
Abstract
Contaminated sediments represent an important management problem that also concerns their remediation. Indeed, port dredging activities produce huge volumes of contaminated sediments that, in turn, require proper handling because of their quantity of inorganic and organic substances. Conventional management-remediation strategies of polluted sediment involve sediment washing, electron-chemical separation, and thermal treatment. Recently, bioremediation strategies have also been proposed as a promising answer to the problem of contaminated sediments. In this context, fungi are pioneer microorganisms known to bioconcentrate, bioaccumulate, and biostabilize heavy metals. These capabilities suggest the potential to employ indigenous fungal strains to remediate polluted port sediments. In the framework of the European Project SEDITERRA (Guidelines for the sustainable treatment of dredged sediments in the Marittimo area), the aim of this paper is to characterize the fungal communities of port sediments of Genoa and present an innovative mycoremediation protocol to evaluate the capability of indigenous fungal strains in the heavy metal remediation. In this study, Penicillium expansum Link and Paecilomyces formosus (Sakag., May. Inoue & Tada) Houbraken & Samson have been selected as fungal species for the mycoremediation treatments. The protocol requires a fungal membrane system and the results highlight efficient bioremoval of Cu and Zn from sediments.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The contamination of marine sediments related to port activities represents one of the most actual and global scale environmental problems, since high population densities, industrial activities, and hydrographical characteristics often favor the accumulation of high concentrations of contaminants in estuaries, harbors, ports, and coastal marine zones (Akcil et al. 2015; Fathollahzadeh et al. 2014; Tavakoly Sany et al. 2013; Walker et al. 2013b). Sediment contamination is often due to both organic (i.e., sludge; polycyclic aromatic hydrocarbons, PAHs; polychlorinated biphenyls, PCBs; etc.) and inorganic pollutants (mostly metals; Akcil et al. 2015; Fathollahzadeh et al. 2014). In one study, Fathollahzadeh et al. (2014) describe the most important pressures posed by human activities in coastal regions: the discharge of urban/industrial storm water runoff, domestic/industrial wastewater, diffuse pollution from agricultural fields, and port/harbor-related activities like shipping and painting of boat/ship surfaces. In particular, contaminants tend to accumulate in the bottom sediments of all aquatic systems, where they can be classified as potentially hazardous materials (Abdollahi et al. 2013; Buruaem et al. 2012; Garcia-Orellana et al. 2011).
Due to the need for periodical or capital dredging activities to maintain navigational depth in ports and waterways (Vagge et al. 2018; Cutroneo et al. 2015; OSPAR Commission 2014; Walker et al. 2013a), contaminated sediments represent an important management problem. In many cases, particularly in the USA and Norway, contaminated dredged sediments are confined in aquatic disposal cells designed for long-term isolation by using layers of capping materials (Ruiz and Schroeder 2007). For example, in the port of Hong Kong, one of the world-leading port cities, more than 1 Mm3 of marine contaminated sediment is generated annually with dredging operations (Wang et al. 2018). These sediments are historically highly contaminated with polychlorinated biphenyls (PCBs) and polycyclic aromatic hydrocarbons (PAHs) and are disposed of off-site in contaminated mud pits with clay covers, which require long-term monitoring to avoid secondary pollution (Wang et al. 2018). Contaminated sediments are often disposed of in landfills or confined disposal facilities. For example, 3 Mm3 of contaminated sediments were dredged in the Port of Genoa (Italy; Cutroneo et al. 2014). However, the disposal of dredged sediments is expensive and the costs rapidly increase if the sediment contaminants exceed applicable guidelines, resulting in more expensive disposal in containment cells (Walker et al. 2013b). Mulligan et al. (2001) showed that in the USA, approximately 10% of sediments in underlying waters are contaminated; there are 300 Mm3 of sediments that are dredged every year and among them, 3–12 Mm3 are highly contaminated. On the other hand, in Europe, Iannelli et al. (2012) reported that approximately 100–200 Mm3 of contaminated sediments are dredged yearly for different purposes. These past works identified how port dredging activities produce huge volumes of contaminated sediments that require proper handling due their load of inorganic and organic substances (OSPAR Commission 2014). Therefore, the extent of contaminated dredged sediments represents a crucial and current research topic.
