
Abstract
Because of human impacts, lowland rivers are among the most degraded running water ecosystems, with their floodplains being the center of human activity. Recently, many programs to restore running water ecosystems have been undertaken using various methods in streams and rivers of North America, Europe, and Far East Asia. However, research and knowledge on the effects of river restoration in lowland rivers are limited around the world. The restoration project involving the first reconstruction of a meandering channel in Asia has been conducted in a lowland river section of the Shibetsu River, northern Japan. We review the geomorphologic and hydraulic characteristics of lowland rivers and their environments for macroinvertebrates and discuss approaches to restoring macroinvertebrate communities in lowland rivers, using insights from the restoration project in the Shibetsu River. It is concluded that the recovery of macroinvertebrate assemblages in channelized lowland rivers requires the implementation of restoration methods to create stable substrates.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Many rivers and streams have been channelized for various purposes across the world. Channelization reduces the heterogeneity of physical habitats in streams and rivers into a homogeneous one. Numerous studies have shown that a loss of habitat diversity caused by channelization has decreased the abundance and taxon richness of macroinvertebrates (e.g., Quinn et al. 1992; Bis et al. 2000; Negishi et al. 2002). Alteration of physical habitats by channelization is the most significant threat to biodiversity and river ecosystem function worldwide (Allan and Flecker 1993; Rosemberg et al. 2000; Nakamura et al. 2002; Nakamura and Yamada 2005). River restoration and rehabilitation to restore biodiversity and ecosystem function appear to be a popular arena of political, social, and scientific investment in Europe and North America. Several restoration programs have recently been conducted in streams and rivers in Far East Asia, such as in South Korea (Shin and Lee 2006) and Japan (Nakamura 2003).
Few other lotic ecosystems have been more altered by human impacts than lowland rivers of temperate regions, because their floodplains have been the center of human activity (Nilsson et al. 2005). In addition, channelization of lowland rivers in these regions has a long history, and therefore, little is known of their original condition. Lack of knowledge on the reference condition complicates the restoration of lowland rivers. However, many restoration projects have endeavored to restore ecosystem processes in lowland rivers (e.g., Brookes and Shields 1996; Waal et al. 1998).
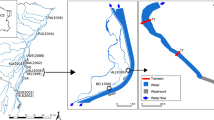
The lower segment of the Shibetsu River in northern Japan historically meandered through its floodplain. Channelization work between the 1950s and 1970s for flood control and farmland reclamation eliminated the physical complexity of in-stream habitats and caused extirpation of native fish species in the Shibetsu River (Hirai and Kuga 2005). Growing interest in river health in Japan has led to a program to reconstruct meanders in the Shibetsu River to restore the river ecosystem (Nakamura 2003). Because of lack of experience in meander restoration in Japan, a small-scale pilot project was launched to test construction methods and examine the response of aquatic biota, about 8.5 km upstream from the river mouth.
Although the number of studies on the responses of stream and river ecosystems to river restoration has increased (Ormerod 2004), the number dealing with lowland rivers is limited. Thus, approaches to restoring lowland river ecosystems have been limited. The objective of this study is to synthesize the findings of previous basic and applied works, which contributes to the success of future restoration projects in lowland rivers. In this paper, we focus on macroinvertebrates, which play an important role in the structure and function of lotic ecosystems (e.g., Covich et al. 1999; Allan and Castillo 2007) and are frequently used as indicators of the ecological status of rivers to evaluate the efficacy of river restoration (Karr and Chu 1999). We initially review the geomorphic and hydraulic characteristics of lowland rivers and their traits as macroinvertebrate habitats. We then describe the responses of macroinvertebrate communities to river restoration in the lower segment of the Shibetsu River, and finally discuss approaches to restoring macroinvertebrate communities in lowland rivers.
Characteristics of lowland rivers as macroinvertebrate habitats
In general, lowland rivers have higher-order channels within an unconstrained valley segment and exhibit less turbulence than mountain streams with a higher gradient (Brussock et al. 1985; Bisson et al. 2006). Reaches of lowland rivers have little longitudinal change in their physical environment, compared with features such as riffle-pool sequences in mountain streams. In contrast to longitudinal change, lateral variations such as asymmetrical cross-section are greater in lowland rivers, because point bars are formed at meander bends by meandering flows. This reach type consists of low-gradient, meandering channels with a predominantly fine substrates (Church 2002). Sediment movement occurs not only during floods, but also in base-flow conditions in thalweg of lowland rivers. This continuous transport of sediments forming an unstable streambed is one of the main characteristics of lowland rivers.
