
Abstract
Lygodium microphyllum is an invasive exotic plant species taking over many sites in freshwater and moist habitats in Florida. Managing it has been a significant challenge for land resource managers and researchers due to its extensive rapid invasion. To assess the effects of soil pH on growth, nutrient uptake, and mycorrhizal colonization in the roots of L. microphyllum, we conducted a 60-day greenhouse experiment by growing it in pots filled with pH-adjusted soils to a range from 4.5 to 8.0. L. microphyllum was able to survive and grow at all soil pH levels; however, final biomass, relative growth rate, photosynthesis, and specific leaf area were all greater in soil pH 5.5–6.5 compared to the other treatments. Correspondingly, nitrogen concentration was also related to these four plant parameters. Root colonization by mycorrhizal fungi was higher in soil pH 5.5–7.5 and lowest for plants growing in 4.5 or 8.0 and was correlated with plant growth parameters as well as elemental concentration in the leaves. Soil pH 8.0 was not strong enough for a pronounced growth decline, thus further increasing soil pH could provide a desired outcome and merit further investigation, although its potential negative impact on native flora (both plants and microorganisms) would need to be assessed.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil pH is an important factor for plant growth, as it affects nutrient availability, nutrient toxicity, and has a direct effect on the protoplasm of plant root cells (Rorison 1980; Alam et al. 1999). It also affects the abundance and activity of soil organisms (from microorganisms to arthropods) responsible for transformations of nutrients (De Boer and Kowalchuk 2001; Nicol et al. 2008). Since most mineral nutrients are readily available to plants when soil pH is near neutral (pH = 6.5–7.5), species richness is high in such neutral soils, declining in both acidic and alkaline soils (Grime 1973; Gould and Walker 1999; Pausas and Austin 2001). Soil pH further influences the fate of chemicals, nutrients, and pesticides/herbicides added to the soil (Liu et al. 2001). Past research has shown that the species diversity is low in most acidic soils (Dupré et al. 2002) as essential nutrients (such as Ca, Mg, K, PO4, and Mo) exist in unavailable forms to plants causing nutrient deficiency (Larcher 2003). Likewise, due to the higher sensitivity of nitrate bacteria, nitrification is significantly slowed down with faster rates of ammonia oxidation than the oxidation of nitrite (Smith et al. 1997). This results in the accumulation of nitrite, which can be toxic to plant and microorganisms in acidic soil (Black 1957; Shen et al. 2003). In strongly acidic soils, certain ions (Al3+, Cu2+, Fe3+, Mn2+) rise to levels toxic for the majority of plants (Foy 1992; Kinraide 1993; Silva 2012). Additionally, acidic soils have high cation exchange capacity, and promote leaching of nutrients resulting in soil unfavorable for plant growth (Johnson 2002). At the other extreme, alkaline soils tend to be unfavorable for plant growth with iron, manganese, and phosphate deficiency (Marschner 1995; Tyler 1999) creating an unfavorable condition for plant growth. Marschner (1995) suggests that in alkaline soils, boron can rise to phytotoxic concentrations.
Plants differ enormously in their degree of tolerance to changes in soil characteristics (pH, moisture content, etc.): some have a narrow tolerance for one variable but a wide tolerance for others (Hill and Ramsay 1977). Weedy species collected from different climate zones show large growth differences when planted in soils with pH ranging from 4.8 to 6.4 (Buchanan et al. 1975). Stephenson and Rechcigl (1991) found that many weedy species grew significantly better when soil pH increased from 4.5 to 5.4, with good growth maintained at pH of 5.5 and above.
Lygodium microphyllum is an invasive exotic plant species taking over many sites in freshwater and moist habitats in Florida. It has the ability to grow in varying hydrological (Gandiaga et al. 2009), nutrient (Volin et al. 2010), and light conditions (Volin et al. 2004). Analysis of soil samples from both its native range and invaded region has shown that although L. microphyllum grows in highly acidic soils in its native range in Australia, it is thriving in close-to-neutral soils in Florida (Soti et al. 2014). Additionally, the roots of L. microphyllum were heavily colonized by mycorrhizal fungi which assist the plant to absorb nutrients, specifically P, leading to almost three times higher biomass accumulation in mycorrhizal plants compared to non-mycorrhizal plants (Soti et al. 2014). Furthermore, the level of mycorrhizal colonization was related to soil pH: a higher degree of mycorrhizal colonization is present in plants from the slightly acidic soils (pH 5.5–6.0) in the invaded regions compared with those from the highly acidic soil (pH 4–4.5) in the native regions (Soti et al. 2014).
