
Abstract
We examined the effects of N addition and rhizome severing on sexual and clonal reproduction and their trade-off in a rhizome clonal grass, Leymus chinensis (Trin.) Tzvel. We discovered that N addition not only greatly increased the quantity and biomass of Leymus chinensis ramets, but also promoted ramet production by increasing the tillers of the plant, while abated the other alternative clonal propagation strategy of rhizome elongation. However, N addition did not affect the rhizome biomass and it significantly reduced the flowering probability, the individual seed mass, and the seed number. Rhizome severing did not markedly affect the quantity and the weight of ramets, the individual seed mass or the seed number, but greatly decreased the rhizome biomass. A significantly negative relation was found between sexual and clonal propagation in Leymus chinensis population upon N addition. We showed that the nutrient availability can modify the sexual versus clonal reproductive trade-off and the clonal propagation strategy. Intense ramet production characteristics of clonal growth will largely affect the sexual reproductive capacity and intensify intraclonal competition, thereby influencing their genetic diversity, spatial colonizing ability and life history strategy.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Clonal plant generally has two reproductive modes: sexual reproduction and clonal propagation (Koivunen et al. 2004). However, most clonal plants mainly rely on clonal propagation, and are pretty low in sexual reproduction and renewal (Eckert 2002). Some even abandon the sexual process (Philbrick and Les 1996). Thus, vigorous clonal growth may influence the evolution of plant life history in many aspects (Zhang and Zhang 2006). Sexual reproduction and clonal propagation are expected to compete for the same resources, i.e., there should be a trade-off between the two reproductive modes (Cook 1985; Sackville Hamilton et al. 1987; Eriksson 1997; Prati and Schmid 2000). Moreover, the cost of these two reproductive modes may be expressed in terms of future reproduction, growth or survival (Koivunen et al. 2004).
The relationship between sexual and clonal reproduction may be highly plastic in response to different environments (Sultan 2000; Fischer and van Kleunen 2002), leading to a variable ratio between both reproductive modes (Weppler and Stocklin 2005). Some researches reported that under growth-limiting conditions, clonal plants tend to allocate proportionally more biomass to clonal propagation and less to sexual reproduction (Abrahamson 1980; Cook 1985; Eriksson 1997). However, Sakai (1995) and Gardner and Mangel (1999) predicted that favorable habitats should promote clonal growth over sexual reproduction. The main cause of this contradiction is that the trade-off between sexual and asexual reproduction is not only affected by resource level (Sutherland and Vickery 1988; Piquot et al. 1998), but also influenced by plant size (Hartnett 1990; Cain and Damman 1997), ramet density (Humphrey and Pyke 1998; Van Kleunen et al. 2001), successional status (Cain and Damman 1997), and age (Piquot et al. 1998; López et al. 2001), which has been demonstrated in different species. Therefore, more research work is necessary to understand the controls on the trade-off between sexual and asexual reproduction.
Leymus chinensis, endemic to the east of Eurasian Continent, has long strong rhizomes and vigorous vegetative propagation, giving rise to extensively spreading clones and often forming monodominant stands. It possesses abundant ecological and genetic diversity (Wang 2004); thus, it is an ideal model plant for carrying out research into adaptive evolution and population ecology of clonal plants (Zhu 2004). For L. chinensis, rhizomes are channels of the ramet reproduction and for inter-ramet nutrient and water transport. Wang et al. (2004a) applied 14C isotope technique to study the photosynthate translocation among ramets, showing that 14C-photosynthate translocates not only between mother ramets and daughter ramets, but also between flowering ramets and vegetative ramets of the same generation. However, there are few researches on the effect on L. chinensis sexual and asexual reproduction and the trade-off relation between them after rhizome severing among ramets.
