
Abstract
The combustion of fossil fuels and associated warmer temperatures are causing a global increase in the availability of soil nutrients such as nitrogen. This will have pronounced effects on plants at northern latitudes that are adapted to low nutrient conditions. An experiment in northern Canada set up in 1990 has investigated the effects of long-term nutrient enrichment (fertilizer addition) and mammalian herbivore exclusion (fencing) on an understory plant community. We used this experiment to assess how 22 years of fertilization has affected investment in sexual reproduction in four herbaceous understory species. We measured reproductive output at the plot level (proportion of plants flowering) for four species (Mertensia paniculata, Epilobium angustifolium, Achillea millefolium and Festuca altaica) and at the individual plant level (biomass allocation to flower parts) for M. paniculata and E. angustifolium. Fertilization increased the probability of flowering for A. millefolium and resulted in a higher allocation of biomass to flower parts for E. angustifolium. Sexual reproduction in M. paniculata and F. altaica was largely unaffected by increased nutrient supply, and, as expected, herbivore exclusion had almost no effect on any species. Whereas plants in northern ecosystems currently reproduce mainly through clonal growth, rapidly changing environmental conditions and warmer temperatures will likely result in increased benefits of sexual reproduction. This could give a competitive advantage to species such as A. millefolium and E. angustifolium that increase investment in sexual reproduction when released from nutrient limitation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Since the industrial revolution, anthropogenic activities have caused a dramatic increase in the amount of available, reactive nitrogen in the global nitrogen cycle (Galloway et al. 2008). The combustion of fossil fuels, combined with faster mineralization rates of key nutrients resulting from warmer temperatures, are causing higher levels of soil nitrogen worldwide (Rustad et al. 2001; Galloway et al. 2008). Northern ecosystems may be especially impacted by this input of nutrients because they are often severely nutrient-limited and are projected to experience the greatest warming as a result of climate change (Aerts et al. 2006; Pachauri and Reisinger 2007). Although many studies focus on the effects of soil nutrients at the community level, these may overlook some critical processes and responses at the individual level (e.g. physiological processes or mortality). These responses are often the first to be detected and can have far-reaching effects as drivers of change at the community level (e.g. species reordering) (Niu et al. 2008; Smith et al. 2009). Sexual reproduction in plants has received attention in recent years as one such process that can be sensitive to environmental changes and may have important consequences at the community level (Fu et al. 2010; Fremlin et al. 2011; Klady et al. 2011; Moulton and Gough 2011).
Soil nutrient availability has long been known to influence relative investment in sexual and vegetative reproduction in clonal species (Harper and Ogden 1970). However, attempts to model how changing soil nutrient levels affects this trade-off have produced conflicting predictions. One hypothesis is that plants should decrease sexual reproduction in favor of clonal growth when soil nutrients are abundant because clonal growth produces proximate and genetically identical offspring with a proven genotype that can take advantage of favorable local conditions (Sakai 1995; Fu et al. 2010). Conversely, plants may invest more resources into sexual reproduction when conditions improve because flower and seed production provide a fitness advantage but are costly to make, so when nutrients are abundant and plants can afford to produce them, they should do so (Loehle 1987; Gardner and Mangel 1999). Experimental evidence is similarly conflicting; some researchers have reported increased investment in sexual reproduction when plants are fertilized (Dormann and Woodin 2002; Kawano and Nagai 2005; Burkle and Irwin 2009) while others reported decreased investment (Niu et al. 2008; Bai et al. 2009; Huang et al. 2009).
It is not surprising that studies investigating the effects of nutrient addition on sexual reproduction have produced conflicting results, as results may depend on species identity, competition intensity and patchiness of resources (Loehle 1987; Sakai 1995; Gardner and Mangel 1999; Fu et al. 2010). In addition, the measured effects of soil nutrients on investment in sexual reproduction may also vary due to differences in methods used to assess reproductive effort. Accepted proxies for reproductive effort include probability of flowering (Bai et al. 2009; Moulton and Gough 2011), number of buds, flowers, seeds or reproductive tillers produced per plant (Burkle and Irwin 2009; Hautier et al. 2009) and biomass allocated to reproductive parts (Sugiyama and Bazzaz 2002; Huang et al. 2009). Although there is no consensus on the most ecologically relevant method for assessing investment in sexual reproduction, biomass allocation to reproductive parts has a long history of use since it was introduced to plant ecologists by Harper (1967). More recent advances have emphasized the importance of untangling the sometimes confounding affects caused by the inherent relationship between plant size and investment in reproductive parts that exists for many species (Samson and Werk 1986; Weiner 2004). As a result of this relationship, plants treated with fertilizer may alter allocation of biomass to reproductive parts simply as a result of being larger, rather than due to a true treatment effect (Samson and Werk 1986; Weiner 2004). By considering how treatments affect the relationship between plant size and investment in reproduction, rather than simply measuring how the ratios of reproductive to total biomass are altered, it is possible to distinguish changes in reproductive allocation resulting from changes in plant size versus actual treatment effects (Weiner 2004).
