
Abstract
Rapid urbanization and the absence of efficient water management policies are increasingly degrading the water quality of tropical reservoirs in developing countries. The small tropical reservoir of Cointzio, located in the Trans-Mexican Volcanic Belt, is a warm monomictic water body (surface area = 6 km2 with short water residence time <1 year) that is strategic to the drinking water supply of the city of Morelia and to downstream irrigation during the dry season (6 months of the year). The reservoir faces two threats: (a) reduced water storage capacity due to sediment accumulation and (b) eutrophication caused by excess nutrients that likely come from untreated wastewaters in the upstream watershed. Intensive field measurements of water and sediment were conducted in 2009 to characterize the trophic status of the reservoir and to estimate nitrogen (N) and phosphorus (P) sources, total suspended sediment (TSS) (N), (P), and carbon (C) loads, and their accumulation or removal in the reservoir. We found that point sources represent the majority of N and P inputs to the reservoir. The trophic status is clearly eutrophic given the high chlorophyll a peaks (up to 70 µg L−1) and a long period of anoxia (from May to October). Most of the TSS, C, N, and P were conveyed to the reservoir between June and October during the wet season. The TSS yield from the watershed was estimated at 35 ± 19 t km−2 year−1, of which more than 90 % was trapped in the reservoir (sediment accumulation rate = 7800 ± 2100 g m−2 of reservoir year−1). The export load of C, N, and P downstream at the reservoir outlet was reduced by 31, 46 and 30 % respectively in comparison to the load at the reservoir inlet. This study reveals the effect of climatic seasonality on inputs to tropical reservoirs and accumulation; it also highlights the need to both reduce nutrient input to combat eutrophication and mitigate erosion to maintain the water storage capacity of the reservoir over the long term.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The functioning of reservoirs is strongly influenced by human activities that occur upstream (Kennedy et al. 2003). Large amounts of sediments, organic matter, and nutrients are efficiently trapped in reservoirs, which can lead to eutrophication (Garnier et al. 1999; Donohue and Molinos 2009) and loss of storage capacity (Maneux et al. 2001; Vörösmarty et al. 2003; Rãdoane and Rãdoane 2005; Dang et al. 2010). On a global scale, reservoirs have been identified as the most significant sink of suspended sediment (TSS), carbon (C), nitrogen (N), and phosphorus (P) in inland waters, reducing nutrient fluxes from upland to downstream ecosystems and coastal regions (Cole et al. 2007; Friedl and Wüest 2002; Bosch and Allan 2008; Seitzinger et al. 2010). However, these studies showed large disparities among regions of the world and reservoir sizes. Although most studies have focused on large reservoirs, small reservoirs are also of great interest on a global scale (Syvitski et al. 2005). Based on the global database of inland waters developed by Lehner and Döll (2004), the total surface area of reservoirs worldwide was estimated at 2.5 × 105 km2 (Harrison et al. 2009). The total area of small reservoirs (<50 km2) accounted for 40 % of the total global reservoir area. The authors point out that small reservoirs can retain up to 45 % of the total N retained by all the world’s reservoirs Furthermore, the number of small reservoirs in tropical areas is increasingly important due to the numerous impoundment projects that are currently underway in developing countries (Tranvik et al. 2009). Tropical regions are characterized by contrasted wet and dry seasons, which have important consequences for the hydrological processes that occur in small reservoirs. These systems are highly vulnerable to eutrophication, especially during the extended dry and warm season when nutrients accumulate and algal blooms increase (Burford et al. 2012). Furthermore, the warmer temperatures that characterize tropical regions may lead to substantial greenhouse gas emissions (CO2 and CH4) through intense mineralization of the carbon present in reservoirs (Guérin et al. 2008; Chanudet et al. 2011). In spite of the importance of tropical reservoirs, literature about them remains limited (Kunz et al. 2011). Therefore a clear need exists to provide additional knowledge of small tropical reservoirs influenced by the strong seasonality of incoming water, TSS, and nutrient fluxes.
In tropical areas, water quality management has become an increasingly important issue in developing countries such as Mexico. The release of untreated wastewater into aquatic ecosystems is a common practice in many tropical countries. This practice is of great concern because the volume of wastewater produced is increasing due to rapid urbanization and economic growth (Le et al. 2015). According to various studies in Mexico, most Mexican lakes and reservoirs continue to deteriorate; the aquatic ecosystems exhibit ecological consequences such as eutrophication (Bravo-Inclán et al. 2011). Mexican authorities concede that water pollution is one of the most serious challenges for sustainable water resource management; water pollution also represents one of the most important concerns of local populations (Berrera Camacho and Bravo Espinosa 2009). The development of wastewater treatment infrastructure in Mexico remains insufficient, both in large cities and in smaller rural settlements (Ramírez-Zierold et al. 2010). Alcocer and Bernal-Brooks (2010) recently provided an overview of the status of lakes and reservoirs in Mexico, particularly within the Trans-Mexican Volcanic Belt (TMVB), where our study site is located. The authors highlighted the lack of data on Mexican rivers, indicating that knowledge about linkages between sediment and nutrient sources within upstream watersheds of reservoirs is limited in Mexico. Hence, the implementation of monitoring networks of discharge and nutrients, as well as the establishment of policy and mitigation strategies of point and non-point pollution sources are necessary to solve water quality problems in the country.
This study focuses on the small tropical reservoir of Cointzio (state of Michoacán), which is used to supply drinking water to the city of Morelia and for irrigation purposes. This reservoir is threatened by sediment accumulation and eutrophication that may increase water treatment costs and decrease the sustainability of regional water resources. On the regional scale, the water quality and quantity flowing out of the Cointzio reservoir also affects the downstream Cuitzeo endorheic laguna, which is of great hydrological and ecological interest because it is the second largest lake in Mexico (Alcocer and Bernal-Brooks 2010). The main objectives of this work were to (1) identify and quantify the N and P sources from the watershed to the reservoir, (2) determine the trophic status of the reservoir, (3) characterize the seasonality of TSS, C, N, and P inputs, and (4) assess the reservoir’s trapping efficiency and evaluate the annual accumulation rates of TSS, C, N, and P in the reservoir.
Materials and methods
Study area
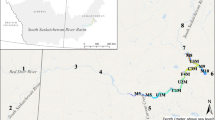
The Cointzio reservoir (19.622°N, −101.256°W) is located in the southern part of the Mexican Central Plateau on the TMVB at an altitude of 1990 masl (Fig. 1). The region has a sub-humid climate with mean annual rainfall of 810 mm, and air temperature fluctuates between 18 and 35 °C, according to data collected at the Morelia meteorological station (data from Servicio Nacional de Meteorología de México). Rainfall is concentrated during the wet season from June to October while the period between November and May is dry (period 1956–2001; Gratiot et al. 2010). The Cointzio reservoir was built in 1940. Its storage capacity is 66 × 106 m3 (most recent bathymetry in 2005; Susperregui 2008) with a maximal surface area of 6 km2 for a maximum depth of 29 m. The Cointzio reservoir is an essential component of the drinking water supply (20 % i.e. 21 × 106 m3, data from Organismo Operador de Agua Potable de Morelia) for the city of Morelia (700,000 inhabitants); it is also used for agricultural irrigation during the dry season.

Map of the Cointzio watershed and reservoir: location of sampling sites (geographical positions in UTM)
The Cointzio watershed drains an area of 630 km2 in which elevations range between 1990 and 3440 masl. The only perennial river is the Rio Grande de Morelia. At the reservoir outlet this river continues its course downstream to the Cuitzeo endorheic laguna (375 km2) (Allende et al. 2009). The majority of the Cointzio watershed is either forested (30 %) or cultivated (43 %) (López-Granados et al. 2013). The mean population density is 68 inh. km−2; population is 43,000 inhabitants (López-Granados et al. 2013). Soils are mostly volcanic (Andosols, Acrisols) and are highly degraded in parts of the watershed where intense erosion takes place during the wet season (Duvert et al. 2010).
Watershed survey
To determine the origin of nutrient pollution upstream of the reservoir, eight watershed sampling sites were selected according to land use, population density, and location in the river network (Fig. 1; Table 2). Sites 2 and 4 located downstream of the largest villages were typical of domestic point sources, whereas sites 3 and 5 were characteristic of diffuse sources in low population density areas and mixed forest/agriculture land uses. Site 1 was located downstream of a small village and pasture areas. Site 6 was representative of degraded agricultural lands affected by severe erosion. Site 7 was located along the main course of the Rio Grande de Morelia. During 2009, monthly water samples were collected just below the water surface in the middle reach of the river using 2-L polypropylene bottles. Water samples were kept refrigerated at 4 °C during transport to the laboratory where they were analyzed for nutrients and TSS. Discharge (salt dilution gauging method, Hudson and Fraser 2005) and dissolved oxygen (DO) were measured in the field using a multi-parameter Hydrolab MS5 probe.