In the Inner-Mongolian arid and semi-arid grasslands, growth and reproduction of L. chinensis are often limited by N deficiency in the soil. Although many researchers demonstrated that N addition could markedly improve aboveground productivity of the plant (Zhu 2004), few reported the effect of N addition on sexual and asexual reproduction and their trade-off relation. Thus, the present experiment examined the responses of L. chinensis population to N supply and rhizome severing. In particular, we address the following questions: (1) whether or not sexual reproduction and clonal reproduction of L. chinensis are affected by N addition or by rhizome severing? (2) since sexual and clonal reproduction are thought to compete for resources (Thompson and Eckert 2004) and may be affected by N addition and rhizome severing, is there a trade-off relation between such two reproductive modes in L. chinensis population, and can such trade-off relation be modified by N addition or by rhizome severing? On the basis of this, the mechanisms of regeneration and adaptation to changing environments for L. chinensis will be further discussed.
Materials
Study site
The experiment was carried out in Duolun county of Xilingol League, Inner Mongolia (41°46′–42°36′N, 115°51′–116°54′E), which is on the southern edge of Hunshandake sandlands. The topography is dominated by low foothills within an elevation range of 1,150–1,800 m. The main soil type of the region is chestnut soil, accounting for 70% of the total area. Other soil types, such as aeolian sandy soil, meadow soil and chernozem, occur sparsely in some azonal habitats. This area is a typical middle temperate zone characterized by a semiarid continental monsoon climate, with a mean annual air temperature of 1.6°C and a frost-free period of about 100 days. The accumulated temperature of ≥10°C is 1,917.9°C, the mean temperature of the warmest month (July) is 18.7°C, while that of the coldest month (January) is −18.3°C. The mean annual precipitation is about 385.5 mm, while the mean potential evaporation is 1,748.0 mm. Vegetation of the region is dominated by typical steppes, and the dominant species are L. chinensis, Stipa spp., Agropyron cristatum, Artemisia frigida and Cleistogenes squarrosa, accompanied by secondary forests, thickets and sandy communities.
Study species
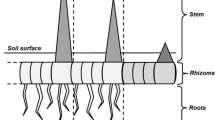
Leymus chinensis (Trin.) Tzvel., a perennial species of Gramineae, is widely distributed in the eastern region of the Eurasian steppe zone including the outer Baikal area of Russia, northern and eastern parts of the People’s Republic of Mongolia, the Northeast China Plain, the Northern China Plain, and the Inner Mongolian Plateau of China. In the middle and eastern parts of Asia, the total area of L. chinensis grasslands is about 420,000 km2, with about 220,000 km2 in China (Inner-Mongolia and Ning Xia Investigation and Survey Team of the Chinese Academy of Sciences 1985). Among all the plant species in the region, wide-spread L. chinensis is a most important forage grass, for it has greatest production and highest nutrient value (Zhu 2004). In the experimental area, L. chinensis begins turning green in late April, tillering in late June, flowering in mid-July, and seeding in late August. Tillering nodes of the plant are important clonal reproduction structures and can reproduce for four times at best within one growing season. Some of buds generated by tillering nodes grow upwards and form daughter ramets, while the others grow horizontally and form rhizomes (Yang et al. 1995). Rhizomes are also important clonal reproduction organs of L. chinensis and their diameters are usually 2–3 mm and their internode lengths are 2–6 cm. They distribute in the soil layer of 5–15 cm, living for about 4 years. Rhizome buds are either in dormancy, or grow upwards to form new ramets, or grow horizontally to form new rhizomes (Zhu 2004). We term every individual shoot plus its roots and rhizomes (rhizome fragments) as a ramet, and term ear-bearing shoots as sexually reproductive shoots.