Although plant species in northern regions are mostly long-lived perennials that rely almost exclusively on clonal growth, future environmental change could, for several reasons, cause sexual reproduction to become more important for plants at high latitudes. As species’ ranges change, the genetic recombination and dispersal associated with sexual reproduction will likely give sexually reproductive plants an advantage in colonizing and adapting to new areas (Aerts et al. 2006; Steltzer et al. 2008; Hedhly et al. 2009). In addition, more soil nitrogen and higher temperatures may improve conditions in a previously harsh germination environment, further enhancing the role of sexual reproduction (Moulton and Gough 2011). Investment in sexual reproduction could thus become a more important determinant of species success in northern biomes in the future. Therefore, a better understanding of the effects of increasing soil nutrients on the reproduction of common species in this biome is essential for conservation and management planning.
This study uses a long-term fertilizer and herbivore exclusion experiment in northern Canada to investigate the effects of 22 years of nutrient enrichment on investment in sexual reproduction in four boreal forest understory species. During this experimental period, plots treated with fertilizer have undergone major changes in plant community composition, with plant species richness declining (Turkington et al. 2002; DeKoning 2011). However, almost no effects of herbivore exclusion have ever been detected in these plots, despite the presence of three peaks in densities of snowshoe hares (the major herbivore in the system) during the study period (Turkington et al. 2002; DeKoning 2011; Fremlin et al. 2011). For this reason, we focus primarily on the effects of nutrient addition, although all parameters were also analysed for fencing effects. Here, we use a snap-shot data set collected in the 22nd year of the experiment to address two questions:
-
1.
Has fertilization altered investment in reproduction for any of four focal species (Mertensia paniculata, Epilobium angustifolium, Achillea millefolium and Festuca altaica)?
-
2.
Are previous assessments of these species’ responses to the treatments consistent with current findings or do long-term trends differ from short-term responses?
Methods
Study site
The study site is in the boreal forest near Kluane Lake in southwestern Yukon Territory. Daily mean temperatures range from 4.7–18.0 °C during the June–August growing season and the area has an annual mean precipitation of 290 mm (Environment Canada 2012). The soil at the study site has low fertility, with levels of key nutrients in mineral soil falling within the ranges of: total N 0–5 ppm, P 3–32 ppm, K 53–824 ppm. The forest is moderately open (45–60 % canopy cover, 160–220 stems/ha) and is dominated by white spruce (Picea glauca (Moench) Voss). In addition to the understory species examined in this study, arctic lupine (Lupinus arcticus S. Wats.), twin-flower (Linnaea borealis L.) and bearberry (Arctostaphylos uva-ursi (L.) Spreng.) are also common. The snowshoe hare (Lepus americanus Erxleben) is the main herbivore in the system, and experiences an 8–12 year population cycle with densities of up to 200 hares/km2 in high years (Krebs et al. 2001). In 2011, when data for this study were collected, snowshoe hares were at a low point in their cycle with a density of 10 hares/km2 (Krebs et al. 2012).
Experimental design
The site has sixteen 5 × 2.5 m2 plots with fully crossed fertilizer and fencing (exclosure) treatments (+/0 fertilizer, +/0 fencing), for a total of four treatments. Fertilizer application increases soil nutrients and fences exclude snowshoe hares and other mammalian herbivores. Plots were placed in semi-open areas within the understory with no rooted trees, and treatments were randomly assigned to each plot for a total of four replicates of each treatment. Since 1990, fences have been maintained and granular fertilizer (N:P:K 35:10:5) has been applied to all fertilized plots each year immediately after snow melt, between mid-May and early June. Fertilizer is applied at a rate of 0.625 kg per 5 × 2.5 m2 plot per year, resulting in a total addition of 17.5 g N/m2/year, 5 g P/m2/year and 2.5 g K/m2/year, a rate consistent with some other fertilizer experiments in northern ecosystems (Kellner 1993; Gough et al. 2002).