Inorganic and organic substances contained in contaminant sediments are toxic for marine organisms. However, while heavy metals are persistent in the environment, some organic constituents, such as PAHs, Tri(n-butyl)tin (TBTs), or PCBs, can be degraded with processes that also may take several years, giving rise to components or elements that are still dangerous for organisms in some cases (such as TBTs that can degrade into the less toxic derivatives di(n-butyl)tin (DBT) and mono(n-butyl)tin (MBT) (Cruz et al. 2015). These products can also degrade into Sn, which is highly harmful for life (Howe and Watts 2005). In other cases, the degradation products are not dangerous (such as PAHs; Coates et al. 1997). Metals can only be transformed into more soluble or insoluble substances and/or into less toxic species (Caplat et al. 2015); as such, the majority of remediation strategies aim to increase metal solubility and stability, reducing their bioavailability (Akcil et al. 2015). Traditional remediation technologies of polluted sediments involve washing, electron-chemical separation, and thermal treatment (Peng et al. 2009; Dermont et al. 2008), but recently, bioremediation strategies appear to be a promising solution to the problem of dredged sediments (Akcil et al. 2015; Fonti et al. 2015; Tabak et al. 2008). Bioremediation technologies consist of stimulating and exploiting live organisms (i.e., fungi and bacteria), which are components of natural biogeochemical cycles. These organisms can exploit natural biological and metabolic processes leading to bioimmobilization and biostabilization of metals (Akcil et al. 2015). In particular, both macro- and microfungi can modulate their microenvironment pH around the hyphae in order to better absorb inorganic compounds. As such, they can co-absorb other elements with similar chemical and physical characteristics (although they are useless or toxic), when mineral nutrients are scarce (Falandysz et al. 2018; Roccotiello et al. 2015). However, until now, the majority of bioremediation studies of marine sediments were conducted with bacteria (Johnson 2013; Tabak et al. 2008). Few studies have considered the role and bioremediation capability of fungi. Macro- and microfungi are active participants in soil ecosystems and pioneer organisms are able to survive in extreme conditions and colonize every kind of environment (Cecchi et al. 2018; Falandysz et al. 2018; Gadd et al. 2012). According to the definition proposed by Kohlmeyer and Kohlmeyer (1979), fungi in marine environments can be divided into obligate and facultative marine fungi. Obligate marine fungi can grow and sporulate only in a marine or estuarine habitat, while facultative marine fungi can live in freshwater and terrestrial fungi can grow and possibly sporulate also in the marine environment (Kohlmeyer and Kohlmeyer 1979). However, facultative marine fungi can often synthesize secondary metabolites different from those produced by their terrestrial counterparts (Bhakuni and Rawat 2006). This property of some facultative marine fungal strains not only show their metabolic activity, but also indicate that many of those fungi are not present as dormant spores, but play a central and active role in the marine microbial ecosystem (Damare et al. 2012). Fungal enzymes and organic acids interact with inorganic contaminants, allowing for fungal bioconcentration, bioaccumulation, and biostabilization of heavy metals (Cecchi et al. 2017a; Cecchi et al. 2017b; Gadd et al. 2012). This result suggests the possibility to employ indigenous fungal strains to remediate polluted port sediments. In the literature, few studies deal with fungal strains than can successfully stabilize metals in soils (Li et al. 2014); similarly, there is a lack of work that addresses fungal biosorption and bioaccumulation as a bioremediation strategy for port sediments.
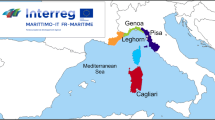
In the framework of the European Interreg Italy-France 2014-2020 Maritime Project SEDITERRA “Guidelines for the sustainable treatment of dredged sediments in the Marittimo area” the Department of Environmental, Earth, and Life Sciences (DISTAV) of the University of Genoa, Partner of the Project, is carrying out a pilot activity on sediments dredged in the Port of Genoa (North-western Italy; Fig. 1). Concerning the mycological aspects to the project, the goal is to both characterize the fungal communities of sediments and devise an innovative mycoremediation protocol to evaluate the heavy metal remediation capability of indigenous fungal strains. This paper is devoted to present the abovementioned pilot activities and show the preliminary mycological results.

Port of Genoa and point of sediment collection (black circle) in the inner part of the port
Materials and methods
Sampling and sediment characterization
In the inner part of the Port of Genoa, 30 kg of sediments were collected with a 5-L steel Van Veen grab. Three liters of seawater were collected with a 10-L Niskin bottle for subsequent analyses in the laboratory.
One kilogram of sediment (sediment at the initial time: S0) was immediately analyzed for initial contamination of heavy metals. The sample was first digested following the US Environmental Protection Agency (USEPA) Method 3050B (EPA 1996) and then analyzed with inductively coupled plasma–optical emission spectrometry (ICP-OES) following the EPA Method 6010D (EPA 2014).
A portion of the sediment (200 g) was analyzed to determine the grain size characteristics. Grain size analysis of the coarse fraction (particle diameter (∅) > 63 μm) was obtained using the procedure followed in Capello et al. (2016). The analysis of the fine fraction (∅ < 63 μm) was carried out using a Coulter Counter® Multisizer 3 (Beckman Coulter, Inc.). Moreover, another portion of sediment (15 g) was analyzed to determine the percentage of the organic (OF) and inorganic fraction (IF). The sediment (with a known weight; S) was burned in an ISCO muffle (ISM320 mod.) for 3 h at 550 °C to remove the OF; the unburned fraction (considered to be the IF) was subsequently weighed. The OF was determined as the difference between S and IF. The value of the grain size classes and OF and IF were expressed as a percentage.