Experimental treatments
In early September 2003, an experimental plot of 15 ha was set up and enclosed to avoid disturbance by livestock or humans. The topography of the plot was very flat and the vegetation was nearly pure and uniform L. chinensis grassland. Within the plot, 15 square experimental subplots with the size of 40 m2 were established. Subplots were chosen to be as similar as one another in vegetation and at least 1 m apart. Fifteen subplots were further randomly assigned to three treatments: Control, N addition (application rate is 32 N m−2 and the applied fertilizer is carbamide) and rhizome severing, giving five replicates for each treatment. In the rhizome severing treatment, we cut the soil into 15 cm deep to severe the rhizomes with a flat shovel (20 × 20 × 0.2 cm thick) along 39 lines which are 20 cm apart and parallel to the short sides of subplots, for rhizomes lie horizontally at about 10 cm under the ground surface (Wang et al. 2004b). In N addition treatment, N was applied at the rate of 32 g N m−2, which has been proved to be the optimal amount for biomass production by a long-term N addition experiment (Pan et al. 2005) conducted in Xilingol L. chinensis grassland. N addition and rhizome severing were carried out only once, on a single day of September 11, 2003.
Sampling and measurement
During the L. chinensis seed-mature period (August) in 2004 and 2005, we selected a 50 cm × 50 cm quadrat in each subplot to measure the number of ramets and of sexually reproductive shoots, then cut off all the ramets within each quadrat along the ground surface, and brought them into the lab, recorded the seed number. Finally all the vegetative shoots, all the seeds and vegetative parts of sexually reproductive shoots were dried in the oven (48 h, 70°C) and weighed. In the same quadrat, rhizome and root samples were taken with soil-sampling drill (30 cm in length, 12.5 cm in diameter). The collected samples were brought to laboratory and washed with tap water, and then rhizomes and roots were picked out. Roots were dipped into 2,3,5-phenyltetrazolium chloride (TTC) solution with the concentration of 0.5%, and placed in dark for 24 h (Wang et al. 1995). After that, we distinguished live roots and dead roots by color, discarded the dead roots. Finally, rhizomes and live roots were dried in the oven (48 h, 70°C) and weighed.
The ratio of number of sexually reproductive shoots to total ramet number in each subplot is used to reflect the flowering probability, and the ratio of total seed weight to total seed number is applied to calculate the individual seed mass. The flowering probability, seed number, and individual seed mass were used as estimates of sexual reproduction, while ramet number, ramet biomass, and rhizome biomass were taken as estimates of clonal propagation. The interrelation between the proportion of total biomass allocation to seeds and the proportion of total biomass allocation to ramets and rhizomes was used to reflect the trade-off between sexual and clonal reproduction. Total aboveground biomass was the total combination of ramet biomass and sexually reproductive shoot biomass, while the total belowground biomass incorporated the biomass of roots and rhizomes.
Data analysis
One-way analysis of variance (ANOVA) was used to analyze the effect of N addition or rhizome severing on the number of ramets, biomass of ramets, rhizome biomass, flowering probability, individual seed mass, seed number, and the number of ramets developed from rhizome nodes and from tiller bases. In all cases, significant levels of differences between means were determined by using a Bonferroni multiple-means comparison test at P < 0.05. Linear regression analysis (REG procedure) was performed to analyze the relation between the proportion of total biomass allocated to seeds and the proportion of total biomass allocation to ramets and rhizomes. All statistical tests were carried out with the Prism program (Graph Pad Software, San Diego).
Results
Clonal propagation
Both number and biomass of ramets were significantly affected by N addition, while no traits but rhizome biomass of 2005 was affected by rhizome severing (Fig. 1). During the two-year long experiment, the number of ramets in N addition treatment was significantly larger than those in control and rhizome severing (P < 0.01). The comparison showed no obvious difference in the number of ramets between the control and rhizome severing treatments (Fig. 1a). Likewise, biomass of ramets in N addition treatment (362.7 g m−2 in 2004 and 211.4 g m−2 in 2005 during the peak biomass period) was also significantly higher than that in either control or rhizome severing treatment (F = 33.25, P < 0.01), whereas biomass of ramets in rhizome severing treatment had no significant difference compared to that in control (Fig. 1b).