In a lowland river in Germany, Brunke et al. (2002) found that the shifting sand habitat was located in the deep midstream floor of the river, where flow velocity and transport capacity for sediment were high. The abundance and richness of macroinvertebrates in the shifting sand habitat was much lower than in other habitat types such as woody debris, roots, and stable sand located at the stream’s margin with low-flow conditions. Macroinvertebrates in the deep water riverbed at midstream (i.e., thalweg) were depauperate in lowland rivers of Canada (Rempel et al. 2000) and Japan (Nakano and Nakamura 2006a). Shear velocity, which affects riverbed stability, increased with increasing water depth, and was negatively related to the density and richness of macroinvertebrates in these lowland rivers (Rempel et al. 1999; Nakamura et al. 2008). These previous studies indicated that riverbed stability is an important factor determining macroinvertebrate distributions in lowland rivers. In contrast to lowland streams and rivers, macroinvertebrate density and richness were highest in the midstream of riffles, where shear velocity was highest in mountain streams, probably because high shear flow plays an important role in maintaining interstitial spaces between coarse substrates that are suitable for macroinvertebrates (Nakano and Nakamura 2006a). This suggests that the difference in the relationship between macroinvertebrate assemblages and shear velocity between mountain streams and lowland rivers is attributable to streambed stability during base-flow conditions.
Woody debris provides various habitats for macroinvertebrates from mountain streams to lowland rivers (Benke and Wallace 2003). Stabilized woody debris can itself be a major habitat, especially in lowland rivers, because of a lack of stable substrates there (e.g., Benke et al. 1984; Rabeni and Hoel 2000). As well as woody debris, rip-rap generally supports a high abundance and diversity of macroinvertebrates in a lowland river (Brunke et al. 2002). Natural stable substrates including woody debris, roots projecting from the bank, and large stones can be major habitats for macroinvertebrates in lowland rivers. Johnson et al. (2003) showed the importance of large woody debris as a macroinvertebrate habitat and indicated negative impacts of woody debris removal on macroinvertebrate communities in low-gradient streams. In addition, bank protection decreases the recruitment of woody debris and large stones from the stream bank. Furthermore, several previous studies have evaluated the impacts of woody debris removal, including increased water velocity (Gregory 1992), decreased channel stability (Bilby 1984; Heede 1985), and enhanced scouring of sediments (Beschta 1979). Thus, bank protection and woody debris removal for transportation enhancement significantly decrease stable substrates and riverbed stability.
Approaches to restoring macroinvertebrates in lowland rivers
The pilot project to restore a meander course in the Shibetsu River was completed in March 2002 (Fig. 1). Several studies have reported the effects of meander restoration on macroinvertebrate assemblages in lowland rivers of Europe (Friberg et al. 1994, 1998; Biggs et al. 1998). However, relationships between changes in the physical environment and the macroinvertebrate assemblage structure are still ambiguous, because none of these studies analyzed changes in hydraulic parameters associated with re-meandering. To evaluate the effects of changes in physical environments by meander restoration on macroinvertebrates, abundance, richness, and distributions of macroinvertebrates were compared among three reaches: a restored meandering reach (restoration reach), a channelized reach (control reach), and a naturally meandering reach (reference reach), from June 2002 to June 2004 (Nakano and Nakamura 2008). The channelized reach was located upstream of the restoration reach of the Shibetsu River. The reference reach was located in the lower segment of the Nishibetsu River, flowing immediately south of the Shibetsu River. Streambed materials in both rivers consisted mainly of sand and gravel, with an average diameter of approximately 12 mm in the Shibetsu River, and 10 mm in the Nishibetsu River. Shifting sands were observed on the streambed for water depths of more than 50 cm in both rivers. The restoration and reference reaches had asymmetrical cross-section at their channel bends and showed high cross-sectional diversity in physical variables. When five sampling events in the study area were carried out in base flow condition (June 2002, November 2002, June 2003, November 2003, and June 2004), total taxa richness across a reach of two meandering reaches was always higher than that of the