
Abstract
Many coastal rivers have a system of cascade reservoirs, but the role of these reservoirs in regulating nutrient transport from watershed to coast is still unknown. In this study, phosphorus (P) in surface water and top sediment was investigated along the North Jiulong River (southeast China) under three hydrological conditions (high flow, medium flow and low flow) in 2012–2013, and P dynamics in a cascade reservoir (Xipi Reservoir) were studied on a monthly scale. Results showed that the concentrations of dissolved reactive phosphorus (DRP) consistently decreased longitudinally in the upper river with the lowest values observed in the section of cascade reservoirs, likely due to tributary inputs and in situ uptakes. The decrease was most rapid during base flow when DRP was highest in the free-flowing river section and lowest in cascade reservoirs. Results from monthly monitoring on the Xipi Reservoir showed general downriver decreases in DRP, total particulate phosphorus (TPP) and total phosphorus (TP) in the riverine zone and transition zone. Mass balance results on an annual basis suggest that the Xipi Reservoir (lacustrine zone) was an overall sink for TPP (6 % retention) but somewhat a source of DRP (−0.3 %) with TP retention (1 %). Even scaled up to the whole cascade reservoir system, P retention was low compared with worldwide reservoirs, which we ascribe to the high P loading and short hydraulic residence time. Nevertheless, major processes controlling P retention in coastal rivers with cascade reservoirs varied from sedimentation in the dry-cold season to biotic transformation in the wet-warm season, thereby affecting loading and composition of P from watershed to the coast. This study highlights the hydrological controls on the role of cascade reservoirs in regulating P retention and downriver fluxes in different seasons.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phosphorus (P) is a limiting nutrient for primary productivity in aquatic systems. Land use changes (e.g., agriculture and urbanization) and global warming can lead to increasing P loading from watersheds (Duan et al. 2012), resulting in eutrophication and other negative effects in both fresh water and coastal sea (Conley and Paerl 2009, Moss et al. 2011, Tysmans et al. 2013). Recent reports on delivery and cycling of P in rivers (Stamm et al. 2014, Withers and Jarvie 2008, Zieliński and Jekatierynczuk-Rudczyk 2015) have shown that P concentration, composition (and thus reactivity), and patterns of delivery (timing and relation to river hydrology) are largely determined by sources of P and in-stream processes. P transport from headwater to downriver depends on a variety of physico-chemical processes to different extents due to stream size, stream geomorphology and anthropogenic pressures (Withers and Jarvie 2008). For example, P fluxes are typically regulated by seasonal changes in runoff and residence time that are highly relevant to hydrological conditions (Lu et al. 2011, Oliver et al. 2014). Understanding how hydrological conditions regulate P delivery and cycling along the river channel is essential to develop proper management strategies to mitigate eutrophication problems.
More than 70,000 large dams have been built worldwide and this number will continue to increase due to growing water stress and demand for energy (Maavara et al. 2015). Dam constructions have substantially altered hydrological conditions and nutrient biogeochemical cycles and seaward fluxes (Cook et al. 2010, Humborg et al. 1997, Jin et al. 2013, Syvitski et al. 2005, Van Cappellen and Maavara 2016). Damming alters not only the quantity and timing of fluvial P fluxes to downstream aquatic systems, but also the form and bioavailability of P (Withers and Jarvie 2008). Moreover, the insertion of reservoirs into a riverine landscape may change longitudinal patterns of nutrients, but the degree of the changes depends on reservoir characteristics (size, residence time, location, ambient environments, etc.) (Stanley and Doyle 2002). Several studies have explored the impact of dam construction on P retention (Evans et al. 2004, Némery and Garnier 2007, Oliver et al. 2014, Stanley and Doyle 2002, Tysmans et al. 2013, Yao et al. 2009), but most of them targeted a single reservoir. Very little research has addressed the hydrological controls on the role of cascade reservoirs in regulating riverine P retention and fluxes except for some studies on sediment P (Liu et al. 2015, Ouyang et al. 2011). Integrating detailed information on P cycling in reservoirs into a holistic perspective of the whole river system is necessary toward improved understanding of how P is exported from watershed to coast.
Numerous dams have been constructed for the purpose of hydropower generation, irrigation and drinkable water production in subtropical mountainous rivers in Southeast China where runoff is high due to the Asian monsoon climate. The objective of this study was to examine: 1) the role of reservoirs on P fluxes as a sink, a source, or a transformer, and 2) how watershed hydrology (or stream flow) affects the role of the reservoirs. During 2012 and 2013 we carried out extensive measurements along the river network (tributary-reservoir-river) under three hydrological conditions (high flow, medium flow and low flow) in the North Jiulong River, and conducted intensive investigations in one of the reservoirs across a river-reservoir gradient. We hypothesized that hydrology controls P retention and flux from rivers via changing sedimentation, algal P uptakes or P release from sediment in the cascade reservoir.
