
Abstract
Algal bloom could drastically influence the nutrient cycling in lakes. To understand how the internal nutrient release responds to algal bloom decay, water and sediment columns were sampled at 22 sites from four distinct regions of China’s eutrophic Lake Taihu and incubated in the laboratory to examine the influence of massive algal bloom decay on nutrient release from sediment. The column experiment involved three treatments: (1) water and sediment (WS); (2) water and algal bloom (WA); and (3) water, sediment, and algal bloom (WSA). Concentrations of dissolved oxygen (DO), total nitrogen (TN), total phosphorus (TP), ammonium (NH +4 -N), and orthophosphate (PO 3−4 -P) were recorded during incubation. The decay of algal material caused a more rapid decrease in DO than in the algae-free controls and led to significant increases in NH +4 -N and PO 3−4 -P in the water. The presence of algae during the incubation had a regionally variable effect on sediment nutrient profiles. In the absence of decaying algae (treatment WS), sediment nutrient concentrations decreased during the incubation. In the presence of blooms (WSA), sediments from the river mouth released P to the overlying water, while sediments from other regions absorbed surplus P from the water. This experiment showed that large-scale algal decay will dramatically affect nutrient cycling at the sediment–water interface and would potentially transfer the function of sediment as “container” or “supplier” in Taihu, although oxygen exchange with atmosphere in lake water was stronger than in columns. The magnitude of the effect depends on the physical–chemical character of the sediments.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
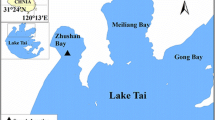
Eutrophication is a worldwide problem in freshwater ecosystems, where it often manifests as harmful algal blooms (HABs). Lakes in most parts of the world, including Asia, North and South America, Europe, and Africa, are increasingly affected by HABs, with implications for drinking water supplies, fisheries, and recreational use (Likens 1971; Reynolds 1987; Paerl 1988; Qin et al. 2010). Lake Taihu (meaning “large water” and referred to as “Taihu” in this paper) is the third largest freshwater lake in China, covering 2,338 km2 and having a volume of 4.4 billion m3 (Pu and Yan 1998; Qin et al. 2007). Taihu is located in the Yangtze River delta, the most rapidly developing region in China (Fig. 1), with approximately 40 million people living within its watershed. Over the past three decades, excessive nutrient inputs have led to the appearance and persistence of massive blooms of toxin-producing cyanobacteria, which have seriously degraded water quality and impacted adversely on human use of the lake (Guo 2007; Qin et al. 2007).

Sites for sampling in Lake Taihu
The development and subsequent decay of massive algal blooms in highly polluted regions of Taihu can cause “black spot” events in which dissolved oxygen (DO) is depleted throughout the water column (Liu 2009) and very high COD and ammonium concentrations can result (Chen 2011; Lu and Ma 2010). This phenomenon can last for more than 1 week and has happened nearly every year in the lake, and the coverage area could reach several square kilometers. In the northwestern part of Taihu, buoyant blooms accumulate throughout much of the year as a result of high nutrient inputs and prevailing winds (Fan et al. 1998; Cheng 2010). This is also the area of the lake where black spot events are most frequent. Black spot events caused the highly publicized drinking water crisis in the city of Wuxi in 2007, which directly affected the drinking water supply of some two million inhabitants (Guo 2007; Kong et al. 2007). In recent years, black spot events have occurred more widely in Taihu according to our field observation, even in some regions bloom happened not so frequently, like the northeastern bay of the lake, because once the bloom flowed into the bay or along the shoreline, and degraded undisturbed for a long time, the black spot event possibly came out.