channelized reach (Nakano et al. 2005; Nakano and Nakamura 2008). Figure 2 shows the cross-sectional distribution of physical variables (water depth and shear velocity) and macroinvertebrate community structure (taxa richness and total density) in June 2004. Sampling points of physical variables were located at nine equidistant positions along a transect established in each reach, numbered from the left bank (the edges of the left and right bank being 0 and 10, respectively). Four macroinvertebrate samples (400 cm2) were collected from a sampling location (1.0 × 1.0 m quadrat) established at each alternate sampling point (with an odd number). Macroinvertebrate abundance and richness were highest at the shallowest habitats in the two meandering reaches (Fig. 2). In addition, almost all taxa observed in the two meandering reaches were concentrated in the shallowest margins of point bars created at the inside of the bend. In contrast with the meandering reaches, the channelized reach showed a trapezoidal cross-section with a uniform water depth, and had no concentration of macroinvertebrates at any cross-sectional locations (Fig. 2). Macroinvertebrate composition has been improved by the meander restoration through the formation of edge habitat with a shallow depth (Nakano and Nakamura 2008). Caddisflies, Padunia forcipata, which are mainly found in habitats with slow flow conditions were abundant in the edge habitat (Nakano and Nakamura 2006b). In contrast, Ceratopogonidae, which probably tolerate unstable streambeds, were the most abundant taxa in the control reach. Shear velocity increased with water depth and had a negative association with macroinvertebrate abundance and richness (Fig. 3). Even in base flow conditions, increases in shear velocity could induce movement of streambed sediment in lowland rivers. These results suggest that the recovery in macroinvertebrate abundance and diversity in the restoration reach occurred through the creation of a stable riverbed suitable for macroinvertebrates around the stream margins with shallow depth (i.e., low shear stress).

Photograph of the experimental restored meandering reach in the Shibetsu River, northern Japan

Cross-sectional distribution of water depth (top row), shear velocity (second row), taxa richness (third row), and total density (bottom row) at three reaches (reference, restoration, and control) in June 2004. The restoration and the control reaches are in the Shibetsu River. The reference reach is in the Nishibetsu River, located immediately south of the Shibetsu River (modified from Nakano and Nakamura 2008). The same lower-case letters indicate no significant difference (Tukey HSD test, p > 0.05)

Macroinvertebrates density (a) and taxa richness (b) in relation to shear velocity at each sampling point in the three study reaches. The line corresponds to the regression equation (modified from Nakano and Nakamura 2008)
As mentioned above, stable substrates are major habitats for macroinvertebrates in lowland rivers with fine substrates. Thus, large woody debris additions could be another rehabilitation option. Large woody debris additions may stabilize bed materials and provide suitable habitats for macroinvertebrates, although how much bed material could be stabilized with large woody debris are unknown. In our study, bank erosion on the outside of the bend promoted uprooting and falling of riparian trees into the river. Thus, meandering channel flows create two major habitats for macroinvertebrates: the marginal habitat with a stable riverbed at the edges of the point bar (inside of bend), and the wood habitats (trunks, roots, and branches) formed at the meander scroll (outside of the bend) in lowland rivers (Fig. 4). Recovery of the original meandering channel and natural flow regime could be the best way to restore macroinvertebrate assemblages in a channelized lowland river. However, reconstruction of meanders may not be feasible because of social and economic constraints. In particular, setback of artificial dikes to create spaces for a meander channel is one of the most difficult approaches available in the floodplain of lowland rivers. In such a situation, installation of in-stream structures consisting of large woody debris (Erskine and Webb 2003; Brookes et al. 2004; Spanhoff et al. 2006) and boulders (Negishi and Richardson 2003) could serve as an alternative. Nakano and Nakamura (2006b) also demonstrated that placement of groynes created a stable edge habitat and increased the abundance and diversity of macroinvertebrates in a channelized lowland river. We conclude that, in order to recover macroinvertebrate communities in channelized lowland rivers, a variety of restoration methods stabilizing substrates should be implemented.