Materials and methods
Description of the study river
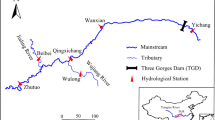
The Jiulong River is a subtropical river (drainage area 14,741 km2) located in southeast China and influenced by a monsoon climate (Fig. 1). Two main tributaries of the river (North River and West River) discharge about 12.4 billion m3 yr.−1 of water into Xiamen Bay. During the last few decades, over 120 dam reservoirs have been constructed within the watershed, with six major dams located in the middle of the main stem of the North River. The North Jiulong River has been turned into a cascade reservoir system and was selected as the study site. The North Jiulong River watershed covers four cities/countries (Longyan, Zhangping, Hua’an, Changtai) and a part of Zhangzhou City. The total population is 1.5 million, 43 % of whom live in urban areas. The other counties are predominantly agricultural and forest land and have a relatively low population density, with the exception of the more densely populated downriver area close to the estuary (Changtai and Zhangzhou) (Chen et al. 2015). Pig farming in the upper North River (Longyan City) has increased markedly since the late 1990s, but the absence of efficient treatment systems for animal wastes has resulted in large amounts of nutrient being discharged to the North River (Chen et al. 2013). Sewage and industrial waste water have been partly treated in Longyan since the early 2000s, mainly for reduction of chemical oxygen demand (COD), but the amount of nutrient discharged to waterbodies has continued to increase (Chen et al. 2013). Land use comprises 70 % forest including upland orchard, 16 % arable land, 3 % urban and residential land, 2 % water, and 1 % bare and grass land (Chen et al. 2015). The topography in the North Jiulong River watershed is mostly hilly (91 % of the catchment area has an altitude greater than 200 m, and 40 % has a slope greater than 25°). The main watershed soil type is lateritic red soil (mainly on hills), followed by paddy soil (alongside rivers) and latosolic red soil (mainly in the valley). Lateritic red soil is one of the soils forming in subtropical and tropical regions and is rich in Fe2O3/MnO2.

Map of North Jiulong River (left) and Xipi Reservoir (right) showing sampling sites
Xipi, a cascade reservoir located in the upper-middle North River, was chosen to make detailed exploration of P behavior and retention along the river-reservoir gradient (Fig. 1). Xipi Reservoir is 8.5 km long and 125 m wide on average. The mean annual air temperature is 20.9 °C and annual precipitation is about 1600 mm. According to the water depth and geomorphology, the reservoir was defined as three regions: riverine zone (4.9–8.5 km upstream of Xipi dam), transitional zone (2.5–4.9 km) and lacustrine zone (0–2.5 km). This study mainly targeted the lacustrine zone, with an open water area of 0.34 km2, mean and maximum depth of 15 m and 28 m respectively, and a capacity of 389 × 104 m3. The hydraulic residence time (HRT) of this zone varied from 4 h to 26 h during the study period, typical for a “run-of-the-river” reservoir.
Sampling and lab analysis
Discrete surface (0.5 m) water samples were collected from upstream to lower river in the North Jiulong River (Fig. 1). A total of 16 main stem sites were selected encompassing the free-flowing river section and reservoirs, and 11 sites located in the outlet of main tributaries were also sampled. Three field surveys were carried out in March 2012 (medium flow), June 2012 (high flow), and January 2013 (low flow). For the lacustrine zone of Xipi Reservoir, monthly field surveys from May 2012 through April 2013 were carried out at seven sites (X1-X7) plus one tributary site (XT) close to the upper lacustrine zone. To understand the spatial pattern of P in the whole river-reservoir system, monthly surveys were extended to include the transitional zone (X8-X12) and riverine zone (X13-X18) from November 2012 to April 2013. Water temperatures were determined in-situ using a WTW TetraCon® 325 probe. Surface sediments at sites X1-X7 (other sites had no sediment or there was too much sand for the sample to be used) were collected with a Petite Ponar grab sampler during the sampling on May and December 2012.
Surface (0.5 m) water samples were collected in polyethylene bottles and stored in a cool box and delivered to the laboratory. All samples were filtered through preweighed 0.45 μm nucleopore membranes, and frozen at −20 °C before analysis. Using a standard colorimetric method (SEPA 2002), we measured dissolved reactive phosphorus (DRP) and total P (TP), following oxidization with 4 % alkaline potassium persulfate. P concentrations are presented in μmol L−1 and detection limits were 0.03 μmol P L−1. The precision for P components was estimated by repeated determinations of 10 % of the samples and the relative error was less than 5 %. The filtered GF/F membranes were oven-dried (105 °C overnight) to constant weight, and then were analyzed for total particulate P (TPP) after being combusted in a muffle furnace (550 °C for 1.5 h) (converted all P to phosphate) and extracted with 1 mol L−1 HCl. Another set of water samples were filtered through 25 mm GF/F membranes, then the filters were extracted with 90 % acetone for measurement of Chl-a (μg L−1) using a Turner fluorescence spectrophotometer (Parsons et al. 1984).
The sediment samples were dried in a freeze-dryer, and ground to powder using a pestle and then hand-sieved through a 150 μm sieve. The fractions less than 150 μm were homogenized prior to analysis. Sediment P compositions were sequentially analyzed using the Standard Measurement and Test (SMT) procedure (Ruban et al. 1999). The SMT procedure comprises five steps corresponding to five components, i.e. NaOH-extractable P (P bound to Al, Fe and Mn oxides and hydroxides), HCl-extractable P (P associated with Ca), organic P, inorganic P, and concentrated HCl-extractable P (total P). Contents of carbon (C) and nitrogen (N) in sediment were determined using a CHNS/O elementary analyzer (PE2400 SERIES II). The relative error of the duplicate samples was less than 3 % (n = 3).