Study species
Four common understory species were selected for this study: bluebell (Mertensia paniculata (Aiton) G. Don), fireweed (Epilobium angustifolium L. s.l.), yarrow (Achillea millefolium L. var. borealis (Bong) Farwell) and northern rough fescue (Festuca altaica Torr.). These species are all native herbaceous perennials. Achillea millefolium and M. paniculata have two distinct growth forms: rosettes (not sexually reproductive) consisting of several leaves and no stems, and erect (sexually reproductive). Epilobium angustifolium has only an erect form. Festuca altaica is a bunch grass and each bunch may or may not include a reproductive ramet. Mertensia paniculata, E. angustifolium and A. millefolium are all capable of reproducing asexually through the spread of rhizomes, whereas F. altaica is a densely-tufted bunchgrass that grows from fibrous roots. Due to the clonal nature of these species, an “individual” may include ramets of the same clone, as identification of individuals was not genetically-based.
Density of reproductive individuals
The number of reproductive stems of each species was counted in each plot during that species’ period of peak flowering (M. paniculata on June 26th, F. altaica on July 9th, A. millefolium on July 19th). Because fertilization advances the phenology of E. angustifolium by 7 days in these plots (Fremlin et al. 2011), E. angustifolium was counted in fertilized plots on August 1st and in unfertilized plots on August 8th. Stems were considered reproductive if any buds, flowers, fruits or seeds were visible. The percent cover of each species in all plots was measured using a pin-drop method between July 11th–13th 2011 for use as a covariate in the analysis of number of reproductive individuals.
Biomass allocation to reproductive parts
Due to an absence of reproductive individuals of F. altaica and A. millefolium in many plots, reproductive individuals could not be analyzed for these species. Five reproductive individuals of M. paniculata and E. angustifolium were randomly selected from each plot for the measurement of reproductive traits. To avoid edge effects, plants were not sampled within 20 cm of the edge of the plot. Plants were also excluded if they were within 15 cm of a previously selected individual of the same species to minimize the chances of selecting two individuals of the same clone. Sampling was conducted during the period of peak flowering for each species; Mertensia paniculata were harvested on June 26th, fertilized E. angustifolium on August 1st and unfertilized E. angustifolium on August 8th. All target plants were divided into vegetative tissues (leaves and stems) and reproductive parts, oven dried at 60 °C for 48 h and weighed. Reproductive parts included all petals, sepals, bracts, open or closed achenes (E. angustifolium) or nutlets (M. paniculata).
Number of buds, flower and fruits
All flowers and flower buds on each individual of M. paniculata and E. angustifolium harvested for biomass measurements were counted prior to harvest. On E. angustifolium, fruits, flowers and buds appeared simultaneously on the same plant and therefore fruits were also counted. Epilobium angustifolium was considered to be in fruit when the petals had fallen off a developed achene. Due to a lack of replicates of fruiting individuals in all plots, this experiment could not be used to measure the effect of fertilizer on seeds (weight, viability and germination) for any species. However an assessment of the effects of fertilizer on germination of E. angustifolium seeds that was made using individuals from outside of these plots did not detect any effects of fertilizer on germination for this species (Grainger and Turkington 2012).
Data analysis
Each species was analyzed independently for all analyses. Fertilizer and fencing effects on the number of reproductive individuals per plot were assessed using two ANCOVAs (one for fertilizer and one for fencing), with species abundance used as a covariate. Analyses of biomass allocation and number of buds, flowers and fruits were based on mean values for five individuals in each plot. Plots containing fewer than three reproductive individuals of a species were excluded analysis for that species. Fertilizer and fencing effects on biomass allocation to reproductive parts were assessed using two separate ANCOVAs (one for fertilizer and one for fencing) for each species, with total plant biomass used as a covariate. Count data (number of buds, fruits, flowers) were analyzed using a two-way ANOVA fitted with a generalized linear model, and a quasipoisson distribution was used because of overdispersion. Data were log or rank transformed to meet model assumptions. All data analyses were conducted using the R statistical package (R Development Core Team 2010).
