to the inhibition of several key neuronal enzymes and enhanced rate of lipid peroxidation (Shivarajashankara et al. 2002). Recent studies have shown that F can also be toxic to the immune system, including macrophages.
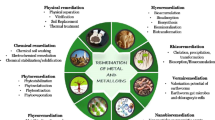
Fluoride remediation strategies
Fluoride is non-biodegradable in nature, therefore causes ecological destruction, gets accumulated in tissues and requires extensive measures for its clean-up (Rose and Marier 1977). Because it is poorly digested by the living organisms (Abdallah et al. 2006), F shows negative effects on plants, animals and human health after entering into the food chain (Stevens et al. 2000). The level of toxicity rendered by the F in different living forms demands effective ways for the ecological removal and remediation of this toxicant. Some of the known and adopted strategies of its removal includes physiochemical and biological approaches.
Physiochemical approaches
Several physiochemical procedures such as reverse osmosis (Sehn 2008), electrodialysis (Jayarathne et al. 2014), adsorption (Halder et al. 2015), ion exchange (Rangel et al. 2015), Nalgonda technique, biosorption (Mukherjee and Halder 2016), nanofiltration, etc., have previously been reported for F removal. Although, these methodologies have many advantages, they are allied with numerous negative side effects as well like high initial cost, water desalination, higher waste production, excessive electricity utilization, problem of waste discarding and inadequacy to remove all the pollutants (Gentil and Fick 2016). These problems necessitate to uncover the clean and cost-effective alternative approaches for F removal and remediation.
Biological approaches
Biological methods concerns with the involvement of living organisms like plants and microorganisms or their products for the treatment/removal of toxicants from the environment. The concept of bioremediation is largely exploited in different ways which helps in the conversion of organic contaminants into simpler forms such as carbon dioxide and water by the intensive action of plants/microorganisms as part of their metabolic processes (Sarkar et al. 2006). They have the potency to resist different pollutants by acquiring procedures like bioaccumulation, biotransformation, biosorption, etc. (Mukherjee et al. 2017). Researchers have reported both aerobic and anaerobic degradation pathways for the mineralization and stabilization of organic contaminants in the atmosphere. Biological treatments are more advantageous because of their cost-effectiveness, operational simplicity and less sludge production. The use of natural resources makes the process environment friendly and highly acceptable even for large-scale treatments (Doble and Kumar 2005).
Phytoremediation
It refers to promising green technology that utilizes plants to clean soil, air and water contaminated with hazardous materials (Kamran et al. 2014). It involves several processes like phytostabilization, phytovolatalization, phytoextraction, phytostimulation and phytodegradation for dealing with the pollutants, amongst which phytostabilization and phytoextraction have already been reported for F remediation.
Phytostabilization
It is mediated by plants that have ability to inhabit the contaminated soil. The direct action of root exudates immobilizes F by trapping in the soil matrices, thereby limiting its solubility. Also, the roots prevent the migration of F due to deflation and erosion by adsorption and precipitation, respectively, within the root zone (Pollard et al. 2014). Abdallah et al. (2006) studied the affinity of grapes to equilibrate F accumulation in its leaf margins by an equivalent Ca accumulation and suggested the entrapping of F as CaF2, which was not able to disturb plant metabolism. This study favoured the non-translocation of F through phloem towards lower plant organs.
Fluoride tolerant plants may have the ability to deactivate it better than the sensitive ones. Probable mechanisms includes shifting towards F insensitive metabolic pathways; complexation of F with organic compounds and their removal from the site of enzyme inhibition; reaction with cationic sites; sequestration in vacuoles and translocation to the leaf surfaces, etc. (Stevens 1996; Thijs et al. 2016). Some of the plants are able to decrease internal F by exporting it to the exterior surfaces of leaves (Baunthiyal and Ranghar 2013).
Phytoextraction (phytosequestration)
It involves absorption, translocation and accumulation of contaminants in the aerial parts. The process of extraction takes place through roots, where contaminants are initially absorbed from their immediate environment and then translocated into various harvestable parts (shoots, leaves, etc.) (Petzold et al. 2015). This approach usually involves hyperaccumulators or plants that can accumulate 0.1% or higher of contaminants on DM basis (Baker et al. 2000; Pollard et al. 2014). Plant species having bioconcentration factor (ratio of contaminant concentration in plant root to soil), translocation factor (ratio of contaminant concentration in plant shoot to root) and enrichment factor (ratio of contaminant concentration in plant shoot to soil) values greater than 1 would be considered as hyperaccumulators (Lorestani et al. 2011). A number of plants are known today which are having promising ability to accumulate F (Table 2). However, many plant species have been reported to accumulate F from the soil, but none have been commercially exploited for this purpose. Screening of potential hyperaccumulators for high F tolerance and resistivity is essentially required in the present time period. Rapidly growing high biomass crop with an extensive root system and showing least toxicity could be raised to remediate F from the soil (Baunthiyal and Ranghar 2013). Correspondingly, the aquatic plant species can be easily utilized to remediate F from the polluted water bodies. Baunthiyal and Sharma (2012) demonstrated the potential of eight tree species of semi-arid region for hyperaccumulation of F, out of which Prosopis juliflora (Sw.) DC., was shown to remove significant amount of F from groundwater and soil.
Fluoride tolerant and resistant plants have inherent cellular mechanisms to reduce its toxic effects. In regard, Saini et al. (2012) examined the organ-wise accumulation of F in P. juliflora. Their results showed that mentioned species accumulated highest amount of F in roots. The bioaccumulation and translocation factor values were found to be > 1 which illustrated high tolerance and accumulation efficiency of plants for F. In another study, 17 species were exposed to varying concentrations (2.5 to 10 mg L−1) of HF, out of which only three species, Camellia japonica L., Pittosporum tobira Thunb. and Saccharum officinarum L. were able to remove F efficiently. S. officinarum showed maximum removal capability, suggesting the activation of some detoxification process in the cell to withstand F concentrations (Santos-Díaz and Zamora-Pedrazaa 2010).