Excessive anthropogenic nutrient loading, especially nitrogen (N) and phosphorus (P), promotes algal blooms (Reynolds and Walsby 1975; Smith 1983; Xu et al. 2010; Abell et al. 2010). Controlling nutrient input is the most effective way of reducing the risk of blooms (Brookes and Carey 2011), and significant efforts have made to reduce the external nutrient loads in Taihu (Qin et al. 2006; Qin et al. 2010), but the blooms have as yet shown no sign of decline. Sediments are an important source of nutrients in shallow lakes (Sas 1989; Marsden 1989; Jeppesen et al. 1991; Søndergaard et al. 1999, 2003), and blooms have been shown to stimulate the release of nutrients from the sediment (Xie et al. 2003). Taihu is very shallow (~1.9 m average depth) and sediment–water interactions have been shown to play an important role in its nutrient cycling dynamics (Qin et al. 2006; Zhu 2008). Therefore, an investigation into how the formation and decay of blooms affects nutrient cycling at the sediment–water interface is a useful approach to understand and control the eutrophication dynamics of Taihu.
The locations and movements of blooms in Taihu are highly dependent on wind direction and circulation, and black spot events tend to occur where these physical factors force blooms to accumulate, making it difficult to understand the process of nutrient release in such a big lake through field observation. Instead, an experimental approach was taken to determine the potential influence of the bloom on sediment nutrient release. Previous experiments with sediment from three sites in bloom-impacted Lake Taihu found that decaying blooms could “pump” nutrients from sediments. The extent to which this occurred was seen to vary from place to place, depending on the physical–chemical properties of sediment (Zhu et al. 2011). However, with only three sites, the results could not be reliably extrapolated to the lake as a whole or used to determine the potential for black spot events in other areas. The primary objective of this study was to understand the potential influence of algal bloom degradation on nutrient release from sediment across the whole lake. By sampling over a range of 22 sites in Taihu, we aimed to improve spatial resolution sufficiently to relate sediment nutrient release data to black spot events. In order to simulate black spot events and estimate the potential for nutrient release, we introduced algal scum to the water surface in sediment–water columns obtained from different regions of the lake and incubated them in dark to enhance algal degradation. We recorded the changes in the physical–chemical parameters of water and sediment during the subsequent incubation to describe the influence of algal bloom degradation on sediment nutrient release.
Materials and methods
Site description
Sediments are distributed unevenly over the lake bed of Taihu (Zhu et al. 2006), and selection of the 22 sample sites was done according to this distribution (Fig. 1). The sites were grouped into four representative regions based on nutrient concentrations in the overlying water: river mouth (RM), northern lake (N), western lake (W), and eastern lake (E). As flow decreases away from the river mouth, nutrients and other contaminants are deposited into the sediment. Since most of the pollution in Taihu is produced in the northwestern catchment, the river mouths in the north and west are always perceived as the most polluted parts of the lake. Furthermore, prevailing summer winds blow from south-east, and the western lake features less embayment and less macrophyte coverage than elsewhere, so total nutrients levels are further elevated by accumulation of blooms and sediment re-suspension. Algal blooms have been concentrated in the northern lake in recent years (Qin et al. 2007). The eastern lake is dominated by floating macrophytes, and it is generally the clearest part of Taihu.
Experimental design
Chlorophyll-a (Chl-a), dissolved oxygen, and pH of lake water were measured in situ with a Yellow Springs Instruments (YSI) 6600 V2 multi-sensor sonde at three different depths, which included “surface” (0.5 m below the water surface), “middle” (near the middle of the water column according to the depth in each site), and “bottom” (0.5 m above the sediment).
Three water–sediment columns (50 cm long and 8.4 cm internal diameter) were sampled at each site on the 2nd (eastern and western lake) and 3rd August 2010 (river mouth and northern lake). In addition, 5 L of lake water was collected from each site. One core from each site was sliced open to determine background nitrogen and phosphorus levels in the sediment. Information on sediment characteristics, including particle size distribution, water content, and porosity, was obtained from former determinations (Qin 2008). The remaining two cores were used in the incubation. Data from the same region (four to seven replicates each) were averaged and standard errors (SE) were calculated for each parameter.
Overlying water was siphoned out of each column, mixed with the 5 L of lake water from each site, and sieved through a 75-μm mesh to exclude most phytoplankton. The water was then returned to the experimental columns for the incubation. The background concentrations of total nitrogen (TN), total phosphorus (TP), dissolved total nitrogen (DTN), dissolved total phosphorus (DTP), and dissolved organic carbon (DOC) in water were determined.