Results
Density of reproductive individuals
There was no effect of fencing on the density of reproductive individuals for any species. Although fertilization increased percent cover of M. paniculata and E. angustifolium, which resulted in more reproductive individuals, when percent cover was considered as a covariate there was no effect of fertilizer on the number of reproductive individuals for either species (Table 1, Fig. 1). Likewise, there was no significant effect of fertilization on the number of reproductive individuals of F. altaica (Table 1, Fig. 1). Achillea millefolium was reproductive only in fertilized plots, therefore fertilization both altered the relationship between percent cover and number of reproductive individuals (created a positive relationship) and increased the number of reproductive individuals even with percent cover considered as a covariate (Table 1, Fig. 1).

The number of reproductive individuals per plot, regressed against percent cover of that species in unfertilized (open symbols, dashed lines) and fertilized plots (closed symbols, solid lines) for four boreal forest understory species. Line equations and R squared values are shown for unfertilized (U) and fertilized (F) plots. Each point represents a plot
Biomass allocation to reproductive parts
There was no effect of fencing on allocation to reproductive parts for either M. paniculata or E. angustifolium (Table 2). Fertilization caused an increase in proportional allocation to reproductive parts in E. angustifolium, but had no effect on M. paniculata (Table 2, Fig. 2).

The relationship between proportional allocation to reproductive parts (as a percentage of total biomass) and total plant biomass for M. paniculata and E. angustifolium in unfertilized (open symbols, dashed lines) and fertilized plots (closed symbols, solid lines). Reproductive biomass includes petals, sepals, bracts and nutlets (M. paniculata) or achenes (E. angustifolium). Line equations and R squared values are shown for unfertilized (U) and fertilized (F) plots. Each point represents a plot average taken from five plants
Number of buds, flowers and fruits
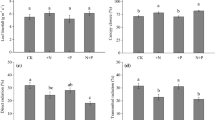
Fertilization resulted in more flower buds on M. paniculata, and more buds and flowers on E. angustifolium (Table 3, Fig. 3). Epilobium angustifolium in fenced plots also had more fruits.

Mean number (±1 SE) of flower buds, flowers and fruits per plant on M. paniculata and E. angustifolium in control (C), exclosure (fenced) (E), fertilized (F) and fertilized + exclosure (FE) plots. Five individuals of each species were sampled in each plot
Discussion
The four species in this study system vary widely in their ability to compete in high nutrient conditions, which has resulted in major changes in community composition in fertilized plots over the past 22 years (Grainger and Turkington 2013). Thus far, most of the observed increases in abundance in fertilized plots by those species that have become dominant have occurred by clonal growth. However, future conditions will likely increase the role of sexual reproduction in successful colonization of northern ecosystems, and differences in species’ investment in sexual reproduction could dictate which species will come to dominate this region (Steltzer et al. 2008; Hedhly et al. 2009). Both A. millefolium and E. angustifolium increased investment in sexual reproduction when fertilized, the former by producing more reproductive individuals and the latter by increasing biomass allocation to reproductive parts. Conversely, reproductive effort for M. paniculata and F. altaica was largely unaffected by the treatments. As previously observed, herbivore exclusion had almost no effect on the reproductive ecology of any of these species (John and Turkington 1995; Fremlin et al. 2011). This is not surprising, because snowshoe hares, which fences were designed to keep out, were at a low population density during 2011 when data for this experiment were collected and were likely not present in high enough numbers for effects of grazing to be detected (Krebs et al. 2012).
Reproductive investment at the plot level
Due to an increase in abundance of M. paniculata and E. angustifolium with fertilization, there were more reproductive individuals of these two species in fertilized plots. However, when abundance was taken into consideration in analysis, fertilizer had no effect on the number of reproductive individuals for either of these species, and the observed increase in number of reproductive individuals was simply a result of increased abundance. This contrasts with John and Turkington’s (1995) finding during a demographic study conducted in these plots between 1990 and 1994 that M. paniculata had a higher probability of flowering in fertilized plots. This could indicate that there has been a decline in reproductive effort for M. paniculata in the 17 years between the two studies.