Sediment-water P exchange experiment
Measurements of sediment P exchange were separately made at two representative sites (X6 and X3) in the lacustrine zone in June (high flow) and December (low flow) 2013 using the SONE (Sediment Oxygen and Nutrient Exchanges) method (Cowan and Boynton 1996). During each sampling, sediment cores were obtained using a UWITEC Sediment corer. Overlying water from approximately 0.5 m above the sediment was prepared and stored in two collapsible, air-tight containers for laboratory incubation. Sediment cores were placed in a temperature-controlled holding tank that was set to in-situ water temperature (27.5 °C and 19.1 °C, respectively). The overlying water in the sediment cores was slowly replaced with fresh ambient bottom water without disturbing the sediments. The incubation lasted 5 h, with 30 mL water samples extracted every hour for measurement of DRP. At the time of water extraction for analysis, the displaced water volume was replaced with collected bottom water. This method has been recently applied in a joint study with Harris et al. (2015).
Data analysis and calculation
Statistical analysis (correlation, regression, ANOVA) was performed using SPSS 16.0 to explore the main factors controlling the spatial and temporal variation of P. To better interpret the results and compare spatial differences, data were divided into groups in different ways. For example, measurements of the North River were grouped into upper tributary (N1-N6), cascade dam reservoir in middle river (N7-N13) and lower river (N14-N16). The Xipi Reservoir was divided into three groups (riverine zone, transitional zone, and lacustrine zone). Temporal variation considered hydrological conditions (low flow, medium flow and high flow). Mean and standard error (±SE) represented the variation of measurements among groups.
To understand P retention by dam reservoir, a rough estimation of P retention (P ret, kg P yr.−1) from the study reservoir (lacustrine zone) was derived by monthly P input (P load) and output (P out) in terms of P from (TP, TPP, DRP), as shown in equation (1), and monthly P retention rate (P rate, %) was calculated using equation (2).
Where C li is the observed monthly P forms (TP, DRP, TPP) concentration (mg P L−1) at the upper site of the lacustrine zone (X7), C oi is observed monthly concentration (mg P L−1) at the furthest downstream site (X1), and Q i is monthly discharge (m3 s−1). In this study, monthly measurement and daily mean flow rate obtained from Xipi hydropower station on samplings dating from May 2012 through April 2013 were used. Given the small size of the reservoir, P load from surface runoff, groundwater discharge and atmospheric deposition over the lacustrine zone were omitted from the P budget. Therefore, P retention was expected to be slightly underestimated. The P budget was calculated month by month but was summed as annual P retention.
Flux of DRP across the sediment-water interface was calculated for each core using equation (3) (Bailey et al. 2011).
where F is flux of sediment DRP [mg P (m2·d)−1], k is the slope of DRP concentration vs. time plot [mg P (L·min)−1], h is the height of overlying water (m). The criteria for accepting, rejecting and modifying the k value refers to Bailey et al. (2011).
We simply estimated P retention by algal uptake relating to phytoplankton growth from the study reservoir (lacustrine zone) by assuming a P:Chl-a ratio (w/w) of 1.384:1. This ratio was derived from N:Chl-a (w/w) = 10 (Malone 1992, Miller et al. 2006) and N:P = 16 (Redfield et al. 1963). Regardless of the species, growth cycle of phytoplankton was assumed to be 10 days (Coles and Jones 2000, Liu et al. 2011). Based on the mean depth of the euphotic layer (here 4 m on average), water area (0.34 km2) and monthly measured Chl-a concentration, total P absorption by phytoplankton growth (kg P yr.−1) was calculated in kg P yr.−1.
Results
Spatio-temporal variations of DRP and Chl-a along the north Jiulong River
DRP concentration in the North River (N1-N16) varied from 0.4 to 17.9 μmol L−1, but showed a consistent longitudinal pattern for all three seasons (Fig. 2). In the section above the cascade reservoir (upper river), DRP concentrations consistently declined downriver across the three seasons. Relative to the upper river (N1-N6, 7.9 ± 1.1 μmol L−1), DRP concentrations in the section with six cascade reservoirs (N7-N13, 1.2 ± 0.2 μmol L−1) were far lower, and showed less spatial variation. DRP concentrations increased slightly in the lower river (sites N14-N16 below the cascade reservoirs, 2.1 ± 0.2 μmol L−1) with the highest values occurring at the second sites. DRP in the tributaries was scattered and in the range 0.2–14.1 μmol L−1, but generally followed the longitudinal trend of the main stem (Fig. 2).

Change of DRP and Chlorophyll-a (Chl-a) concentrations along the North River (N1-N16) across varying hydrological conditions. The river discharge at Zhangping hydrological gauge (close to site N6, 142 km to river outlet) around the sampling date was about 25 m3 s−1 (January 2013), 500 m3 s−1 (June 2012), and 350 m3 s−1 (March 2012). The major dams XQ, XP, ML, HA, TG, LS and JD are Xiaoqi, Xipi, Mianliang, Hua’an, Tiangong, Lishui and Jiangdong, respectively
DRP concentration at the reservoir sites (N7-N13) showed a contrasting seasonal pattern to free-flowing river sites (N1-N6) across the three hydrological conditions (Fig. 2a). At the river sites, DRP concentrations during the low flow period (11.0 ± 2.5 μmol L−1 in upper river and 2.6 ± 1.0 μmol L−1 down river) were significantly higher than during the high flow period (5.0 ± 0.8 μmol L−1 and 2.0 ± 0.2 μmol L−1, respectively). In contrast, DRP at reservoir sites during the low flow period (0.6 ± 0.1 μmol L−1) was lower than that during the high flow period (1.5 ± 0.1 μmol L−1) (Fig. 2a). As a result, the largest longitudinal change in DRP concentration occurred during the low flow period in January 2012, while longitudinal changes during the high flow period in June 2012 were relatively less.
