
Abstract
Background
Entomopathogenic nematodes (EPNs) are tiny parasitic worms that parasitize insects, in which they reproduce. Their foraging behavior has been subject to numerous studies, most of which have proposed that, at short distances, EPNs use chemicals that are emitted directly from the host as host location cues. Carbon dioxide (CO2) in particular has been implicated as an important cue. Recent evidence shows that at longer distances several EPNs take advantage of volatiles that are specifically emitted by roots in response to insect attack. Studies that have revealed these plant-mediated interactions among three trophic levels have been met with some disbelief.
Scope
This review aims to take away this skepticism by summarizing the evidence for a role of root volatiles as foraging cues for EPNs. To reinforce our argument, we conducted olfactometer assays in which we directly compared the attraction of an EPN species to CO2 and two typical inducible root volatiles.
Conclusions
The combination of the ubiquitous gas and a more specific root volatile was found to be considerably more attractive than one of the two alone. Hence, future studies on EPN foraging behavior should take into account that CO2 and plant volatiles may work in synergy as attractants for EPNs. Recent research efforts also reveal prospects of exploiting plant-produced signals to improve the biological control of insect pests in the rhizosphere.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Nematodes are tremendously diverse with close to thirty thousand described species, but estimates go as far as one to twenty million undescribed species (Hugot et al. 2001). Of the known species of these tiny worms, more than half have a parasitic life cycle, many of them plant parasitic (Mitreva et al. 2005). Among the nematodes that are specialized to parasitize insects, only two families (Steinernematidae and Heterorhabditidae) have evolved an active host seeking behavior and are known to carry pathogenic bacteria, which they release into the insect hemolymph leading to the rapid host death (Dillman et al. 2012). Of these so-called entomopathogenic nematodes (EPNs) the genera Steinernema and Heterorhabditis are the most commonly studied (Gaugler 2002). They are produced commercially and used as biological control agents against many soil insect pests (Grewal et al. 2005). The only stage of these obligate parasites that is free-living is the so-called infective juvenile of the third larval instar (Fig. 1). Infective juveniles have to find hosts in the physically and chemically complex soil matrix. When they succeed at this task, they will enter the newly found host, usually via natural openings such as mouth, anus or spiracles. Once inside they release their symbiotic bacteria, which multiply and produce toxins that kill the insect within days. Several generations of EPN develop, mate and lay eggs, while feeding on the bacteria inside the cadaver. When all resources are depleted a new generation of infective juveniles, carrying their symbionts, will burst out and will be faced with the challenge of finding new hosts. EPNs can differ considerably in their behavior and strategies to tackle this challenge (Campbell and Lewis 2002; Kaya and Gaugler 1993; Lewis et al. 2002).

Scanning electron microscopy photograph of an infective juvenile of Heterorhabditis bacteriaphora an entomophatogenic nematode that is commonly used as a biological control agent against various insect pests. The infective juvenile is the only stage that lives freely outside of hosts and relies on cues to locate potential insect hosts (photograph taken by Ivan Hiltpold, University of Neuchâtel)
EPNs are commonly divided in cruiser and ambushers (Lewis 2002). Cruisers actively move about in search of hosts, whereas ambushers stay in one place and wait for an arthropod host to pass by. Ambushers typically show nictating behavior, which means that they sit up straight on their tail, allowing them to readily attach and even swing themselves to hosts that may pass by (Lewis 2002). The behavioral distinction is not absolute and various EPN species may show both cruiser and ambusher behavior. In fact, the classification of EPN species in cruiser and ambusher types is being challenged by Wilson et al. (2012). They propose that many species will show different behaviors depending on the substrate in which they forage and they argue that these differences may depend on the detectability of volatile signals that the EPNs use as foraging cues (Wilson et al. 2012). In all cases, whether cruising or ambushing, chemical signals emitted from the hosts have been shown to be key close-range host detection cues.