The aim of this study was to compare the degree of mycorrhizal colonization, nutrient uptake, biomass accumulation, and growth rate of L. microphyllum at different soil pH levels. Since the existing chemical control method is not very efficient in controlling L. microphyllum, this information may be useful in developing an integrated weed management technique. In this study, we hypothesized that growth of L. microphyllum and the mycorrhizal association will be highest in slightly acidic soils with growth highly reduced (or the plants not surviving) in alkaline soils. We predicted that changing the soil pH can reduce the extensive growth rate of L. microphyllum, with a significant influence in the mycorrhizal fungi in its roots.
Methods
To test the hypothesis, we undertook a greenhouse experiment to investigate the effects of soil pH on various aspects of growth of L. microphyllum. Different soil pH levels were selected to include a wide range of soil pH where L. microphyllum has been reported to grow in its native range in Australia and the invaded regions in Florida (for details see Soti et al. 2014). Fourteen-week-old plants were maintained in pots in the Florida International University greenhouses until they began to sporulate, at which time the experiment (described in detail below) was concluded.
Potting soil
Sandy loam soil from plots in Tree Tops County Park at Davie, Florida was collected for this study; this site was not yet invaded by L. microphyllum but had the potential to be invaded as the plant density increased. Soil was then passed through a 2-mm sieve and air dried at room temperature. Quartzite sand was added to this soil to form a 1:1 soil/sand ratio. A sub sample of the soil was analyzed to determine initial soil characteristics. Soil pH was measured with a pH meter (soil solution ratio 1:2 in water); texture was measured by the hydrometer method; total organic matter was measured based on the standard loss on ignition method (500 °C, 5 h; Storer 1984). A soil neutralization curve (Kellogg et al. 1957) was generated to determine the amount of Ca(OH)2 or S to be added in the potting soil. To generate the curve, 150 g of air-dried soil samples were placed in 120-mL plastic containers and mixed with Ca(OH)2 at rates 0, 1, 2, 3, 4, 5, 6, 7, 8 g Kg−1; elemental S was added at rates 3.5, 4.0, 4.5, 5.0, and 6.0 mg Kg−1, with 5 replicates for each treatment. These soil samples were watered with DI water and incubated for 28 days and the soil pH was measured (soil solution ratio 1:2 in water). The amount of lime or sulfur required to raise the experimental soil pH to the desired level was determined based on the regression equation resulting from pH measurement of the incubated soils. The rate of Ca(OH)2 and S added is given in Table 1.
The soil was a sandy loam with 37 % organic matter, 1.10 mg/g total P, 1.2 % total N and pH 5.5 ± 0.2. This soil was divided into 5 subsamples and elemental sulfur was added to lower the pH to 4.5 in one set; no treatment was done in the 5.5 pots, and Ca(OH)2 was added to increase the pH to 6.5, 7.5, and 8.0. Soil samples with elemental sodium or Ca(OH)2 were thoroughly mixed, added to pots and watered with DI water. The soils were allowed to equilibrate for 8 weeks with frequent mixing. Soil pH was measured weekly and after 8 weeks, all pH measurements were within ± 0.3 of the targeted pH value and remained constant throughout the experiment time.
Plant material
Experimental plants were grown from spores following the method used by Lott et al. (2003). Spores of L. microphyllum were disinfected with 1 % bleach and transferred to Petri dishes that contained Parker-Thomson Medium. The plates were placed in an incubator set at 25–27 °C for 10 weeks and were watered with sterile DI water every week. After 10 weeks, individual gametophytes were transferred to fresh Petri dishes. When the sporelings’ roots and leaves developed, 60 plants with similar size were selected and were transplanted to small pots previously filled with sterile sand. These 60 plants were placed in a growth chamber for approximately 4 weeks. The plants were kept very moist, and were watered with half strength Hoagland’s nutrient solution. At time 0 (the beginning of the treatment/transplant date), 20 plants were randomly selected and destructively harvested to develop an allometric relationship discussed below. Thirty plants with similar size were transferred to 2.5-L pots filled with pH modified soil. These plants were grown in the green house for 60 days; watered to saturation biweekly and received 250 mL of half strength Hoagland’s solution weekly. Before the plants were harvested, photosynthesis was measured using a Li-Cor 6400 Portable Photosynthesis System (Li-Cor Biosciences, Lincoln, Nebraska) on two fully grown leaves per plant in all the treatments. Measurements were taken at leaf temperatures ranging from 34 to 38 °C; CO2 concentration of 400 µmol mol−1 and photosynthetic photon flux was at 600 µmol m−2 s−1. Dry mass-based photosynthesis was calculated as photosynthesis × specific leaf area (SLA).