Microbial remediation
Soil rejuvenation after removing its pollutants and contaminants by using bacteria or fungi is termed as microbial remediation. These microbes utilize the contaminants and then degrade them for energy and reproduction.
Microbial remediation is performed by three processes:
Natural attenuation which takes place naturally with indigenous soil microorganisms
Biostimulation by providing external nutrients, moisture and an ideal pH to the microorganisms present in the soil
Bioaugmentation by the use of externally introduced microorganisms in situations where the naturally occurring microbes go dead because of the intensity of the contamination
Bacterial remediation of fluoride
Fluoride affects bacterial cells in many ways: firstly, by directly inhibiting the enzymes; secondly, by forming phosphate analogues like alumino-fluoride and beryllium fluoride complex: and lastly, by forming HF which acts as a transmembrane proton transporter, resulting in uncoupling of oxidative phosphorylation (Poulsen 2011). It has been studied that contamination by atmospheric F causes 80% decrease in soil bacterial biomass (Tscherko and Kandeler 1997). Fluoride mainly exists in combination with proton as HF, which readily cross through the cell membrane and acts as a proton conductor (Chouhan et al. 2012). The contaminant may disrupt the microbial membrane; however, the cell sometimes defends itself by forming an outer covering of hydrophobic layer having solvent efflux pumps over the membrane (Dixit et al. 2015). In spite of the cell deterioration induced by F, microorganisms have developed diverse machinery such as sorption, mineralization, uptake and accumulation, extracellular precipitation, enzymatic transformation and efflux from the cell, for adapting F toxicity (Hussein et al. 2004). These abilities could be sufficiently utilized for successful remediation of F by making it less accessible and toxic for the living forms.
Bacteria can acquire either transient or stable F resistance. Transient resistance is related to the horizontal transfer of plasmids between two cells, while stable F resistance is believed to be due to the chromosomal mutations (Liao et al. 2017). Inhibitory effects of F on intracellular metabolism depend upon the influx of HF which after diffusion dissociates into the H+ and F− (Hamilton 1990). This intracellular H+ and F− inhibit F-ATPase and enolase enzymes which adversely affects glycolysis pathway and acid tolerance of bacteria. These enzymes are believed to be mutated in the F-resistant bacteria (van Loveren et al. 2008). Also, bacteria can efficiently reduce F concentration because their cell wall is comprised of amines, sulphhydryl, carboxylates and phosphates which are metal-binding groups and help in adherence of the F ions to their surfaces. Certain bacterial species are tolerant to higher concentration of F since they can build their resistance through various mechanisms like bioaccumulation, biotransformation, etc. (Juwarkar and Yadav 2010). Different genera of bacteria have been reported which can tolerate high F concentrations along with the potential to remove it (Table 3). These bacteria can survive in F-contaminated soil and provides a new opportunity to develop bioremediation techniques which will be a novel way of alleviating F toxicity.
In 2012, Breaker discovered two gene families: crcB and eriCF which had identical biochemical roles. Both these families encoded F exporters which were directly related to the F resistance in microorganisms. To support this, authors constructed a reporter gene by joining crcB motif RNA (crcB gene) of Bacillus cereus and a homologous eriC motif RNA (eriC gene) of Pseudomonas syringae to a lacZ gene, and transferred this reporter construct into an Escherichia coli after mutating its natural lacZ gene. A high rate of expression was seen when cells were allowed to grow on a medium supplemented with higher concentration of F (Baker et al. 2012; Breaker 2012). Men et al. (2016) observed two eriCF genes in Streptococcus mutans, namely, perA and perB which were also involved in F resistance and has been approved by gene knockout study and gene regulation analysis. Studies revealed that the S. mutans became 100-fold more sensitive to F when both the genes were knocked out. Further, gene regulation studies proved that a single mutation in the promoter region can significantly upregulate the expression of both the genes, thereby conferring F resistance in S. Mutans (Liao et al. 2016; Men et al. 2016; Murata and Hanada 2016).
Mycoremediation
Mycoremediation is a method of environmental clean-up which relies on fungi and their enzyme systems. Fungi are metabolically and ecologically diverse organisms which have unique ability to degrade different types of pollutants. Furthermore, these also have ability to form extended mycelial networks, to absorb pollutants as their growth substrates; the process is more likely known as mycosorption. Fungi which are native to the contaminated sites play important role in modification and manipulation of the bioavailability of chemicals, thereby decreasing the allied risks. Their intracellular and extracellular catabolic enzymes make them well suited for bioremediation process which facilitates degradation of a broad range of pollutants and absorption of complex carbohydrates without hydrolysis. Application of fungal biosorbents like Pleurotus ostreatus (Ramanaiah et al. 2007), Aspergillus penicillioides, Mucor racemosus (Prajapat et al. 2010) and Pleurotus eryngii (Amin et al. 2015) for F removal have already been reported. The other fungal species which are used as tools in remediation process includes Trichoderma longibrachiatum, P. ostreatus, Rhizopus arrhizus, Trametes hirsute, T. versicolor, Lentinus edodes, Aspergillus niger, Aspergillus flavus, Aspergillus terreus, Cladosporium resinae and Phanerochaete chrysosporium (Cecchi et al. 2019). Fungi present the most significant class of soil biomass; however, they have not been exploited to their fullest for the treatment of contaminated environments.