One of the most important of such cues is CO2. This omnipresent gas is emitted by most living organisms, including soil dwelling insects, but also plant roots. CO2 has been shown to attract various EPNs (Gaugler et al. 1980; Hallem et al. 2011; Lewis et al. 1993; O’Halloran and Burnell 2003). Yet, the ubiquitousness of CO2 suggests that it is not a very reliable cue. It will be released from many sources, including non-hosts, and its release from a host can even be negatively correlated with host quality, as already infected hosts may release more CO2 than uninfected hosts (Ramos-Rodriguez et al. 2007). It seems therefore more likely that CO2 predominantly serves as a response activator that alerts EPNs to the general presence of living organisms and may enhance their responsiveness to other, more specific and more reliable cues, similar to how CO2 increases responsiveness of mosquitoes to host odors (Dekker et al. 2005; Turner et al. 2011). The strategy to respond to generic signals in combination with more specific signals may be a highly effective way of finding resources and can be expected to be employed by many organisms. For the purpose of this review we tested if this might indeed also be the case for EPNs (see later).
For EPNs, the most reliable cues would be the ones that are specifically emitted by potential host insects. Yet, the strong selective pressure on insects to not give away their presence and therefore not produce such signals, poses an interesting dilemma for natural enemies, which has been described by Vet and Dicke (1992) as the reliability-detectability dilemma. In their seminal paper these authors argue that parasitic wasps that search for hosts in which they lay their eggs might use cues from their hosts in their foraging efforts, but these cues will only be available in minute amounts and therefore hard to detect. On the other hand, volatiles emitted by plants that are under insect attack are released in very large amounts, but are less reliable indicators of the specific presence of hosts. Many parasitic wasps are capable of learning by association and link host presence to specific odors, thereby increasing the likelihood that they can distinguish reliable plant signals from unreliable ones. Although olfactory learning has been reported also for nematodes (Nuttley et al. 2002; Torayama et al. 2007), EPNs do not have the possibility to use associative learning to improve their foraging efficiency because they only have one shot at finding a host. Nevertheless, as for parasitic wasps, volatiles that are emitted from insect-damaged plant tissues may also serve as useful and relatively reliable cues for EPN to locate environments with potential hosts, especially if they can be used in combination with more specific close range cues.
The evidence for attraction to herbivore-induced root volatiles
Wang and Gaugler (1998) conducted one of the earliest studies to show attraction of EPNs to volatiles from roots. They found that infective juveniles of Steinernema glaseri and Heterorhabditis bacteriophora are attracted to whole and wounded grass roots, prompting the authors to suggest “that these nematodes may use cues from grass roots and root wounds for host habitat finding”.
Our own work on EPN foraging behavior was motivated by the work by van Tol et al. (2001). They studied the attraction of the EPN Heterorhabditis megidis to roots of a coniferous plant (Thuja occidentalis), which were found to be particularly attractive when they had been damaged by larvae of the weevil Otiorhynchus sulcatus. Similarly, Boff et al. (2001) found strong attraction of H. megidis to strawberry roots that had been damaged by the same weevil species. In this latter case there was no attraction to the larvae themselves, strongly suggesting a key role for induced root volatiles. A study by Cutler and Webster (2003) also found an effect of plant root exudates on the foraging efficiency of EPNs. They did not always find a positive effect, which may have been due to the fact that the roots were not subjected to insect damage.
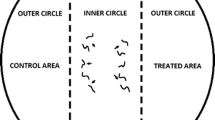
Another motivation for our investigations into the foraging behavior of EPNs is their potential as biological control agents against the western corn rootworm (WCR), Diabrotica virgifera virgifera, a major pest of maize roots that has been accidentally introduced several times into Europe, starting in the early 80’s (Szalai et al. 2011; Miller et al. 2005; Vidal et al. 2005). A better understanding of how EPNs locate host insects can help to develop strategies to augment their efficacy in controlling soil pests (see below). Having studied herbivore-induced volatiles as aboveground attractants for parasitic wasps for several decades (e.g. Tamo et al. 2006; Turlings et al. 2004, 1998, 1990), we logically suspected an active role of the plant in the attraction of EPNs to weevil-infested roots reported by van Tol et al. (2001) and Boff et al. (2001). The results from these studies not only implied EPN attraction to plant-produced signals, but also that these attractants act over considerably longer distances than commonly assumed. The two-choice PVC Y-tubes olfactometers that were used in their studies had a starting distance of about 20 cm between odor sources and nematodes. With this in mind, we developed an all-glass six-arm olfactometer (Fig. 2). The system allows for highly standardized testing of multiple odor sources at the same time and has additional advantage that it is easily cleaned and has no contaminants from unnatural materials. The only non-glass parts are the (also inert) Teflon connectors with each a very fine metal screen that keep the EPNs from entering the outer pots. For each test, the whole system is filled with moist sand or soil and the six outer pots receive potential odor sources, in most cases in our experiments these include plants. The EPNs, a few thousand at a time, are released in the central pot. One to 3 days after release the olfactometer is disconnected and the sand/soil in each of the glass connector arms is placed in a separate Bearman extractor funnel (Baermann 1917; Hass et al. 1999) in order to recuperate and count the number of nematodes that had ventured in each arm. These numbers are used as a measurement of the attractiveness of each odor that was tested.