Algal bloom scum was collected from Lake Taihu in the Mashan Channel and concentrated by draining through a 75-μm mesh to about 100 mg/L. Three treatments were set up for each sampling site: (1) water–sediment columns (WS) with half water (20 cm) and half sediment (20 cm), (2) water columns (40 cm) with 100 mL algal scum added (WA), and (3) water–sediment columns with 50 mL algal scum added (WSA). The concentration of Chl-a in water from the WA and WSA columns was about 5 mg/L, which is similar to that recorded in field observations during previous Taihu black spot events (Li et al. 2011; Qin et al. 2010).
All the columns were immersed in an opaque square vat filled with lake water, kept at a constant 25°C, with the tops of the columns extending 10 cm above the water line. Columns were covered with aluminum foil and black cloth to exclude light, except during sampling. Columns were sampled nine times, first on day 0 (the initial value), then after 1, 3, 5, 6, 7, 9, 11, and 17 days. Twenty milliliters of water overlying the sediment was withdrawn through a tube in each column. Samples were used to determine concentrations of orthophosphate (PO 3−4 -P) and ammonium (NH +4 -N). In addition, water was sampled on the first (day 0) and last day (day 17) to determine TN and TP, DTN and DTP, and Chl-a. Twenty milliliters of lake water from each site was poured gently into tubes correspondingly along the wall after sampling, making the water volume constant. DO, pH, and water temperature were measured with YSI 6600 V2 multi-sensor sonde. After the incubation, all the sediment cores were sliced across at 3 cm from the surface to determine sediment TN, TP, P bound to Fe (Fe–P), and total organic carbon (TOC) contents.
Chemical analysis
Concentrations of nutrients and Chl-a in the water column were determined according to the textbook of Standard of Lake Eutrophication Survey of China (Jin and Tu 1990). The concentration of PO 3−4 -P in filtered (through GF/F glass membrane) water was determined by spectrophotometry at 700 nm following the molybdenum blue method (Murphy and Riley 1962). DTP in filtered water and TP concentrations were analyzed as for PO 3−4 -P, following digestion with alkaline potassium persulfate (K2S2O8 + NaOH). DTN in filtered water and TN concentrations were analyzed after digestion at a wavelength of 210 nm. NH +4 -N concentration was measured using a flow injection (Skalar SAN++, and Delft in the Netherlands) analyzer. Chl-a concentration was determined by spectrophotometry at wavelengths of 665 nm and 750 nm, following extraction with hot 90 % ethanol (Jespersen and Christoffersen 1987). DOC concentration was determined by the high-temperature combustion on a TOC-V CPN (Shimadzu, and Kyoto in Japan) analyzer. Sediments were air-dried and ground into powder until they could pass a 150-μm mesh. A 0.02 g aliquot of powder was put it into 25 mL deionized water to determine sediment TN and TP with the same method as used for the water samples. The powder was also used to determine Fe–P content by SMT sequential extraction method (Ruban et al. 2001) and TOC content according to GB (national standard method in Chinese) 7857-87 in China (Zhang and Xu 1984).
Statistical analysis
Mean value and standard error calculations and construction of graphics were all carried out using Microsoft Excel 2007. The differences in nutrient levels in different treatments or different regions (Tables 2 and 3) were analyzed by Wilcoxon signed rank test of non-parametric test with SPSS (Statistical Program for Social Sciences) 13.0 software, and significance levels reported as * (significant, 0.01 ≤ p < 0.05) or ** (highly significant, p < 0.01).
Results and discussion
Water and sediment quality at different sites in Taihu
Background values for water quality in different sampling areas are shown in Table 1. Chl-a level was highest in samples taken near the river mouth, and decreased into northern, western, and eastern parts of lake, respectively. DO and pH were correspondingly highest at or near the river mouth. The values of all parameters measured in the water column differed from surface water to bottom water, with the biggest differences within samples taken at the river mouth. In most of the lake, the values of all parameters were higher in surface waters than bottom water, but the reverse was true in the eastern region, where highest values were found at the bottom. This phenomenon may be attributable to the presence of submerged plants in the eastern lake. TN and TP concentrations in the river mouth and western lake were higher than those in the northern and eastern lake. The polluted river was clearly responsible for the large nutrient loads recorded at the river mouth, and high rates of sediment resuspension by wind-induced waves in the western lake increased nutrient levels there. DTN concentration was highest in the western lake and a little lower at the river mouth, which could be due to the uptake of algal growth in the river mouth region in summer. DTP was a little higher at the river mouth than in other regions. DOC also showed highest values at the river mouth.