Harvest and plant nutrient analysis
Two harvests were conducted during this study: at time 0, and after 60 days. The allometric relationship between stem length and total mass (R2 = 0.92) from the initial harvest was developed to estimate the initial plant mass of the experimental plants and to calculate the relative growth rate (RGR) (Gandiaga et al. 2009). The RGR (mg g−1 day−1) was calculated for each individual plant used for the experiment following the method by Evans (1972):
where M1 and M2 are the dry biomass at the beginning of the experiment (T1) and end of the experiment (T2), respectively. After each harvest, individual plants’ roots, rhizomes, stem, and leaves were separated and leaf area was measured with a leaf area meter to calculate the SLA (leaf area/total leaf mass). The separated plant parts were oven dried (1 week at 65 °C) to constant mass and weighed to determine the leaf mass ratio (LMR) (leaf mass/total plant mass), stem (rachis) mass ratio (SMR) (stem mass/total plant mass), rhizome mass ratio (RhiMR) (rhizome mass/total plant mass), and root mass ratio (RMR) (root mass/total plant mass); and RGR.
The above ground parts of the plants (shoot tissue) were analyzed for nutrient content. The oven-dried tissues were carefully ground by hand using a mortar and a pestle. Samples underwent acid digestion using Method 3050B (U.S. Environmental Protection Agency 1996). One gram of finely ground plant tissue sample was transferred to a large glass tube and mixed with 10 mL of 30 % HNO3. The tubes were covered with a vapor recovery system and heated to 95 ± 5 °C and refluxed for 10 min under the hood in a heating block maintained with a Partlow Mic 6000 Profile Process Controller (Despatch Industries, Minneapolis, MN). After cooling to 40 °C, 5 mL of concentrated HNO3 was added and the sample was heated again until no brown fumes were given off. After cooling to 40 °C, 2 mL of DI water and 3 mL of 30 % H2O2 were added and heated until the effervescence subsided. The samples were cooled and diluted to 100 mL with DI water, centrifuged at 2000 rpm for 10 min and filtered with Whatman No. 41 filter paper and analyzed with an ICP-OES (Inductively Coupled Plasma-Optical Emission Spectrometry) at USDA ARS, Homestead, Florida.
Mycorrhizal colonization
Before drying, forty-five 1.5-cm root fragments were collected from each plant, and the colonization of AMF was quantified following the magnified intersect method described by (McGonigle et al. 1990). Roots were cleared in 15 % KOH at 70 °C for 4 h, rinsed twice with water, bleached with ammoniated H2O2, and acidified with 1 N HCl. Staining was done using Trypan blue in acidic glycerol at 80 °C for 20 min. The stained roots were mounted parallel on slides in lactoglycerol covered with coverslip and examined for fungal structures at × 100 magnification. Root segments were considered mycorrhizal if extratadical mycelium, intraradical hyphe, arbuscules, or vesicles were observed.
Experimental design and data analysis
The experimental design was a randomized complete block with five pH treatments and six replicates. It was a single-factor experiment investigating the effects of pH on plant growth, nutrient accumulation, and level of mycorrhizal colonization. After the harvest at 60 days, regression analysis was done to examine the influence of initial plant mass on RGR and its morphological, allocational, and physiological determinants (e.g., McConnaughay and Coleman 1999; Volin et al. 2002; Kruger and Volin 2006). Regression analysis indicated that RGR was negatively correlated to the natural log (ln) of initial plant mass (P < 0.001). Additionally, RMR final harvest was significantly related (P < 0.05) to final plant mass. Therefore, treatment effect on RGR was analyzed after it was normalized for variation in plant mass using analysis of covariance. All of the variables in the five pH treatments were then compared with one-way ANOVA for significance at P ≤ 0.05. Means were separated using Tukey HSD test. Correlation analysis between total biomass, RGR SLA, and leaf concentration of Al, Ca, P, N, and Fe was done to determine the effects of leaf elemental status on plant growth. Regression analysis was done to analyze the relationship between the plant growth parameters and N concentration in the leaves. In addition, regression analysis was done to examine the relationship between RGR and its determinants (morphological: SLA; physiological: photosynthesis; and biomass allocation: LMR, RMR, RhiMR, SMR). All analyses were performed with SAS Version 9.2 software (SAS Institute 2009).