Two major habitats (stable riverbed formed at the inside of the bend and wood habitat from fallen trees at the outside of the bend) for macroinvertebrates created by meandering flows in lowland rivers
References
Allan JD, Flecker AS (1993) Biodiversity conservation in running waters. Bioscience 43:32–43
Allan JD, Castillo MM (2007) Stream ecology, 2nd edn, Springer, Dordrecht
Benke AC, Wallace JB (2003) Influence of wood on invertebrate communities in streams and rivers. In: Gregory S, Boyer K, Gurnell A (eds) The ecology and management of wood in world rivers. American Fisheries Society Symposium, vol 37, pp 149–177
Benke AC, Van Arsdall TC Jr, Gillespie DM, Parrish FK (1984) Invertebrate productivity in a subtropical blackwater river: the importance of habitat and life history. Ecol Monogr 54:25–63
Beschta RL (1979) Debris removal and its effects on sedimentation in an Oregon Coast Range stream. Northwest Sci 53:71–77
Biggs J, Corfield A, Gron P, Hansen HO, Walker D, Whitfield M, Williams P (1998) Restoration of the rivers Brede, Cole and Skerne: a joint Danish and British EU-LIFE demonstration project, V—short-term impacts on the conservation value of aquatic macroinvertebrate and macrophyte assemblages. Aquat Conserv Mar Freshw Ecosyst 8:241–255
Bilby RE (1984) Removal of woody debris may affect stream channel stability. J For 82:609–613
Bis B, Zdanowicz A, Zalewski M (2000) Effects of catchment properties on hydrochemistry, habitat complexity and invertebrate community structure in a lowland river. Hydrobiologia 133:367–387
Bisson PA, Montgomery DR, Buffington JM (2006) Valley segments, stream reaches, and channel units. In: Hauer FR, Lamberti GA (eds) Methods in stream ecology. Academic, Amsterdam, pp 23–49
Brookes A, Shields FD (1996) River channel restoration: guiding principles for sustainable projects. Wiley, New York
Brookes AP, Gehrke PC, Jansen JD, Abbe TB (2004) Experimental reintroduction of woody debris on the Williams River, NSW: geomorphic and ecological responses. River Res Appl 20:513–536
Brunke M, Hoffmann A, Pusch M (2002) Association between invertebrate assemblages and mesohabitats in a lowland river (Spree, Germany): a chance for predictions? Arch Hydrobiol 154:239–259
Brussock PP, Brown AV, Dixon JC (1985) Channel form and stream ecosystem models. Water Resour Bull 21:859–866
Church M (2002) Geomorphic thresholds in riverine landscapes. Freshw Biol 47:541–557
Covich AP, Palmer MA, Crowl TA (1999) The role of benthic invertebrate species in freshwater ecosystems. Bioscience 49:119–127
Erskine WD, Webb AA (2003) Desnagging to resnagging: new directions in river rehabilitation in Southeastern Australia. River Res Appl 19:233–249
Friberg N, Kronvang B, Svendsen LM, Hansen HO (1994) Restoration of a channelized reach of the River Gelsa, Denmark: effects on the macroinvertebrate community. Aquat Conserv Mar Freshw Ecosyst 4:289–296
Friberg N, Kronvang B, Hansen HO, Svendsen LM (1998) Long-teram, habitat-specific response of a macroinvertebrate community to river restoration. Aquat Conserv Mar Freshw Ecosyst 8:87–99
Gregory KJ (1992) Vegetation and river channel process interactions. In: Boon PJ, Calow P, Petts GE (eds) River conservation and management. Wiley, Chichester, pp 255–269
Heede BH (1985) Channel adjustments to the removal of log steps: an experiment in a mountain stream. Environ Manage 9:427–432
Hirai Y, Kuga K (2005) The approach to nature restoration project for the Shibetsu River. Ecol Civil Eng 7:143–150 (in Japanese with English abstract)
Johnson LB, Breneman DH, Richards C (2003) Macroinvertebrate community structure and function associated with large wood in low gradient streams. River Res Appl 19:199–218
Karr JR, Chu EW (1999) Restoring life in running waters. Island, Washington DC
Nakamura F (2003) Restoration strategies for rivers, floodplain and wetland in Kusiro Mire and Shibetsu River, northern Japan. Ecol Civil Eng 5:217–232 (in Japanese with English abstract)
Nakamura F, Yamada H (2005) The effects of pasture development on the ecological functions of riparian forests in Hokkaido in northern Japan. Ecol Eng 24:539–550