Moreover, the sediment of the Port of Genoa was previously employed for fungal characterization by means of the dilution plate technique (Greco et al. 2018). Marine Rose Bengal Agar (MRBA; Sigma Chemical Co.) medium was prepared using sterilized port seawater. Fungi were counted as CFUs (colony-forming units) to calculate CFU g−1 dry soil. The sediment, diluted with sterile seawater 1:10 (w/v), was shaken by hand or mechanically for 10–20 min. The suspension was further diluted by a factor of 10; then, 1 ml aliquots of the suspensions (10−3, 10−2, and 10−1) were pipetted into Petri dishes and mixed with 10 ml of undercooled medium, which included antibiotics (MRBA) to prevent bacterial growth. Each soil sample was diluted and plated in three plates of MRBA for each dilution factor. After inoculum, plates were incubated in the dark at 24 °C and monitored on a weekly basis. The fungal colonies grown on the plates were isolated and maintained in axenic cultures (Greco et al. 2018). These strains were studied with a polyphasic approach (morphological, physiological, and molecular) (Samson et al. 2010). Morphological and physiological identification was carried out through cultivating each isolated fungal strain on different culture media (i.e., Potato Dextrose Agar, Malt Extract Agar, Czapek Yeast Agar, Czapek Dox Agar, etc.). Then, the initial detection of fungal structures was carried out with a stereomicroscope (× 10–50). The strains were subsequently identified using a micromorphological (× 100–100) analysis. Specific literature was used for the morphological and physiological fungal identification: Pitt (1979), for the genus Penicillium; Raper and Fennel (1965), Klich (2002), Samson et al. (2011), for Aspergillus; Bissett (1984, 1991a, b, c), Rifai (1969), for Trichoderma; Crous et al. (2007), Bensch et al. (2010, 2012), for Cladosporium; and Domsch et al. (2007), for strains not ascribable to other taxonomical groups. Genomic DNA was extracted from 100 mg of fresh fungal culture using a modified Cetyl Trimethylammonium Bromide (CTAB) method (Doyle and Doyle 1987). The amplification of the β-tubulin gene was performed using Bt2a and Bt2b primers (Glass and Donaldson 1995), while universal primers ITS1F/ITS4 were used for ITS region amplification (Gardes and Bruns 1993; White et al. 1990). Polymerase chain reaction (PCR) products were purified and sequenced using Macrogen Inc. (Seoul, Republic of Korea). Sequence assembly and editing were performed by means of SequencherR (Gene Codes Corporation, version 5.2). The taxonomic assignment of the sequenced samples was carried out using the BLASTN algorithm to compare the sequences obtained in the present study against the GenBank database. We took a conservative approach to species-level assignment (identity ≥ 97%) and verified the result accuracy by studying the macro- and micromorphological features of the colonies. The isolated strains were conserved in the culture collection of the Laboratory of Mycology of DISTAV (University of Genoa, Italy). These cultures were maintained by agar slants with periodic transfer and then cryopreserved (− 20 °C). The sequences obtained were deposited in GenBank with accession numbers: MG604357 (Penicillium expansum) and MK330005 (Paecilomyces formosus) (Greco et al. 2018).
Pilot mycological activity
Three plastic boxes were prepared with 5 kg of port sediment in order to check the mycoremediation activities of heavy metals. The sediment was covered with sterile cellophane membrane (M) in all the three boxes, and two boxes (1 and 2) were inoculated with 1 l of a fungal starter mix of selected autochthonous fungal species. A portion of the original membrane (M0) was analyzed at the initial time of the experiment (T0) to evaluate any pre-existing heavy metals contaminations (M0T0).
Fungal starter mixes were prepared using Malt Extract liquid (ME, Sigma Chemical Co.) medium inoculated with the conidia of the selected species. The conidia were counted with a Burker chamber to quantify the inoculum (8 × 105 conidia ml−1). The fungal mix was cultivated in a rotary shaker with 140 rpm rotation speed at 24 °C for 3 to 5 days to form ripe pellets (Wang et al. 2013) (Fig. 2). The boxes were stored at 19 °C and maintained at 60% relative humidity.