Chl-a concentration ranged from 0.8 to 20.2 μg L−1 in the North River (N1-N16). In contrast to DRP, no downriver decreases in Chl-a concentrations were observed in the upper river, and there was no significant difference between the upper river sites (2.5 ± 1.1 μg L−1) and cascade reservoir sites (2.7 ± 1.8 μg L−1) (Fig. 2b, Table 1). Chl-a values were significantly higher at the downriver sites (8.8 ± 7.4 μg L−1) regardless of hydrological conditions. Chl-a concentrations were lowest during the high-flow condition across the whole river sections. The highest Chl-a concentrations at the last two lower river sites were observed during the lowest flow conditions, while the highest Chl-a concentrations at the cascade reservoir sites occurred during medium flows (Fig. 2b and Table 1).
Monthly variations of P and Chl-a in Xipi reservoir
Monthly measurement provided more details on seasonal changes in river discharge, water temperature, P and Chl-a concentration in the lacustrine zone of Xipi Reservoir (Fig. 3). While temperature was highest in July and lowest in December, flow was highest May and June 2012 and lowest from October 2012 to March 2013. DRP concentration was spiky and showed no clear seasonal trend or correlation with flow or temperature (Fig. 3a). TPP concentration (0.59–3.46 μmol L−1) decreased from the highest value in May (3.04 ± 0.11 μmol L−1), when river flow was highest, to the lowest in October (0.66 ± 0.02 μmol L−1); peak values were also observed in January (2.00 ± 0.06 μmol L−1) when there was a minor flow peak (Fig. 3b). The seasonal pattern of TP generally followed that of TPP but was also affected by DRP. The proportion of DRP in TP was 18 %–66 %, with more than six months exceeding 50 % in the study period. TPP accounted for 30 %–79 % of TP, with the highest value in May and lowest value in February. Chl-a had a peak average value of 18.7 μg L−1 in July when lowest DRP was observed, but Chl-a and DRP were not correlated in general (Fig. 3d). Interesting, the two Chl-a concentration peaks in July and February followed the peaks in TPP concentration in May/June and January.

Changes in concentrations of phosphorus forms and Chl-a in lacustrine zone of Xipi Reservoir in different months. In Fig. (d), temperature was monthly mean water temperature of the seven sites, and discharge was monthly mean discharge of Xipi Dam
Along the river-reservoir gradient (X1-X18), DRP, TP, and TPP concentrations all showed an apparently declining trend during this period (Fig. 4a-c). In particular, DRP, TPP and TP declined from the riverine zone (1.34 ± 0.13, 1.71 ± 0.08, 2.83 ± 0.10 μmol L−1, respectively) to transitional zone (1.20 ± 0.15, 1.50 ± 0.12, 2.64 ± 0.11 μmol L−1) and lacustrine zone (1.19 ± 0.13, 1.30 ± 0.07, 2.57 ± 0.12 μmol L−1) (Fig. 4a-c). An exception occurred in February when elevated values of DRP and TP concentrations in the lacustrine zone (mean DRP was 2.38 μmol L−1and TP was 3.59 μmol L−1) were recorded relative to the riverine zone (mean 1.81 and 3.35 μmol L−1) and transitional zone (mean 1.47 and 3.04 μmol L−1). Chl-a concentrations also decreased downstream in the riverine zone and transitional zone, but downstream increases in Chl-a occurred in the lacustrine zone (Fig. 4d). Chl-a changed from 3.6 ± 0.3 μg L−1 in the riverine zone to 3.1 ± 0.3 μg L−1 in the transitional zone and 4.3 ± 0.5 μg L−1 in the lacustrine zone (Fig. 4d).

Spatial variation of phosphorus and Chl-a along the river-reservoir gradient in Xipi Reservoir during the period November 2012 to April 2013. XT in the x-axis shows the tributary site close to the upper lacustrine zone
Sediment P content in the lacustrine zone
Sediment P content and composition varied between May and December (Fig. 5). Fe/Al-P content in May (432 ± 57 mg kg−1) and percentage (58 %) was significantly lower than in December (644 ± 41 mg kg−1 and 69 %) (Fig. 5b), while OP content and percentage in May (198 ± 13 mg kg−1 and 26 %) was greater than in December (161 ± 11 mg kg−1 and 17 %) (Fig. 5d). Ca-P changed little both in content (124–134 mg kg−1) and fraction (14 %–16 %) between May and December (Fig. 5c).

Spatial and temporal distributions of TP, Fe/Al-P, Ca-P and OP contents in lacustrine zone of Xipi Reservoir sediments. Inset pies show the average percentage compositions of sediment P forms in May and December 2012
Overall, the spatial pattern of sediment TP was shaped by Fe/Al-P because it was the dominant form (Fe/Al-P accounted for more than 50 % of TP). Under high flow condition (May, 2012), content of all P forms in sediment increased longitudinally, but declined (except Ca-P) at site X1, which is close to the dam (Fig. 5a). A minor spatial variability of sediment P was observed in the low flow period (December, 2012).
Sediment-water flux of DRP in the lacustrine zone
Under high flow condition (June 2013), the flux of sediment DRP at site X6 was −31.2 mg (m2·d)−1 (negative value means P diffusion from water column into sediment). The result from site X3 was not statistically significant but DRP concentration in the water column decreased over the incubation period compared with the initial concentration (Fig. 6a). In contrast, under low flow condition (December 2013), the flux of sediment DRP at site X3 was 5.1 mg (m2·d)−1 (positive value means P release from sediment to water column). The result from site X6 was not statistically significant but DRP concentration in water column over the incubation period was greater than the initial value (Fig. 6b).