Reservoir inflow and outflow
Given the low population density around the reservoir banks and the absence of other rivers, we considered the Rio Grande de Morelia River to be the predominant source of C, N, P, and TSS. Sampling site 8 was located at the outlet of the Cointzio watershed, downstream from the Santiago Undameo township (Fig. 1). The Undameo gauging station (Parshall flume to provide a control of the hydraulic section) was built in 1940 by the Comisión Nacional Del Agua (CONAGUA). A stage-discharge rating curve was also established and has been regularly adjusted. Since 2007 water levels have been measured at a 5-min time-step with a Thalimede OTT water level gauge; the CONAGUA rating curve has then been used to determine water discharge time series (Duvert et al. 2011). The Undameo gauging station physically separates the outlet of the Cointzio watershed from the inlet to the Cointzio reservoir. To calculate (C, N, P, TSS) annual loads, surface sampling was conducted using a bucket in the middle of the river (4 m width); samples were stored at 4 °C in a cooler during transport to the laboratory. The sampling frequency was daily for TSS measurements (since 2007) and weekly for C, N, and P measurements (in 2009).
At the reservoir outlet, CONAGUA measured discharges in the water uptake pipe immediately downstream of the reservoir on a daily basis, and additionally in the irrigation canal downstream of the spillway during the dry period. A meteorological station located atop the dam was used for measurement of evaporation and precipitation. For this study discharge, evaporation, and precipitation data for the period 2007–2009 were collected at CONAGUA. Daily TSS data were additionally obtained from CONAGUA for the period 2007–2008. In 2009 we sampled daily for TSS measurements and weekly for C, N, and P measurements. All water samples were collected by bucket from the water uptake pipe and the irrigation canal and stored at 4 °C in a cooler during transport to the laboratory.
Reservoir sampling
To assess spatial and temporal dynamics within the reservoir, vertical distributions of temperature (°C), DO (mg L−1), and turbidity (NTU) were measured using a multi-parameter Hydrolab MS5 probe. NTU measurements correlated well with TSS values (R2 = 0.97 data not shown). During 2009, surveys were conducted at 15 longitudinal stations along the reservoir (Fig. 1) at a fortnightly to monthly frequency. At each station, Secchi depth was measured using a Secchi disk. The deepest station P27 and the middle station P6 were also sampled at different depths on the vertical axis, (surface = 0.1, 1, 2, 5, 10, 15, 20 m and bottom = 0.3 m from the bottom) using a 2-L Niskin bottle. Each water sample was collected in a 2-L polypropylene recipient for TSS, C, N, and P measurements and chlorophyll a analysis. Samples were kept at 4 °C in a cooler during transport to the laboratory. Bottom sediment samples were collected with a Van Veen grab sampler on two occasions: during the dry season (19 May 2009) and at the end of the wet season (13 October 2009). Six stations along the longitudinal profile (P27, P13, P11, P6, P47, and P3) were chosen to assess the C, N, and P contents in deposited sediments. We assumed that sampling of the first 2 cm was representative of the sediment surface layer. Sediments were oven-dried at 60 °C for 24 h in the laboratory.
Water and sediment analyses
After sampling, water was filtered through GF/F Whatman glass fibre filters (0.7 µm porosity) in the laboratory within 3–6 h and frozen prior to dissolved nutrient analysis. Unfiltered water samples were also frozen for further total nutrient analysis. Total suspended solid (TSS) was weighed on GF/F Whatman glass fibre filters (dried 2 h at 105 °C) and expressed per unit volume filtered. Reproducibility for replicate was better than 3 %. Chlorophyll a was analyzed after filtration on GF/C Whatman glass fibre filters using the methanol extraction method of Holm-Hansen and Riemann (1978). Reproducibility for replicate was better than 3 %. Particulate organic carbon (POC) analyses were performed after filtering on GF/F Whatman glass fibre filters (ignited at 500 °C). Filtrates were kept for dissolved organic carbon (DOC) analysis. Filters were treated with HCl (2 N) to remove carbonates and dried at 60 °C for one night (Etcheber et al. 2007). POC was then determined on dry filters by combustion in a LECO CS 125 analyser (Etcheber et al. 2007). POC concentration (mg L−1) was then obtained by multiplying POC (mg g−1) and TSS (mg L−1). DOC was analyzed on filtered water using an OC-V Shimadzu analyzer (Sugimura and Suzuki 1988). Analytical accuracy and reproducibility of carbon analysis was better than 5 % (Coynel et al. 2005). P-tot and N-tot were analyzed on unfiltered water samples using a persulfate digestion process and standard colorimetric method (American Public Health Association; APHA 1995). Dissolved nutrients (PO4 3−, NH4 +, NO3 −) were analyzed on filtered water samples using standard colorimetric methods (APHA 1995). Reproducibility for replicate measurements was better than 5 % for all total and dissolved nutrient analysis.
The total particulate P (TPP) content of sediment was determined using a high temperature/HCl extraction technique (Aspila et al. 1976; Némery and Garnier 2007) prior to phosphate measurement by colorimetric method (Murphy and Riley 1962). To estimate particulate inorganic P (PIP), the analysis was similar to that of TPP, except that the high temperature organic P mineralization was omitted. Particulate organic P (POP) was determined by calculating the difference between TPP and PIP (Svendsen et al. 1993). Total carbon (TC) and total nitrogen (TN) contents were measured by CHN analysis using a CN-analyser FlashEA 1112 (Thermo Fisher Sci., MA, USA). Based on the catchment geology, which is primarily composed of volcanic and acid soils, inorganic C (IC) in sediment was expected to be low (Covaleda et al. 2011). To confirm this hypothesis, carbonates were analyzed using the calcimetric method (Robertson et al. 1999) and all values were <0.01 % C (CaCO3) (data not shown). Hence measured TC was thereafter considered to be organic carbon (OC). Reproducibility for replicate measurement was better than 5 % for OC, TN, TPP, and PIP.
Accumulation in the reservoir
In previous studies, the sediment deposition rate was estimated using three sediment cores sampled in 2006 at P27, P11, and P47 (Susperregui 2008). Stratigraphy and chronology using radioisotopic dating (210Pb) made it possible to reconstitute historical sediment deposition through 1974. Annual cycles of deposition were shown using RX intensity and particle sizing, and these results fitted well with the seasonality of inflow discharge at Undameo (Susperregui 2008). During the wet season (June to October), a large quantity of sediment is transported towards the bottom of the reservoir by hyperpycnal flows after flood events and it accumulates at depth. Comparison between cores showed homogeneous deposition within the reservoir (Susperregui 2008). The deposition surface area was estimated at 2 km2 based on a bathymetric map and particle size distribution of bottom sediments, which showed low variability at greater depths (D50 < 30 µm, Susperregui et al. 2009). The annual deposition rate was estimated at 2.5 ± 0.5 cm year−1 and the mean bulk density of sediment was 550 kg m−3, for an annual sedimentation rate of 14,000 ± 3000 g m−2 year−1 (Susperregui 2008). Sediment accumulation (Sedacc) was calculated as follows:
where Sedacc in t year−1, sedimentation rate = 14,000 ± 3000 g m−2 year−1, surface = 2 km2 and K is the unit conversion factor.
Mean contents of C, N, and P in deposited sediment during dry and wet periods were then assumed to be the same as settling particles. The significance of differences between the two seasons was tested using statistical U test (Mann–Whitney U test using XLSTAT software). Annual C accumulation (Cacc), N accumulation (Nacc) and P accumulation (Pacc) were calculated by multiplying Sedacc times mean annual OC, TN, and TPP content (Table 4). Accumulation uncertainties were evaluated based on the square root of the sum of the square of uncertainties of Sedacc and mean annual OC, TN, and TPP content.
To ensure comparability with other studies, the sediment, C, N, and P accumulation rates were normalized to the mean reservoir surface area for 2009 (3.6 km2) (Table 5).
Load calculations and uncertainties
Input and output TSS loads were calculated as the product of instantaneous TSS concentrations times the instantaneous discharge (for the output, we considered the daily discharge provided by CONAGUA to be the same as instantaneous discharge, on average). The TSS load (HL in t day−1) is calculated as follows:
where Qi is the instantaneous discharge (m3 s−1), Ci is the instantaneous TSS concentration (kg m−3).
The load (L in t) is the sum of daily loads during the time duration considered:
At the Undameo monitoring station, Duvert et al. (2010) assessed the uncertainty of the TSS load at ±20 % for the daily sampling strategy carried out in 2009. This uncertainty was assumed to be the same in 2007 and 2008 at both the Undameo station and the reservoir outlet.
The instantaneous P-tot, N-tot, and carbon loads resulted from the product of instantaneous concentrations in the water times the discharge measured at the same time. From the weekly sampling database, the P-tot, N-tot, and carbon loads (L) expressed in t year−1 were calculated according to the load-estimating procedure previously described by Verhoff et al. (1980) and recommended by Walling and Webb (1985):
where K is a conversion factor for the time duration and unit conversion, Ci is the instantaneous concentration in the water (g m−3), Qi is the instantaneous discharge (m3 s−1), and Qm is the mean annual discharge for 2009 (m3 s−1). The P-tot, N-tot, and carbon loads had 95 % confidence intervals (CI), calculated as described by Hope et al. (1997) and recommended when (Eq. 3) was used (Dawson et al. 2011; Némery et al. 2013). The total carbon (TOC) load was calculated as the sum of POC and DOC loads; the uncertainty of the TOC load was the square root of the sum of the square of uncertainties of POC and DOC loads described above.