Sediment properties vary in the different sampling areas (Qin 2008). Sediments in most parts of the lake are silt or clay, only in eastern lake range from silt to sand. Particle size distribution in the north is around 11–15 μm and in the south it ranges from 15 to 27 μm. Water content is lower in river mouth and western lake (35–65 %), and higher in northeastern and southeastern lake (50–80 %). Correspondingly, sediment porosity is higher in river mouth and western lake (1.3–1.8 g/m3), and lower in northeastern and southeastern lake (1.1–1.4 g/m3). TOC content shows highest value (0.91–1.61 %) in southeastern lake, where it is covered with floating macrophytes, and shows lowest value (0.66–0.91 %) in southwestern lake. TOC content in sediment from northern lake and river mouth is 0.71–1.23 % and 0.94–1.33 %, respectively.
Changes of Chl-a, DO concentrations, and pH in overlying water during incubation
Chl-a, which represents the biomass of phytoplankton, decreased gradually during the dark incubations of sediment cores. The final Chl-a concentrations in the WA columns from in each region were 1,733 μg/L (RM), 1,725 μg/L (N), 1,917 μg/L (W), and 2,496 μg/L (E); in the WSA columns, the values were 690 μg/L (RM), 1,660 μg/L (N), 1,607 μg/L (W), and 1,611 μg/L (E), respectively. The decreases in Chl-a suggested that more than two thirds of algal biomass had declined, and with greater declines in WSA than in WA columns, in all regions. Chl-a levels in both WA and WSA columns declined most in samples from the river mouth, but variability between regions was higher in the WSA treatment than in WA. In the WSA treatments, nearly 90 % of algae decayed in columns sampled from river mouth, whereas samples from the eastern lake lost only half their algae.
DO concentrations in overlying water decreased strongly in both WA and WSA treatments, with similar changes observed in each of the four regions (Fig. 2). Rapid declines from around 50 % to 0 % were observed between days 3 and 5 of incubation and levels remained close to 0 % for the remainder of the experiment. DO levels showed least variation in WS columns and the lowest values in samples obtained from river mouth.

DO, PO 3—4 P, and NH +4 -N concentration in overlying water in each treatment
The presence or absence of algae had a marked effect on the pH of overlying water. In water–sediment columns without algae, pH increased slightly over the first few days then decreased again back to the initial value. In contrast, in columns with added algae, pH showed an initial reduction to around 7, where it remained in WA columns, while rallying later in WSA.
Influence of algal bloom degradation on nutrients in overlying water
The water in the WSA columns became black and smelly during incubation, while in WA columns the water went very green. In the absence of algal scum (WS), PO 3−4 -P concentrations in the overlying water remained stable at about 0.022–0.066 mg/L in samples from the northern, western, and eastern lake, while showing a relatively strong increase to 1.216 mg/L in samples from the river mouth. In both WA and WSA treatments, meanwhile, PO 3−4 -P concentrations exhibited an enormous increase (Fig. 2). The initial concentration of PO 3−4 -P averaged 0.003 mg/L, a value that typically increased slowly during the first days of incubation. Only in samples from the river mouth did it show a considerable increment in first 3 days (Fig. 3). Coinciding with algal decay later in the incubation period, PO 3−4 -P concentrations showed a large increase in the WA and WSA columns. WA samples from different regions showed very similar trends, while WSA columns showed some regional variation, with the highest values in samples from the river mouth. Final PO 3−4 -P concentrations reached 7.138–7.686 mg/L in WA and 5.290–7.848 mg/L in WSA, some 1,800–2,600 times the initial values. Apparently, the regional variations in the final concentrations were smaller in the WA treatment than in WSA. In columns obtained from the river mouth, the final PO 3−4 -P concentration in WA columns was slightly lower than for WSA, while elsewhere PO 3−4 -P levels were always higher in WA than in WSA.