Penicillium expansum and Paecilomyces formosus pellets in Malt Extract (ME) medium after 3 days
Samples of sediment and colonized membrane were collected from the boxes in order to evaluate their metal content after 15 (T1), 30 (T2), and 60 (T3) days. The concentration of metals was measured in triplicate for quality control and the percentage coefficients of the relative standard deviation were typically below 10%, only reaching maximum values of about 25% only for those concentrations close to the detection limit of elements. Due to the high standard deviation values of some metals, only the most abundant ones (Al, Cu, Zn, Fe, and Ni) were studied. The fungal bioaccumulation capability was evaluated by computing the recovery efficiency (RE), defined as the percentage ratio between the element concentration in the membrane (eM) and the initial element concentration in the sediment (eS0): RE = (eM ÷ eS0) × 100 (Di Piazza et al. 2017). The control membrane RE value (M0T0) represents the natural RE of the membrane employed in this study. Moreover, RE was calculated for each inoculated box after 15 (T1), 30 (T2), and 60 (T3) days (M1T1, M1T2, M1T3 and M2T1, M2T2, and M2T3, respectively). These values represent the RE given by the membrane absorption (M0T0) and the activity of the fungi plus the membrane system. Finally, the difference between all the RE values of the membrane inoculated with fungi (M1T1, M1T2, M1T3, M2T1, M2T2, and M2T3) and the RE values of the control membrane (M0T0) was calculated for box 1 and 2 (D1T1, D1T2, D1T3 and D2T1, D2T2, D2T3, respectively). This difference shows that RE values only increase due to the bioaccumulation activity of the fungal-membrane system.
Results and discussions
The results of the initial chemical and physical analyses of the sediment are summarized in Table 1. The concentration of Ni exceeds the Italian Law limit (DMI 56/14 2009). Sediment is characterized by coarse grain size (∅ > 63 μm; 87%) instead of fine grain size (∅ < 63 μm; 13%), and, in particular, by fine sand (125 < ∅ < 250 μm). Moreover, the sediment is almost completely composed of inorganic material (IF equal to 97%). The results of both chemical and dimensional analyses of sediments at T0 agree with what was observed by Cutroneo et al. (2017) in the same part of the Port of Genoa (Fig. 1), thus showing stability in the chemical and physical characteristics of sediments in this part of the port.
The results of this study highlight how contaminated marine sediments are colonized by fungi. Among all of the isolated strains, Aspergillus, Penicillium, and Trichoderma are the most common genera in the bottom sediments, with about 150 CFUs are counted on MRBA plates (Greco et al. 2018). The majority of the isolated strains by Greco et al. 2018 are able to live in both terrestrial and marine environments; no typical and/or solely marine fungi were found. Some isolated species are known for their capability to degrade organic pollutant-contaminants and bioaccumulate heavy metals, such as Paecilomyces formosus (Aftab et al. 2017) and Penicillium expansum (Di Piazza et al. 2017; Zotti et al. 2014). Therefore, these species were selected for the mycoremediation pilot activity. Our morphological results of these species are similar to data reported in specific bibliographies (Samson et al. 2009; Domsch et al. 2007; Pitt 1979; Ram 1968). In particular, our selected strain of Paecilomyces formosus easily grows in all common laboratory media, but in particular is characterized by effuse and floccose mycelium in Czapek Dextrose Agar (CZDA), Malt Extract Agar (MEA), and Czapek Yeast Agar (CYA). Conidia are ellipsoidal to ovoid, smooth, with general sizes of (4.8–)9.1(–12) × (2.4–)4(–4.8) μm and (7.2–)9.6(–12) × (3.6–)4(–4.8) μm in MEA and CYA, respectively. Both selected species are known to colonize aquatic and marine environments: Penicillium expansum was already isolated from extreme and polluted environments, such as saline beaches, soils with high salinity, polluted streams, and river sediments (Domsch et al. 2007), while Paecilomyces formosus was isolated from marine substrates as sponges (Samson et al. 2009).
Recent studies have shown how autochthonous microfungi isolated from metal contaminated areas are characterized by a tolerance to high metals and capability to bioconcentrate (Cecchi et al. 2017a; Cecchi et al. 2017b; Zotti et al. 2014). These studies were generally carried out in vitro using sterile microporous cellophane membranes, which allow the physical (not chemical) mycelia separation from the solid media enriched with metals (Cecchi et al. 2017a; Cecchi et al. 2017b; Di Piazza et al. 2017; Zotti et al. 2014). However, the employment of this membrane in pilot mycoremediation protocols has never been investigated. The results show that selected microfungi could grow on the membrane and colonize it over 1 week. Table 2 shows the RE values of the control membrane (M0T0) and the membrane inoculated with fungi after T1, T2, and T3 in box 1 and 2 (M1T1, M1T2, M3T3 and M2T1, M2T2, M2T3, respectively). Table 2 also allows for the comparison of these data with an increase in the fungal-membrane system RE during pilot activity in box 1 and 2 (D1T1, D1T2, D1T3 and D2T1, D2T2, D2T3, respectively). The control membrane is characterized by absorption capability (Table 2). Indeed, the membrane initially absorbs metal ions in solution, while after the fungal inoculum, it appears to increase the absorption capability, owing to the active bioaccumulation and passive bioabsorption of metals in the fungal cells and on the fungal wall, respectively. Bioaccumulation is a property of living organisms and is influenced by physical, chemical, and biological mechanisms including intra- and extra-cellular processes (Fomina and Gadd 2014; Gadd et al. 2012; Dhankhar and Hooda 2011). On the other hand, bioabsorption has been defined by most researchers as a passive and metabolically-independent process (Fomina and Gadd 2014; Malik 2004) and can be performed using dead biomass or fragments of cells and tissues. The majority of metal removal studies choose to use the dead biomass strategy (bioabsorption; Fomina and Gadd 2014); however, the living microorganism alternative not only allows for the sorption, transport, complexation, and transformation of metals and metalloids but also promotes the degradation of organic pollutants to contribute to the overall decontamination process (Svobodová and Novotný 2018).