Results
Hydrology
Patterns of riverine input into the reservoir were strongly influenced by the regional climatic regime. Two periods could be distinguished based on 2007–2009 observations. From November to May, the dry season, water input was quite low (minimum Q was between 0.18 and 0.31 m3 s−1) while reservoir output was at a maximum due to irrigation demand (maximum Q was 7–8 m3 s−1 Fig. 2a, b). From June to October, the wet season, riverine input was at its maximum (Q was 8–18 m3 s−1), which allowed the annual filling of the reservoir (Fig. 2c). All along the year, the reservoir provided water for drinking water supply (Q = 0.67 ± 0.09 m3 s−1). Water input from the watershed decreased from 49 × 106 m3 in 2007 to 42 × 106 m3 in 2009 mainly due to decreased precipitation (from 750 mm in 2007 to 690 mm in 2009). For the same period reservoir output decreased from 60 × 106 to 43 × 106 m3, indicating a progressive water deficit from year to year. The mean annual water volume in the reservoir decreased from 54 × 106 m3 in 2007 to 40 × 106 m3 in 2009. Between 2007 and 2009, evaporation (11–15.5 % of mean reservoir volume) was twice as high as precipitation (6 % of mean reservoir volume) (Table 1). Therefore the residence time of water in Cointzio was estimated at 1.03–0.81 year in 2007 and 0.90–0.83 year in 2009 (Table 1). Hydrological conditions during the period 2007–2009 (precipitation and Qmax between 750 and 690 mm year−1 and 8–18 m3 s−1 respectively, Table 1; Fig. 2a) were in the low range of the long-term time series (1956–2001, 650–1200 mm year−1, Qmax 10–40 m3 s−1) as shown in Gratiot et al. (2010). Years 2007, 2008, and 2009 were therefore typical of dry years. In 2009, the Cointzio reservoir had a lower water level (minimum of 21 m) than in 2007 and 2008; the 2009 water level minimum was also in the low range of the water level fluctuations from 1991 to 2005 (Fig. 2c).

Seasonal time-series of a water discharge (m3 s−1) and TSS inflow (mg L−1), b water discharge and TSS outflow (mg L−1), and c the volume of the Cointzio Reservoir for 2007–2009 (106 m3, incomplete year in 2007, mean, minimum and maximum volumes are given for the period 1991–2005; data CONAGUA)
Nutrient pollution levels within the watershed
Maximum P-tot, PO4 3−, and NH4 + concentrations were observed downstream of highly populated villages (Table 2). The highest mean P-PO4 3− (2.2 ± 1.6 mg L−1), N-NH4 + (5.9 ± 3.8 mg L−1), and N-NO3 − (2.6 ± 1.2 mg L−1) concentrations were observed at site 2 where we also measured the lowest DO (2.3 ± 1.2 mg L−1). Site 4 presented the highest mean P-tot concentration (4.4 ± 5.4 mg L−1) and also high concentrations of P-PO4 3− (0.37 ± 0.23 mg L−1) and N-NH4 + (1.23 ± 1.07 mg L−1). N-NO3 − concentrations remained quite low (<3 mg L−1) at all sites. Site 5 had the best water quality, with very low concentrations of P-PO4 3− (0.05 ± 0.03 mg L−1), N-NH4 + (0.06 ± 0.11 mg L−1), and N-NO3 − (0.78 ± 0.41 mg L−1). Sites 7 and 8 were quite similar in terms of nutrient concentrations. Mean concentrations at site 8 remained high, especially for P-tot (0.38 ± 0.23 mg L−1), P-PO4 3− (0.11 ± 0.07 mg L−1), and N-NH4 + (0.15 ± 0.10 mg L−1).
River input and reservoir output
TSS input was similar for the 3 years (≈22,200 ± 4 400 t year−1, Table 1). Maximum TSS concentration was observed during the wet season with peaks >10,000 mg L−1 (Fig. 2a, Supplementary material 1). Consequently TSS input showed a strong seasonality. TSS input during the wet season (June–October) represented more than 95 % of the annual TSS input for the 3 years (shown only for 2009, Fig. 3a). TSS output was found to be comparable for the 3 years and one order of magnitude smaller than TSS input (between 1800 and 2500 ± 400–500 t year−1, Table 1). Moreover, TSS output concentrations never exceeded 600 mg L−1 (Fig. 2b; Supplementary material 2). In 2009 mean input concentrations of P-tot (0.38 ± 0.23 mg L−1), N-tot (2.1 ± 1.0 mg L−1), and TOC (POC + DOC = 32.6 ± 26.5 mg L−1) were one and a half times higher than their output concentrations (Supplementary materials 1 and 2). The input concentrations of P-tot, N-tot, and TOC increased with increased discharge and TSS concentration during the wet season. For instance, maximum P-tot (0.94 mg L−1) and N-tot (5.9 mg L−1) were measured on the 24 June 2009 (Supplementary material 1) during the second largest TSS peak (Fig. 2a). TOC was at a maximum (173.5 mg L−1) during the extreme TSS peak on the 2 June 2009 (Fig. 2a, Supplementary material 1). Therefore P-tot, N-tot, and TOC inputs to the reservoir showed a strong seasonality concomitantly with flood events and TSS inputs (Fig. 3).

Seasonal variation of TSS, P-tot (including P-PO4 3−), N-tot (including N-NO3 −, N-NH4 +), POC, and DOC inputs to the Cointzio reservoir (t day−1) in 2009
P-tot input was 20 ± 4 t year−1 (P-PO4 3− accounted for 26 %, Table 3). P-tot input during the wet season (June to October) represented 84 % of the annual P-tot input and showed the same seasonal trend as TSS input (Fig. 3b). P-tot export downstream decreased by 30 % during transfer through the reservoir (P-tot output was 12 ± 3 t year−1 with the same proportion of P-PO4 3−, i.e. 25 %). N-tot input was 98 ± 17 t year−1 (61 % as N-NO3 − and 5.5 % as N-NH4 +) and export downstream of the reservoir was reduced by 46 % (output was 53 ± 15 t year−1, 73.6 % as N-NO3 − and 10 % as N-NH4 +) (Table 3). NO3 −export downstream of the reservoir was much lower (about 30 %) than the export of NH4 + (Table 3). TOC export downstream of the reservoir decreased by 31 % between the input (1617 ± 340 t year−1, 25 % as POC) and the output (1115 ± 167 t year−1, 5 % as POC). For TOC, the difference between the input and output is mainly due to POC reduction. As for P-tot, most of the N-tot (81 %) and TOC inputs (80 %) were concentrated during the wet season (Fig. 3c, d).
Overall, the reservoir received most of the TSS, TOC, N-tot, and P-tot inputs during the 5 months between June and October. During the rest of the year, inputs were very low. As a result of high discharges and high TSS concentrations, September was the most exceptional period in terms of input (about 50 % of TSS, 30 % of P-tot and N-tot, and 40 % of TOC annual inputs).
Internal reservoir dynamics
During 2009, the water level fluctuated 7 m between the maximum water level in January and the minimum in June (Fig. 4), as a result of irrigation output during the dry season (Fig. 2b). The 15 vertical temperature and DO profiles measured along the longitudinal axis did not reveal significant horizontal heterogeneities (data not shown). P27 was thereafter considered to be representative of the entire reservoir. The spatiotemporal dynamics of temperature, chlorophyll a, TSS, and DO at P27 are presented in Fig. 4. Temperature fluctuated between 14 and 22 °C. The reservoir behaved as a warm monomictic system with a progressive thermic stratification building up between April and October (Fig. 4a). This buildup was followed by complete vertical mixing of the water column in early November. During the stratified period DO decreased drastically from 4 to ~0 mg L−1 in the hypolimnion (Fig. 4b). The influence of hyperpycnal flows was observed between July and October when floods occurred in the watershed and filled the reservoir with turbid water (Fig. 4c). This input led to an increase of TSS from 50 to 250 mg L−1 in the hypolimnion layer of the reservoir (Fig. 4c). As a result of the input and increased TSS, the Secchi depth dropped from 31 cm in February to a minimum of 11 cm in September (R2 = 0.67 between TSS and Secchi depth, Fig. 5a).

Seasonal variation of a temperature, b TSS, c DO and d Chlorophyll a at the deepest station P27 in 2009

Comparison of seasonal variations in a Secchi depth, b chlorophyll a (surface and bottom), and c POC (surface and bottom) in the water column at the two sampling stations P6 and P27
Chlorophyll a was high in the first five meters of the water column during the dry season (January–June) (mean values of 30 ± 19 µg L−1 and maximum 70 µg L−1 observed in March, Fig. 4d). Concentrations then dropped drastically to 4 µg L−1 in the entire water column between July and December at the same time as the Secchi depth measurements decreased (Fig. 5a, b). Given the low correlation between chlorophyll a and TSS (R2 = 0.03) and between chlorophyll a and Secchi depth (R2 = 0.03) most particles were from TSS inputs rather than algae. The temporal variability of POC showed the same trend as chlorophyll a (R2 = 0.61, Fig. 5c). There was a good agreement in POC values between P6 and P27 (R2 = 0.63, Fig. 5c). POC followed the chlorophyll a dynamics with maximum values of 300–400 mg g−1 at the surface from January to June (mean 200 ± 100 mg g−1, Fig. 5c). With the arrival of the first floods in June, POC decreased simultaneously with the decrease in chlorophyll a, to a minimum value of 10 mg g−1 in December (mean 29 ± 18 mg g−1 between July and December).