Increments of PO 3−4 -P concentration change over time for each region in WA (above) and WSA (below). The black parts in the figure represent the percentage of PO 3−4 -P increment in first three days. PO 3−4 -P did not show comparable increment during day 0-3 in WA, while showed a significant increase in RM and a slight increase in N in WSA. The light gray, white and dark gray parts represent the percentage of PO 3−4 -P increment during day 3-6, 6-9 and 9-17, respectively
NH +4 -N concentrations in overlying water also showed a strong increase after 3 days, with WA samples from different areas exhibiting a similar increasing trend while in the WSA treatments, samples from the river mouth yielded higher values. In the WS columns, however, concentrations of NH +4 -N increased in all cases and reached the comparatively high value of 2.925 mg/L in samples from the river mouth (Fig. 2). In contrast to the trend observed for PO 3−4 -P, NH +4 -N concentrations did not increase consistently. They sometimes decreased during the final days of the incubation, especially in WS treatments. This may have been due to denitrification, considering the low DO and high NH +4 -N values in water. The final NH +4 -N concentrations reached 42.018–53.153 mg/L in WA and 32.370–71.575 mg/L in WSA, 1,600–3,500 times the initial values. Variation between WA samples from the four regions was again much smaller than for WSA.
TP and TN concentrations in water were increased dramatically. Final TP concentrations in the WA columns reached 9.490–11.138 mg/L for the four regions of the lake and TN reached 138.551–164.293 mg/L. Values for WSA treatments were lower in general, with final TP reaching 8.654–11.317 mg/L in the four regions and TN reaching 121.034–146.545 mg/L.
Compared to the WS columns, which lacked any additional algal detritus, samples in the WA and WSA columns exhibited a strong decrease in DO and an increase in nutrient levels in overlying water. Significant between-treatment differences in the concentrations of PO 3−4 -P and NH +4 -N in overlying water for the four regions are shown in Table 2. Nearly all the differences in PO 3−4 -P levels between WS and WA, and between WS and WSA, rate the presence of algae as significant (p < 0.05) or highly significant (p < 0.01), while nearly all the differences between WA and WSA were not significant (p ≥ 0.05). This indicates that a dense algal bloom will have a very considerable effect on levels of PO 3−4 -P in lake water, regardless of the presence or absence of underlying sediments. Nutrient levels increase dramatically as accumulated algae die off and degrade. For NH +4 -N, nearly all the differences between treatments were significant. In columns containing algae, levels of NH +4 -N in overlying water in were much higher than in WS columns.
However, the response to algal decay varied somewhat in different regions of the lake. Significant regional differences in the concentrations of PO 3−4 -P and NH +4 -N in overlying water from all three treatments are shown in Table 3. Highly significant regional differences in nutrient levels occurred in WS, while the differences between each pair of regions were more significant in WSA columns than in WA. The observed differences in nutrient levels between regions in WS and WSA reflect the different physical and chemical characters of the respective sediments. Among the columns with no sediment (WA), only samples from the cleaner eastern lake exhibited PO 3−4 -P levels that differed significantly from those elsewhere. This difference does not appear to be caused by variations in lake water quality. TP was much higher in sediments from the river mouth than other regions before incubation, and showed a marked decline after incubation in WSA columns, while levels in other samples all increased, resulting in large differences in PO 3−4 -P concentrations of overlying water between RM-N, RM-W, and RM-E. A significant difference in NH +4 -N concentrations was observed between water overlying western lake samples and that from the other regions. The western lake yielded the lowest TN contents in sediment and showed the lowest increase in water NH +4 -N concentration in the WS treatment.