Drawing of a belowground six-arm olfactometer as used to demonstrate the importance of inducible root volatiles for the attraction of EPNs. Drawing by Thomas Degen (www.thomas-degen.ch) modified from Rasmann et al. (2005)
The first olfactometer assay revealed that maize plants that had been subjected to root damage by WCR larvae were considerably more attractive to infective juveniles of Heterorhabditis megidis than undamaged maize plants or plants with mechanically damaged root (Rasmann et al. 2005). This result prompted us to investigate the volatiles emitted from maize roots. Using various volatile collection and analytical techniques it was found that the main difference between undamaged and WCR-damaged maize roots was the release of the sesquiterpene (E)-β-caryophyllene, which was only released by the latter plants (Rasmann et al. 2005). Using synthetic (E)-β-caryophyllene in an olfactometer assay showed that the compound is indeed highly attractive to H. megidis (Rasmann et al. 2005). Interestingly, American maize lines do not produce this compound, neither from the leaves (Degen et al. 2004), nor from their roots (Köllner et al. 2008). This offered a perfect opportunity to test the importance of (E)-β-caryophyllene for EPN attraction in vivo. A field test in which we compared a variety with and a variety without (E)-β-caryophyllene emission confirmed that the compound helps to guide EPNs towards WCR-infested maize roots under field conditions. The nematodes were able to locate hosts 50 cm away from the plants, confirming their long-distance foraging ability. The superior attraction by the emitting maize line resulted in a dramatic five-fold higher infection rate of WCR larvae on their roots than the non-emitting line (Rasmann et al. 2005). This first identification of a plant-produced EPN attractant led to several follow-up studies that revealed that roots of other plant species also release volatiles that attract EPN, but not all EPN species respond similarly (Hiltpold et al. 2010a; Rasmann and Turlings 2008).
Moving away from agronomic plants, Rasmann et al. (2011) showed that common milkweed Asclepias syriaca, when under root herbivore attack by Tetraopes tetraophthalmus (Coleoptera: Cerambycidae), also increases its belowground emission of volatile organic compounds. This too was associated with higher attraction of the EPN H. bacteriophora in laboratory experiments, but in this case the highly complex blend of organic compounds exuded from roots of milkweeds impedes the assessment of the exact attractant for EPNs. Subsequent field trials showed that the same nematode reduces survival of T. tetraophthalmus larvae and, as a result, restore aboveground plant biomass to control levels (i.e. as without herbivores) (Rasmann et al. 2011). This is one of the first demonstrations that a natural enemy of herbivores may contribute to positive selection on plants for signal production in response to insect feeding. Independently from the above studies on maize and milkweed, Ali et al. (2010, 2011) found that three species of EPNs are highly attracted to citrus roots damaged by larvae of the root weevil Diaprepes abbreviates. They found that roots of citrus plants emit a simple blend of terpenoids either constitutively or only after herbivore damage, in both cases correlating with nematode attraction toward the root volatiles (Ali et al. 2011). They found that the main compounds that are attractive to EPNs might also attract plant parasitic nematodes, which poses a significant cost to the plant. This could be one of the explanations why roots only emit the volatile when they are actually under insect attack. Interestingly, and in agreement with this notion, the only cultivar of citrus that Ali et al. (2011) found to release the nematode attractants constitutively, was also found to be resistant to phytopathogenic nematodes.
The most recent evidence that plant volatiles are attractive to EPN comes from a paper by Hallem et al. (2011), in which they tested a whole suite of volatile compounds, most of which known as plant volatiles, and found that infective juveniles of H. bacteriophora and S. carpocapsae are attracted to most of them. Combined, the results of these studies suggest that EPNs have a well-developed chemosensory apparatus, particularly sensitive to plant-derived compounds, which they readily exploit for successful host foraging (Rasmann et al. 2012).