NO3 −, NH4 + and PO4 3− concentrations at P6 and P27 were similar and showed comparable seasonal trends at P6 and P27 (R2 = 0.63, R2 = 0.83, R2 = 0.15, respectively, Fig. 6). Concentrations were slightly higher in the hypolimnion than in the epilimnion but remained within the same order of magnitude. PO4 3− concentration was low during the dry period (0.05 ± 0.05 mg L−1 on average from January to May) (Fig. 6a). NH4 + decreased rapidly from January to March and remained low until May (0.04 ± 0.02 mg L−1 at minimum in May) (Fig. 6c). The decrease in NH4 + and the low concentration of PO4 3− were concomitant to the maximum chlorophyll a that was observed during the dry period, Fig. 4d). Overall, PO4 3− and NH4 + increased more than tenfold beginning in May and remained high until destratification in late October when a sharp decrease in PO4 3− and NH4 + was observed (Fig. 6a, c). NO3 − increased slightly by a factor of two (2) throughout the year (Fig. 6b). Correlation between chlorophyll a and NH4 +and PO4 3− (R2 = 0.23 and 0.11 respectively) was higher than correlation between chlorophyll a and Secchi depth (R2 = 0.03) indicating that nutrients were the most important driver of algal production.

Comparison of seasonal variations in a P-PO4 3−, b N-NO3 −, and c N-NH4 + in the water column at the two sampling stations P6 and P27 (surface and bottom)
C, N, and P contents in sediments and accumulation
The OC, TN, and TPP contents of sediment showed minor variability along the longitudinal transect, except at P3, which was located close to mouth of the Rio Grande de Morelia River. At all other sampling sites, individual values were in the range of the standard deviation of the mean (Table 4). Sediments collected during the wet season had significantly lower mean content of TPP compared to sediments collected during the dry season (0.21 ± 0.03 compared to 0.12 ± 0.02 mg P g−1, p value <0.001). This difference was explained by a decrease in POP (0.17 ± 0.03 against 0.07 ± 0.01 mg P g−1) rather than PIP as the latter remained within the same order of magnitude (0.04 ± 0.01 versus 0.05 ± 0.01 mg P g−1). Dry season contents of OC (11.1 ± 1.4 mg C g−1) and TN (1.0 ± 0.1 mg N g−1) were similar to their concentrations in the wet season (10.2 ± 0.1 mg C g−1 and 0.09 ± 0.1 mg N g−1 respectively for OC and TN, p-value >0.05) (Table 4). The TN content in sediments remained low (~1 mg g−1). Resulting C:N ratio was 11 ± 1 on both dates.
Cacc was estimated at 298 ± 67 t C year−1, Nacc at 26 ± 6 t N year−1, and Pacc at 5 ± 1 g P year−1. C, N, and P accumulation rates are presented in Table 5 for comparison with accumulation rates in other tropical reservoirs.
Discussion
Nutrient sources within the watershed and reservoir trophic status
Point sources from domestic wastewater clearly dominated in the Cointzio watershed. Nutrient emission originated primarily from the most populated villages; the highest concentrations were observed around these villages (site 2 and 4, Table 2). This conclusion is in agreement with two previous studies on the Cointzio reservoir (López López and Dávalos-Lind 1998; Ramírez-Olvera et al. 2004). Both studies identified wastewater discharge as the major nutrient inputs into this reservoir. It is known that the villages located upstream have no wastewater treatment plants (Avila Garcia 2006).
Our results showed that N and P concentrations were low in agricultural sub-watersheds (sites 3 and 5, Table 2) compared with sub-watersheds affected by domestic wastewater. Nitrate concentrations remained low within the entire watershed (mean NO3 − of 1.5 ± 0.7 mg N-NO3 − L−1) give the European numerical standard of water quality (Water Framework Directive, 2000/60/EC 2000; Bouraoui and Grizzetti 2014). These results are in good agreement with the study that Bravo-Espinosa et al. (2009) conducted in the eastern part of the Cointzio watershed (site 6 Huertitas sub-basin, Table 2). These authors estimated very low nitrate loss in runoff waters from cultivated soils, in the range of 0.1–0.6 mg N-NO3 − L−1. Diffuse sources from agricultural soils are also generally linked to erosion processes that constitute a global concern (Quinton et al. 2010). A recent study using fingerprinting methods showed the prevailing contribution to sediment load at the Cointzio watershed outlet to be degraded soils (Evrard et al. 2013). Those soils (mainly Acrisols and Andosols) are very poor in C, N, and P content (Bravo-Espinosa et al. 2009), again indicating the low contribution from diffuse sources in this study. Other sources such as N and P atmospheric deposition can be considered very low given the high ratio between the watershed and the reservoir areas (watershed area/reservoir area >100); this suggests a stronger influence of watershed inputs than atmospheric inputs to the reservoir (Susperregui et al. 2009). A rough estimation of bulk atmospheric deposition (wet and dry) from a study in the nearby Valle de Bravo in central Mexico (16 kg P km−2 year−1 and 550 kg N km−2 year−1, Ramírez-Zierold et al. 2010) confirms that N and P depositions can be considered very low in the case of the Cointzio reservoir (0.06 t P year−1 and 2 t N year−1 i.e. 0.3 and 2 % of P-tot and N-tot inputs respectively, calculation was done using mean annual area of reservoir of 3.6 km2).
To support the assumption of the predominance of point sources, we calculated the theoretical domestic load based on the physiological per capita production of P (1–1.5 g P day−1, Billen et al. 2007). Given the total population of the watershed (43,000 inhabitants), direct point sources would represent an annual load of 15.5–23.5 t P. This is the same order of magnitude as the value at the outlet of the watershed (site 8), i.e. 20 ± 4 t P (Table 3). P domestic inputs from direct wastewater release would then explain the P-tot load reaching the reservoir.
To characterize the trophic status of the Cointzio reservoir, we applied the P trophic model for warm-water tropical lakes and reservoirs first proposed by Salas and Martino (1991). This approach makes it possible to estimate the trophic status of reservoirs from the annual P load and water residence time in the reservoir. Applying this method to the Cointzio reservoir led to an annual P load of 0.5 ± 0.1 g m−3 year−1 (ratio between input P-tot load of 20 ± 4 t P and mean annual reservoir volume of 40 × 106 m3 in 2009). Given the residence time of about 1 year (Table 1), the Cointzio reservoir was classified as eutrophic. According to the maximum chlorophyll a concentration (up to 70 µg L−1, Fig. 5b) the Cointzio reservoir appeared to be a highly productive system. The measurements of POC and chlorophyll a made it possible to calculate the C:chl a ratio for the reservoir (51 ± 19; R = 0.78). This ratio is the slope of the linear regression between POC concentrations and chlorophyll a (Garnier et al. 1989). Based on the algal growth rate (0.26 day−1) measured in the Cointzio reservoir by López López and Dávalos-Lind (1998), mean chlorophyll a (30 ± 19 µg L−1) converted to C from this C:chl a ratio and the 0–5 m layer (17 × 106 m3) from January to July, algal C uptake is estimated at 1400 ± 450 t C year−1. The annual primary production rate would then be 390 ± 130 g C m−2 year−1, which again categorizes the Cointzio reservoir as eutrophic, according to the lake trophic state classification (eutrophic >365 g C m−2 year−1; Wetzel 2001).
The Cointzio reservoir is eutrophic as are many neighboring lakes and reservoirs in the TMVB region: Lake Chapala (de Anda et al. 2011), the Lago de Guadalupe (Sepulveda-Jauregui et al. 2013), and numerous others (Bravo-Inclán et al. 2011). For example, a recent study on the main reservoir in the Valle de Bravo, which provides water to Mexico City, showed that N and P loads in the Valle de Bravo reservoir increased two to threefold between 1992 and 2005 (Ramírez-Zierold et al. 2010). The Valle de Bravo reservoir is eutrophic as a result of local township sewage and agriculture diffuse sources from the watershed. Lake Pátzcuaro, located near the Cointzio reservoir, has been facing severe eutrophication problems for at least 20 years (Chacón-Torres 1993; Rosas et al. 1993). Mijangos Carro et al. (2008) estimated the contribution of point and diffuse sources from 13 surrounding sub-watersheds (total surface of 933 km2). They concluded that urban nutrient loads were of high concern in towns adjacent to lakes whereas diffuse sources predominated in eroded and agricultural catchments. The oligotrophic Lake Zirahuen, located in the same region, has been spared from eutrophication so far but the study by Chacón-Torres and Rosas-Monge (2008) alerted the public about the urgency to collect and treat incoming wastewater to preserve this ecosystem.
According to the above-cited studies within the TMVB region and due to the obvious link between nutrient loads and eutrophication of reservoirs in tropical areas, implementation of best management practices (BMPs) in upstream watersheds is a crucial need. Traditional recommendations, as made by Ramírez-Zierold et al. (2010), are to reduce N and P inputs from wastewater (by treating sewage) and from agricultural diffuse sources (by regulating fertilizer use and inputs from livestock). In the case of the Cointzio watershed, the priority issue is a drastic reduction of nutrient point sources from domestic effluents (collection and treatment), which would benefit the water quality, as recently shown by the study of Doan et al. (2015). This mitigation would also have positive effects on the downstream Cuitzeo laguna, which has suffered from hypereutrophication and fish-fauna ecological impacts for many years, particularly when the laguna dried up during severe droughts in the early 1990s (Soto-Galera et al. 1999).