The data of this study show that the five analyzed metals were absorbed, albeit to a different degree, from the complex membrane plus fungi (Table 2; Fig. 3). An increasing uptake of metals by the membrane-fungi system from T1 to T3 is highlighted in the case of Zn in box 1 and 2 and Al and Ni in box 2 (Table 2). Results suggest that the metal uptake was highest after 60 days (T3) for all the metals except for Cu in box 2, for which it was highest after 30 days (T2) (Table 2 and Fig. 3). Moreover, results display how the membrane uptake of metals by fungi increased particularly for Cu and Zn (RE was equal to 75% and 207%, respectively). These are considered two of the most studied metals regarding fungal bioaccumulation and bioabsorption in vitro (Cecchi et al. 2018; Goltapeh et al. 2013; Zotti et al. 2014). Cu and Zn have a biological role in fungal metabolism and are characterized by specific metal-binding compounds. In particular, Cu detoxification in fungal cells depends on sequestration in the cytosol by induced molecules, which includes low-molecular-weight cysteine-rich proteins (metallothioneins) and peptides derived from glutathione (phytochelatins). On the other hand, Zn shows a higher affinity for organic acids (e.g., citric and ossalic acids) (Goltapeh et al. 2013). However, the majority of applied works deal with the employment of fungal mycelium in wastewater treatment (Svobodová and Novotný 2018), whereas few studies have investigated the potential role of fungi in soil and sediment decontamination. Li et al. (2014) show how Penicillium chrysogenum Thom and rice straw are promising method against metal bioavailability (e.g., Cu and Cd) and improve soil microbial properties and plant yield in heavy-metal-polluted agricultural soils. Nevertheless, one unsolved problems is represented by the possibility to recover fungal mycelia after metal mycoremediation treatment. In fact, while organic pollutants may be degraded into minor toxicity compounds, heavy metals still persist in the environment. Many studies show how autochthonous fungi may stabilize toxic metals in situ (Li and Yu 2015), yet when contamination degree is too high, metal removal represents the only solution.

Recovery efficiency (RE, in %) of metals given by the fungal-membrane system of box 1 (D1) and box 2 (D2) during the different times (T1, T2, and T3) of the mycological pilot activity
Conclusion
There is a need for the application of a new green remediation technique instead of the traditional methods for the global problem of dredged sediments. The mycological pilot activity carried out in this study demonstrates that port dredged sediments can be treated by using mycoremediation protocols. Currently, the application of biological strategies, such as fungal strategies, for metal removal from the sediments is complicated by various problems (e.g., time for pilot scale studies, research, development, and technology commercialization) (Brierley 2010). However, the complex of Penicillium expansum Link and Paecilomyces formosus (Sakag., May. Inoue & Tada) Houbraken & Samson selected after the fungal community characterization of the Port of Genoa (Greco et al. 2018), with the cellophane membrane leading to sediment decontamination. The five metals analyzed in this study (Al, Fe, Cu, Zn, and Ni) were absorbed with varying degrees; in particular, Cu and Zn were actively removed (RE was equal to 75% and 207%, respectively). Metal membrane uptake was highest after 60 days of fungal activity. After the removal of the membrane, the special waste requires disposal. However, the membrane could undergo a mycomining process to recover metals with economic interest and value. The exploitation of fungi that show high adaptation, biodegradation, and accumulation capacity not only represents an innovative and sustainable possible solution but also allows for the reduction in the costs of transporting and disposing sediments ex situ. Moreover, the creation of a consortium that sums metal absorption by an absorbent membrane, together with active fungi bioaccumulation, leads to a more efficient decontamination system and a lower amount of special wastes.