Temporal variation of DRP concentrations in the overlying water and sediment-water fluxes. Open circles represent outliers based on data modifying criteria as defined by Bailey et al. (2011)
Annual P budget for the lacustrine zone
The annual TP input and output of the lacustrine zone was estimated to be 3501 × 103 kg P and 3466 × 103 kg P. In this case, the annual TP retained (total P removal) was 35 × 103 kg P, only 1 % of TP load (Table 2). TP retention was larger in June (21 % of TP input), and lower in September (−38 % of TP input). In terms of DRP, a slight additional release (−0.3 % of DRP input) was observed at an annual scale. More DRP was retained in November (68 % of DRP input), but a large fraction of DRP (−85 % of DRP input) was released in January, and as much as 115 × 103 kg TPP was retained, approximately 6 % of TPP input. A larger fraction of TPP was retained in April (24 % of TPP input) but less in July (−50 % of TPP input). An annual retention of 6 % for TPP was estimated. The annual P removal by phytoplankton was roughly estimated to be 3.40 × 103 kg P, accounting for 10 % of annual total P retention.
Discussion
Controls on P delivery in free-flowing river sections
It seems that longitudinal patterns of DRP in free-flowing river sections were largely shaped by watershed pollution influenced by tributary inputs. As previously reported (Chen et al. 2013, Chen et al. 2012), the headwater sites of the upper river (N1-N4) receive high P loads from human and animal waste. This is consistent with the highest DRP concentrations in the tributary and main stem of this river section in this study (Fig. 2a). The large anthropogenic P loading have also resulted in a high sediment P content in upper river sites (Lu et al. 2013). Moreover, the sharp decline of DRP along the river channel (N5-N7) can be ascribed to the “dilution” by tributaries water with low DRP concentration (mean DRP concentration of NA3, NA4 and NA5 was 1.42 μmol L−1, 0.85 μmol L−1 and 0.91 μmol L−1, respectively) (Fig. 2a). Similarly, the lowest DRP concentrations at the reservoir sites (N7-N13) were consistent with the lowest DRP concentrations in tributaries in this river segment, and the increases in DRP concentrations below the reservoirs coincided with higher DRP concentrations in tributaries in this river segment than the last. Relative to watershed landuse and tributary inputs, the effect of phytoplankton uptake on the longitudinal DRP pattern seemed less important. For example, Chl-a concentrations in the upper river and reservoir sites were low, although there were large decreases in DRP, while DRP increased in the lower river despite elevated Chl-a concentrations (Fig. 2). Distinguishing the effect on longitudinal P variability of cascade reservoirs from that of watershed landuse along this river warrants further investigation.
Controls on temporal changes in DRP concentration in free-flow river reaches seemed influenced by the river hydrological regime. DRP concentration in upper river sites was highest in low flow conditions and lowest in high flow conditions, suggesting dominance of point sources and storm water dilution due to the monsoon climate. This is consistent with the fact that very little wastewater is treated in the watershed and point pollution was the main source of DRP in the North River (Chen et al. 2013, Chen et al. 2012). Interestingly, different seasonal patterns of DRP were observed at reservoir sites (Fig. 2a). The contrasting seasonal changes in DRP concentration between river sites (N1-N6) and dam reservoir sites (N7-N13) suggest different controls: hydrological dilution in the river section versus in situ transformation in the cascade reservoirs.
Hydrological controls on P transformation in Xipi reservoir
It is known that spatiotemporal variability of P in reservoirs is a combined effect of physical (e.g., sedimentation) and biogeochemical (e.g., algal uptake, remineralization and sediment release) processes that is a function of river hydrology. A large amount of sediment and particulate matter can be transported from the catchment into reservoirs and a fraction can be retained in reservoirs due to reduced flow. In this study, mean TPP concentration in surface water of the lacustrine zone was positively correlated with the discharge (r = 0.75, p < 0.01), suggesting hydrological control on TPP. In high flow conditions (for example in May), a large amount of particle-bound P might be washed from upland soil by strong rainfall (our investigation was conducted after a heavy rainfall event in May 2012), and the peak values of TPP concentrations in the reservoir corresponded to the heavy precipitation event in the watershed (Fig. 3d). In addition, we found a trend of increasing TPP retention with flow, probably because coarser particles during high flows can be easily and readily retained. The negative correlation between surface water TPP concentration and the sediment Fe/Al-P (r = 0.82, p < 0.05) supported the occurrence of coarser particles during high flows, because coarse particles were generally depleted in Fe/Al-P. Moreover, the effect of sedimentation on TPP retention was also supported by the longitudinal decreases in TPP concentrations across the Xipi Reservoir, especially in the river and transition zones during low flow conditions (Fig. 4b). The occurrence of these longitudinal decreases in TPP can be attributed to reduced flow across the river-reservoir gradient.