Comparisons of number (a) and biomass (b) of ramets and rhizomes biomass (c) of Leymus chinensis population among control, N addition (32 g N m−2) and rhizome severing treatments. Means (±Ι standard error) are shown. Letters above bars indicate significant differences at P < 0.05, based on Bonferroni’s multiple comparison test
In the first year, rhizome biomass in N addition and rhizome severing treatments was only a little lower than that in control. But in the second year, rhizome biomass in rhizome severing treatment was markedly lower than that in control (P < 0.05), while rhizome biomass in N addition treatment was not distinctly different from that in control (Fig. 1c).
The mean number of ramets produced by tiller bases in N addition treatment was 257 (No. m−2), markedly higher than those in control and rhizome severing treatment (Fig. 2), but there were no obvious difference between rhizome severing treatment and the control. However, the mean number of ramets produced by rhizome nodes in the control was 84 (No. m−2), obviously higher than those in N addition and rhizome severing treatments, while the ramet number in N addition treatment was not markedly different from that in rhizome severing treatment. The ramets developed from tiller bases accounted for 78.4% of all ramets in N addition treatment, on average 20.7% higher than that in control (P < 0.001) (Fig. 2).

Effects of N addition (32 g N m−2) and rhizome severing treatment on the number of ramets developed from rhizomes and from tiller bases of Leymus chinensis on August 19, 2005. Means with same letter are not significantly different (P < 0.05)
Sexual reproduction
The effects of N addition on the flowering probability, the individual seed mass and the seed number are almost the same. In N addition treatment, the flowering probability was sharply decreased by 34.5% (P < 0.05) and 29.5% (P < 0.05) in 2004 and 2005 (Fig. 3a), the mean seed mass was reduced by 27.54% and 30.45% (Fig. 3b), and the mean seed number was reduced by 25.2% (P < 0.05) and 23.8% (P < 0.05), respectively (Fig. 3c).

Comparisons of flowering probability (a), individual seed mass (b) and seeds number (c) of Leymus chinensis population among control, N addition (32 g N m−2) and rhizome severing treatments. Means (± Ι standard error) are shown. Letters above bars indicate significant differences at P < 0.05, based on Bonferroni’s multiple comparison test
In rhizome severing treatment, the flowering probability, the individual seed mass and the seed number were lower than those in control in the first year, and there was a great difference of the flowering probability (P < 0.05) between the two treatments. The flowering probability, the individual seed mass and the seed number were a little higher in rhizome severing treatment than those in control in the second year (Fig. 3), but without significant difference. The flowering probability and the number of seeds varied a lot between the 2 years due to the environmental fluctuation.
Relationship between clonal and sexual reproduction
There was a significant negative correlation between proportional biomass allocation to seeds and that to ramets and rhizomes in N addition treatment (P < 0.05), but not in control and rhizome severing treatment (Fig. 4).