References
Abdollahi S, Raoufi Z, Faghiri I, Savari A, Nikpour Y, Mansouri A (2013) Contamination levels and spatial distributions of heavy metals and PAHs in surface sediment of Imam Khomeini Port, Persian Gulf, Iran. Mar Pollut Bull 71:336–345
Aftab K, Akhtar K, Kausar A, Khaliq S, Nisar N, Umbreen H, Iqbal M (2017) Fungal strains isolation, identification and application for the recovery of Zn (II) ions. J Photochem Photobiol B 175:282–290
Akcil A, Erust C, Ozdemiroglu S, Fonti V, Beolchini F (2015) A review of approaches and techniques used in aquatic contaminated sediments: metal removal and stabilization by chemical and biotechnological processes. J Clean Prod 86:24–36
Bensch K, Groenewald JZ, Dijksterhuis J, Starink-Willemse M, Andersen B, Summerell BA, Shin HD, Dugan FM, Schroers HJ, Braun U, Crous PW (2010) Species and ecological diversity within the Cladosporium cladosporoides complex (Davidiellaceae, Capnoidiales). Stud Mycol 67(1):1–94
Bensch K, Braun U, Groenewald J, Crous P (2012) The genus Cladosporium. Stud Mycol 72:1–401
Bhakuni DS and Rawat DS (2006) Bioactive Marine Natural Products. Springer Science & Business Media.
Bissett J (1984) A revision of the genus Trichoderma. I. Section Longibrachiatum sect. Nov. Can J Bot 62(5):924–931
Bissett J (1991a) A revision of the genus Trichoderma. II. Infrageneric classification. Can J Bot 69(11):2357–2372
Bissett J (1991b) A revision of the genus Trichoderma. III. Section Pachybasium. Can J Bot 69(11):2373–2417
Bissett J (1991c) A revision of the genus Trichoderma. IV. Additional notes on section Longibrachiatum. Can J Bot 69(11):2418–2420
Brierley CL (2010) Biohydrometallurgical prospects. Hydrometallurgy 104(3-4):324–328
Buruaem LM, Hortellani MA, Sarkis JE, Costa-Lotufo LV, Abessa DMS (2012) Contamination of port zone sediments by metals from Large Marine Ecosystems of Brazil. Mar Pollut Bull 64:479–488
Capello M, Cutroneo L, Consani S, Dinelli E, Vagge G, Carbone C (2016) Marine sediment contamination and dynamics at the mouth of a contaminated torrent: the case of the Gromolo Torrent (Sestri Levante, north-western Italy). Mar Pollut Bull 109:128–141
Caplat C, Texier H, Barillier D, Lelievre C (2015) Heavy metals mobility in harbor contaminated sediments: the case of Port-en-Bessin. Mar Pollut Bull 50:504–511
Cecchi G, Marescotti P, Di Piazza S, Zotti M (2017a) Native fungi as metal remediators: silver mycoaccumulation from metal contaminated wasterock dumps (Libiola Mine, Italy). J Environ Sci Health B 52(3):191–195
Cecchi G, Roccotiello E, Di Piazza S, Riggi A, Mariotti MG, Zotti M (2017b) Assessment of Ni accumulation capability by fungi for a possible approach to remove metals from soils and waters. J Environ Sci Health B 52(3):166–170
Cecchi G, Marescotti P, Di Piazza S, Lucchetti G, Mariotti MG, Zotti M (2018) Gypsum biomineralization in sulphide-rich hardpans by a native Trichoderma harzianum Rifai strain. Geomicrobiol J 35(3):209–214
Coates JD, Woodward J, Allen J, Philp P, Lovley DR (1997) Anaerobic degradation of polycyclic aromatic hydrocarbons and alkanes in petroleum-contaminated marine harbor sediments. Appl Environ Microbiol 63(9):3589–3593
Crous PW, Braun U, Schubert K, Groenewald JZ (2007) Delimiting Cladosporium from morphologically similar genera. Stud Mycol 58(1):33–56
Cruz A, Anselmo AM, Suzuki S, Mendo S (2015) Tributyltin (TBT): A review on microbial resistance and degradation. Environ Sci Technol 45(9):970–1006
Cutroneo L, Massa F, Castellano M, Canepa G, Costa S, Povero P, Tucci S, Capello M (2014) Technical and public approaches to involve dredging stakeholders and citizens in the development of a port area. Environ Earth Sci 72:3159–3171
Cutroneo L, Castellano M, Carbone C, Consani S, Gaino F, Tucci S, Magrì S, Povero P, Bertolotto RM, Canepa G, Capello M (2015) Evaluation of the boundary condition influence on PAH concentrations in the water column during the sediment dredging of a port. Mar Pollut Bull 101:583–593
Cutroneo L, Carbone C, Consani S, Vagge G, Canepa G, Capello M (2017) Environmental complexity of a port: evidence from circulation of the water masses, and composition and contamination of bottom sediments. Mar Pollut Bull 119:184–194
Damare S, Singh P, Raghukumar S (2012) Biotechnology of marine fungi. Prog Mol Subcell Biol 53:277–297
Dermont G, Bergeron M, Mercier G, Richer-Lafleche M (2008) Soil washing for metal removal: a review of physical/chemical technologies and field applications. J Hazard Mater 152:1–31
Dhankhar R, Hooda A (2011) Fungal biosorption–an alternative to meet the challenges of heavy metal pollution in aqueous solutions. Environ Technol 32(5):467–491
Di Piazza S, Cecchi G, Cardinale AM, Carbone C, Mariotti MG, Giovine M, Zotti M (2017) Penicillium expansum Link strain for a biometallurgical method to recover REEs from WEEE. Waste Manag 60:596–600
Domsch KH, Gams W, Anderson TH (2007) Compendium of soil fungi, 2nd taxonomically revised edition by W." Gams. IHW, Eching.