This regional context in Mexico can be generalized to many other developing countries in the inter-tropical area that are facing increasing population and lack of wastewater treatment. The implementation of BMPs listed above are widely recommended in numerous studies worldwide (Brazil, India, Sri Lanka, China and others; Reddy 2005).
Effect of flood events on reservoir functioning
In the Cointzio reservoir, a large proportion (>80 %) of the TSS, TOC, N-tot, and P-tot annual inputs occurred during the wet season (June–October). The seasonality of the inputs had a strong effect on the internal functioning of the reservoir. The most important feature was the very high TSS input during the wet season, which induced a rapid decrease of light penetration due to high turbidity. Turbidity also remained elevated during the dry season; the range of Secchi depths in the Cointzio reservoir (0.11–0.30 m) was among the lowest of the tropical lakes and reservoirs of the region. For comparison, the Secchi value is 13–19 m in the clear Lake Zirahuen (Martinez-Almeida and Tavera 2005) and 3.2–5.8 m in the eutrophic reservoir of Valle de Bravo (Merino-Ibarra et al. 2008). As a result of the decreased Secchi depth, a rapid drop of chlorophyll a was observed. In the comparable eutrophic reservoir of Valle de Bravo, chlorophyll a remained high during the entire stratified period (Merino-Iberra et al. 2008). Moreover, in eutrophic systems a decreased Secchi depth is generally correlated with increased chlorophyll a, which induces light absorption (Vollenweider 1968). This is not the case in the Cointzio reservoir, where turbidity was high during the entire year, especially after flood events, which maintained low light penetration affecting the seasonality of primary production.
Increased nutrient levels in the reservoir were primarily driven by large inputs of nutrients during flood events (Fig. 3). These inputs quickly and strongly enriched the reservoir to a high concentration level of PO4 3− and NH4 + in both the epilimnion and hypolimnion (Fig. 6). During stratification in tropical systems, nutrient depletion in the epilimnion due to algal uptake is generally observed, together with enrichment in the hypolimnion due to benthic mineralization and benthic desorption under anoxic conditions (Kunz et al. 2011; Merino-Iberra et al. 2008, Burford et al. 2012). Benthic desorption under anoxic conditions has been observed in most of the warm monomictic Mexican lakes and reservoirs listed by Alcocer and Bernal-Brooks (2010); in these water bodies, the hypolimnion becomes anoxic during the stratification period, which leads to PIP release from sediments and additional PO4 3− in the hypolimnion. In the Cointzio reservoir PO4 3− and NH4 + had already begun to increase prior to the arrival of floods that began in May and continuing over the course of stratification. The long and intense anoxia observed in the Cointzio reservoir from May to October (Fig. 4b) is consistent with intense benthic mineralization but little evidence of PIP desorption was observed because PIP in deposited sediment did not vary between dry and wet season. This behavior may be the consequence of the Acrisol-origin of sediments transported during flood events (Evrard et al. 2013). These soils typically have an elevated P adsorption capacity caused by the presence of iron and aluminium (Parfitt and Clayden 1991). In lakes and reservoirs with high iron and aluminium concentrations, hypolimnic anoxia does not necessarily lead to P release (Gächter and Müller 2003). In the Cointzio reservoir, aluminium and iron concentrations were elevated, regularly exceeding the standard for drinking water supply (unpublished data from Oaapas). In addition to benthic mineralization, floods would have a significant effect on PO4 3− concentration, whereas desorption of PIP from sediments appeared to be a negligible process. The concomitant effect of flood events and benthic mineralization on the increased hypolimnic concentrations was also highlighted in the Itezhi-Tezhi reservoir in Zambia (Kunz et al. 2011) but in the Zambian study nutrient concentrations remained stratified in the water column while in our study the increase was observed not only in the hypolimnion but also in the epilimnion. This could be the consequence of a wind effect that may have driven nutrient movement from the hypolimnion to the epilimnion as a result of boundary mixing caused by internal waves (Merino-Iberra et al. 2008). This effect is likely to occur in the Cointzio reservoir given the high diurnal wind variability (Susperregui 2008). The observed decrease in PO4 3− and NH4 + in October can be explained by the complete mixing of the water column and also by a dilution effect due to increased reservoir volume (Fig. 2c). Given the high interannual variability of precipitation and discharge in the Cointzio watershed, the effect of flood events is probably much more pronounced in wet years (Gratiot et al. 2010). This flooding effect is enhanced in tropical areas where seasonality is highly marked (Burford et al. 2012; Syvitski et al. 2014).
Sediment trapping efficiency and sediment accumulation
The Cointzio reservoir acted as a TSS sink. Sediment retention exceeded 90 % of incoming TSS. The difference between TSS input and output was estimated at about 20,000 t year−1 for the 3 years studied (Table 1). According to the classification proposed by Vörösmarty et al. (2003), the trapping efficiency of the Cointzio reservoir ranks in the highest range of anthropogenic sediment dam retention (80–100 %). Sediment retention based on differences between TSS inputs and outputs was significantly different from the Sedacc estimate (28,000 ± 6000 t year−1, Table 5). This difference can be explained by the particularly dry years of the study period (2007–2009) whereas sediment accumulation was estimated on the basis of an average value of long-term deposition obtained from sediment cores that spanned the years 1974–2006. TSS input was estimated at 22,000 t year−1 for the 3 years of our study (i.e. 35 ± 19 t km−2 year−1). The historical database of discharge and TSS at the watershed outlet (1973–1985, Oaapas, data not shown) enabled us to calculate a TSS input of up to 150,000 t year−1 i.e. 240 t km−2 year−1 for some wet years (1976 and 1981 for instance, Susperregui 2008). This indicates a considerable interannual variability of TSS input to the reservoir, which is driven by the hydrological conditions. This variability was also observed by Susperregui (2008) using sediment core data. Another factor that may be involved in TSS input variability is land use changes that occurred in the Cointzio watershed in recent decades. López-Granados et al. (2013) identified significant land use changes between 1986 and 1996 leading to increased reforested areas and shrubland progression. The sediment accumulation rate deduced from our study was 7 800 ± 2 100 g m−2 year−1 (Table 5). This value was of the same order of magnitude as that estimated on the global scale for small reservoirs (7700 g m−2 year−1) whereas large reservoirs presented sediment accumulation rates of about 10,000 g m−2 year−1 (Syvitski et al. 2005, Table 5). Compared to other tropical reservoirs listed in Table 5, the sediment accumulation rate was higher in the Cointzio reservoir than in the Itezhi-Tezhi reservoir (900 g m−2 year−1, Kunz et al. 2011, Table 5) or the Pampulha reservoir in Brazil (320 g m−2 year−1, Torres et al. 2007). Major differences in the sediment accumulation rate among reservoirs can be explained by the position of bottom gates and spillways and also by reservoir morphology (depth and surface area) and water residence time in the reservoir (Cunha et al. 2014). For instance, the combination of shallow water depth and high residence time can favor sediment accumulation. The lower sediment accumulation rate for the Pampulha reservoir can be explained by low water residence time (0.2 year) and a low specific TSS load (20 t km−2 year−1). In the case of Cointzio, even with a low residence time (<1 year) and substantial depth, the reservoir exhibited very efficient retention and a high sediment accumulation rate. These findings are likely a consequence of the high seasonality of the TSS input, which generated dense hyperpycnal flows and led to rapid sediment trapping.
Sediment accumulation is a key issue for management of the Cointzio reservoir because the reservoir has already lost 25 % (i.e. 22 × 106 m3) of its initial storage capacity after construction in 1940 (initial capacity was 88 × 106 m3, Susperregui 2008). The most promising way to mitigate a decrease in reservoir storage capacity would be to reduce TSS inputs by limiting upstream erosion. Alternative agronomic practices such as the planting of agave forest and native species are being implemented locally in the Cointzio watershed to prevent severe soil erosion (Schwilch et al. 2012); these practices may have long-term positive effects on TSS input to the reservoir. Modifying reservoir morphology or its hydraulic processes to reduce sediment accumulation rates appears to be unachievable.
C, N, and P trapping efficiency and accumulation
The Cointzio reservoir significantly reduced the export of TOC downstream of the dam (31 %) (Table 3). Cacc was estimated at 298 ± 67 t C year−1 (Table 5), which may explain about 60 % of this reduction, indicating potential additional C removal by mineralization. Our field data did not allow us to quantify C removal by mineralization. However, it is well known that aerobic and anaerobic mineralization in tropical reservoirs can generate CO2 and CH4 and emit them into the atmosphere (Guérin et al. 2006). The long and intense period of anoxia observed in the Cointzio reservoir clearly indicates high aerobic mineralization and also conditions favorable to methanogenesis. Additional loss of C via CO2 and CH4 emission would most likely occur in the Cointzio reservoir. The methanogenesis process, which has been widely demonstrated in tropical reservoirs, is of general concern for warming on the global scale (Barros et al. 2011). To assess these processes in the Cointzio reservoir, additional study based on a modeling approach is underway (Doan et al. 2014 pers. comm.).