Relationship between sexual and clonal reproduction in control, N addition (32 g N m−2) and rhizome severing treatments
Discussion
N addition modifies the clonal propagation strategy
Plants are expected to show a high degree of clonal expansion under good environmental conditions because they can arrive at a maximum through monopolizing the space and resources (Prati and Schmid 2000). Nutrient application was reported to be able to activate production of new rhizomes (see Hutchings and de Kroon 1994). Our results also demonstrated that N addition could significantly increase the number and the biomass of ramets in L. chinensis population, but did not affect the rhizome biomass (Fig. 1). Usually, rhizome clonal grass has two alternative ways to generate vegetative offsprings (clonal ramets): by developing from rhizome buds and tiller buds, which originates from rhizome nodes and older tiller bases, respectively. As a typical clonal plant, L. chinensis mainly relies on vegetative reproduction by rhizomes to expand clone and niche space under natural conditions (Yang et al. 1995). Furthermore, whatever architecture the L. chinensis clone has, it must occupy the growth space first, then increase the number of ramets in clonal patches (Zhu 2004). Yang et al. (1995) reported that L. chinensis clone occupied the niche space by elongating rhizomes which could grow further away from mother ramets to produce more daughter ramets, thereby realized the population recruitment. This implies an important clonal reproduction strategy: competition between daughter ramets is avoided to some extent and that between daughter and mother ramets reduced at the same time. Therefore, plants can utilize resources (nutrients and water) more effectively, occupy and expand the growing space more sufficiently (Cook 1985). However, our research has demonstrated that N addition can significantly increase the number of ramets of L. chinensis population (Fig. 1a), but preferentially by the development from the tiller-base-originated buds (Fig. 2). Noble et al. (1979) also found that nutrient addition increased the density of shoot populations of Carex arenaria. This suggested that the clonal reproduction strategy through rhizome elongating was altered by N addition. In relatively good habitats, the recruitment of L. chinensis population is usually realized by ramets developed from tiller bases, which can cut down the costs of ramet production because considerable materials and energy input into rhizomes could be saved. On the other hand, this may be considered to be another form of foraging behavior (Hutchings and de Kroon 1994), which enabled the plant to place their offsprings in most favorable habitats. While on the contrary, i.e., under unfavorable environmental conditions, plants may invoke plasticity that help escape from these adverse conditions by increasing the proportion of rhizome-originated buds, which are more effective to get the clone spread than those from tiller bases (Wang 2004). The underlying mechanism for such foraging behavior is probably the adjustment of these two alternative ways to generate offsprings (daughter ramets).
Rhizome severing affected clonal propagation
Wang et al. (2004a) reported that photosynthate could translocate not only between mother ramets and daughter ramets, but also between flowering ramets and vegetative ramets of the same generation. However, we did not discover any significant effect on the ramet quantity and the biomass of L. chinensis after rhizome severing. Similarly, Wang et al. (2004b) also reported that there was no clonal integration between ramets in the rhizome severing treatment at population level. In 2004, the rhizome biomass sharply decreased, probably caused by the death of some rhizomes after rhizome severing.
Sexual versus clonal reproduction trade-off
Clonal plants can facultatively switch between sexual and asexual reproduction in plastic response to the environment (Gardner and Mangel 1999). Although, Van Groenendael and Habekotte (1988) reported that under the resource limitation most clonal species showed a remarkable increase in the proportional allocation to clonal reproduction and a concomitant decrease in allocation to seed production (Harper 1977; Abrahamson 1980). But Loehle (1987) model predicted that more favorable conditions would decrease the cost of sexual reproduction, leading to the increase of seed output, while harsh conditions, by decreasing seedling establishment, would curtail seed production (Van Zandt et al. 2003). However, our data suggest that N addition significantly reduced the flowering probability, mass of single L. chinensis seed and the number of seeds in the population (Fig. 2). Sakai (1995) and Gardner and Mangel (1999) predicted that sexual reproduction would be minimized and clonal growth would be maximized in the most favorable habitats, which is in agreement with our findings of the experimental result in N addition treatment. The effect of rhizome severing on the flowering probability, mass of single L. chinensis seed and the number of seeds differed in the 2 years. Flowering probability, seed mass, and seed number were decreased in the first year while increased in the second year by rhizome severing. Nevertheless, except the great difference in flowering probability between rhizome severing and the control in the first year, no significant difference was examined in the mass of single seed and the seed number. What underlay such different responses between years is likely that sexual reproduction of L. chinensis is relatively sensitive to the fluctuation of temperature and rainfall (Zhu 2004).