Phycoremediation
Microbial usage for bioremediation has certain limitations such as bacterial sludge produced is of very little use and it also needs a long-standing commitment (Wang et al. 2002). Many of the microorganisms are also associated with the production of toxins and complex end products which are difficult to tackle. For this reason, nowadays phycoremediation has become a potential method for bioremediation (Cho et al. 2011). Algae have the capability of growing in any atmosphere, including domestic, municipal and industrial wastewaters. It could withdraw nutrients for growth from the pollutants; thereby decreasing the level of pollution and contributing to environmental clean-up (Mahapatra et al. 2014). On the other hand, algal biomass has affluent amount of hydrocarbons, polysaccharides and lipids which can be used for the production of biofuel. Many physiochemical and microbial remediation work has previously been done for removing F from soil and water, but little has been reported using microalgae (Bhatnagar and Bhatnagar 2000). There is a recent report on the use of a cyanobacterial strain Starria zimbabweensis for remediation of F-contaminated waste water. A maximum of 66.6% of F was removed following this strain from synthetic wastewater having 10 mg L−1 initial F concentration (Ramachandra et al. 2013; Biswas et al. 2018). Other algal strains which were used as biosorbents were Spirogyra sp.-IO2 and Chara fragilis (Levy and Strauss 1973; Mohan et al. 2007).
Plant-microbe interaction approach
In general, plants are in symbiotic relationship with soil microbes during their growth and development processes. These microorganisms residing in the rhizosphere impose various beneficial effects on the host plant following different mechanisms such as nitrogen fixation and nodulation. These microbes are referred as plant growth-promoting rhizobacteria (PGPR). They are likely to protect the health of plants in an eco-friendly manner, and their interactions with plants are utilized commercially, which have scientific implications for sustainable agriculture (Gouda et al. 2018).
Plant-microbe interaction acts as a low-cost biotechnological approach for ecosystem renaissance and for the F remediation. Most of the F accumulating plants interact with root-associated microbes to subsist in the toxic and adverse environment (Paul 2007). Efficacy of bioremediation may be partial, particularly when soils are polluted with recalcitrant organic molecules; hence, the combined use of microbes and plants can boost the efficiency (Thijs et al. 2016). Plant roots supply microaerophilic conditions, and their exudates increase the binding and availability of organic contaminants for the soil microbes. This process accelerates the degradability of contaminants by upregulating the microbial metabolism and by altering the soil pH or solubility of contaminants. Possible pathways for enhanced biodegradation of F by root exudates are (1) direct degradation by plant enzymes; (2) secretion of surfactants for enhanced bioavailability of pollutants; (3) utilization of structural analogy and co-metabolic processes and (4) continuous flow of nutrients and energy to stimulate whole process (Martin et al. 2014). Pseudomonas fluorescence has been seen to improve ability of P. juliflora for remediation of F by increasing the mineral content and biomass (Chaudhary and Khan 2016). Bacillus sp. also has capability to remove F from soil and has been reported as PGPR by many researchers (Sheng et al. 2008; Pal et al. 2014). These bacteria reduce F concentration from their surroundings by accumulating it inside their cells. This ability of tolerating high concentration of F may provide opportunity to develop a new bioremediation technique and a novel way of removing this xenobiotic from contaminated soil and water.
Roles of PGPR in phytoremediation
The PGPR promote phytoremediation either directly or indirectly. Direct process comprised of solubilization, bioavailability and final accumulation of contaminants by plants. In contrast, indirect processes involves plant growth enhancement by the production of essential enzymes and volatile organic compounds and by prevention from phytopathogens.
Direct process
Microbes present in the soil are involved in different reactions and metabolic processes that occur in the biogeochemical cycling of nutrients, maintenance of soil structure and detoxification of pollutants (Khan et al. 2010). Various PGPR produce siderophores at extreme environmental conditions including scarcity of nutrients or presence of elevated concentrations of pollutants, thus increasing their bio-accessibility in the rhizosphere through a complex reaction (Rajkumar et al. 2012). Low molecular weight organic acids like gluconic, oxalic and citric acids produced by PGPR helps in the mobilization of pollutants such as F, thereby increasing their uptake in plants (Ullah et al. 2015). Under nutrient deficient and adverse conditions, PGPR have the potential to provide essential nutrients to the plants. For instance, nitrogen is about 78% in the atmosphere but is unavailable to plants. Diazotrophic bacteria have the ability to fix atmospheric nitrogen, even under abiotic stresses. Moreover, PGPR play significant role in mineral uptake, stomatal regulation, osmotic modification and adaptation of root morphology (Compant et al. 2005; Ouzounidou et al. 2006). Biosurfactants are additional important metabolites produced by PGPR that may have the potential to improve F mobilization and phytoremediation. These biosurfactants might form complexes with F like other metals at the soil interface, desorbing it from the soil matrix and thus increasing its solubility and bioavailability for plants (Gadd 2010).
Indirect process
Pollutants present in the rhizosphere affect nutrient uptake, consequently reducing plant growth. The PGPR provide tolerance to plants by protecting them from phytopathogens and by contributing to enhanced biomass production. This not only helps the plants to grow and flourish in the contaminated ecosystem but also facilitate them to remove immediate pollutants (Luo et al. 2012). Plants may have altered physiology and biochemistry owing to numerous environmental stresses. In such conditions, many PGPR have the capability of producing phytohormones like indole-acetic acid, cytokinins and gibberellins, thereby improving hormone levels and plant responses to stresses (Glick 2012). Collectively, these bacteria improve plant growth in polluted soil, which in turn facilitates the phytoremediation process.
Technologies to understand plant-microbe interaction during fluoride remediation
Transgenic technologies
The potential of plant-microbe interactions for F remediation can be further improved by accomplishment of genetic modifications both in soil bacteria and in plant species.
Omics approaches
Plant responses towards environmental changes comprise of several means which include changes in gene expression (transcriptome), defence responses and also accumulation of protein(s) that can degrade pollutants or metabolites. Nowadays, omics technologies are widely used to understand the cellular processes, genetic control and signalling networks involved in plant responses to environmental stresses. For this, next-generation sequencing technology in addition to mass spectrometry has made a noteworthy contribution to figure out how plant-microbe interaction occurs and also to resolve problems related to soil remediation (Deshmukh et al. 2014; Rabara et al. 2017). The omics technology also has the potentiality to understand the gene-protein networks involved in bioremediation/phytoremediation and can be further utilized for the treatment of F-contaminated ecosystems. Additionally, they can be pooled with other new diagnostic technologies such as mathematical modelling or network biology for the better understanding of the biological processes implicated in the plant-microbe interaction.