The synergizing role of CO2
For the purpose of this review we decided to conduct a series of olfactometer assays in order to test the relative importance of CO2 versus herbivore-induced root volatiles for the attraction of EPN. We tested the attractiveness to the compounds individually, but also in combination to reveal possible additive or synergistic effects. Using H. megidis, we tested, in addition to CO2, two volatiles that are commonly emitted from insect-damaged roots, (E)-β-caryophyllene (Hiltpold et al. 2010c; Rasmann et al. 2005) and dimethyl disulfide (Ferry et al. 2007). As described above, 6-arm olfactometers (Fig. 2) were filled with moist sand (10 % water). Capillaries protruding from volatile dispensers (Hiltpold et al. 2010a) were inserted in the sand of three of the outer pots alternated with three sand-only pots. A first experiment included a pot that received a dispenser delivering (E)-β-caryophyllene, a pot that received a dispenser emitting CO2, a pot that received both such dispensers, and three control pots without dispensers. The second experiment was similar yet changing the (E)-β-caryophyllene dispenser with a one delivering dimethyl disulfide. Dispensers were prepared as described by D’Alessandro et al. (2006) by placing 100 mg of glass wool in a glass vial (1 ml Microlitre vial, Gerstel AG, Sursee, CH) on which we dispensed 300 μl of either synthetic (E)-β-caryophyllene or dimethyl disulfide (Sigma-Aldrich, Basel, CH). In parallel, 300 mg of CO2 generating material (sodium hydrogencarbonate and citric acid 3:1) was placed in the carbon dioxide dispensers and 50 μl of water was injected into the vial to trigger the release of CO2. All vials were closed with a septum lid and a 100 μl glass capillary (Hirschmann Laborgeräte GmbH, Eberstadt, DE) was inserted through the septum. The dispensers were then placed upside-down with the capillaries inserted in the sand of the assigned pots. Immediately after inserting the dispensers, 2000 H. megidis (Becker Underwood, UK) nematodes were released in the center of the device. The next day (24 h later), the olfactometers were disassembled and the nematodes were recovered from sand and counted as described by Rasmann et al. (2005). Both experiments were repeated six times for each synthetic compound and in both experiments we found differential nematode attraction ((E)-β-caryophyllene: ANOVA, F 3,33 = 10.58, P > 0.001, Fig. 3a; dimethyl disulfide: ANOVA, F 3,35 = 43.89, P > 0.001, Fig. 3b). H. megidis was significantly attracted toward CO2 as compared to empty pots, confirming that it is an attractant for this nematode. Moreover, both (E)-β-caryophyllene and dimethyl disulfide were as attractive to the nematodes as CO2. Interestingly, however, the nematodes were far more attracted to a combination of CO2 with either one of the two plant volatiles. To the best of our knowledge, this synergistic effect between CO2 and plant volatiles on EPN attraction was as yet unknown. These findings should not only settle the discussion on the relative importance of CO2 versus plant volatiles as attractants for EPNs, but may also encourage researchers to, from now on, consider CO2 as an response enhancer (Turner et al. 2011) that should be studied in combination with other attractants. It is important to note that besides typical wound-induced volatiles, plant roots secrete a great variety of other compounds of lesser volatility (Bertin et al. 2003), which may also play a role in short-range attraction, especially if they too are specifically emitted in response to herbivory.

Carbon dioxide and induced plant volatiles synergistically attract the EPN H. megidis. In a three choice situation including CO2 and (E)-β-caryophyllene (EβC) a) and CO2 and dimethyl disulfide (DMDS) b). In both cases, no differences were measured in the attractiveness of the single volatiles. However, a synergistic effect is observed when both CO2 and either (E)-β-caryophyllene or dimethyl disulfide were offered and significantly more nematodes chose these particular arms. Bars represent the standard error on the mean (SEM) and letters indicate statistical differences (p < 0.05)
Sesquiterpenes are signals with ideal diffusion properties
One other argument for the signaling function of herbivore-induced root volatiles is their exceptional diffusion rate in the soil matrix (Hiltpold and Turlings 2008). Naively, we initially were surprised to observe that a non-polar compound like (E)-β-caryophyllene attracted nematodes under subterranean conditions. Because it dissolves poorly in water we assumed that the sesquiterpene would also poorly travel in soil. The opposite turned out to be the case. By testing the diffusion properties of a series of common maize leaf volatiles of variable molecular weight and polarity, it was found that (E)-β-caryophyllene was one of the best diffusing compounds, in particular in sand with low humidity (Hiltpold and Turlings 2008). These results clearly imply that the compound travels in the gaseous phase and that water is more an obstacle rather than a facilitator of diffusion. The only tested compound that diffused even better than (E)-β-caryophyllene was β−copaene, another sesquiterpene that can also be observed in the volatile emissions of maize roots, but is more “expensive” to produce for the plant (Hiltpold and Turlings 2008).