Our estimation of the C accumulation rate was 83 ± 19 g C m−2 year−1 (Table 5), which was higher than the value estimated at the Itezhi-Tezhi reservoir, for instance (62 g C m−2 year−1, Kunz et al. 2011, Table 5). This difference can be explained by the higher sediment accumulation rate in the Cointzio reservoir and substantial allochthonous C inputs during the wet season. The opposite phenomenon was observed in the Pampulha reservoir in Brazil, which exported C rather than accumulating it (Torres et al. 2007, Table 5). The high productivity of this reservoir was identified as the main factor for explaining the C exportation. In the Cointzio reservoir the low mean C:N ratio in deposited sediment (11 ± 1, Table 4) indicated that deposited sediment may contain a significant proportion of autochthonous C (Park et al. 2009). However, the very low OC content in sediment emphasized the probable dominance of allochthonous C in the C accumulation, which should be confirmed by a complementary study on C fingerprinting. C accumulation can therefore vary widely among reservoirs and may be linked to the allochthonous C inputs and sediment trapping efficiency (Tranvik et al. 2009).
The difference between N-tot input and output indicated that N export downstream of the reservoir was reduced by 46 %. Sixty percent (60 %) of this reduction can be explained by Nacc, which was estimated at 26 ± 6 t N year−1 (Table 5). The N accumulation rate (7.2 ± 1.7 g N m−2 year−1) was slightly lower than the estimated N accumulation rate on the global scale (11 ± g N m−2 year−1, Beusen et al. 2005, Table 5) and as the N accumulation rate estimated in the Valle de Bravo reservoir (Table 5). The N accumulation rate can vary widely among reservoirs because of the difference in TSS trapping efficiency or water residence time, as highlighted previously for sediment accumulation (Cunha et al. 2014). Moreover, the estimation of total N removal in reservoirs is much more complex because N cycling is dynamic; it involves not only accumulation but also numerous biological processes (i.e. denitrification, N fixation, nitrification, Wetzel 2001). Denitrification processes are particularly responsible for N removal, with the production of N2, which is known to be an important process in explaining the mass balance of nitrogen in aquatic tropical ecosystems (Lewis 2002). In the Cointzio reservoir, the reduction of 30 % that was observed between NO3 − input and output suggests possible denitrification. In the eutrophic Valle de Bravo reservoir, for instance, N removal by denitrification was equivalent to N accumulation (Ramírez-Zierold et al. 2010). The long and intense anoxia in Cointzio was likely to favor denitrification. N removal by denitrification would then be of importance to achieve the N mass balance in the Cointzio reservoir. This assumption is consistent with the NiRReLa global model results of Harrison et al. (2009), who identified this central region of Mexico as having a high N removal rate potential. Further studies are needed to confirm our hypothesis. In addition, N fixation may also be an important process since N-fixing Cyanobacteria (Oscillatoria lacustris) were identified as the dominant group in the Cointzio reservoir during the dry season (Ramírez-Olvera et al. 2004).
The Cointzio reservoir acted as a P sink, with an estimated 30 % reduction of the P-tot export downstream of the reservoir. Pacc was estimated at 5 ± 1 t P year−1; this result explains a large proportion (>60 %) of the reduction (Table 5). However the difference in P inputs and outputs is not completely balanced by Pacc. This may be due to an underestimation of Pacc resulting from a possible underestimation of the PIP content in sediment by the HCl extraction method used. As mentioned above, the sediments of the reservoir of Cointzio may have a high P adsorption capacity. HCl would then not be strong enough to properly extract aluminium- and iron-bound P. Complementary analysis using Total HF-HClO4 extraction was conducted on four samples (data not presented), which resulted in systematically higher estimates of P content.
P trapping is commonly correlated with TSS retention and P is largely retained through sedimentation (Beusen et al. 2005). In the case of the Cointzio reservoir, although TSS retention was >90 %, the proportion of P retained remained moderate. P retention in reservoirs is also influenced by their water residence time (Kõiv et al. 2011). The longer the residence time, the higher the internal P recycling, which in turn leads to increased P retention. In the large eutrophic tropical reservoir of Castanhão in Brazil, which has a residence time of about 10 years, P retention was estimated at 98 % (Molisani et al. 2013). Therefore low water residence time in the Cointzio reservoir may explain the moderate P trapping efficiency. However, the water residence time is not the only factor that may explain P trapping efficiency. For instance, Torres et al. (2007) calculated P retention at 81 % in the Pampulha reservoir, which had a very low water residence time (0.2 year) and shallow waters. The latter feature can also favor the P trapping efficiency (Kõiv et al. 2011).
The P accumulation rate for the Cointzio reservoir (1.4 ± 0.3 g P m−2 year−1, Table 5) was slightly higher than the P accumulation rate at both the Pampulha reservoir (1.2 g P m−2 year−1) and the Itezhi-Tezhi reservoir (0.8 g P m−2 year−1). The latter has similar water residence time (0.7 year) and 60 % P trapping efficiency (Kunz et al. 2011, Table 5). The P accumulation rate was only 1/4 of the P accumulation rate in the neighboring Valle de Bravo reservoir (5.5 g P m−2 year−1), which has a water residence time of 1.58 y and retained 85 % of P input (Ramírez-Zierold et al. 2010, Table 5). The difference can be explained by the P input to the Valle Bravo reservoir (121 t P year−1), which was six times higher than P input to the Cointzio reservoir (20 t P year−1) although it has a similar upstream watershed area. Apart from the morphologically induced characteristics of the reservoirs discussed above, P retention also depends on watershed P inputs. Based on an analysis of 54 lakes and reservoirs in various climate regions around the world, Kõiv et al. (2011) found a strong correlation between the specific external P load and the reservoir P retention in P per m2. A comparison among several reservoirs showed that, regardless of the wide variety of reservoirs, the level of P input is the main cause of high P retention. This observation is consistent with the 6-year study in the Wivenhoe reservoir (Australia) conducted by Burford et al. (2012), who concluded that annual P retention variability was largely driven by the level of P input during each year.
Conclusions
In this study we assessed nutrient and sediment sources and their retention in a small tropical reservoir located in Central Mexico. High nutrient inputs from point sources to the Cointzio reservoir were the majot driver of its eutrophication, as shown by high chlorophyll a concentrations and the long and intense period of hypolimnic anoxia observed. Our results also indicate a pronounced seasonality in upstream inputs to the reservoir. Alternating dry and wet seasons strongly influenced the processes occurring within the reservoir. The reservoir was a trap for incoming sediment-bound nutrients and carbon capable of maintaining its eutrophic state over the long term.
This study has important implications for local management of the Cointzio reservoir, because it illustrates the strong influence of watershed-emitted untreated point sources on reservoir eutrophication. Also, even with low water residence times, efficient sediment trapping in the reservoir is likely to threaten the sustainability of its water storage capacity. As a result, the key challenge for local authorities is to reduce sediment and nutrient inputs to the reservoir by adopting best management practices in the watershed (i.e. domestic wastewater treatment, soil erosion mitigation strategies).
Our findings also have broader implications for all reservoir managers in tropical areas. Because eutrophication is an issue of concern for a large number of tropical lakes and reservoirs, point source reduction should be the highest water-management priority in tropical regions. This focus is particularly important because rapid population growth and increasing peri-urban settlement in these regions are likely to further exacerbate nutrient and sediment inputs to small reservoirs. In the context of increasing climate uncertainty, there is a critical need to implement effective mitigation strategies because climate change may also accelerate eutrophication risk in the coming decades. Overall, future research should focus on long-term, detailed biogeochemical monitoring of small tropical reservoirs, as well as their linkages with upstream watersheds. This research is a necessary step towards an improved understanding of the processes that occur in these systems and the driving forces behind them.