Transcriptomics and proteomics
Transcriptomic means the study of transcriptome, the complete set of RNA transcripts produced by the genome under specific condition or in specific cells. Comparison of transcriptomes by using high-throughput methods like microarray analysis allows the identification of genes that are differentially expressed in distinct cell population or in response to different treatments and stresses. With the help of transcriptomics, improvement can be done in plants and microbes which could further be used for phytoremediation. Through genetic engineering, manipulation in gene expression pattern can be done in plants responsible for (i) uptake and transportation of different pollutants and (ii) converting pollutants to less toxic and volatile forms (Kotrba et al. 2009; Mosa et al. 2016).
Bacterial cells use various RNA and proteins to reduce F toxicity which could possibly serve as objectives for compounds that increases the uptake or release of F by bacteria. The crcB gene is associated with F riboswitches, and thus, compounds that can block the F exporter, encoded by this gene might serve as antimicrobial agent when combined with F. One can exploit components of the F sensory and resistance systems to manipulate cellular concentration of F. For example, F-specific biochemical tools such as F riboswitches, when fused with reporter genes, can act as convenient in vivo sensor of F detection. Similarly, model bacterial species can be genetically altered to make them more sensitive or resistant to F by knocking out or overexpressing exporters or other toxicity mitigation proteins (Baker et al. 2012).
In addition, this technology can identify several metabolic pathways that are modified in response to stresses and can also be exploited in transcriptome profiling to identify upregulated or downregulated genes under stress conditions. A transcriptomic technology is also used to detect and alter the PGPR genes that may be upregulated or downregulated under polluted environmental conditions and can be utilized to enhance the phytoremediation process.
Proteomics is an analytical technique used to identify changes in the protein expression of hyperaccumulator plants and microbes in response to stresses. The utilization of various proteomic tools and technologies has made easy the characterization of plant and environmental interactions, expanding our knowledge of these processes in the future. It has been previously reviewed that in response to various environmental stimuli, plant proteins are involved in establishing many types of plant-bacterial interactions, which includes both, resistance towards pathogenic bacteria and symbiotic relations for nutrient availability (Gamalero and Glick 2011). Chouhan et al. (2012) detected an extra protein band of 30 kDa in Micrococcus luteus and three extra bands with molecular weights of 20, 22 and 25 kDa in Pseudomonas aeruginosa grown in media having F than grown in F lacking media. Understanding the crucial involvement of these proteins and their detailed profiling by proteomic approach may help in understanding the mechanisms evolved in plants for battling with environmental contaminants. Further, a relative proteomics study of plant-microbe interactions in a polluted environment can discover key players of phytoremediation (Chouhan et al. 2012).
Metagenomics
A number of new encroachments in science and technology have always been adapting in the field of bioremediation for establishment of better environment, and in that progression, metagenomics is considered as one of the better emerging technologies. Metagenomics is a tactical approach to identify microbes at the genomic level. For the detection of specific pollutant degrading microbe, researchers isolate the microbial genome directly from the contaminated sample, sequence and then perform reverse transcriptase real-time PCR (RT-qPCR) to quantify the expression of specific genes involved in the degradation of a pollutant (Deverapalli and Kumawath 2015). Screening and identification of metagenomes from the contaminated environments are essential in this study. Various studies have showed that the interest in the metagenomics-based bioremediation has been gradually increased and has proved to be the best adaptation which leads to establishment of a non-toxic and pure environment. The approach can be successfully applied for the identification of F-resistant strains and understanding their gene profiles best adapted for F remediation.
Nanotechnology
These days nanotechnology mediated bioremediation is in practice widely. Researchers have shown that nanosized zerovalent ions have the potential to degrade organic pollutants like atrazine, molinate and chlorpyrifos (Zhang 2003; Ghormade et al. 2011). It also affects the uptake of organic pollutants like trichloroethylene in Populus deltoides W. Bartram ex Marshall via fullerenes (Ma and Wang 2010).
Amongst various physiochemical methods, adsorption offers potential benefits particularly in places where water supply is limited and in small-scale community level or household level where water treatment is required. This process is less energy intensive and offers other advantages like ease of operation, flexibility, simplicity of design and cost-effectiveness (Sarkar et al. 2006; Kumar et al. 2011). Many materials like alumina, iron-based oxides, rare metal oxides, activated carbon, etc., have been tested as possible adsorbents for removal of F from water (Stanic et al. 2014). In recent years, application of nanomaterials in environmental applications is getting more importance due to their advantages such as high reactivity and high surface area to volume ratio (Tang and Lo 2013). Christina and Viswanathan (2015) developed iron oxide nanoparticles which were later immobilized in sodium alginate matrix and studied for their F removal potential. These nanoparticles exhibited a maximum adsorption capacity of ~ 58 mg g−1 F for an initial concentration of 40 mg L−1 F at pH 5. In another study, bacterial-surfactin-mediated hydroxyapatite (Hap) nanoparticle was able to remove F from water. Findings of Bazrafshan et al. (2016) showed copper oxide nanoparticles as effective adsorbent of F.
Nanoparticles of various metals are used for plant growth as well as remediation of silver, gold, titanium, silicon, etc. A report showed that silver nanoparticles amplified the PGPR activity which then increased the root area and length of host plant (Sajid et al. 2006). The root-shoot ratio was also changed with the implementation of silver nanoparticles. The plants irrigated with municipal wastewater had higher activities of peroxidase and catalase which were further augmented by the solicitation of silver nanoparticles. Application of silver nanoparticles also modulated the levels of abscisic acid (34%), indole acetic acid (55%) and gibberellic acid (82%). These also increased the proline (70%), encountered oxidative stress and augmented the bioremediation potential of PGPR for lead, cadmium and nickel (Lippmann et al. 1995; Cardoso et al. 2005). Municipal wastewater needs to be treated with PGPR and silver nanoparticles prior to its use for irrigation. This aims for the better growth of the plant and enhanced bioremediation of toxic substances like F (Khan and Bano 2018). Researches proposed that Sol-Gel synthesized titanium oxide nanoparticles stably attach with PGPRs of plant roots and encourage bacterial performance (Timmusk et al. 2018).