Hydrological controls on P retention via biological processes were illustrated by changes in Chl-a concentration. Algal uptake of DRP, which is desorbed from particles, released from sediment or regenerated from organic matter decomposition, is one of several critical processes occurring in the lacustrine zone (Essington and Carpenter 2000, Sterner 2008). In this study, we found Chl-a concentration in the lacustrine zone of Xipi Reservoir was higher in July (18.7 ± 0.9 μg L−1) and February (8.2 ± 1.5 μg L−1) (Fig. 3d), one to two months after the peak values of TPP concentrations (Fig. 3b). Most likely, the elevated Chl-a concentrations in the Xipi Reservoir were the result of enhanced P inputs from watershed sources that were mobilized by precipitation events, because P is the limiting nutrient in freshwater systems. In addition to this watershed-reservoir coupling, algal uptake of DRP also occurred in the reservoir, supported by the relationship between Chl-a and DRP and P mass balance. For example, both Chl-a (Fig. 3d) and most retention rates of DRP were higher in medium flow (1 %–36 % of DRP input), and there was a negative correlation between Chl-a and DRP (r = 0.80, p < 0.001) during this period. Moreover, elevated water temperature may favor algal growth and uptake of DRP, as the measured data in July and February suggested (Fig. 3d). Unlike medium flow, DRP uptake in the high flow period might be limited because algal growth could be restrained (lowest Chl-a in May and June 2012 when discharges were fairly high, Fig. 3d), probably due to high turbidity and short water residence time (HRT was 4 h). Under low flow conditions, long water residence time (HRT over 10 h) was expected to promote DRP uptake due to algal growth, and a negative correlation between Chl-a and DRP (r = 0.52, p < 0.001) was also observed in low flow conditions.
DRP release from sediments or regeneration from organic P also seemed to vary with hydrological condition. The occurrence of DRP release from sediments in the lacustrine zone was possible because there was more Fe/Al-P (usually bound with fine particles) (Fig. 5a), possibly as a result of sediment sorting along the river-reservoir transit (Horowitz and Elrick 1987). Release of DRP from sediment to water column may more likely occur during the low flow period (December 2012), shown as a positive flux of DRP during the incubation experiment (Fig. 6b), because both magnitude and fraction of Fe/Al-P in surface sediments in the low flow period (December 2012) was generally greater than in the high flow period (May 2012) (Fig. 5a). Other studies also showed that DRP release can also be enhanced under lowered redox potential and anoxic conditions, which generally occurs during the low flow period (Boström et al. 1988, Lake et al. 2007). In addition, large particulate organic P deposits to sediments during the high flow period could be remineralized during subsequent low flow conditions. As a result of redox mobilization and organic P mineralization with oxygen depletion, release of DRP through desorption of Fe/Al-P from sediments occurred. During the low flow period, most months had negative DRP retention rates, ranging from −85 % to −9 % (Table 2). We speculate that regeneration from organic P to DRP in the water column during the low flow period was another important process leading to an increase in DRP. Elevated DRP concentrations in the lacustrine zone during low flows (January and February) relative to the lower values in river and transition zone (Fig. 4a) further suggest hydrological controls on in-situ transformation in Xipi Reservoir.
The role of cascade reservoirs in regulating P export from river to coast
Results of P mass balances of the Xipi Reservoir could be scaled up to the whole cascade reservoir system. As shown by the annual P budget for the lacustrine zone (Table 2), Xipi Reservoir was a net sink for TPP, and approximately 6 % of the TPP input from the upper river was retained by the single reservoir (lacustrine zone) on an annual basis. If we assume that the other reservoirs had the same TPP retention rate (that is, 94 % of the initial TPP input can be passed on to the next reservoir), then only 69 % of the initial TPP was exported from the whole cascade reservoir system, while 31 % of the input was retained. However, because Xipi Reservoir was a net source for DRP (−0.3 %) due to in-situ transformation (e.g., deposition, sediment release or regeneration from organic matter), and DRP was usually the dominant form in TP, net retention of TP in the reservoir was offset and annual retention rates of TP were only 1 %. Scaling to the whole cascade reservoir system with the same method as TPP, only 6 % of the initial TP input from the upper river was retained.
The overall TP retention rate of this study (1 % for single reservoir and 6 % for the whole system) was less than previous estimates. For larger reservoirs, such as the Three Gorges Reservoir in the Yangtze River, over 70 % of the sediment load and more than half of the P load is intercepted by deposition in the reservoir (Huang et al. 2015), much higher than the results found in this study. A high fraction of P (60 % of inputs) was found to be retained by a subtropical reservoir over a 6 year period (Burford et al. 2012), while the small tropical reservoir of Cointzio in the southern Mexican Central Plateau with a water residence time less than one year reduced 30 % of the P load (Némery et al. 2016). As much as 79 % of the annual total P inputs to the Nui Coc reservoir (Vietnam) were deposited or eliminated (Le et al. 2014). Based on the comparison with the above studies, we speculated that the possible reason for the low TP retention in Xipi Reservoir is the high P loading and the relatively short residence time (average HRT less than one day) that does not allow for a complete deposition of TPP, due to the small reservoir size and shallow river channel. The sediment budget suggested that the erosional and depositional processes could be shifted for the same stretch of river as it is affected by a sequence of dams (Lu et al. 2015). As sediments are closely related to P transport and cycling, how cascade dams affect the P export for the whole river watershed deserves further detailed study.