Nakamura F, Jitsu M, Kameyama S, Mizugaki S (2002) Change in riparian forests in the Kushiro Mire, Japan, associated with stream channelization. River Res Appl 18:65–79
Nakamura F, Kawaguchi Y, Nakano D, Yamada H (2008) Ecological responses to anthropogenic alternations of gravel-bed rivers in Japan, from floodplain river segments to the microhabitat scale: a review. In: Habersack H, Piegay H, Rinaldi M (eds) Gravel-bed rivers VI: from process understanding to river restoration. Elsevier, Amsterdam, pp 501–523
Nakano D, Nakamura F (2006a) Lateral variation of lotic macroinvertebrate community from the headwater to lowland meandering stream in the Nishibetsu River basin, northern Japan. Verh Internat Verein Limnol 29:1377–1382
Nakano D, Nakamura F (2006b) Responses of macroinvertebrate communities to river restoration in a channelized segment of the Shibetsu River, northern Japan. River Res Appl 22:681–689
Nakano D, Nakamura F (2008) The significance of meandering channel morphology on the diversity and abundance of macroinvertebrates in a lowland river in Japan. Aquat Conserv Mar Freshw Ecosyst (in press)
Nakano D, Nunokawa M, Nakamura F (2005) Changes in distribution and structure of macroinvertebrate community before and after re-meandering experiment. Ecol Civil Eng 7:173–186 (in Japanese with English abstract)
Negishi JN, Richardson JS (2003) Responses of organic matter and macroinvertebrates to placements of boulder clusters in a small stream of southwestern British Columbia, Canada. Can J Fish Aquat Sci 60:247–258
Negishi JN, Inoue M, Nunokawa M (2002) Effects of canalization on stream habitat in relation to a spate and flow refugia for macroinvertebrates in northern Japan. Freshw Biol 47:1515–1529
Nilsson C, Reidy CA, Dynesius M, Revenga C (2005) Fragmentation and flow regulation of the world’s large river systems. Science 308:405–408
Ormerod SJ (2004) The golden age of river restoration science? Aquat Conserv Mar Freshw Ecosyst 14:543–549
Quinn JM, Williamson RB, Smith RK, Vickers ML (1992) Effects of riparian grazing and channelization on streams in Southland, New Zealand 2. Benthic invertebrates. N Z J Mar Freshw Res 26:259–273
Rabeni CF, Hoel SM (2000) The imporatance of woody debris to benthic invertebrates in two Missouri prairie streams. Verh Internat Verein Limnol 27:1499–1502
Rempel LL, Richardson JS, Healey MC (1999) Flow refugia for benthic macroinvertebrates during flooding of a large river. J N Am Benthol Soc 18:34–48
Rempel LL, Richardson JS, Healey MC (2000) Macroinvertebrate community structure along gradients of hydraulic and sedimentary conditions in a large gravel-bed river. Freshw Biol 45:57–73
Rosemberg DM, Mccully P, Pringle CM (2000) Global-scale environmental effects of hydrological alternations: introduction. Bioscience 50: 746–751
Shin J-H, Lee I-K (2006) Cheong Gye Cheon restoration in Seoul, Korea. Proc Inst Civil Eng Civil Eng 159:162–170
Spänhoff B, Riss W, Jäkel P, Dakkak N, Meyer EI (2006) Effects of an experimental enrichment of instream habitat heterogeneity on the stream bed morphology and chironomid community of a straightened section in a sandy lowland stream. Environ Manage 37:247–257
Waal LC, Large ARG, Wade PM (1998) Rehabilitation of rivers. Wiley, New York
Acknowledgments
The research into responses of macroinvertebrates to river restoration in the Shibetsu River was supported by the “River Ecology Research Group of Japan” on the Shibetsu River and Grants in Aid for Scientific Research (Nos. 17380100, 18201008, and 19208013) from the Ministry of Education, Science, and Culture, Japan. Daisuke Nakano is supported by MEXT through Special Coordination Funds for Promoting Science and Technology.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nakano, D., Nagayama, S., Kawaguchi, Y. et al. River restoration for macroinvertebrate communities in lowland rivers: insights from restorations of the Shibetsu River, north Japan. Landscape Ecol Eng 4, 63–68 (2008). https://doi.org/10.1007/s11355-008-0038-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11355-008-0038-3