Diffusion rates are obviously dependent on the substrate in which they are released and it can be expected that the better the diffusion the better the volatile signals serve EPN foraging in their efforts. Indeed, the success rates in terms of infectivity and persistence of EPNs can be highly dependent on soil type (Koppenhöfer and Fuzy 2006; Kruitbos et al. 2010). Besides factors such as humidity and pH, the soil texture and presence of organic matter will also influence the diffusion of host location signals. Indeed, the study conducted by Hiltpold and Turlings (2008) revealed that plant volatiles diffuse over considerably longer distances in sand than in soil. This is very nicely reflected in a study by Torr et al. (2004), who found that S. feltiae and H. megidis (but not S. carpocapsae) responded well to chemical cues in sand, but not in peat. Similarly, Kruitbos et al. (2010) also compared EPN foraging in peat and sand and for H. megidis found that it “dispersed well in both media, but only showed taxis towards hosts in sand”. These observations match well with the results from the diffusion experiments and imply that the effectiveness of the signals may be greatly impaired in soil types that contain high levels of chemical activity. In this respect, substrates with different types of sorption properties (e.g. polar versus non-polar) may be useful in the isolation and identification of other soil-borne attractants (D’Alessandro and Turlings 2005).
Exploiting herbivore-induced plant volatiles for better root pest control
As mentioned, one main reason for us to investigate the foraging behavior of EPN is their potential as biological control agents against WCR. The larvae of this beetle are the most destructive insect pest of maize in the USA, where they cost an estimated two billion US$ annually in damages and control measures (Mitchell 2011). Since its accidental introductions in Europe, WCR has rapidly spread and more recently has invaded various Western European countries (Miller et al. 2005), it is estimated that it will cost European farmers at least 0.5 million Euros annually (Wesseler and Fall 2010). EPNs have great potential as control agents against this and other root pests (Toepfer et al. 2009). A good understanding of the role of plant volatiles in plant protection in general may help to develop novel strategies to protect crops from insect pests (Degenhardt et al. 2003; Hiltpold and Turlings 2012; Pickett et al. 2006; Turlings and Ton 2006) and this may be particularly true for the effectiveness of biological control agents, such as EPNs. For aboveground signals, enhancement through genetic transformation has been found to be possible and it has been shown in laboratory and in greenhouse setups that such manipulation can improve the attractiveness of plants to predatory mites (Kappers et al. 2005) and parasitic wasps (Fontana et al. 2011; Schnee et al. 2006). The identification of the maize-produced EPN-attractant root signal (E)-β-caryophyllene offered another excellent opportunity to test the potential of such an approach.
The fact that American maize varieties have apparently lost the ability to release (E)-β-caryophyllene (Degen et al. 2004; Köllner et al. 2008; Rasmann et al. 2005) allowed us to validate the notion that plant signals can be used and possibly manipulated to improve the control of pest insects. The signal was restored in a non-producing line by transforming it with an (E)-β-caryophyllene synthase gene that had been identified from oregano, Origanum vulgare (Crocoll et al. 2010). The successful transformation resulted in the release of the sesquiterpene in equivalent amounts as a normally producing maize line, after WCR attack. However, in contrast to non-transformed, (E)-β-caryophyllene–emitting maize, the release was constitutive and occurred systemically throughout the transformed plants, from the roots as well as the leaves (Degenhardt et al. 2009). An extensive field experiment was conducted in which it was shown that transformed plants received considerably more EPN-mediated protection against WCR-inflicted root damage than non-transformed isogenic equivalents (Degenhardt et al. 2009). It should be noted that the constitutive release of the compound is of little use for realistic agronomic use, as the compound may also lure in pests and its continuous production may burdens the plant with a significant physiological cost. It is therefore envisioned that the approach can be improved by placing an inducible promoter in front of the (E)-β-caryophyllene-synthase gene.