References
Alcocer J, Bernal-Brooks FW (2010) Limnology in Mexico. Hydrobiology 644:15–68
Allende TC, Mendoza ME, López Granados EM, Morales Manilla LM (2009) Hydrogeographical regionalisation: an approach for evaluating the effects of land cover change in watersheds. A case study in the Cuitzeo Lake watershed, Central Mexico. Water Res Manag 23:2587–2603
American Public Health Association (APHA) (1995) Standard methods for the examination of water and wastewater. Greenburg AE, Clesceri LS, Eaton AD (eds) 20th edn. American Public Health Association, Washington, DC
Aspila KI, Agemian H, Chau ASY (1976) A semi-automated method for the determination of inorganic, organic and total phosphate in sediments. Analyst 101:187–197
Avila Garcia P (2006) Water, society and environment in the history of one Mexican city. Environ Urban 8(1):129–140. doi:10.1177/0956247806063969
Barros N, Cole JJ, Tranvik LJ, Prairie YT, Bastviken D, Huszar VLM, del Giorgio P, Roland F (2011) Carbon emission from hydroelectric reservoirs linked to reservoir age and latitude. Nat Geosci 4:593–596
Berrera Camacho G, Bravo Espinosa M (2009) La planificación del territorio, gestión de recursos o gestión de conflictos. El Caso de Cointzio, Michoacán CONACYT report
Beusen AHW, Dekkers ALM, Bouwman AF, Ludwig W, Harrison J (2005) Estimation of global river transport of sediments and associated particulate C, N, and P. Glob Biogeochem Cycles 19:GB4S05. doi:10.1029/2005GB002453
Billen G, Garnier J, Némery J, Sebilo M, Sferratore A, Barles S, Benoit P, Benoît M (2007) A long-term view of nutrient transfers through the Seine river continuum. Sci Tot Environ 375:80–97
Bosch NS, Allan JD (2008) The influence of impoundments on nutrient budgets in two catchments of Southeastern Michigan. Biogeochemistry 87:325–338
Bouraoui F, Grizzetti B (2014) Modelling mitigation options to reduce diffuse nitrogen water pollution from agriculture. Sci Tot Environ 468–469:1267–1277
Bravo-Espinosa M, Mendoza ME, Medina-Orozco L, Prat C, García-Oliva F, López-Granados E (2009) Runoff, soil loss, and nutrient depletion under traditional and alternative cropping systems in the Transmexican Volcanic Belt, Central Mexico. Land Degrad Dev 20(6):640–653
Bravo-Inclán LA, Olvera-Viascán V, Sánchez-Chávez JJ, Saldaña-Fabela P, Tomasini-Ortiz AC (2011) Trophic state assessment in warm-water tropical lakes and reservoirs of the central region of Mexico. In: Van Bochove EPA, Vanrolleghem PA, Chambers G, Thériault B. Novotná and Burkart MR (eds) Issues and solutions to diffuse pollution: 14th international conference of the IWA Diffuse Pollution Specialist Group, DIPCON 2010 Québec, Canada, p 495
Burford MA, Green SA, Cook AJ, Johnson SA, Kerr JG, O’Brien KR (2012) Sources and fate of nutrients in a subtropical reservoir. Aquat Sci 74:179–190
Chacón-Torres A (1993) Lake Pátzcuaro, Mexico: watershed and water quality deterioration in a tropical high-altitude Latin American lake. Lake Reserv Manag 8:37–47
Chacón-Torres A, Rosas-Monge C (2008) Water quality characteristics of a high altitude oligotrophic Mexican lake. Aquat Ecosy Health Manag 1:237–243
Chanudet V, Descloux S, Harby A, Sundt H, Brakstad BHHO, Serça D, Guérin F (2011) Gross CO2 and CH4 emissions from the Nam Ngum and Nam Leuk sub-tropical reservoirs in Lao PDR. Sci Total Environ 409:5382–5391
Cole JJ, Prairie YT, Caraco NF, McDowell WH, Tranvik LJ, Striegl RG, Duarte CM, Kortelainen P, Downing JA, Middelburg JJ (2007) Plumbing the global carbon cycle: integrating inland waters into the terrestrial carbon budget. Ecosystems 10:172–185
Covaleda S, Gallardo JF, Garcıá-Oliva F, Kirchmann H, Prat C, Bravo M, Etchevers JD (2011) Land-use effects on the distribution of soil organic carbon within particle-size fractions of volcanic soils in the Transmexican Volcanic Belt (Mexico). Soil Use Manag 27:186–194
Coynel A, Etcheber H, Abril G, Maneux E, Dumas J, Hurtrez JE (2005) Contribution of small mountainous rivers to particulate organic carbon input in the Bay of Biscay. Biogeochem 74:151–171
Cunha DGF, de Carmo Calijuri MC, Dodds WK (2014) Trends in nutrient and sediment retention in Great Plains reservoirs (USA). Environ Monit Assess 186:1143–1155
Dang TH, Coynel A, Orange D, Blanc G, Etcheber H, Le LA (2010) Long term monitoring (1960–2008) of the river-sediment transport in the Red River Watershed (Vietnam): temporal variability and dam-reservoir impact. Sci Total Environ 408:4654–4664
Dawson JCC, Tetzlaff D, Speed M, Hrachowitz M, Soulsby C (2011) Seasonal controls on DOC dynamics in nested upland catchments in NE Scotland. Hydrol Proc 25:164–1658
de Anda J, Shear H, Maniak U, Riedel G (2011) Phosphates in Lake Chapala, Mexico. Lakes Reserv: Res Manag 6:313–321
Doan TKP, Némery J, Schmid M, Gratiot N (2015) Eutrophication of turbid tropical reservoirs: scenarios of evolution of the reservoir of Cointzio. Ecol Inform, Mexico. doi:10.1016/j.ecoinf.2015.01.006
Donohue I, Molinos JG (2009) Impacts of increased sediment loads on the ecology of lakes. Biol Rev 84:517–531
Duvert C, Gratiot N, Evrard O, Navratil O, Némery J, Prat C, Esteves M (2010) Drivers of erosion and suspended sediment transport in three headwater catchments. Geomorphology 123(3–4):243–256
Duvert C, Gratiot N, Némery J, Burgos A, Navratil O (2011) Sub-daily variability of suspended sediment fluxes in small mountainous catchments—implications for community-based river monitoring. Hydrol Earth Syst Sci 15:703–713
Etcheber H, Taillez A, Abril G, Garnier J, Servais P, Moatar F, Commarieu MV (2007) Particulate organic carbon in the estuarine turbidity maxima of the Gironde, Loire and Seine estuaries: origin and lability. Hydrobiology 588:245–259
Evrard O, Poulenard J, Némery J, Ayrault S, Gratiot N, Duvert C, Prat C, Lefèvre I, Bonté P, Esteves M (2013) Tracing sediment sources in a tropical highland catchment of central Mexico by using conventional and alternative fingerprinting methods. Hydrol Proc 27:911–922
Friedl G, Wüest A (2002) Disrupting biogeochemical cycles—consequences of damming. Aquat Sc 64:55–65
Gächter R, Müller B (2003) Why the phosphorus retention of lakes does not necessarily depend on the oxygen supply to their sediment surface. Limnol Ocean 48(2):929–933
Garnier J, Blanc P, Benest D (1989) Estimating a Carbon/chlorophyll ratio in nannoplankton (Creteil Lake, S-E Paris, France). Water Resour Bull 25:751–754
Garnier J, Leporcq B, Sanchez N, Philippon X (1999) Biogeochemical mass-balances (C, N, P, Si) in three large reservoirs of the Seine Basin (France). Biogeochemistry 47:119–146
Gratiot N, Duvert C, Collet L, Vinson D, Némery J, Saenz-Romero C (2010) Increase in surface runoff in the central mountains of Mexico: lessons from the past and predictive scenario for the next century. Hydrol Earth Syst Sci 14(2):291–300
Guérin F, Abril G, Richard S, Burban B, Reynouard C, Seyler P, Delmas R (2006) Methane and carbon dioxide emissions from tropical reservoirs: significance of downstream rivers. Geophys Res Lett 33:L21407. doi:10.1029/2006GL027929
Guérin F, Abril G, de Junet A, Bonnet MP (2008) Anaerobic decomposition of tropical soils and plant material: implication for the CO2 and CH4 budget of the Petit Saut Reservoir. Appl Geochem 23:2272–2283
Harrison JA, Maranger RJ, Alexander RB, Giblin AE, Jacinthe PA, Mayorga E, Seitzinger SP, Sobota DJ, Wollheim WM (2009) The regional and global significance of nitrogen removal in lakes and reservoirs. Biogeochemistry 93:143–157
Holm-Hansen O, Riemann B (1978) Chlorophyll a determination: improvements in methodology. Oikos 30:438–447
Hope D, Billett MF, Cresser MS (1997) Exports of organic carbon in two river systems in NE Scotland. J Hydrol 193:61–82
Hudson R, Fraser J (2005) Introduction to salt dilution gauging for streamflow measurement part IV: the mass balance (or dry injection) method. Watershed Manag Bull 9(1):6–12
Kennedy RH, Tundisi JG, Straškrábová V, Lind OT, Hejzlar J (2003) Reservoirs and the limnologist’s growing role in sustainable water resource management. Hydrobiology 504:xi–xii
Kõiv T, Nõges T, Laas A (2011) Phosphorus retention as a function of external loading, hydraulic turnover time, area and relative depth in 54 lakes and reservoirs. Hydrobiology 660:105–115
Kunz MJ, Wüest A, Wehrli B, Landert J, Senn DB (2011) Impact of a large tropical reservoir on riverine transport of sediment, carbon, and nutrients to downstream wetlands. Water Resour Res 47:W12531. doi:10.1029/2011WR010996
Le TPQ, Billen G, Garnier J, Chau VM (2015) Long-term biogeochemical functioning of the Red River (Vietnam): past and present situations. Reg Environ Change 15(2):329–339. doi:10.1007/s10113-014-0646-4
Lehner B, Döll P (2004) Development and validation of a global database of lakes, reservoirs and wetlands. J Hydrol 296:1–22
Lewis WM (2002) Estimation of background nitrogen yields for North America by use of benchmark watersheds. Biogeochemistry 57–58:375–385