Seedling recruitment has been considered to be rare in clonal plants (Eriksson 1989; 1992). Although Wang (1987) reported that L. chinensis has a long-life seed bank in the soil, we did not observe seedling recruitment in all the treated plots during the two-year experimental period. Yang and Zhang (1992) did not observe L. chinensis seedlings in the whole growing season in natural grassland in the Songnen Plain, probably due to the fact that L. chinensis is a photophilic plant with slender and short-lived seedlings, which dies soon after seeding. Thus, it is very difficult for seedlings to live in a L. chinensis dominated community (Yang and Zhang 1992; Yang et al. 1995). L. chinensis has been characterized by its low seed setting rate and germination capacity (Wang 1987). Despite that sexual reproduction still has its own significance. Firstly, the seed dispersal provides the possibility to further expand the population area. Secondly, seeds are able to tolerate more severe environments compared to ramets and seedlings. Even if the habitat is changed catastrophically, thanks to the seed bank, the population still has the opportunity to survive and flourish again.
The correlation between sexual and clonal reproduction may be highly plastic in response to different environments (Sultan 2000; Fischer and van Kleunen 2002). This has been corroborated by our findings that N addition could alter the relationship between the two reproductive modes, which are negatively correlated as declared in the present experiment and several others (Sutherland and Vickery 1988; Westley 1993; Muir 1995; Sun et al. 2001). For clonal plants, sexual versus clonal reproductive trade-off is of great significance in terms of ecology and evolution. In general, clonal plants rely more on clonal reproduction under favorable conditions (N addition treatment in the present study for an example) than under unfavorable ones, probably because the sexual reproduction is much more costly than clonal reproduction (López-Almansa et al. 2003). The favorable environment reduces the needs for progeny genetic diversity to be selected. Under unfavorable conditions, sexual reproduction will be enhanced or even outweigh the clonal reproduction, as to produce more genetic diversity to be chosen by the adverse or stressed environments. This is considered an important mechanism for clonal plants to adapt to the environment. However, vigorous clonal growth may have various effects on life history evolution. For example, some clonal plants exhibit very weak or even null sexual reproductive capacity because of overwhelming clonal growth (Philbrick and Les 1996), and consequently their genetic diversity, spatial colonizing ability, and life history strategy will be affected. In addition, ramet densification caused by clonal growth will intensify the intraclonal competition, which may affect the resource allocation between clonal growth and the sexual reproduction, and ultimately influences plant fitness components including genet size and distribution, reproductive value, size, and genetic structure of populations (Heywoods 1986). Therefore, to explore which genetic or ecological factors have effects on the sexual versus clonal reproductive tradeoff and to what extent they operate are still the challenging problems for plant ecologists.
References
Abrahamson WG (1980) Demography and vegetative reproduction. In: Solbrig OT (ed) Demography and evolution in plant populations. Blackwell, Oxford, UK, pp 89–106
Cain ML, Damman H (1997) Clonal growth and ramet performance in the woodland herb, Asarum canadense. J Ecol 85:883–897. doi:10.2307/2960609
Cook RE (1985) Growth and development in clonal plant populations. In: Jackson JBC, Buss LW, Cook RE (eds) Population biology and evolution of clonal organisms. Yale University Press, New Haven, pp 259–296