In addition to differences in overall quantities of nutrients, different regions also showed different rates of nutrient increase. Figure 3 shows incremental percentages of PO 3−4 -P accumulation at days 0–3, days 3–6, days 6–9, and days 9–17 of incubation. In the WA treatment, the increments for each of the four periods were almost the same for the different regions. Only in the river mouth did it increase slightly in the first 3 days. The WSA treatment showed that in the presence of sediment, PO 3−4 -P in the overlying water increased considerably in river mouth samples over the first 3 days. There was also a slight increase in the northern lake, while hardly any increases were observed in samples from the western and eastern regions. Algal decay led to a particularly rapid elevation of nutrient levels in river mouth samples. PO 3−4 -P concentrations in the river mouth differed significantly between algal treatments with and without sediment (Table 2), though this could be due to different response times. The increments of NH +4 -N accumulation in the river mouth WSA treatment were also disproportionately large in the early days, but the difference was not as pronounced as that in PO 3−4 -P concentration.
Influence of algal bloom degradation on nutrients in sediment
Sediment nutrient levels before and after incubation are shown in Fig. 4. After 17 days’ incubation, TN and TP had been released from sediments in WS columns in almost all regions, in line with the findings of an earlier investigation (Zhang et al. 2006). As a store and supplier of nutrients, sediment can be influenced by external stimulation. Nutrient storage in sediment is stable when external loading is high, but flexible at lower levels so that nutrients can be released when they are needed (Søndergaard et al. 2001). In Lake Taihu, nutrients are generally more concentrated in sediment than in water, and thus have a tendency to be released into overlying water even when there is no disturbance. In the absence of algae (WS), TN and TP levels recorded at the end of the incubation had nearly always decreased. Nutrient release from sediment was highest in the river mouth area, while initial concentrations of N and P were the highest in overlying water and especially in sediment. The greatest increase of N and P in water was recorded there.

TN and TP contents in sediment before incubation (background value) and after incubation in WS and WSA
In the presence of algae, sediment nutrient contents increased in most regions, leading to a disparity between WS and WSA columns. We used sediment porosity values for each region (Qin 2008) to convert the units of nutrient concentration in sediment from milligrams per kilogram into milligrams per liter and thus to show differences in phosphorus concentration between water and sediment (Fig. 5). The TP changes in sediment observed in the WSA treatment compared to background levels seemed relatively small (Fig. 4) and did not show statistically significant differences (p > 0.05), but the changes were comparable with differences of PO 3−4 -P in final water between treatments WA and WSA (Fig. 5), which indicated the differences of available phosphorus concentration in water column between WA and WSA may be induced by phosphorus exchange between water and sediment. We also used changes in post-incubation sediment nutrient levels (Fig. 6) to indicate the risk of nutrient release from sediment during a black spot event. The changes in sediment nutrient levels after incubation with algal scum varied from region to region. The decay of algal material in the WA and WSA columns lead to elevated post-incubation levels of sediment TN in samples from all regions, and TP levels increased in all sediments but those from the river mouth.

Changes in P levels in water and sediment after incubation of samples from four regions. The white bars represent the differences of PO 3−4 -P concentration between WSA and WA in overlying water. PO 3−4 -P concentration in WSA was higher than in WA in river mouth and lower in three other regions. The black bars represent the increments of TP in sediment after incubation

Changes in TP and TN concentrations in sediment after incubation
The degradation of algae consumes much oxygen, creating anoxic conditions in which Fe–P is easy to release from sediment. Fe–P content in sediment from river mouth decreased for 21 % (from 493 to 374 mg/g) and TP content decreased for 4 % after incubation. However, the products of algal decay lead to a surplus of nutrients in the water, and changes in the ratio of water- to sediment-bound nutrient, which may also influence nutrient cycling (Søndergaard et al. 2003). The Fe–P content in other three regions increased 26 % (from 107 to 132 mg/g) and TP increased 5 % after incubation. Nitrogen was released from sediments from almost all regions during algal decay. In a previous experiment, using a less concentrated algal scum, nutrients were released from sediments in the river mouth and other areas where phytoplankton was dominant, while sediment nutrient levels increased in areas dominated by macrophytes (Zhu et al. 2011). Nutrient cycling at the sediment–water interface was influenced by both DO and by nutrient concentration gradients that resulted from algal decay.