Achillea millefolium was the only species that had more flowering individuals in fertilized plots even when abundance was taken into account. Achillea millefolium only flowered in fertilized plots and no flowering individuals were seen in the forest surrounding the plots. Nams et al. (1993) also reported an extremely low number of reproductive A. millefolium in unfertilized plots and an increased number of flowering individuals with fertilizer treatment. This species may have a minimum nutrient requirement for reproduction that is only attained in fertilized plots.
Allocation to reproduction at the individual plant level
Despite having more buds in fertilized plots, there was no effect of fertilization on allocation to reproduction for M. paniculata when plant size was taken into account. These results highlight the importance of considering the effect of plant size on allocation to sexual reproduction; if we had only counted the number of buds and flowers per plant, we could have erroneously concluded that allocation to reproduction had increased in M. paniculata. In contrast, fertilized E. angustifolium allocated more biomass to reproductive parts when fertilized, even when plant size was taken into account. Although plants in fertilized plots were indeed larger than their unfertilized counterparts, it is unlikely that the observed increase in allocation to reproductive tissue was apparent plasticity as a result of increased plant size, as there was no relationship between plant size and allocation to reproductive parts for this species. Rather, this increased allocation to reproductive parts was a direct fertilizer treatment effect. This increased allocation was also reflected in higher numbers of both buds and flowers on fertilized plants of this species. Of studies that took size into account when assessing nutrient effects on allocation to reproduction, Sugiyama and Bazzaz (2002) reported that nutrient addition resulted in increased allocation to reproduction in Abutilon theophrasti, whereas Huang et al. (2009) found decreased allocation to reproductive parts under high nutrient conditions in Corispermum macrocarpum. Response to fertilization appears to be species-specific, and increased investment in sexual reproduction of a widespread species such as E. angustifolium could have far-reaching consequences for vegetation dynamics in the boreal forest. Although germination rates for E. angustifolium were not affected by nutrient enrichment (Grainger and Turkington 2012), warmer temperatures could increase the probability of successful germination, giving this more sexually reproductive species a competitive in the future.
Limitations
Clonal growth by rhizomes is currently the main mode of reproduction for three of the four species we considered, and fertilizer addition can also affect this form of reproduction (Bai et al. 2009; Liu et al. 2009). Thus, an examination of the effects of nutrient enrichment on clonal growth would have been an ideal complement to this assessment of changes in sexual reproduction. However, due to the long-term nature of this experiment and the destructiveness of sampling the rhizomes of perennial species, exploring this avenue of investigation was not possible. Future assessments of reproductive responses to fertilizer treatment in northern biomes should attempt to measure the effects on clonal growth, wherever possible.
Competition is an important and possibly confounding variable in this study because neighbour density can affect a plant’s productivity, abundance and allocation to reproduction (Goldberg and Barton 1992; Treberg and Turkington 2010). Fertilization increased productivity, which led to increased competition from densely packed neighbours in fertilized plots. Turkington et al. (2002) reported an increase in F. altaica abundance in the first 10 years of this experiment, after which the species began to decline, which was likely a response to competition from tall neighbours (M. paniculata and E. angustifolium) that took several years to dominate the fertilized plots, rather than a direct effect of fertilizer (Turkington et al. 2002). Likewise, because nutrients were added at approximately the same time each year, interspecific differences in the timing of nutrient uptake could have affected competition for the added resources and determined how much nutrients were received by each species (Veresoglou and Fitter 1984). However, we found that investment to sexual reproduction was unaffected by isolating E. angustifolium from above-ground competition (data not reported). Other studies have also failed to detect any effect of density on allocation to reproduction (Zhang et al. 1996; Huang et al. 2009). Although the impact of competition intensity likely varies between fertilized and unfertilized plots as well as between species, the resulting effect on allocation to reproduction is likely not as substantial as the direct effects of treatment with fertilizer.
Conclusions
This study and comparisons with earlier results from the same area highlight the need for long-term experiments. Some earlier results, such as an increase in reproductive A. millefolium in fertilized plots and the lack of effects of herbivore exclusion, were corroborated in the current study and indicate long-term trends (Nams et al. 1993; Turkington et al. 2002). However other findings, such as the early increase in F. altaica abundance and M. paniculata probability of flowering in fertilized plots, were not confirmed, indicating shifting dynamics of this community during more than two decades of treatment (John and Turkington 1995; Turkington et al. 2002). Use of an analysis that takes plant size into consideration has further clarified the effects of increased nutrients on the reproductive ecology of these four species and highlighted subtleties that may previously have been overlooked.