Eckert CG (2002) The loss of sex in clonal plants. Evol Ecol 15:501–520. doi:10.1023/A:1016005519651
Eriksson O (1989) Seedling dynamics and life histories in clonal plants. Oikos 55:231–238. doi:10.2307/3565427
Eriksson O (1992) Evolution of seed dispersal and recruitment in clonal plants. Oikos 63:439–448. doi:10.2307/3544970
Eriksson O (1997) Clonal life histories and the evolution of seed recruitment. In: de Kroon H, van Groenendael J (eds) The ecology and evolution of clonal plants. Backhuys, Leiden, pp 211–226
Fischer M, van Kleunen M (2002) On the evolution of clonal plant life histories. Evol Ecol 15:565–582. doi:10.1023/A:1016013721469
Gardner SN, Mangel M (1999) Modeling investments in seed, clonal offspring, and translocation in a clonal plant. Ecology 80:1202–1220
Harper JL (1977) Population biology of plants. Academic Press, San Diego, CA
Hartnett DC (1990) Size-dependent allocation to sexual and vegetative reproduction in four clonal composites. Oecologia 84:254–259
Heywoods JS (1986) The effect of plant size variation on genetic drift in population of annuals. Am Nat 127:851–861. doi:10.1086/284529
Humphrey LD, Pyke DA (1998) Demographic and growth responses of a guerrilla and a phalanx perennial grass in competitive mixtures. J Ecol 86:854–865. doi:10.1046/j.1365-2745.1998.8650854.x
Hutchings MJ, De Kroon H (1994) Foraging in plants: the role of morphological plasticity in resource acquisition. Adv Ecol Res 25:159–238. doi:10.1016/S0065-2504(08)60215-9
Inner-Mongolia, Ning Xia Investigation and Survey Team of the Chinese Academy of Sciences (1985) The vegetation of Inner Mongolia. Science Press, Beijing, pp 516–527
Koivunen S, Saikkonen K, Vuorisalo T, Mutikainen P (2004) Heavy metals modify costs of reproduction and clonal growth in the stoloniferous herb Potentilla anserine. Evol Ecol 18:541–561. doi:10.1007/s10682-004-5143-7
Loehle C (1987) Partitioning of reproductive effort in clonal plants: a benefit-cost model. Oikos 49:199–208. doi:10.2307/3566027
López F, Fungairino S, Heras P, Serrano J, Acosta F (2001) Age changes in the vegetative vs. reproductive allocation by module demographic strategies in a perennial plant. Plant Ecol 157:13–21. doi:10.1023/A:1014597832475
López-Almansa JC, Pannell JR, Gil L (2003) Female sterility in Ulmus minor (Ulmaceae): a hypothesis invoking the cost of sex in a clonal plant. Am J Bot 90:603–609. doi:10.3732/ajb.90.4.603
Muir AM (1995) The cost of reproduction to the clonal herb Asarum canadense (wild ginger). Can J Bot 73:1683–1686. doi:10.1139/b95-182
Noble JC, Bell AD, Harper JL (1979) The population biology of plants with clonal growth. I. The morphology and structural demography of Carex arenarla. J Ecol 67:983–1008. doi:10.2307/2259224
Pan QM, Bai YF, Han XG, Yang JC (2005) Effects on nitrogen additions on Leymus chinensis population in typical steppe of Inner Mongolia. Acta Phytoecol Sin 29:311–317
Philbrick CT, Les DH (1996) Evolution of aquatic angiosperm reproductive systems. Bioscience 46:813–826. doi:10.2307/1312967
Piquot Y, Petit D, Valero M, Cugen J, de Laguerie P, Vernet P (1998) Variation in asexual and sexual reproduction among young and old populations of the perennial macrophyte Sparganium erectum. Oikos 82:139–148. doi:10.2307/3546924
Prati D, Schmid B (2000) Genetic differentiation of life-history traits within populations of the clonal plant Ranunculus reptans. Oikos 90:442–456. doi:10.1034/j.1600-0706.2000.900303.x
Sackville Hamilton NR, Schmid B, Harper JL (1987) Life-history concepts and the population biology of clonal organisms. Proc R Soc Lond B Biol Sci 232:35–57
Sakai S (1995) Optimal resource allocation to vegetative and sexual reproduction of a plant growing in a spatially varying environment. J Theor Biol 175:271–282. doi:10.1006/jtbi.1995.0141
Sultan SE (2000) Phenotypic plasticity for plant development, function and life history. Trends Plant Sci 5:537–542. doi:10.1016/S1360-1385(00)01797-0