Besides improving the signal itself, it is also possible to enhance the responsiveness of EPNs to the signals. The sheer numbers that EPNs produce each generation of new infective juveniles, as well as the short generation time of only a few weeks make them ideal subjects for selection for improved host finding and infectivity, as has been repeatedly validated (Gaugler and Campbell 1991; Gaugler et al. 1989; Griffin and Downes 1994). The feasibility of selecting for increased responsiveness to a plant-derived host finding cue was again demonstrated with the maize root volatile (E)-β-caryophyllene (Hiltpold et al. 2010a). Using the 6-arm olfactometer and the above-mentioned dispensers infective juveniles of H. bacteriophora were selected for increased responsiveness to (E)-β-caryophyllene (Hiltpold et al. 2010a). Unlike H. medidis, H. bacteriophora normally does not respond to this compound (Hiltpold et al. 2010c; Rasmann and Turlings 2008), but it took only six generations of selection to obtain a strain with a high degree of responsiveness (Hiltpold et al. 2010a). In a field trial in Hungary this strain was shown to be more effective at killing WCR larvae near a (E)-β-caryophyllene-emitting maize variety than the original strain, but no such difference between the strains was observed near a non-emitting line (Hiltpold et al. 2010a), whereas the persistence of both strains was equivalent (Hiltpold et al. 2010b).
The selection experiment also revealed that the responsiveness to (E)-β-caryophyllene is unlikely to be very specific, as the selected strain was also found to exhibit increased responsiveness to another sesquiterpene, (E)-β-farnesene, but less so to the monoterpene α-pinene (Hiltpold et al. 2010a). These data, combined with those from the diffusion study (Hiltpold and Turlings 2008), suggest that the attractiveness of plant volatiles is not specific, but rather depends on the facility with which a compound diffuses in soil.
With ways to improve signal strength and responsiveness to these signals, the only remaining obstacle towards effective control of root pests with EPNs is the cumbersome and labor-intensive field application process. EPNs can be readily reared in large numbers, but getting these numbers in good condition applied in the soil has rarely been a success (Shapiro-Ilan et al. 2006). In this same issue, Hiltpold et al. introduce a novel EPN application method that is based on EPNs-containing capsules. The precise formulation of the capsule shell allows for controlled escape of the EPNs. Theoretically sowing equipment can be adapted such that capsules can be “planted” during sowing in the field and therefore be applied with much less effort and less EPN loss than conventional methods. The use of capsules could be particularly effective if they contain attractants and stimulants that lure the pest towards the capsules and thereby towards their death.
Conclusions
The foraging behavior of entomopathogenic nematodes is tremendously diverse and can even vary considerably among different genotypes within one species. Increasing evidence suggests that root-produced signals can play a key role in luring EPNs into the rhizosphere of plants that are under attack by root herbivores. At closer distance to potential hosts, it is likely that other, host-derived chemical cues become more important. Field evidence obtained with maize plants that have lost the ability to emit an EPN attractant show the importance of such signals for successful host location by EPN. This has important implications for the use of EPN as biological control agents. Not only can their effectiveness be improved by enhancing the signals in crop varieties, high genetic variability in EPN responsiveness to signals also creates the possibility to rapidly select for enhanced responsiveness to specific signals. Combining these and other strategies that improve EPN virulence and release methods is very likely to lead to highly effective control of root pests.