Conclusion
In a subtropical area such as the Jiulong River watershed with a monsoon climate, the strong seasonal variation of precipitation determines the river discharge but is regulated by cascade dams. The hydrological condition influences P retention and down river export by changing the water residence time and associated in-stream processes. Changes in the role of cascade reservoirs on P retention and potential transformation processes with hydrological and meteorological condition are summarized in Fig. 7. Low flow conditions with low temperatures facilitated deposition of particulate P but restrained algal uptake, which resulted in a high retention of TPP in the dam reservoir. High flow delivers large amounts of sediment-bound P and retention is high. Medium flow and elevated temperatures favor algal uptake of DRP and addition of TPP as a result. Biotic uptake might control the retention of DRP in medium or high flow conditions with warm temperatures, while regeneration of organic P and release of sediment P are likely to account for the addition of DRP in the water column under low flow conditions. Therefore, P fluxes to down river and coast varied among P forms and with season. In summary, this river with cascade reservoirs appears to deliver large DRP in low flow periods but relatively more TPP in medium flow periods to the lower river. This finding presents an important implication for coastal ecology with respect to P availability and stoichiometry. In the context of increasing human and climate perturbation, dam-regulation of rivers is anticipated to increase the uncertainty associated with P export from river to coast in terms of fluxes and timing. This study highlights the hydrological controls on the role of cascade reservoirs in regulating P retention and downriver fluxes in different seasons. Future efforts should be made to link riverine P delivery, cycling and coastal ecosystem response.

A schematic showing hydrological controls on the role of cascade reservoirs in regulating phosphorus retention and downriver fluxes under an Asian monsoon climate
References
Bailey EM, Boynton WR, Ceballos MAC (2011) Monitoring of sediment oxgen and nutrient exchange measurements in Eastern Bay, Tangier sound and tributaries in support of TDML Development 2009. USA: Maryland Department of the Environment, UMCES report TS-610-11-CBL: 18–19
Boström B, Persson G, Broberg B (1988) Bioavailability of different phosphorus forms in freshwater systems. Hydrobiologia 170:133–155
Burford MA, Green SA, Cook AJ, Johnson SA, Kerr JG, O’Brien KR (2012) Sources and fate of nutrients in a subtropical reservoir. Aquat Sci 74:179–190
Chen NW, Wang LJ, Liu H, Wu JZ, Liu T (2012) A spatio-temporal correlation analysis of water quality and economic growth in the Jiulong River basin. Journal of Ecology and Rural Environment 28:19–25
Chen NW, Peng BR, Hong HS, Turyaheebwa N, Cui SH, Mo XJ (2013) Nutrient enrichment and N:P ratio decline in a coastal bay–river system in Southeast China: the need for a dual nutrient (N and P) management strategy. Ocean & Coastal Management 81:7–13
Chen NW, Wu YQ, Chen ZH, Hong HS (2015) Phosphorus export during storm events from a human perturbed watershed, Southeast China: implications for coastal ecology. Estuar Coast Shelf Sci 166:178–188
Coles JF, Jones RC (2000) Effect of temperature on photosynthesis-light response and growth of four phytoplankton species isolated from a tidal freshwater river. J Phycol 36:7–16
Conley DJ, Paerl HW (2009) Controlling eutrophication nitrogen and phosphorus. Science 323:1014–1015
Cook PLM, Aldridge KT, Lamontagne S, Brookes JD (2010) Retention of nitrogen, phosphorus and silicon in a large semi-arid riverine lake system. Biogeochemistry 99:49–63
Cowan JLW, Boynton WR (1996) Sediment-water oxygen and nutrient exchanges along the longitudinal axis of Chesapeake Bay: seasonal patterns, controlling factors and ecological significance. Estuaries 19:562–580
Duan SW, Kaushal SS, Groffman PM, Band LE, Belt KT (2012) Phosphorus export across an urban to rural gradient in the Chesapeake Bay watershed. J Geophys Res-Biogeosci 117:1–12
Essington TE, Carpenter SR (2000) Mini-review: nutrient cycling in lakes and streams: insights from a comparative analysis. Ecosystems 3:131–143
Evans DJ, Johnes PJ, Lawrence DS (2004) Physico-chemical controls on phosphorus cycling in two lowland streams. Part 2-the sediment phase. Sci Total Environ 329:165–182
Harris LA, Hodgkins CLS, Day MC, Austin D, Testa JM, Boynton W, Van Der Tak L, Chen NW (2015) Optimizing recovery of eutrophic estuaries: impact of destratification and re-aeration on nutrient and dissolved oxygen dynamics. Ecol Eng 75:470–483
Horowitz AJ, Elrick KA (1987) The relationship of stream sediment surface area, grain size and composition to trace element chemistry. Appl Geochem 2:437–451
Huang L, Fang H, Reible D (2015) Mathematical model for interactions and transport of phosphorus and sediment in the three gorges reservoir. Water Res 85:393–403
Humborg C, Ittekkot V, Cociasu A, Bodungen BV (1997) Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure. Nature:385–388
Jin GZ, S-i O, Amano A, Saito M, Shimizu Y, Satou T (2013) Effects of dam construction on sediment phosphorus variation in a semi-enclosed bay of the Seto Inland Sea, Japan. Estuar Coast Shelf Sci 135:191–200
Lake BA, Coolidge KM, Norton SA, Amirbahman A (2007) Factors contributing to the internal loading of phosphorus from anoxic sediments in six Maine, USA, lakes. Sci Total Environ 373:534–541