Results
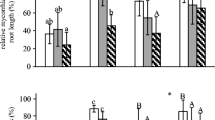
A significant effect of soil pH was visible on L. microphyllum growth, nutrient uptake, and degree of mycorrhizal colonization in its roots, despite the small sample size (n = 6) and short duration (60 days) of this experiment. Total final plant mass was greatest in plants grown in soil with pH 6.5 and 5.5, and these were more than twice the biomass of plants grown in pH 8.0 (Fig. 1a). The growth of L. microphyllum was greater in pH 5.5 and 6.5 compared to the strongly acidic and alkaline soils (Fig. 1b). Surprisingly, biomass allocation to the above ground parts was not influenced by soil pH (data not shown). There was, however, difference in biomass allocation to the belowground parts. Relative growth rate and biomass allocation patterns were different among the pH treatments (Fig. 1b–d). Correspondingly, this pattern was found for RGR, which increased with increasing soil pH from 4.5 to 5.5, remained unchanged at 6.5, and gradually declined with increasing soil pH, with lowest RGR at soil pH 8.0 (Fig. 1b).

Mean (± SE) final biomass (a); mean (± SE) relative growth rate (RGR) (b); mean (± SE) root mass ratio (RMR) (c); and mean (± SE) rhizome mass ratio (RhiMR) (d), measured at the end of 60 days from plants grown at different soil pH levels. Similar letters represent no significant difference at P < 0.05
Plants in soil pH 5.5 and 6.5 had higher SLA than the other plants (Fig. 2). The influence of soil pH also was strongly reflected in the photosynthetic rates, which showed the same response as SLA (Fig. 3). As a result of higher SLA and area-based photosynthesis, mass-based photosynthesis was also higher in plants grown in pH 5.5 and 6.5 (Fig. 3b).

Mean (± SE) value of area-based photosynthesis at different pH levels. Similar letters represent no significant difference at P ≤ 0.05

Mean (± SE) value of area-based photosynthesis (a); mass-based photosynthesis (b) at different pH levels. Similar letters represent no significant difference at P ≤ 0.05
Element concentration and uptake
Soil pH affected the concentrations of Al, Ca, Fe, and N in the leaf tissue of L. microphyllum (Table 2), while it did not have any influence on the leaf concentration of P, K, Mg, Mn, and Zn. Plants grown in soil pH 4.5 had higher concentration of Al and Fe and lower concentrations of N than plants grown in higher soil pH. Similarly plants grown in pH 8.0 had highest concentration of Ca. There was a strong positive relationship between biomass (R 2 = 0.61, P < 0.0001), RGR (R 2 = 0.65, P < 0.0001), SLA (R 2 = 0.55, P < 0.0001) photosynthesis (R 2 = 0.64, P < 0.0001)) and RhiMR (R 2 = 0.63, P < 0.0001) and leaf concentration of N when all the treatments were pooled. However, there were no correlations between the plant growth parameters and leaf concentration of Al, Ca, Fe, Mg, Mn, P, and Zn.
Mycorrhizal colonization
As expected, soil pH also had an effect on the degree of mycorrhizal colonization (Fig. 4). The degree of colonization was highest at pH 5.5 with no difference at pH 6.5 and 7.5, while the degree of colonization was lower at both pH 4.5 and pH 8.0. There was no significant difference in the mycorrhizal structures such as vesicles, arbuscules, and hyphae among the four soil pH levels.

Mean value (± SE) of degree of mycorrhizal colonization at different pH levels. Similar letters represent no significant difference at P ≤ 0.05
When the pH treatments were analyzed independently there was no strong correlation between the root colonization by mycorrhizal fungi and plant growth parameters or the leaf concentration of elements. However, when the samples were pooled there was a strong correlation between the degree of mycorrhizal colonization and plant growth parameters as well as the leaf element status (Table 3). Additionally, there was a correlation between the degree of mycorrhizal colonization and element uptake by L. microphyllum.
Discussion
Soil pH had a significant effect on the growth, biomass allocation, nutrient uptake, and mycorrhizal colonization in L. microphyllum. At soil pH 4.5 and 8.0, the plants were noticeably smaller and grew less vigorously compared to the other treatments. Soil pH 5.5 and 6.5 provided the most favorable conditions for the nutrient uptake, growth, and biomass accumulation. This result was expected for L. microphyllum because extensive growth is seen in slightly acidic soils of Florida. Few other ferns in Florida have been reported to prefer soil pH close to 6.0; field study by Van Loan (2006) showed that Lygodium japonicum, another pest plant species in Florida, was present in sites with soil pH 6.0. Similarly, Mathur et al. (1982) reported that the fern Rumohra adiantiformis requires soil pH between 5.5 and 6.0 for optimal growth. However, as opposed to our expectation, the plants grown in alkaline soils survived, maintained a fair growth rate and produced fertile fronds.