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissues. Phytochem Bull 19:11–15
Falandysz J, Mędyk M, Treu R (2018) Bio-concentration potential and associations of heavy metals in Amanita muscaria (L.) Lam. from northern regions of Poland. Environ Sci Pollut Res 25:25190–25206
Fathollahzadeh H, Kaczala F, Bhatnagar A, Hogland W (2014) Speciation of metals in contaminated sediments can enhance metal mobility due to changes of bacterial diversity. Water Res 68:637–650
Fomina M, Gadd GM (2014) Biosorption: current perspectives on concept, definition and application. Bioresour Technol 160:3–14
Fonti V, Beolchini F, Rocchetti L, Dell'Anno A (2015) Bioremediation of contaminated marine sediments from Oskarshamn Harbor, Oskarshamn, Sweden. Environ Sci Pollut R 21:2455–2464
Gadd GM, Rhee YJ, Stephenson K, Wei Z (2012) Geomycology: metals, actinides and biominerals. Environ Microbiol Rep 4(3):270–296
Garcia-Orellana J, Cañas L, Masqué P, Obrador B, Olid C, Pretus J (2011) Chronological reconstruction of metal contamination in the Port of Maó (Minorca, Spain). Mar Pollut Bull 62:1632–1640
Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts. Mol Ecol 2(2):113–118
Glass NL, Donaldson GC (1995) Development of primer sets designed for used with the PCR to amplify conserved genes from filamentous ascomycetes. Appl Environ Microbiol 61(4):1323–1330
Goltapeh EM, Danesh YR, Varma A (2013) (Eds.). Fungi as bioremediators (Vol. 32). Springer Science & Business Media.
Greco G, Cecchi G, Di Piazza S, Cutroneo L, Capello M, Zotti M (2018) Fungal characterisation of a contaminated marine environment: the case of the Port of Genoa (North-Western Italy). Webbia 73(1):97–106
Howe P, Watts P (2005) Tin and inorganic tin compounds. Concise International Chemical Assessment Document 65. World Health Organization, pp. 81.
Iannelli R, Bianchi V, Macci C, Peruzzi E, Chiellini C, Petroni G, Masciandaro G (2012) Assessment of pollution impact on biological activity and structure of seabed bacterial communities in the Port of Livorno (Italy). Sci Total Environ 426:56–64
Italian Ministerial Decree 56 of 14 April 2009. Criteri tecnici per il monitoraggio dei corpi idrici – Articolo 75, Dlgs 152/2006.
Johnson DB (2013) Development and application of biotechnologies in the metal mining industry. Environ Sci Pollut R 20:7768–7776
Klich MA (2002) Identification of common Aspergillus species, Utrecht, Netherlands: Centraalbureau voor Schimmelcultures 116 pp..
Kohlmeyer J, Kohlmeyer E (1979) Marine Mycology: The Higher Fungi. Academic Press, New York, p 690
Li WW, Yu HQ (2015) Stimulating sediment bioremediation with benthic microbial fuel cells. Biotechnol Adv 33:1–12
Li M, Tian X, Liu RZ, Chen WL, Cai P, Rong XM, Dai K, Huang QY (2014) Combined application of rice straw and fungus Penicillium Chrysogenum to remediate heavy-metal-contaminated. Soil Sediment Contam 23(3):328–338
Malik A (2004) Metal bioremediation through growing cells. Environ Int 30(2):261–278
Mulligan CN, Yong RN, Gibbs BF (2001) An evaluation of technologies for the heavy metal remediation of dredged sediments. J Hazard Mater 85:145–163
OSPAR Commission (2014) OSPAR Guidelines for the Management of Dredged Material at Sea. Draft Summary Record - EIHA, Annex 7, pp. 34. https://dredging.org/media/ceda/org/documents/guidance/ospar/ospar-dredged-materialguidelines_for%20london-2014.pdf
Peng J-F, Song Y-H, Yuan P, Cui X-Y, Qiu G-L (2009) The remediation of heavy metals contaminated sediment. J Hazard Mater 161:633–640
Pitt JL (1979) The genus Penicillium and its teleomorphic states Eupenicillium and Talaromyces. Academic Press, New York, 634 pp..