References
Ali JG, Alborn HT, Stelinski LL (2010) Subterranean herbivore-induced volatiles released by citrus roots upon feeding by Diaprepes abbreviatus recruit entomopathogenic nematodes. J Chem Ecol 36(4):361–368
Ali JG, Alborn HT, Stelinski LL (2011) Constitutive and induced subterranean plant volatiles attract both entomopathogenic and plant parasitic nematodes. J Ecol 99(1):26–35
Baermann G (1917) Eine enifache Methode Zur Auffindung von Anklyostomum (Nematoden) larven in Erdproben. GeneeskTijdschrNed-Indie 57:131–137
Bertin C, Yang XH, Weston LA (2003) The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 256(1):67–83
Boff MIC, Zoon FC, Smits PH (2001) Orientation of Heterorhabditis megidis to insect hosts and plant roots in a Y-tube sand olfactometer. Entomologia Experimentalis et Applicata 98(3):329–337
Campbell JF, Lewis EE (2002) Entomopathogenic nematode host- search strategies. In: Lewis EE, Campbell JF, Sukhdeo MVK (eds) The behavioural ecology of parasites. CABI Publishing, Wallingford, pp 13–38
Crocoll C, Asbach J, Novak J, Gershenzon J, Degenhardt J (2010) Terpene synthases of oregano (Origanum vulgare L.) and their roles in the pathway and regulation of terpene biosynthesis. Plant Mol Biol 73:587–603
Cutler GC, Webster JM (2003) Host-finding ability of three entomopathogenic nematode isolates in the presence of plant roots. Nematology 5:601–608
D’Alessandro M, Turlings TCJ (2005) In Situ modification of herbivore-induced plant odors: a novel approach to study the attractiveness of volatile organic compounds to parasitic wasps. Chem Senses 30:739–753
D’Alessandro M, Held M, Triponez Y, Turlings TCJ (2006) The role of indole and other shikimic acid derived maize volatiles in the attraction of two parasitic wasps. J Chem Ecol 32(12):2733–2748
Degen T, Dillmann C, Marion-Poll F, Turlings TCJ (2004) High genetic variability of herbivore-induced volatile emission within a broad range of maize inbred lines. Plant Physiol 135(4):1928–1938
Degenhardt J, Gershenzon J, Baldwin IT, Kessler A (2003) Attracting friends to feast on foes: engineering terpene emission to make crop plants more attractive to herbivore enemies. Curr Opin Biotechnol 14(2):169–176
Degenhardt J, Hiltpold I, Köllner TG, Frey M, Gierl A, Gershenzon J, Hibbard BE, Ellersieck MR, Turlings TCJ (2009) Restoring a maize root signal that attracts insect-killing nematodes to control a major pest. Proc Natl Acad Sci U S A 106(32):13213–13218
Dekker T, Geier M, Carde RT (2005) Carbon dioxide instantly sensitizes female yellow fever mosquitoes to human skin odours. J Exp Biol 208(15):2963–2972
Dillman AR, Chaston JM, Adams BJ, Ciche TA, Goodrich-Blair H, Stock SP, Sternberg PW (2012) An entomopathogenic nematode by any other name. Plos Pathog 8(3):e1002527. doi:10.1371/journal.ppat.1002527
Ferry A, Dugravot S, Delattre T, Christides JP, Auger J, Bagneres AG, Poinsot D, Cortesero AM (2007) Identification of a widespread monomolecular odor differentially attractive to several Delia radicum ground-dwelling predators in the field. J Chem Ecol 33(11):2064–2077
Fontana A, Held M, Fantaye CA, Turlings TCJ, Degenhardt J, Gershenzon J (2011) Attractiveness of constitutive and herbivore-induced sesquiterpene blends of maize to the parasitic wasp Cotesia marginiventris (Cresson). J Chem Ecol 37(6):582–591
Gaugler R (2002) Entomopathogenic nematology. CABI Publishing, Wallingford
Gaugler R, Campbell JF (1991) Selection for enhanced host-finding of scarab larvae (Coleoptera, Scarabaeidae) in an entomopathogenic nematode. Environ Entomol 20(2):700–706
Gaugler R, Lebeck L, Nakagaki B, Boush GM (1980) Orientation of the entomogenous nematode Neoaplectana carpocapsae to carbon-dioxide. Environ Entomol 9(5):649–652
Gaugler R, Campbell JF, McGuire TR (1989) Selection for host-finding in Steinernema feltiae. J Invertebr Pathol 54(3):363–372
Grewal PS, Ehlers RU, Shapiro DI (2005) Nematodes as biocontrol agents. CABI Publishing, Wallingford
Griffin CT, Downes MJ (1994) Selection of Heterorhabditis sp. for improved infectivity at low temperatures. In: Burnell AM, Ehlers RU, Masson JP (eds) Genetics of entomopathogenic nematode-bacterium complexes. European Commission Publication EUR 15681 EN, Luxembourg, pp 120–128