Future prospects
Studies on the approaches of F remediation have provided a greater understanding of the use of different biological agents with unique capabilities in treating F contamination in a more pronounced way. The diversified and unmatching performances of the microorganisms and plant varieties look forward in designing new strategies for improving their efficacy. Further, strain discovery, improvement and selection of new isolates will not only create new avenues in treating F but would also expand towards broader spectrum of action. Formulation of PGP microbial consortia and transgenic strains for F remediation can be accomplished to boost their survivability into the environment for more efficient results (Islam et al. 2013; Ojuederie and Babalola 2017). New recombinant DNA techniques can be used to engineer F-degrading microorganisms with targeted mode of action. Moreover, developing new methods for regulating gene expression and the use of targeted and random mutagenesis which increases the activity of biodegrading enzymes in these organisms is highly desirable. Besides this, more research is needed to completely understand the metabolic pathways in transgenic plants and microbes so as to discover their impact and side effects when used in bioremediation. Likewise, hyperaccumulator plants can also be genetically enhanced for effective extraction of F from the soil environment by the process of phytoextraction for efficient phytoremediation (Ojuederie and Babalola 2017).
Conclusions
The prominent and solemn effects of F on the biosphere are a matter of concern and require a worldwide attention to deliver solutions for its secure removal from the environment. While many physiochemical techniques have been developed, bioremediation is a safe and innovative method for remediating this toxicant. Phytoremediation with PGPR is an emerging technology that appears to solve problems without any hazardous effects. These soil bacteria transform pollutants into soluble and bioavailable forms, which consequently facilitates phytoremediation. Other than this, chemical and biological synthesis of metal nanoparticles, formation of bacterial consortium and the use of transgenics are gaining momentum for the cautious and efficient removal of F from the environment. Even though the above association proved as beneficial approaches for F handling, more intuitive discovery is necessary to optimize the plant-microbe interaction which requires a well-orchestrated combination of various omics technologies along with skilful implementation of gene editing and other transgenic approaches to introduce foreign genes.
References
Abdallah FB, Elloumi N, Mezghani I, Garrec JP, Boukhris M (2006) Industrial fluoride pollution of Jerbi grape leaves and the distribution of F, Ca, mg and P in them. Fluoride 39:43–48
Ali S, Thakur SK, Sarkar A, Shekhar S (2016) Worldwide contamination of water by fluoride. Environ Chem Lett 14:291–315
Amin F, Talpur FN, Baloucha A, Surhioa MA, Bhutto MA (2015) Biosorption of fluoride from aqueous solution by white-rot fungus Pleurotus eryngii ATCC 90888. Environ Nanotechnol Monit Manage 3:30–37
Arlappa N, Qureshi A, Srinivas R (2013) Fluorosis in India: an overview. Int J Res Dev Health 1:97–102
Baker AJM, McGrath SP, Reeves RD, Smith JAC (2000) Metal hyper accumulator plants: a review of the ecology and physiology of a biological resource for phytoremediation of metal-polluted soils. Phytorem Contaminated Soil Water:85–107
Baker JL, Sudarsan N, Weinberg Z, Roth A, Stockbridge RB, Breaker RR (2012) Widespread genetic switches and toxicity resistance proteins for fluoride. Science 335:233–235
Banerjee G, Sengupta A, Roy T, Banerjee PP, Chattopadhyay A, Raya AK (2016) Isolation and characterization of fluoride resistant bacterial strains from fluoride endemic areas of West Bengal: assessment of their fluoride absorption efficiency. Fluoride 49:414–425
Barbier O, Arreola-Mendoza L, Del Razo LM (2010) Molecular mechanisms of fluoride toxicity. Chem Biodivers 188:319–333
Baunthiyal M, Ranghar S (2013) Accumulation of fluoride by plants: potential for phytoremediation. Clean Soil Air Water 43:127–132
Baunthiyal M, Sharma V (2012) Phytoremediation of fluoride contaminated water and soil: a search for fluoride hyperaccumulators. J Agri Tech 8:1965–1978
Bazrafshan E, Balarak D, Panahi AH, Kamani H, Mahvi AH (2016) Fluoride removal from aqueous solutions by cupricoxide nanoparticles. Fluoride 49:233–244
Bhatnagar M, Bhatnagar A (2000) Algal and cyanobacterial responses to fluoride. Fluoride 33:55–65
Biswas G, Thakurta SG, Chakrabarty J, Adhikari K, Dutta S (2018) Evaluation of fluoride bioremediation and production of biomolecules by living cyanobacteria under fluoride stress condition. Ecotox Environ Saf 148:26–36
Breaker RR (2012) New insight on the response of bacteria to fluoride. Caries Res 46:78–81
Cardoso PF, Priscila LG, Rui AG, Leonardo OM, Ricardo AA (2005) Response of Crotalaria juncea to nickel exposure. J Plant Physiol 17:267–272
Cecchi G, Vagge G, Cutroneo L, Greco G, Di Piazza S (2019) Fungi as potential tool for polluted port sediment remediation. Environ Sci Pollut Res 26:35602–35609. https://doi.org/10.1007/s11356-019-04844-5
Chaudhary K, Khan S (2016) Physiochemical characterization of fluoride contaminated soil and its microbe-assisted bioremediation by Prosopis juliflora. J Plant Biol Soil Health 3:8