If global nutrient enrichment and higher temperatures increase the benefits of sexual reproduction and the likelihood of successful germination in this northern ecosystem, E. angustifolium and A. millefolium could become increasingly dominant while M. paniculata and F. altaica could lose the competitive advantage that were initially reported from this study site. Continued monitoring of the response of such widespread and abundant species to environmental change is essential. By using long-term monitoring sites, developing and embracing new methods and incorporating past findings into our research, we improve our ability to predict vegetation responses to environmental change.
References
Aerts R, Cornelissen JHC, Dorrepaal E (2006) Plant performance in a warmer world: general responses of plants from cold, northern biomes and the importance of winter and spring events. Plant Ecol 182:63–77
Bai W, Sun X, Wang Z, Li L (2009) Nitrogen addition and rhizome severing modify clonal growth and reproductive modes of Leymus chinensis population. Plant Ecol 205:13–21
Burkle LA, Irwin RE (2009) The effects of nutrient addition on floral characters and pollination in two subalpine plants, Ipomopsis aggregata and Linum lewisii. Plant Ecol 203:83–98
Canada Environment (2012) Canadian climate normals 1971–2000 for burwash. National Climate Data and Information Archive, Yukon Territory
DeKoning PK (2011) Consequences and recovery after nutrient enrichment and herbivore reduction in the boreal forest understory. Dissertation, University of British Columbia
Dormann C, Woodin S (2002) Climate change in the arctic: using plant functional types in a meta-analysis of field experiments. Funct Ecol 16:4–17
Fremlin KM, McLaren JR, DeSandoli L, Turkington R (2011) The effects of fertilization and herbivory on the phenology of the understory vegetation of the boreal forest in Northwestern Canada. Arct Antarct Alp Res 43:389–396
Fu L, Wang S, Liu Z, Nijs I, Ma K, Li Z (2010) Effects of resource availability on the trade-off between seed and vegetative reproduction. J Plant Ecol 3:251–258
Galloway JN, Townsend AR, Erisman JW, Bekunda M, Cai Z, Freney JR, Martinelli LA, Seitzinger SP, Sutton MA (2008) Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320:889–892
Gardner SN, Mangel M (1999) Modeling investments in seeds, clonal offspring, and translocation in a clonal plant. Ecol 80:1202–1220
Goldberg DE, Barton AM (1992) Patterns and consequences of interspecific competition in natural communities: a review of field experiments with plants. Am Nat 139:771–801
Gough L, Wookey P, Shaver G (2002) Dry heath arctic tundra responses to long-term nutrient and light manipulation. Arct Antarct Alp Res 34(2):211–218
Grainger TN, Turkington R (2012) Germinability of Epilobium angustifoium seeds from plants treated annually with fertilizer for 22 years. Davidsonia 22:2–7
Grainger TN, Turkington R (2013) Mechanisms for success after long-term nutrient enrichment in a boreal forest understory. PLoS One 8:e61229
Harper J (1967) A Darwinian approach to plant ecology. J Anim Ecol 36:495–518
Harper JL, Ogden J (1970) The reproductive strategy of higher plants: I. The concept of strategy with special reference to Senecio vulgaris L. J Ecol 58:681–698
Hautier Y, Randin CF, Stöcklin J, Guisan A (2009) Changes in reproductive investment with altitude in an alpine plant. J Plant Ecol 2:125–134
Hedhly A, Hormaza JI, Herrero M (2009) Global warming and sexual plant reproduction. Trends Plant Sci 14:30–36
Huang YHY, Zhao XZX, Zhou DZD, Luo YLY, Mao WMW (2009) Allometry of Corispermum macrocarpum in response to soil nutrient, water, and population density. Botany 88:13–19
John E, Turkington R (1995) Herbaceous vegetation in the understorey of the boreal forest: does nutrient supply or snowshoe hare herbivory regulate species composition and abundance? J Ecol 83:581–590
Kawano S, Nagai Y (2005) Regulatory mechanisms of reproductive effort in plants 1. Plasticity in reproductive energy allocation and propagule output of Helianthus annuus L. (Compositae) cultivated at varying densities and nitrogen levels. Plant Species Biol 1:1–18