In its native range, L. microphyllum grows in highly acidic soils (soil pH range 3.9–4.7) (Soti et al. 2014); this adaptive capability of L. microphyllum to acidic soils was visible in our study. Contrary to our expectation plants grown in soil pH 8.0 maintained a fair growth rate. High SLA and RGR are reported as typical characteristics of invasive species and remain highly associated with invasiveness (Grotkopp and Rejmánek 2007). The high RGR of plants grown in soil pH 5.5 and 6.5 corresponded to the higher mass-based photosynthesis, which resulted from the increased SLA as well as area-based photosynthesis (Lambers and Poorter 1992). Our results indicate that variation in RGR was correlated to the variation in photosynthetic capacity and SLA of L. microphyllum.
Our results show that the biomass allocation to the belowground structures was different across the different pH treatments. An unexpected result was that plants grown in strongly acidic soils allocated the highest biomass to the roots compared to the plants grown at higher pH values at the cost of biomass allocation to the rhizomes. This higher biomass allocation to the roots in acidic soils is different that the common assumption that Al toxicity in acidic soil causes a significant reduction in root growth by inhibiting cell division in the root apical meristem (Ryan et al. 1993; Crawford and Wilkens 1998) resulting in reduced water and nutrient uptake. Abrahamsen (1983) suggested that certain plant species have the ability to translocate the Al absorbed from roots to other parts of plant to avoid Al toxicity; element concentration in the roots were not measured in our study but this may be one explanation for the extensive root growth in the plants in soil pH 4.5 and would need to be substantiated in future research. Additionally, there is a possibility of root exudates by L. microphyllum as a defense mechanism to Al toxicity. This ability to avoid Al toxicity in acidic soils could in part explain the extensive growth of L. microphyllum in the sand mine spoils with high levels of Al and Fe in central Florida (Soti, pers. obser.). Schindelbeck and Riha (1988) and Kidd and Proctor (2001) have found that decrease in soil pH caused an increased biomass allocation to roots; Bates et al. (2002) found that when the soil pH was lower than 4.4 the root, shoot ratio increased in Vitis labruscana L. plants.
Phenotypic plasticity is one of the key characteristics of invasive plants which allow them to adapt to a wide range of habitat types (Claridge and Franklin 2002). Previous studies have shown that L. microphyllum is extremely plastic in its ability to respond to myriad environmental conditions, including plasticity in reproduction, physiology, biomass allocation, and morphology (see: Lott et al. 2003; Gandiaga et al. 2009; Volin et al. 2004, 2010). Our results suggest the possibility of a strong phenotypic plasticity in L. microphyllum in response to changes in growth environment. Similar phenotypic plasticity in response to environmental conditions has been reported in Melaleuca quinquenervia, a flowering tree which shares habitat with L. microphyllum in the native range in Australia and is invasive in most of the regions in south Florida, where 97 % of its variation was accounted by soil pH (Kaufman and Smouse 2001).
Soil pH had a significant effect on the element uptake by L. microphyllum. Higher concentration of Al in the leaves of plants growing in acidic soils did not substantially alter the growth parameters of L. microphyllum. It is reported that Al and Mn toxicity occur in soil when the pH is below 4.8 (Slattery et al. 1999), but L. microphyllum plants grown in strongly acidic soils did not show any sign of toxicity. Phosphorus is reported to form insoluble compounds under high soil pH conditions, causing P deficiency in plants (Shen et al. 2011). In our study there was no significant difference in the leaf concentration of P among the various soil pH treatments; this could have been in part influenced by arbuscular mycorrhizal fungi (AMF).