Cho S, Luong TT, Lee D, Lee T (2011) Reuse of effluent water from a municipal wastewater treatment plant in microalgae cultivation for biofuel production. Bioresour Technol 102:8639–8645
Choubisa SL, Sompura K (1996) Dental fluorosis in tribal villages of Dungarpur district (Rajasthan). Pollut Res 15:45–47
Chouhan S, Lomash V, Flora SJS (2009) Fluoride-induced changes in haem biosynthesis pathway, neurological variables and tissue histopathology of rats. Appl Toxicol 30:63–73
Chouhan S, Tuteja U, Flora SJS (2012) Isolation, identification and characterization of fluoride resistant bacteria: possible role in bioremediation. J App Biochem Microbiol 48:43–50
Christina E, Viswanathan P (2015) Development of a novel nano-biosorbent for the removal of fluoride from water. J Chem Eng 23:924–933
Compant S, Duffy B, Nowak J, Clément C, Barka EA (2005) Use of plant growth promoting bacteria for biocontrol of plant diseases: principles mechanisms of action, and future prospects. Appl Environ Microbiol 71:4951–4959
Cooke JA, Johnson MS, Davidson AW, Bradshaw AD (1976) Fluoride in plants colonising fluorspar mine waste in the peak district and Weardale. Environ Pollut 11:10–13
Davis RD (1980) Uptake of fluoride by rye-grass in soil treated with sewage sludge. Environ Pollut 1:277–284
Deshmukh R, Sonah H, Patil G, Chen W, Prince S, Mutava R, Vuong T, Valliyodan B, Nguyen HT (2014) Integrating omic approaches for abiotic stress tolerance in soybean. Front Plant Sci 5:244
Deverapalli P, Kumawath RN (2015) Metagenomics- a technological drift in bioremediation. In: Shiomi N (ed) advances in bioremediation of wastewater and polluted soil. InTech, Europe. https://doi.org/10.5772/60749
Dixit R, Wasiullah MD, Pandiyan K, Singh UB, Sahu A, Shukla R, Singh BP, Rai JP, Sharma PK, Lade H, Paul D (2015) Bioremediation of heavy metals from soil and aquatic environmental: an overview of principles and criteria of fundamental processes. Sustain 7:2189–2212
Doble M, Kumar A (2005) Biotreatment of industrial effluents. Elsevier, Butterworth-Heinemann
Elloumi N, Abdallah FB, Mezghani I, Rhouma A, Boukhris M (2005) Effect of fluoride on almond seedlings in culture solution. Fluoride 38:193–198
Ericsson Y (1970) Fluorides and human health. Lancet 2:351–352
Gadd GM (2010) Metals: minerals and microbes: geomicrobiology and bioremediation. Microbiol 156:609–643
Gamalero E, Glick BR (2011) Mechanisms used by plant growth-promoting Bacteria. In: Maheshwari D (ed) Bacteria in agrobiology: plant nutrient management. Springer, Berlin
Gentil FG, Fick J (2016) Algal cultivation in urban wastewater: an efficient way to reduce pharmaceutical pollutants. J Appl Phycol29:255–262
Ghormade V, Deshpande MV, Paknikar KM (2011) Perspectives for nano-biotechnology enabled protection and nutrition of plants. Biotechnol Adv 29:792–803
Ghosh A, Mukherjee K, Ghosh SK (2013) Sources and toxicity of fluoride in environment. Res Chem Intermed 39:2881–2915
Glick BR (2012) Plant growth-promoting bacteria: mechanisms and applications. Scientifica:1–15
Gouda S, Kerry RG, Das G, Paramithiotis S, Shin HS, Patra JK (2018) Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol Res 206:131–140
Halder GN, Sinha S, Dhawane S (2015) Defluoridation of wastewater using powdered activated carbon developed from Eichhornia crassipes stem: optimization by response surface methodology. Desal Water Treat 56:953–966
Hamilton IR (1990) Biochemical effects of fluoride on oral bacteria. J Dent Res 69:660–667
He LF, Chen JG (2006) DNA damage, apoptosis and cell cycle changes induced by fluoride in rat oral mucosal cells and hepatocytes. World J Gastroenterol 1:144–1148
Hussein H, Moawad H, Farag S (2004) Isolation and characterization of Pseudomonas resistant to heavy metals contaminants. Arab Biotech 7:13–22
Islam MS, Ueno Y, Sikder MT, Kurasaki M (2013) Phytofiltration of arsenic and cadmium from the water environment using Micranthemum umbrosum (GMEL) blake as a hyperaccumulator. Int J Phytoremed 15:1010–1021
Jayarathne A, Weerasooriya R, Chandrajith R (2014) A rapid method for the removal of fluoride in contaminated groundwater using natural crystalline apatite: a laboratory and field study. Environ Earth
Juwarkar A, Yadav S (2010) Bioaccumulation and biotransformation of heavy metals. Biorem Technol 9:266–284
Kamran MA, Amna MR, Mubariz N, Syed JH, Bano A, Javed MT, Munis MFH, Tan Z, Chaudhary HJ (2014) The potential of the flora from different regions of Pakistan in phytoremediation: a review. Environ Sci Pollut Res 21:801–812
Khan N, Bano A (2018) Role of plant growth promoting rhizobacteria and Ag-nano particle in the bioremediation of heavy metals and maize growth under municipal wastewater irrigation. Int J Phytorem 18:211–221
Khan S, Hesham AEL, Qiao M, Rehman S, He JZ (2010) Effects of cd and Pb on soil microbial community structure and activities. Environ Sci Pollut Res 17:288–296
Khandare AL, Rao GS (2006) Uptake of fluoride, aluminium and molybdenum by some vegetables from irrigation water. J Hum Ecol 19:283–288
Khandare RV, Desai SKB, Bhujbal SS, Watharkar AD, Biradar SP, Pawar PK, Govindwar SP (2017) Phytoremediation of fluoride with garden ornamentals Nerium oleander, Portulaca oleracea, and Pogonatherum crinitum. Environ Sci Pollut Res 24:6833–6839
Kotrba P, Najmanova J, Macek T, Ruml T, Mackova M (2009) Genetically modified plants in phytoremediation of heavy metal and metalloid soil and sediment pollution. Biotechnol Adv 27:799–810