Kellner O (1993) Effects on associated flora of sylvicultural nitrogen fertilization repeated at long intervals. J Appl Ecol 30:563–574
Klady RA, Herny GHR, Lemay V (2011) Changes in high arctic tundra plant reproduction in response to long-term experimental warming. Glob Change Biol 17:1611–1624
Krebs CJ, Boonstra R, Boutin S, Sinclair ARE (2001) What drives the 10-year cycle of snowshoe hares? Bioscience 51:25–35
Krebs CJ, O’Donoghue M, Loewen V, Jung T, Gilbert S, Taylor S, Larocque L, Boonstra R, Boutin S and Kenney A (2012) The Community Ecological Monitoring Program Annual Report 2012
Liu F, Chen J-M, Wang Q-F (2009) Trade-offs between sexual and asexual reproduction in a monoecious species Sagittaria pygmaea (Alismataceae): the effect of different nutrient levels. Plant Syst Evol 277:61–65
Loehle C (1987) Partitioning of reproductive effort in clonal plants: a benefit-cost model. Oikos 49:199–208
Moulton CA, Gough L (2011) Effects of soil nutrient availability on the role of sexual reproduction in an Alaskan tundra plant community. Arct Antarct Alp Res 43:612–620
Nams VO, Folkard NFG, Smith JNM (1993) Effects of nitrogen fertilization on several woody and nonwoody boreal forest species. Can J Bot 71:93–97
Niu K, Luo Y, Choler P, Du G (2008) The role of biomass allocation strategy in diversity loss due to fertilization. Basic Appl Ecol 9:485–493
Pachauri RK and Reisinger A (2007) Climate change 2007: synthesis report: part of the working group III contribution to the fourth assessment report of the intergovernmental panel on climate change
R Development Core Team (2010) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rustad L, Campbell J, Marion G, Norby R, Mitchell M, Hartley A, Cornelissen J, Gurevitch J (2001) A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 126:543–562
Sakai S (1995) Optimal resource allocation to vegetative and sexual reproduction of a plant growing in a spatially varying environment. J Theor Biol 175:271–282
Samson DA, Werk KS (1986) Size-dependent effects in the analysis of reproductive effort in plants. Am Nat 127:667–680
Smith MD, Knapp AK, Collins SL (2009) A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change. Ecol 90:3279–3289
Steltzer H, Hufbauer RA, Welker JM, Casalis M, Sullivan PF, Chimner R (2008) Frequent sexual reproduction and high intraspecific variation in Salix arctica: Implications for a terrestrial feedback to climate change in the high arctic. J Geophys Res. doi:10.1029/2007JG000503
Sugiyama S, Bazzaz F (2002) Size dependence of reproductive allocation: the influence of resource availability, competition and genetic identity. Funct Ecol 12:280–288
Treberg MA, Turkington R (2010) Density dependence in an experimental boreal forest understory community. Botany 88:753–764
Turkington R, John E, Watson S, Seccombe-Hett P (2002) The effects of fertilization and herbivory on the herbaceous vegetation of the boreal forest in north-western Canada: a 10-year study. J Ecol 90:325–337
Veresoglou D, Fitter A (1984) Spatial and temporal patterns of growth and nutrient uptake of five co-existing grasses. J Ecol 72:259–272
Weiner J (2004) Allocation, plasticity and allometry in plants. Perspect Plant Ecol Evol Syst 6:207–215
Zhang D, Armitage A, Affolter J, Dirr M (1996) Environmental control of flowering and growth of Achillea millefolium L. summer pastels’. HortScience 31:364–365
Acknowledgments
Funding for this research was provided by the Natural Science and Engineering Research Council of Canada, the Association of Canadian Universities for Northern Studies and the Northern Scientific Training Program. We would also like to thank the staff at Kluane Lake Research Station, Douglas Curley, De Wet van Niekerk and Jennie McLaren for their invaluable assistance. We are grateful to the Kluane and Champagne-Aishihik First Nations for allowing us to do research on their traditional lands.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Grainger, T.N., Turkington, R. Long-term nutrient enrichment differentially affects investment in sexual reproduction in four boreal forest understory species. Plant Ecol 214, 1017–1026 (2013). https://doi.org/10.1007/s11258-013-0227-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-013-0227-5