Le TPQ, Ho CT, Duong TT, Rochelle-Newall E, Dang DK, Hoang TS (2014) Nutrient budgets (N and P) for the Nui Coc reservoir catchment (North Vietnam). Agric Water Manag 142:152–161
Liu Q, Liu S, Zhao H, Deng L, Wang C, Zhao Q, Dong S (2015) The phosphorus speciations in the sediments up- and down-stream of cascade dams along the middle Lancang River. Chemosphere 120:653–659
Liu S, Guo Z, Li T, Huang H, Lin SJ (2011) Photosynthetic efficiency, cell volume, and elemental stoichiometric ratios in Thalassirosira weissflogii under phosphorus limitation. Chin J Oceanol Limnol 29:1048–1056
Lu T, Chen NW, Chen ZH, Wang LJ, Wu JZ (2013) Characteristics of sediment phosphorus in the Jiulong River-reservoir system and its ecological significance. Environmental Science 34:92–98
Lu XX, Li S, He M, Zhou Y, Bei R, Li L, Ziegler AD (2011) Seasonal changes of nutrient fluxes in the upper Changjiang basin: an example of the Longchuanjiang River, China. J Hydrol 405:344–351
Lu XX, Oeurng C, Le TPQ, Thuy DT (2015) Sediment budget as affected by construction of a sequence of dams in the lower Red River, Viet Nam. Geomorphology 248:125–133
Maavara T, Parsons CT, Ridenour C, Stojanovic S, Durr HH, Powley HR, Van Cappellen P (2015) Global phosphorus retention by river damming. P Natl Acad Sci USA 112:15603–15608
Malone TC (1992) Effects of water colum processes on dissolved oxygen, nutrients, phytoplankton, and zooplankton, in oxygen dynamics in the Chesapeake Bay: A Synthesis of recent research, edited by Smith DE, Leffler M, Mackiernan G. United States: Maryland Sea Grant College: 61–112
Miller WD, Harding LW, Adolf JE (2006) Hurricane Isabel generated an unusual fall bloom in Chesapeake Bay. Geophys Res Lett 33:1–4
Moss B, Kosten S, Meerhoff M (2011) Allied attack: climate change and eutrophication. Inland Waters 1:101–105
Némery J, Garnier J (2007) Origin and fate of phosphorus in the seine watershed (France): agricultural and hydrographic P budgets. J Geophys Res 112:1–14
Némery J, Gratiot N, Doan PTK, Duvert C, Alvarado-Villanueva R, Duwig C (2016) Carbon, nitrogen, phosphorus, and sediment sources and retention in a small eutrophic tropical reservoir. Aquat Sci 78:171–189
Oliver AA, Dahlgren RA, Deas ML (2014) The upside-down river: reservoirs, algal blooms, and tributaries affect temporal and spatial patterns in nitrogen and phosphorus in the Klamath River, USA. J Hydrol 519:164–176
Ouyang W, Hao F, Song K, Zhang X (2011) Cascade dam-induced hydrological disturbance and environmental impact in the upper stream of the Yellow River. Water Resour Manag 25:913–927
Parsons TR, Maita Y, Lalli CM (1984) Manual of chemical and biological methods for seawater analysis. Int Rev Hydrobiol 70:903–904
Redfield AC, Ketchum BH, Rechards FA (1963) The influence of organisms on the composition of seawater. The Sea., 2, New York, 26–77
Ruban V, Lopez-Sanchez JF, Pardo P, Rauret G, Muntau H, Quevauviller P (1999) Selection and evaluation of sequential extraction procedures for the determination of phosphorus forms in lake sediment. J Environ Monit 1:51–56
SEPA (2002) State Environm ental Protection Administration of the People's Republic of China. Water and waste water: monitoring and analytical methods. Bejing, P.R. China: Chinese Environment Science Press
Stamm C, Jarvie HP, Scott T (2014) What's more important for managing phosphorus: loads, concentrations or both? Environmental Science & Technology 48:23–24
Stanley EH, Doyle MW (2002) A geomorphic perspective on nutrient retention following dam removal. Bioscience 52:693–701
Sterner RW (2008) On the phosphorus limitation paradigm for lakes. Int Rev Hydrobiol 93:433–445
Syvitski JPM, Vörösmarty CJ, Kettner AJ, Green P (2005) Impact of humans on the flux of terrestrial sediment to the global coastal ocean. Science 308:376–380
Tysmans DJJ, Löhr AJ, Kroeze C, Ivens WPMF, van Wijnen J (2013) Spatial and temporal variability of nutrient retention in river basins: a global inventory. Ecol Indic 34:607–615
Van Cappellen P, Maavara T (2016) Rivers in the Anthropocene: global scale modifications of riverine nutrient fluxes by damming. Ecohydrology & Hydrobiology 16:106–111
Withers PJ, Jarvie HP (2008) Delivery and cycling of phosphorus in rivers: a review. Sci Total Environ 400:379–395
Yao QZ, Yu ZG, Chen HT, Liu PX, Mi TZ (2009) Phosphorus transport and speciation in the Changjiang (Yangtze River) system. Appl Geochem 24:2186–2194
Zieliński P, Jekatierynczuk-Rudczyk E (2015) Comparison of mineral and organic phosphorus forms in regulated and restored section of a small lowland river (NE Poland). Ecohydrology & Hydrobiology 15:125–135
Acknowledgments
This study was supported by the National Natural Science Foundation of China (No. 41376082), the Program for New Century Excellent Talents in University (NCET-13-0496), and the Fundamental Research Funds for the Central Universities (2012121053). We thank Jonathan Vause for his assistance with English editing, and Longjian Wang, Chaochao Yu and Guoxiang Chen for their help with sampling and lab analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Rights and permissions
About this article

Cite this article
Lu, T., Chen, N., Duan, S. et al. Hydrological controls on cascade reservoirs regulating phosphorus retention and downriver fluxes. Environ Sci Pollut Res 23, 24166–24177 (2016). https://doi.org/10.1007/s11356-016-7397-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-016-7397-3