Kumar V, Talreja N, Deva D, Sankararamakrishnan N, Sharma A, Verma N (2011) Development of bi-metal doped micro- and nano multi-functional polymeric adsorbents for the removal of fluoride and arsenic (V) from wastewater. Desalination 282:27–38
Levy LW, Strauss RL (1973) Sur L' insolubilisation de L' Ion fluor a L' etat- de fluorure de calcium chez Chara fragilis. Desvaux CR. Acad Sci 277:181–184
Liao Y, Brandt BW, Zhang M (2016) A single nucleotide change in the promoter mutp enhances fluoride resistance of Streptococcus mutans. Antimicrob Agents Chemother 60:7509–7512
Liao Y, Brandt BW, Li J, Crielaard W, Loveren CV, Deng DM (2017) Fluoride resistance in Streptococcus mutans: a mini review. J Oral Microbiol 9:1344509
Lippmann B, Leinhos V, Bergmann H (1995) Influence of auxin producing rhizobacteria on root morphology and nutrient accumulation of crops, pt. 2: changes in root morphology and nutrient accumulation in maize (Zea mays L.) caused by inoculation with indole-3-acetic acid (IAA) producing Pseudomonas and Acinetobacter strains or IAA applied exogenously. Angew Bot 69:31–36
Lorestani B, Cheraghi M, Yousefi N (2011) Phytoremediation potential of native plants growing on a heavy metals contaminated soil of copper mine in Iran. World Acad Sci Eng Technol 77:377–382
Luo S, Xu T, Chen L, Chen J, Rao C, Xiao X, Wan Y, Zeng G, Long F, Liu C, Liu Y (2012) Endophyte-assisted promotion of biomass production and metal-uptake of energy crop sweet sorghum by plant-growth-promoting endophyte Bacillus sp. SLS18. Appl Microbiol Biotechnol 93:1745–1753
Ma X, Wang C (2010) Fullerene nanoparticle affect the fate and uptake of trichloroethylene in phytoremediation systems. Environ Eng Sci 27:989–992
Mahapatra DM, Chanakya HN, Ramachandra TV (2014) Bioremediation and lipid synthesis through mixotrophic algal consortia in municipal wastewater. Bioresour Technol 168:142–150
Martin BC, George SJ, Price CA, Ryan MH, Tibbett M (2014) The role of root exuded low molecular weight organic anions in facilitating petroleum hydrocarbon degradation: current knowledge and future directions. Sci Total Environ 472:642–653
Men X, Shibata Y, Takeshita T (2016) Identification of anion channels responsible for fluoride resistance in oral Streptococci. PLoS One 11:0165900
Messaitfa A (2008) Fluoride contents in ground waters and the main consumed foods (dates and tea) in southern Algeria region. Environ Geol 55:377–383
Mohan SV, Ramanaiah SV, Rajkumar B, Sarma PN (2007) Removal of fluoride from aqueous phase by biosorption onto algal biosorbent Spirogyra sp.-IO2: sorption mechanism elucidation. J Hazard Mater 141:465–474
Mosa KA, Saadoun I, Kumar K, Helmy M, Dhankher OP (2016) Potential biotechnological strategies for the cleanup of heavy metals and metalloids. Front Plant Sci 7:303
Mukherjee S, Halder G (2016) Assessment of fluoride uptake performance of raw biomass and activated biochar of Colocasia esculenta stem: optimization through response surface methodology. Environ Prog Sustain Energy 35:1305–1316
Mukherjee S, Yadav V, Mondal M, Banerjee S, Halder G (2015) Characterization of fluoride-resistant bacterium Acinetobacter sp. RH5 towards assessment of its water defluoridation capability. Appl Water Sci 7:1923–1930
Mukherjee S, Sahu P, Halder G (2017) Microbial remediation of fluoride-contaminated water via a novel bacterium Providencia vermicola (KX926492). J Env Manage 204:413–423
Mukherjee S, Sahu P, Halder G (2018) Comparative assessment of the fluoride removal capability of immobilized and dead cells of Staphylococcus lentus (KX941098) isolated from contaminated groundwater. Env Progress Sustainable Energy 37:5
Murata T, Hanada N (2016) Contribution of chloride channel permease to fluoride resistance in Streptococcus mutans. FEMS Microbiol Lett 363:101
Ojuederie OB, Babalola OO (2017) Microbial and plant-assisted bioremediation of heavy metal polluted environments: a review. Int J Environ Res Public Health 14:1504
Ouzounidou G, Moustakas M, Symeonidis L, Karataglis S (2006) Response of wheat seedlings to Ni stress: effects of supplemental calcium. Arch Environ Contam Toxicol 50:346–352
Pal KC, Mondal NK, Chatterjee S, Ghosh TS, Datta JK (2014) Characterization of fluoride-tolerant halophilic Bacillus flexus NM25 (HQ875778) isolated from fluoride-affected soil in Birbhum District, West Bengal, India. Environ Monit Assess 186:699–709
Patil SB, Jagdale AD, Patil DB, Kulkarni AS, Govindwar SP (2014) Evaluation of fluoride uptake and accumulation by Portulaca grandiflora hook. And Brassica oleracea Linn.From water. Int J Science Eng Tech Res 3:3474–3477
Paul E (2007) Soil microbiology, ecology, and biochemistry (3rd E). Vadose Zone J 8:1087–1088
Petzold CJ, Chan LJ, Nhan M, Adams PD (2015) Analytics for metabolic engineering. Front Bioeng Biotechnol 3:135
Pollard AJ, Reeves RD, Baker AJM (2014) Facultative hyperaccumulation of heavy metals and metalloids. Plant Sci 218:8–17
Poulsen R (2011) The effect of fluoride pollution on soil microorganisms. Bachelor thesis Faculty of Physical Sciences University of Iceland 42
Prajapat R, Bhatnagar A, Gaur KR, Bajpai V (2010) Fluoride removal from water by sorbing on plant and fungal biomass. Int J Biotechnol 1:43–46
Rabara RC, Tripathi P, Rushton PJ (2017) Comparative metabolome profile between tobacco and soybean grown under water-stressed conditions. Biomed Res Int 3065251:18