Degree of mycorrhizal colonization in the roots of L. microphyllum was significantly low in extremely acidic and alkaline soil. The high degree of mycorrhizal colonization in the roots of soil pH ranging from 5.5 to 7.5 in our greenhouse experiment is comparable to the field study results (Soti et al. 2014). There is some published evidence that plant and mycorrhizal fungi can modify the soil pH in the rhizosphere (Jones et al. 2004). This was not analyzed in our study and merits further research. The negative correlation between degree of mycorrhizal colonization and RMR and the influence of low pH in root growth indicates that the response in root growth could be mediated the mycorrhizal fungi response to soil pH. Mycorrhizal colonization had a strong influence in the growth and N uptake in L. microphyllum. This result is comparable to previous studies on the relationship between mycorrhizal fungi and invasive plant species (Fumanal et al. 2006; Callaway et al. 2008; Soti et al. 2014) Our results indicate that the ability of L. microphyllum to nutrient poor habitats in south Florida could be mediated by its strong association with mycorrhizal fungi in various soil conditions. These findings have important implications for the development of an integrative management technique to reduce the extensive growth of L. microphyllum in the south Florida natural areas.
In our study, the major element influencing plant growth was N, which acts as a direct growth regulator in plants Marschner (1995). Its concentration was significantly higher in plants grown in soil pH 5.5 and 6.5, and strongly correlated with the RGR, photosynthesis, RhiMR, and SLA. A positive correlation between leaf N concentration and RGR, SLA, and photosynthetic capacity have reported for a wide range of plants (Poorter et al. 1990; Poorter and Bergkotte 1992; Reich and Walters 1994; Nielsen et al. 1996; Cornelissen et al. 1997; Reich et al. 1998). This higher concentration of N in soil pH 5.5 and 6.5 is directly correlated with the higher degree of mycorrhizal colonization at these pH levels.
L. microphyllum is reported to have most of the traits of an aggressive invader, including its reproductive characteristics and its lack of a significant pathogens or herbivores in its introduced range (Volin et al. 2004, 2010; Gandiaga et al. 2009; Soti et al. 2014). Our results show that even though soil pH levels 5.5–6.5 were optimal for rapid growth, biomass accumulation and mycorrhizal colonization in roots, L. microphyllum can maintain a fair growth rate over a wide range of soil pH, indicating a continuing threat to most uninvaded sites.
Burning and application of herbicides are the most commonly used methods to control L. microphyllum. Loveless (1959) found that burning raises the soil pH from 4.5 to 5.8 in tree islands of the northern Everglades. We found that increasing soil pH from highly acidic to 5.5–6.5 soil pH provides a more favorable condition for L. microphyllum growth. Prescribed burning, which causes a temporary rise in soil pH, is a widely used method to control L. microphyllum, but our study shows that L. microphyllum could be benefiting from the slight increase in soil pH resulting from fire as well as the release of nutrients that are associated with burning. In our study, L. microphyllum had highest growth at neutral soil pH and began to show a significant decrease at a soil pH of 8.0, likely further growth reductions would happen in even more alkaline soils. Thus, raising soil pH may be a possible management option to explore in the future, but increasing the soil pH would need to be studied carefully for its potential adverse effects to native flora as well, including both native plants and soil microorganisms.
References
Abrahamsen G (1983) Sulphur pollution: Ca, Mg and Al in soil and soil water and possible effects on forest trees. In: Effects of accumulation of air pollutants in forest ecosystems. Springer, New York
Alam SM, Naqvi SSM, Ansari R (1999) Impact of soil pH on nutrient uptake by crop plants. Handbook of Plant and Crop Stress, Madison
Bates TR, Dunst RM, Taft T, Vercant M (2002) The vegetative response of ‘Concord’ grapevines to soil pH. HortScience 37:890–893
Black CA (1957) Soil-plant relationships, 2nd edn. Wiley, New York
Buchanan G, Hoveland C, Harris M (1975) Response of weeds to soil pH. Weed Sci 23:473–477