Shorelines in northwestern parts of Taihu are prone to massive algal blooms, which accumulate and decay, rendering large areas of water hypoxic/anoxic, elevating nutrient levels, and creating the potential for subsequent black spot events, just like the phenomenon in the columns of the experiment. Sediment is actively involved in the process of bloom degradation in this large, shallow lake and can either stimulate or delay the process. Black spot events occurred mainly in the northwestern lake, and we simulate the process in all over the lake to see the potential influence in different regions. According to the results of the experiment, the polluted sediment at river mouth carries the greatest risk of fostering black spot events. Elsewhere in the northwestern lake, however, surplus nutrients resulting from massive algal decay might still be adequately absorbed by sediment.
Conclusions
Based on the experiment, the decay of algal blooms leads to nutrients increase in water overlying the lake bottom and changes in nutrient flux at the sediment–water interface. However, the presence of sediments can influence these changes and, in comparatively clean areas, may even substantially weaken the impact of intense blooms by absorbing the surplus N and P resulting from algal decay. Different regions of Taihu show different responses to algal blooms in terms of overall nutrient levels and rates of nutrient release. The nutrient-rich sediments located near the river mouth carry the greatest risk of accelerating and enhancing black spot anoxia events, thus special attention should be paid to algal accumulations there.
References
Abell JM, Ozkundakci D, Hamilton DP (2010) Nitrogen and phosphorus limitation of phytoplankton growth in New Zealand Lakes: implications for eutrophication control. Ecosystems 13:966–977
Brookes JD, Carey CC (2011) Resilience to blooms. Science 333:46–47
Chen HS (2011) Black water aggregation in Yixing inshore water area of Taihu Lake. Adv Sci Technol Water Resour 31:33–37
Cheng G (2010) The study on distribution characteristics and regional diversities of nitrogen nutrients in Taihu Lake. University of Lanzhou (in Chinese), Dissertation
Fan C, Chen Y, Wu Q (1998) Effect of prevailing wind in summer on distribution of algal bloom in Lake Taihu. Shanghai Environ Sci 17:4–6 (in Chinese)
Guo L (2007) Ecology—doing battle with the green monster of Lake Taihu. Science 317:1166
Jeppesen E, Kristensen P, Jensen J, Søndergaard M, Mortensen E, Lauridsen T (1991) Recovery resilience following a reduction in external phosphorus loading of shallow, eutrophic Danish lakes: duration, regulating factors and methods for overcoming resilience. Memorie dell' Istituto italiano di idrobiologia dott Marco de Marchi 48:127–148
Jespersen AM, Christoffersen K (1987) Measurements of chlorophyll-a from phytoplankton using ethanol as extraction solvent. Archiv Fur Hydrobiol 109:445–454
Jin XC, Tu QY (1990) Standard of lake eutrophication survey of China. China Environmental Science Publishing House, Beijing (in Chinese)
Kong FX, Hu WP, Gu XH, Yang GS, Fan CX, Chen KN (2007) On the cause of cyanophyta bloom and pollution in water intake area and emergency measures in Meiliang Bay, Taihu Lake in 2007. J Lake Sci 19:357–358 (in Chinese)
Li HB, Xing P, Chen MJ, Bian YQ, Wu QL (2011) Short-term bacterial community composition dynamics in response to accumulation and breakdown of Microcystis blooms. Water Res 45:1702–1710
Likens GE (1971) Nutrients and eutrophication. Science 173:873–874
Liu GF (2009) Black spots of algae blooms effects on biogenic elements cycling in interface of sediment–water of lake Taihu and its pre-control measures research. Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences (in Chinese), Dissertation
Lu GH, Ma Q (2010) Analysis on the causes of forming black water cluster in Taihu Lake. Adv Water Sci 20:438–442 (in Chinese)
Marsden MW (1989) Lake restoration by reducing external phosphorus loading: the influence of sediment phosphorus release. Freshw Biol 21:139–162