Rajkumar M, Sandhya S, Prasad MNV, Freitas H (2012) Perspectives of plant associated microbes in heavy metal phytoremediation. Biotechnol Adv 30:1562–1574
Ramachandra TV, Mahapatra DM, Samantray S, Joshi NV (2013) Biofuel from urban wastewater: scope and challenges. Renew Sust Energ Rev 21:767–777
Ramanaiah SV, Mohan SV, Sarma PN (2007) Adsorptive removal of fluoride from aqueous phase using waste fungus (Pleurotus ostreatus 1804) biosorbent: kinetics evaluation. Ecol Eng 31:47–56
Rangel NC, Velázquez ANR, Díaz MSS (2015) Fluoride bioaccumulation by hydroponic cultures of camellia (Camellia japonica spp.) and sugar cane (Saccharum officinarum spp.) chemosphere. 136:56–62
Rose D, Marier JR (1977) National Research Council of Canada, Associate Committee on Scientific Criteria for Environmental Quality NRCC No. J Environ Fluoride:16081
Saini P, Khan S, Baunthiyal M, Sharma V (2012) Organ-wise accumulation of fluoride in Prosopis juliflora and its potential for phytoremediation of fluoride contaminated soil. Chemosphere 89:633–635
Sajid M, Nadeem A, Zahir M, Naveed M, Arshad M, Shahzad SM (2006) Variation in growth and ion uptake of maize due to inoculation with plant growth promoting rhizobacteria under salt stress. Soil & Environ 25:78–84
Santos-Díaz MS, Zamora-Pedrazaa C (2010) Fluoride removal from water by plant species that are tolerant and highly tolerant to hydrogen fluoride. Fluoride 43:150–156
Sarkar M, Banerjee A, Pramanick PP, Sarkar AR (2006) Use of laterite for the removal of fluoride from contaminated drinking water. J Colloid Interface Sci 302:432–441
Sehn P (2008) Fluoride removal with extra low energy reverse osmosis membranes: three years of large scale field experience in Finland. Desalination 223:73–84
Sheng X, Jiang C, He L (2008) Characterization of plant growth–promoting Bacillus edaphicus NBT and its effect on lead uptake by Indian mustard in a lead–amended soil. J Microbiol 54:417–422
Shivarajashankara YM, Shivashankara AR, Bhat PG, Rao SH (2002) Brain lipid peroxidation and antioxidant systems of young rats in chronic fluoride intoxication. Fluoride 35:197–203
Singh M, Verma KK (2013) Influence of fluoride-contaminated irrigation water on physiological responses of poplar seedlings (Populus deltoides L. clone-S7C15). Fluoride 46:83–89
Singh S, Singh J, Singh N (2013) Studies on the impact of fluoride toxicity on growth parameters of Raphanus sativus L. Indian J Sci Res 4:61–63
Stanic MH, Ravancic ME, Flanagan A (2014) A review on adsorption of fluoride from aqueous solution. Materials 7:6317–6366
Stevens DP (1996) Uptake of fluoride by plant roots. Thesis PhD. Dept of soil science. Uni Adelaide Australia
Stevens DP, McLaughlin MJ, Randall PJ, Keerthisinghe G (2000) Effects of fluoride supply on fluoride concentrations in five pasture species: levels required to reach phytotoxic or potentially zootoxic concentrations in plant tissue. Plant Soil 227:223–233
Tang SCN, Lo IMC (2013) Magnetic nanoparticles: essential factors for sustainable environmental applications. Water Res 47:2613–2632
Thijs S, Sillen W, Rineau F, Weyens N, Vangronsveld J (2016) Towards an enhanced understanding of plant–microbiome interactions to improve phytoremediation: engineering the metaorganism. Front Microbiol:7
Timmusk S, Seisenbaeva G, Behers L (2018) Titania (TiO2) nanoparticles enhance the performance of growth-promoting rhizobacteria. Sci Rep:8
Tscherko D, Kandeler E (1997) Ecotoxicological effects of fluorine deposits on microbial biomass and enzyme activities in grasslands. Eur J Soil Sci 48:329–335
Ullah A, Mushtaq H, Ali H, Munis MFH, Javed MT, Chaudhary HJ (2015) Diazotrophs-assisted phytoremediation of heavy metals: a novel approach. Environ Sci Pollut Res 22:2505–2514
van Loveren C, Hoogenkamp MA, Deng DM (2008) Effects of different kinds of fluorides on enolase and ATPase activity of a fluoride-sensitive and fluoride resistant Streptococcus mutans strain. Caries Res 42:429–434
Wang J, Zhao FJ, Meharg AA, Raab A, Feldmann J, McGrath SP (2002) Mechanisms of arsenic hyperaccumulation in Pteris vittata: uptake kinetics, interactions with phosphate, and arsenic speciation. Plant Physiol 130:1552–1561
World Health Organisation (2008) Chemical fact sheets: fluoride, guidelines for drinking water quality (electronic resource), 3rd ed. Incorporation first addendum, vol 1. WHO, Geneva, pp 375–377
Zhang WX (2003) Nanoscale iron particles for environmental remediation: an overview. J Nanopart Res 5:323–332
Zhang M, Wang A, Xia T, He P (2008) Effects of fluoride on DNA damage, S-phase cell-cycle arrest and the expression of NF-kappaB in primary cultured rat hippocampal neurons. Toxicol Lett 179:1–5
Funding
The authors would like to thank University Grants Commission, New Delhi, and Scientific and Engineering Research Board, New Delhi, for awarding fellowships to Priya Katiyar and Neha Pandey, respectively. Authors are also grateful to Department of Science & Technology, New Delhi, for financial support through DST-FIST scheme.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Elena Maestri
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Katiyar, P., Pandey, N. & Sahu, K.K. Biological approaches of fluoride remediation: potential for environmental clean-up. Environ Sci Pollut Res 27, 13044–13055 (2020). https://doi.org/10.1007/s11356-020-08224-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-020-08224-2