López López E, Dávalos-Lind L (1998) Algal growth potential and nutrient limitation in a tropical river-reservoir system of the Central Plateau, Mexico. Aquat Ecosyst Health Manag 1:345–351
López-Granados E, Mendoza ME, González DI (2013) Linking geomorphologic knowledge, RS and GIS techniques for analyzing land cover and land use change: a multitemporal study in the Cointzio watershed, Mexico. Ambients Água Interdiscip J Appl Sci 8(1):18–37. doi:10.4136/ambi-agua.956
Maneux E, Probst JL, Veyssy E, Etcheber H (2001) Assessment of dam trapping efficiency from water residence time: application to fluvial sediment transport in the Adour, Dordogne and Garonne river basins (France). Water Resour Res 37:801–811
Martinez-Almeida V, Tavera R (2005) A hydrobiological study to interpret the presence of desmids in Lake Zirahuén, México. Limnologica 35:61–69
Merino-Iberra M, Monroy-Ríos E, Vilaclara G, Castillo FS, Gallegos ME, Ramírez-Zierold J (2008) Physical and chemical limnology of a wind-swept tropical highland reservoir. Aquat Ecol 42:335–345
Mijangos Carro M, Izurieta Dávila J, Gómez Balandra A, López RH, Huerto Delgadillo R, Sánchez Chávez J, Bravo Inclán L (2008) Importance of diffuse pollution control in the Patzcuaro Lake Basin in Mexico. Water Technol 58(11):2179–2186. doi:10.2166/wst.2008.820
Molisani MM, Becker H, Barroso H, Hijo CAG, Monte TM, Vasconcellos GH, Lacerda LD (2013) The influence of Castanhão reservoir on nutrient and suspended matter transport during rainy season in the ephemeral Jaguaribe river (CE, Brazil). Braz J Biol 73(1):115–123
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 27:31–36
Némery J, Garnier J (2007) Origin and fate of phosphorus in the Seine watershed (France): the agricultural and hydrographic P budget. J Geophys Res Biogeosci 112:G03012. doi:10.1029/2006JG000331
Némery J, Mano V, Coynel A, Etcheber H, Moatar F, Meybeck M, Belleudy P, Poirel A (2013) Carbon and suspended sediment transport in an impounded alpine river (Isère, France). Hydrol Proc 27:2498–2508
Parfitt RL, Clayden B (1991) Andisols—the development of a new order in soil taxonomy. Geoderma 49:181–198
Park HK, Byeon MS, Shin YN, Jung DI (2009) Sources and spatial and temporal characteristics of organic carbon in two large reservoirs with contrasting hydrologic characteristics. Water Resour Res 45:W11418. doi:10.1029/2009WR008043
Quinton JN, Govers G, Van Oost K, Bardgett R (2010) The impact of agricultural soil erosion on biogeochemical cycling. Nat Geosci 3:311–314
Rãdoane M, Rãdoane N (2005) Dams, sediment sources and reservoir silting in Romania. Sci Total Environ 71(1–2):112–125
Ramírez-Olvera MA, Díaz-Argüero M, López-López E (2004) Planktonic crustacean assemblages in a system of three reservoirs in the Mexican Central Plateau: seasonal and spatial patterns. J Freshw Ecol 19(1):25–34
Ramírez-Zierold JA, Merino-Ibarra M, Monroy-Ríos E, Olson M, Castillo FS, Gallegos ME, Vilaclara G (2010) Changing water, phosphorus and nitrogen budgets for Valle de Bravo reservoir, water supply for Mexico City Metropolitan Area. Lake Reserv Manag 26:23–34
Reddy MV (2005) Restoration and Management of Tropical Eutrophic Lakes. Science publisher Inc., Enfield, Plymouth, p 543
Robertson GP, Coleman DC, Bledsoe CS, Sollins P (1999) Standard soil methods for long-term ecological research. Oxford University Press, New York, Oxford, p 480
Rosas I, Velasco A, Belmont R, Baez A, Martínez A (1993) The algal community as an indicator of trophic status in Lake Patzcuaro, Mexico. Environ Poll 80:255–264
Salas HJ, Martino P (1991) A simplified phosphorus trophic state model for warm-water tropical lakes. Water Res 25(3):341–350
Schwilch G, Hessel R, Verzandvoort S (eds) (2012) DESIRE for greener land. Options for sustainable land management in drylands. Publishers University of Bern, CDE, Alterra, Wageningen UR and ISRIC, World Soil Information WOCATFAO, p 250
Seitzinger SP, Mayorga E, Kroeze C, Bouwman AF, Beusen AHW, Billen G, Van Drecht G, Dumont E, Fekete BM, Garnier J, Harrison JA (2010) Global river nutrient export: a scenario analysis of past and future trend. Glob Biogeochem Cycles 24:1–16
Sepulveda-Jauregui A, Hoyos-Santillan J, Gutierrez-Mendieta FJ, Torres-Alvarado R, Dendooven L, Thalasso F (2013) The impact of anthropogenic pollution on limnological characteristics of a subtropical highland reservoir “Lago de Guadalupe”, Mexico. Knowl Manag Aquat Ecosyst 410(4):1–15. doi:10.1051/kmae/2013059
Soto-Galera E, Paulo-Maya J, López-López E, Serna-Hernández JA, Lyons J (1999) Change in Fish Fauna as Indication of Aquatic Ecosystem Condition in Rio Grande de Morelia-Lago de Cuitzeo Basin, Mexico. Environ Manag 24(1):133–140
Sugimura Y, Suzuki Y (1988) A high temperature catalytic oxidation method for non-volatile dissolved organic carbon in seawater by direct injection of a liquid sample. Mar Chem 24:105–131
Susperregui AS (2008) Caractérisation hydro-sédimentaire des retenues de Cointzio et d’Umécuaro (Michoacán, Mexique) comme indicateur du fonctionnement érosif du bassin versant. Hydro-sedimentary characterization of the reservoirs of Cointzio and Umécuaro (Michoacán, Mexico) as an indicator of erosional activity in the watershed. Ph.D. Dissertation Grenoble University, p 289
Susperregui AS, Gratiot N, Esteves M, Prat C (2009) A preliminary hydrosedimentary view of a highly turbid tropical, manmade lake: Cointzio Reservoir (Michoacán, Mexico). Lakes Reserv: Res Manag 14:31–39
Svendsen LM, Redsdorf A, Nørnberg P (1993) Comparison of methods for analysis of organic and inorganic phosphorus in river sediment. Water Res 27:77–83
Syvitski JPM, Vörösmarty JV, Kettner AJ, Green P (2005) Impacts of humans on the load of terrestrial sediments to the global coastal ocean. Science 308:376–380
Syvitski JPM, Cohen S, Kettner AJ, Brakenridge R (2014) How important and different are tropical rivers? An overview. Geomorphology 227:5–17. doi:10.1016/j.geomorph.2014.02.029
Torres IC, Resck RP, Pinto-Coelho RM (2007) Mass balance estimation of nitrogen, carbon, phosphorus and total suspended solids in the urban eutrophic, Pampulha reservoir, Brazil. Acta Limnol Braz 19:79–91
Tranvik LJ, Downing JA, Cotner JB, Loiselle SA, Striegl RG, Ballatore TJ, Dillon P, Finlay K, Fortino K, Knoll LB, Kortelainen PL, Kutser T, Larsen S, Laurion I, Leech DM, McCallister SL, McKnight DM, Melack JM, Overholt E, Porter JA, Prairie Y, Renwick WH, Roland F, Sherman BS, Schindler DW, Sobek S, Tremblay A, Vanni MJ, Verschoor AM, von Wachenfeldt E, Weyhenmeyera GA (2009) Lakes and reservoirs as regulators of carbon cycling and climate. Limnol Oceanogr 54(6):2298–2314
Verhoff FH, Yaksich SM, Melfi DA (1980) River nutrient and chemical transport estimates. J Environ Eng Div 10:591–608
Vollenweider RA (1968) Scientific fundamentals of the eutrophication of lakes and flowing waters, with particular reference to nitrogen and phosphorus as factors of eutrophication. Technical report, DA 5/SCI/68.27, OCDE, Paris, p 250
Vörösmarty JV, Meybeck M, Fekete B, Sharma K, Green P, Syvitski JPM (2003) Anthropogenic sediment retention: major global impact from registered river impoundments. Glob Planet Change 39(1–2):169–190
Walling DE, Webb W (1985) Estimating the discharge of contaminants to coastal waters by rivers: some cautionary comments. Mar Poll Bull 16(12):488–492
Water Framework Directive, 2000/60/EC (2000) Establishing a framework for community action in the field of water policy (OJ (2000) L327/1)
Wetzel RG (2001) Limnology: lake and river ecosystems, 3rd edn. Academic, San Diego, p 1006
Acknowledgments
This work was undertaken in the framework of the European Research project DESIRE (2007–2011) and the French ANR Research project STREAMS (2008–2010). We thank the two PI Christian Prat and Michel Esteves for financial support. We thank the CIGA and CIECO, and UNAM campus of Morelia, for providing access to their laboratory facilities.
We are grateful to Jorge Schondube (UNAM) for kindly providing his wonderful electric boat and CONAGUA for their support in the project. We thank Alexandra Coynel and Henry Etcheber for help with POC analysis at the EPOC laboratory. We also thank students who helped us during field measurements from 2007 to 2009 (Florence Mahé, Natacha Salles, Antonio Munoz-Gaytan, Lila Collet and Anne-Sophie Susperregui). We are grateful to Marie-Paul Bonnet and Martin Schmid for fruitful discussions and comments. We also thank the three anonymous reviewers for their constructive comments.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Némery, J., Gratiot, N., Doan, P.T.K. et al. Carbon, nitrogen, phosphorus, and sediment sources and retention in a small eutrophic tropical reservoir. Aquat Sci 78, 171–189 (2016). https://doi.org/10.1007/s00027-015-0416-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00027-015-0416-5