Sun S, Gao X, Cai Y (2001) Variations in sexual and asexual reproduction of Scirpus mariqueter along an elevational gradient. Ecol Res 16:263–274. doi:10.1046/j.1440-1703.2001.00395.x
Sutherland S, Vickery RKJ (1988) Trade-offs between sexual and asexual reproduction in the genus Mimulus. Oecologia 76:330–335
Thompson FL, Eckert CG (2004) Trade-offs between sexual and clonal reproduction in an aquatic plant: experimental manipulations vs. phenotypic correlations. J Evol Biol 17:581–592. doi:10.1111/j.1420-9101.2004.00701.x
Van Groenendael JM, Habekotte B (1988) Cyperus esculentus L. Biology, population dynamics, and possibilities to control this neophyte. Z PflKrankh PflSchutz Sonderh XI:61–69
Van Kleunen M, Fischer M, Schmid B (2001) Effects of intraspecific competition on size variation and reproductive allocation in a clonal plant. Oikos 94:515–524. doi:10.1034/j.1600-0706.2001.940313.x
Van Zandt PA, Tobler MA, Mouton E, Hasenstein KH, Mopper S (2003) Positive and negative consequences of salinity stress for the growth and reproduction of the clonal plant, Iris hexagona. J Ecol 91:837–846. doi:10.1046/j.1365-2745.2003.00806.x
Wang YS (1987) A preliminary studies on seed banks and propagule dispersal of Leymus chinensis population in forest steppe. Proceedings of international symposium on grassland in forest area, Harbin, China
Wang ZW (2004) Growing strategies of a rhizomatous clonal plant Leymus chinensis (Trin.) Tzvel. in response to defoliation at ramet population level. Postdoctoral thesis, Institute of Botany, Chinese Academy of Sciences, Beijing, pp 1–94
Wang Z, Burch WH, Mou P, Jones RH, Mitchell RJ (1995) Accuracy of visible and ultraviolet light for estimating live root proportions with minirhizotrons. Ecology 76:2330–2334. doi:10.2307/1941705
Wang YS, Hong RM, Huang DM, Teng XH, Li YS, Shiyomi M, Nakamura M (2004a) The translocation of photosynthate between clonal ramets of Leymus chinensis population. Acta Ecol Sin 24:900–907
Wang Z, Li L, Han X, Dong M (2004b) Do rhizome severing and shoot defoliation affect clonal growth of Leymus chinensis at ramet population level? Acta Oecol 26:255–260. doi:10.1016/j.actao.2004.08.007
Weppler T, Stocklin J (2005) Variation of sexual and clonal reproduction in the alpine Geum reptans in contrasting altitudes and successional stages. Basic Appl Ecol 6:305–306. doi:10.1016/j.baae.2005.03.002
Westley LC (1993) The effects of inflorescence bud removal on corm production in Helianthus cormosus, L. (Asocaceae). Ecology 74:2136–2144. doi:10.2307/1940858
Yang YF, Zhang BT (1992) An analysis of seasonal variation of vegetative propagation and the relationship between biomass and population density of an Aneurolepidium chinense in Songnen plain of China. Acta Bot Sin 34:443–449
Yang YF, Liu GC, Zhang BT (1995) An analysis of age structure and the strategy for asexual propagation of Aneurolepidium chinense population. Acta Bot Sin 37:147–153
Zhang YF, Zhang DY (2006) Asexual and sexual reproductive strategies in clonal plants. Acta Phytoecol Sin 30:174–183
Zhu TC (2004) Biological and ecological study of Leymus chinensis. Ji Lin Science and technology press, China, pp 90–154
Acknowledgments
We would like to thank S. H. Song and X. Li for their help in field and laboratory work. This research was supported by the State Key Basic Research Development Program of China (No.2007CB106800), the Knowledge Innovation Major Project of CAS (No.KZCX2-XB2-01), a grant from the National Natural Science Foundation of China (No.30521002 and No.30370268).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bai, W., Sun, X., Wang, Z. et al. Nitrogen addition and rhizome severing modify clonal growth and reproductive modes of Leymus chinensis population. Plant Ecol 205, 13–21 (2009). https://doi.org/10.1007/s11258-009-9595-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-009-9595-2