Ram C (1968) Timber-attacking fungi from the State of Maranhao, Brazil. Some new species of Paecilomyces and its perfect stage Byssochlamys Westl. VIII. Nova Hedwigia 16:305–314
Raper KB, Fennel DI (1965) The genus Aspergillus. The Williams & Wilkins Co., Baltimore, 686 pp..
Rifai MA (1969) A revision of the genus Trichoderma. Mycol Pap 166:1–56
Roccotiello E, Marescotti P, Di Piazza S, Cecchi G, Mariotti MG, Zotti M (2015) Biodiversity in metal contaminated sites—problem and perspective—a case study. In: Lo YH, Blanco JA, Roy S (eds) Biodiversity in Ecosystems—Linking Structure and Function. InTech, Rijeka, pp 581–600
Ruiz CE, Schroeder PR (2007) Capping Design Approaches for Contaminated Sediments and Dredging Residuals: Engineered Reduction in Bioavailability. Fifth LACCEI International Latin American and Caribbean Conference for Engineering and Technology (LACCEI’2007), “Developing Entrepreneurial Engineers for the Sustainable Growth of Latin America and the Caribbean: Education, Innovation, Technology and Practice”. 29 May – 1 June 2007, Tampico, México.
Samson RA, Houbraken J, Varga J, Frisvad JC (2009) Polyphasic taxonomy of the heat resistant ascomycete genus Byssochlamys and its Paecilomyces anamorphs. Persoonia 22:14–27
Samson RA, Houbraken J, Rane U, Frisvad JC, Andersen B (2010) Food and indoor fungi. CBS Laboratory Manual Series 2. 390 p. ISBN: 978-90-70351-82-3.
Samson SA, Varga J, Frisvad JC (2011) Taxonomic studies on the genus Aspergillus. Stud Mycol 69:1–97
Svobodová K, Novotný Č (2018) Bioreactors based on immobilized fungi: bioremediation under non-sterile conditions. Appl Microbiol Biotechnol 102:39–46
Tabak HH, Lens P, Hullebusch ED, Dejonghe W (2008) Developments in bioremediation of soils and sediments polluted with metals and radionuclides. Microbial processes and mechanisms affecting bioremediation of metal contamination and influencing metal toxicity and transport. Rev Environ Sci Biotechnol 4:115–156
Tavakoly Sany SB, Salleh A, Rezayi M, Saadati N, Narimany L, Tehrani GM (2013) Distribution and contamination of heavy metal in the coastal sediments of port Klang, Selangor, Malaysia. Water Air Soil Pollut 224(4):1–18
U.S. EPA (1996) Method 3050B: Acid Digestion of Sediments, Sludges, and Soils. Revision 2. Washington, DC, pp. 12.
U.S. EPA. (2014) Method 6010D (SW-846): Inductively Coupled Plasma-Atomic Emission Spectrometry. Revision 4. Washington, DC, pp. 35.
Vagge G, Cutroneo L, Castellano M, Canepa G, Bertolotto RM, Capello M (2018) The effects of dredging and environmental conditions on concentrations of polycyclic aromatic hydrocarbons in the water column. Mar Pollut Bull 135:704–713
Walker TR, MacAskill D, Weaver P (2013a) Environmental recovery in Sidney Harbour, Nova Scotia: evidence of natural and anthropogenic sediment capping. Mar Pollut Bull 74:446–452
Walker TR, MacLean B, Appleton R, McMillan S, Miles M (2013b) Cost-effective sediment dredge disposal options for small craft harbours in Canada. Remediat J 23(4):123–140
Wang JN, Li A, Yang JX, Wang JH, Guo JB, Ma F, Shi SN, Zhang S, Ren NQ (2013) Mycelial pellet as the biomass carrier for semi-continuous production of bioflocculant. RSC Adv 3:18414–18423
Wang L, Chen L, Tsang DCW, Li J-S, Baek K, Hou D, Ding S, Poon C-S (2018) Recycling dredged sediment into fill materials, partition blocks, and paving blocks: technical and economic assessment. J Clean Prod 199:69–76. https://doi.org/10.1016/j.jclepro.2018.07.165
White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: PCR protocols: a guide to methods and applications, pp 315–322
Zotti M, Di Piazza S, Roccotiello E, Lucchetti G, Mariotti MG, Marescotti P (2014) Microfungi in highly copper-contaminated soils from an abandoned Fe–Cu sulphide mine: growth responses, tolerance and bioaccumulation. Chemosphere 117:471–476
Funding
The present study was funded by the European Interreg Italy-France 2014-2020 Maritime Project SEDITERRA “Guidelines for the sustainable treatment of dredged sediments in the Marittimo area” (CUP I42F17000010006).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Elena Maestri
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Cecchi, G., Vagge, G., Cutroneo, L. et al. Fungi as potential tool for polluted port sediment remediation. Environ Sci Pollut Res 26, 35602–35609 (2019). https://doi.org/10.1007/s11356-019-04844-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-019-04844-5