Callaway RM, Cipollini D, Barto K, Thelen GC, Hallett SG, Prati D, Stinson K, Klironomos J (2008) Novel weapons: invasive plant suppresses fungal mutualists in America but not in its native Europe. Ecology 89:1043–1055
Claridge K, Franklin SB (2002) Compensation and plasticity in an invasive plant species. Biol Invasions 4:339–347
Cornelissen J, Werger M, Castro-Diez P, Van Rheenen J, Rowland A (1997) Foliar nutrients in relation to growth, allocation and leaf traits in seedlings of a wide range of woody plant species and types. Oecologia 111:460–469
Crawford SA, Wilkens S (1998) Effect of aluminium on root elongation in two Australian perennial grasses. Funct Plant Biol 25:165–171
De Boer W, Kowalchuk G (2001) Nitrification in acid soils: micro-organisms and mechanisms. Soil Biol Biochem 33:853–866
Dupré C, Wessberg C, Diekmann M (2002) Species richness in deciduous forests: effects of species pools and environmental variables. J Veg Sci 13(4):505–516
Evans GC (1972) The quantitative analysis of plant growth, vol 1. Univ of California Press, Berkeley
Foy CD (1992) Soil chemical factors limiting plant root growth. In: Hatfield JL, Stewart BA (eds) Limitations to plant root growth. Springer, New York, pp 97–149
Fumanal B, Plenchette C, Chauvel B, Bretagnolle F (2006) Which role can arbuscular mycorrhizal fungi play in the facilitation of Ambrosia artemisiifolia L. invasion in France? Mycorrhiza 17:25–35
Gandiaga S, Volin J, Kruger E, Kitajima K (2009) Effects of hydrology on the growth and physiology of an invasive exotic, Lygodium microphyllum (Old World climbing fern). Weed Res 49:283–290
Gould WA, Walker MD (1999) Plant communities and landscape diversity along a Canadian Arctic river. J Veg Sci 10:537–548
Grime JP (1973) Competitive exclusion in herbaceous vegetation. Nat UK 242:344–347
Grotkopp E, Rejmánek M (2007) High seedling relative growth rate and specific leaf area are traits of invasive species: phylogenetically independent contrasts of woody angiosperms. Am J Bot 94(4):526–532
Hill SB, Ramsay J (1977) Weeds as indicators of soil conditions. The McDonald J 38:8–12
Johnson C (2002) Cation exchange properties of acid forest soils of the northeastern USA. Eur J Soil Sci 53:271–282
Jones DL, Hodge A, Kuzyakov Y (2004) Plant and mycorrhizal regulation of rhizodeposition. New Phytol 163(3):459–480
Kaufman SR, Smouse PE (2001) Comparing indigenous and introduced populations of Melaleuca quinquenervia (Cav.) Blake: response of seedlings to water and pH levels. Oecologia 127:487–494
Kellogg CE, Allaway WH, Barnes CP, Harper VL, Heisig CP, Pierre WH, Pinches HE, Quisenbery KS, Ritchie RG, Thorne W, Stefferud A, Wadleigh CH, Winters E (1957) The Year Book of Agriculture: soil. US Government Printing Office, Washington, DC, p 164
Kidd PS, Proctor J (2001) Why plants grow poorly on very acid soils: are ecologists missing the obvious? J Exp Bot 52:791–799
Kinraide TB (1993) Aluminum enhancement of plant growth in acid rooting media. A case of reciprocal alleviation of toxicity by two toxic cations. Physiol Plant 88:619–625
Kruger EL, Volin JC (2006) Reexamining the empirical relation between plant growth and leaf photosynthesis. Funct Plant Biol 33:421–429
Lambers H, Poorter H (1992) Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences
Larcher W (2003) Physiological plant ecology: ecophysiology and stress physiology of functional groups. Springer, Berlin
Liu F, De Cristofaro A, Violante A (2001) Effect of pH, phosphate and oxalate on the adsorption/desorption of arsenate on/from goethite. Soil Sci 166:197–208
Lott MS, Volin JC, Pemberton RW, Austin DF (2003) The reproductive biology of the invasive ferns Lygodium microphyllum and L. japonicum (Schizaeaceae): implications for invasive potential. Am J Bot 90:1144–1152. doi:10.3732/ajb.90.8.1144
Loveless CM (1959) A study of the vegetation in the Florida Everglades. Ecology 40:1–9
Marschner H (1995) Mineral nutrition of higher plants, 2nd edn. Academic Press, Boston
Mathur DD, Stamps RH, Conover CA (1982) Postharvest wilt and yellowing of leatherleaf fern. Proc Fla State Hort Soc 95:142–143
McConnaughay K, Coleman J (1999) Biomass allocation in plants: ontogeny or optimality? A test along three resource gradients. Ecology 80:2581–2593
McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA (1990) A new method which gives an objective measure of colonization of roots by vesicular—arbuscular mycorrhizal fungi. New Phytol 115(3):495–501