Murphy J, Riley J (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 27:31–36
Paerl HW (1988) Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnol Oceanogr 33:823–847
Pu P, Yan J (1998) Taihu Lake—a large shallow lake in the East China Plain. J Lake Sci 10:1–12 (in Chinese)
Qin BQ (2008) Lake Taihu, China. Springer, New York
Qin B, Zhu G, Zhang L, Luo L, Gao G, Gu B (2006) Estimation of internal nutrient release in large shallow Lake Taihu, China. Sci China Ser D: Earth Sci 49:38–50
Qin B, Xu P, Wu Q, Luo L, Zhang Y (2007) Environmental issues of Lake Taihu, China. Hydrobiol 581:3–14
Qin B, Zhu G, Gao G, Zhang Y, Li W, Paerl H, Carmichael W (2010) A drinking water crisis in Lake Taihu, China: linkage to climatic variability and lake management. Environ Manag 45:105–112
Reynolds CS (1987) Cyanobacterial water blooms. Adv Bot Res 13:67–143
Reynolds CS, Walsby A (1975) Water blooms. Biol Rev 50:437–481
Ruban V, López-Sánchez JF, Pardo P, Rauret G, Muntau H, Quevauviller Ph (2001) Harmonized protocol and certified reference material for the determination of extractable contents of phosphorus in freshwater sediments—a synthesis of recent works. Fresenius J Anal Chem 370:224–228
Sas H (1989) Lake restoration by reduction of nutrient loading: expectations, experiences, extrapolations. Academia Verlag Richarz, Saint Augustine
Smith VH (1983) Low nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. Science 221:669–671
Søndergaard M, Jensen J, Jeppesen E (1999) Internal phosphorus loading in shallow Danish lakes. Hydrobiol 408:145–152
Søndergaard M, Jensen J, Jeppesen E (2001) Retention and internal loading of phosphorus in shallow, eutrophic lakes. Sci World J 1:427–442
Søndergaard M, Jensen J, Jeppesen E (2003) Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiol 506:135–145
Xie LQ, Xie P, Tang HJ (2003) Enhancement of dissolved phosphorus release from sediment to lake water by Microcystis blooms—an enclosure experiment in a hyper-eutrophic, subtropical Chinese lake. Environ Pollut 122:391–399
Xu H, Paerl H, Qin B, Zhu G, Gao G (2010) Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China. Limnol Oceanogr 55:420–432
Zhang W, Xu B (1984) Study methods for forest soil. China Forestry Publishing House, Beijing (in Chinese)
Zhang L, Fan C, Wang J, Zheng C (2006) Space–time dependent variances of ammonia and phosphorus flux on sediment–water interface in Lake Taihu. Environ Sci 27:1537–1543 (in Chinese)
Zhu GW (2008) Eutrophic status and causing factors for a large, shallow and subtropical Lake Taihu, China. J Lake Sci 20:21–26 (in Chinese)
Zhu GW, Qin B, Zhang L (2006) Phosphorus forms and bioavailability of lake sediments in the middle and lower reaches of Yangtze River. Sci China Ser D: Earth Sci 49:28–37
Zhu MY, Zhu GW, Wang YP (2011) Influence of scum of algal bloom on the release of N and P from sediments of Lake Taihu. Environ Sci 32:409–416 (in Chinese)
Acknowledgments
This project was supported by the National Natural Science Foundation of China (41171368) and Chinese Ministry of Science and Technology (2008CB418103, 2012ZX07101-010). We thank the Taihu Laboratory for Lake Ecosystem Research (TLLER) for providing the laboratory and equipment. We would like to thank Feng Longqing for his help with sediment sampling, and Xue Jingchen and Zhang Chengying for their help with water quality analysis in laboratory. We would also like to thank Professor Hans Paerl from Institute of Marine Science, University of North Carolina (IMS–UNC) at Chapel Hill for his help in English writing and his scientific suggestions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Rights and permissions
About this article
Cite this article
Zhu, M., Zhu, G., Zhao, L. et al. Influence of algal bloom degradation on nutrient release at the sediment–water interface in Lake Taihu, China. Environ Sci Pollut Res 20, 1803–1811 (2013). https://doi.org/10.1007/s11356-012-1084-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-012-1084-9