Nicol GW, Leininger S, Schleper C, Prosser JI (2008) The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ Microbiol 10(11):2966–2978
Nielsen SL, Enriquez S, Duarte C, Sand-Jensen K (1996) Scaling maximum growth rates across photosynthetic organisms. Funct Ecol 10:167–175
Pausas JG, Austin MP (2001) Patterns of plant species richness in relation to different environments: an appraisal. J Veg Sci 12:153–166
Poorter H, Bergkotte M (1992) Chemical composition of 24 wild species differing in relative growth rate. Plant, Cell Environ 15:221–229
Poorter H, Remkes C, Lambers H (1990) Carbon and nitrogen economy of 24 wild species differing in relative growth rate. Plant Physiol 94:621–627
Reich P, Walters M (1994) Photosynthesis-nitrogen relations in Amazonian tree species. II. Variation in nitrogen vis-a-vis specific leaf area influences mass- and area-based expressions. Oecologia 97:73–81
Reich P, Ellsworth D, Walters M (1998) Leaf structure (specific leaf area) modulates photosynthesis–nitrogen relations: evidence from within and across species and functional groups. Funct Ecol 12:948–958
Rorison I (1980) The effects of soil acidity on nutrient availability and plant response. In: Anonymous Effects of acid precipitation on terrestrial ecosystems. Springer, New York, pp 283–304
Ryan PR, Ditomaso JM, Kochian LV (1993) Aluminium toxicity in roots: an investigation of spatial sensitivity and the role of the root cap. J Exp Bot 44(2):437–446
Schindelbeck RR, Riha SJ (1988) Soil acidity, and the growth, biomass partitioning and leaf mineral composition of honeylocust (Gleditsia triacanthos L.) seedlings. Tree Physiol 4:361–369
Shen Q, Ran W, Cao Z (2003) Mechanisms of nitrite accumulation occurring in soil nitrification. Chemosphere 50:747–753
Shen J, Yuan L, Zhang J, Li H, Bai Z, Chen X, Zhang W, Zhang F (2011) Phosphorus dynamics: from soil to plant. Plant Physiol 156:997–1005. doi:10.1104/pp.111.175232
Silva S (2012) Aluminium toxicity targets in plants. J Bot. doi:10.1155/2012/219462
Slattery WJ, Conyers MK, Aitken RL (1999) Soil pH, aluminium, manganese and lime requirement. In: Peverill KI, Sparrow LA, Reuter DJ (eds) Soil analysis: an interpretation manual. CSIRO Publishing, Melbourne
Smith RV, Doyle RM, Burns LC, Stevens RJ (1997) A model for nitrite accumulation in soils. Soil Biol Biochem 29(8):1241–1247
Soti PG, Jayachandran K, Purcell M, Volin JC, Kitajima K (2014) Mycorrhizal symbiosis and Lygodium microphyllum invasion in south Florida—a biogeographic comparison. Symbiosis 62(2):81–90
Stephenson RJ, Rechcigl JE (1991) Effects of dolomite and gypsum on weeds. Commun Soil Sci Plant Anal 22:1569–1579
Storer DA (1984) A simple high sample volume ashing procedure for determination of soil organic matter. Commun Soil Sci Plant Anal 15:759–772
Tyler G (1999) Plant distribution and soil-plant interactions on shallow soils. Acta Phytogeogr Suec 84:21–32
U.S. Environmental Protection Agency. 1996. Method 3050B. Acid digestion of sediments, sludges, and soils. http://www.epa.gov/wastes/hazard/testmethods/sw846/online/3_series.htm. Accessed 2 Feb 2013
Van Loan AN (2006) Japanese climbing fern: the insidious “other” Lygodium. Wildland Weeds 9:25–27
Volin JC, Kruger EL, Lindroth RL (2002) Responses of deciduous broadleaf trees to defoliation in a CO2 enriched atmosphere. Tree Physiol 22:435–448
Volin JC, Lott MS, Muss JD, Owen D (2004) Predicting rapid invasion of the Florida Everglades by Old World climbing fern (Lygodium microphyllum). Divers Distrib 10:439–446
Volin JC, Kruger EL, Volin VC, Tobin MF, Kitajima K (2010) Does release from natural belowground enemies help explain the invasiveness of Lygodium microphyllum? A cross-continental comparison. Plant Ecol 208:223–234
Acknowledgments
We thank Steven Oberbauer, Department of Biological Sciences, Florida International University and Stewart T Reed, USDA, ARS, Miami, Florida for their help in analyzing the samples. This research was supported by the Dissertation Evidence Acquisition Fellowship, to Pushpa Soti from the Graduate School, Florida International University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Sarah M Emery.
Rights and permissions
About this article

Cite this article
Soti, P.G., Jayachandran, K., Koptur, S. et al. Effect of soil pH on growth, nutrient uptake, and mycorrhizal colonization in exotic invasive Lygodium microphyllum . Plant Ecol 216, 989–998 (2015). https://doi.org/10.1007/s11258-015-0484-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-015-0484-6