Hallem EA, Dillman AR, Hong AV, Zhang YJ, Yano JM, DeMarco SF, Sternberg PW (2011) A sensory code for host seeking in parasitic nematodes. Curr Biol 21(5):377–383
Hass B, Griffin CT, Downes MJ (1999) Persistence of Heterorhabditis infective juveniles in soil: comparison of extraction and infectivity measurements. J Nematol 31(4):508–516
Hiltpold I, Turlings TCJ (2008) Belowground chemical signalling in maize: when simplicity rhymes with efficiency. J Chem Ecol 34(5):628–635
Hiltpold I, Turlings TCJ (2012) Manipulation of chemically mediated interactions in agricultural soils to enhance the control of crop pest and to improve crop yield. J Chem Ecol. doi:10.1007/s10886-012-0131-9
Hiltpold I, Baroni M, Toepfer S, Kuhlmann U, Turlings TCJ (2010a) Selection of entomopathogenic nematodes for enhanced responsiveness to a volatile root signal helps to control a major root pest. J Exp Biol 213(14):2417–2423
Hiltpold I, Baroni M, Toepfer S, Kuhlmann U, Turlings TCJ (2010b) Selective breeding of entomopathogenic nematodes for enhanced attraction to a root signal did not reduce their establishment or persistence after field release. Plant Signal Behav 5(11):1450–1452
Hiltpold I, Toepfer S, Kuhlmann U, Turlings TCJ (2010c) How maize root volatiles influence the efficacy of entomopathogenic nematodes against the western corn rootworm? Chemoecology 20(2):155–162
Hugot JP, Baujard P, Morand S (2001) Biodiversity in helminths and nematodes as a field of study: an overview. Nematology 3:199–208
Kappers IF, Aharoni A, van Herpen T, Luckerhoff LLP, Dicke M, Bouwmeester HJ (2005) Genetic engineering of terpenoid metabolism attracts bodyguards to Arabidopsis. Science 309(5743):2070–2072
Kaya HK, Gaugler R (1993) Entomopathogenic nematodes. Annu Rev Entomol 38:181–206
Köllner T, Held M, Lenk C, Hiltpold I, Turlings TCJ, Gershenzon J, Degenhardt J (2008) A maize (E)-BETA-caryophyllene synthase implicated in indirect defense responses against herbivores is not expressed in most American maize varieties. Plant Cell 20(2):482–494
Koppenhöfer AM, Fuzy EM (2006) Nematodes for white grub control: effects of soil type and soil moisture on infectivity and persistence. USGA Turfgrass Environ Res Online 5(18):1–10
Kruitbos LM, Heritage S, Hapca S, Wilson MJ (2010) The influence of habitat quality on the foraging strategies of the entomopathogenic nematodes Steinernema carpocapsae and Heterorhabditis megidis. Parasitology 137(2):303–309
Lewis EE (2002) Behavioural ecology. In: Gaugler R (ed) Entomopathogenic nematology. CABI International, pp 205–223
Lewis EE, Gaugler R, Harrison R (1993) Response of cruiser and ambusher entomopathogenic nematodes (Steinernematidae) to host volatile cues. Can J Zool-Rev Canadienne De Zool 71(4):765–769
Lewis EE, Barbarosa B, Gaugler R (2002) Mating and sexual communication by Steinernema carpocapsae (Nemata: Steinernematidae). J Nematol 34(4):328–331
Miller N, Estoup A, Toepfer S, Bourguet D, Lapchin L, Derridj S, Kim KS, Reynaud P, Furlan L, Guillemaud T (2005) Multiple transatlantic introductions of the western corn rootworm. Science 310(5750):992–992
Mitchell P (2011) Costs and benefits of controllin pest Diabrotica in maize in the United States. Paper presented at the 24th IWG Conference, Freiburg, Germany, 24–26 Oct. 2011
Mitreva M, Blaxter ML, Bird DM, McCarter JP (2005) Comparative genomics of nematodes. Trends Genet 21(10):573–581
Nuttley WM, Atkinson-Leadbeater KP, van der Kooy D (2002) Serotonin mediates food-odor associative learning in the nematode Caenorhabditis elegans. Proc Natl Acad Sci U S A 99(19):12449–12454
O’Halloran DM, Burnell AM (2003) An investigation of chemotaxis in the insect parasitic nematode Heterorhabditis bacteriophora. Parasitology 127:375–385
Pickett JA, Bruce TJA, Chamberlain K, Hassanali A, Khan ZR, Matthes MC, Napier JA, Smart LE, Wadhams LJ, Woodcock CM (2006) Plant volatiles yielding new ways to exploit plant defence. In: Dicke M, Takken W (eds) Chemical ecology: from gene to ecosystem. Springer, Dordrecht
Ramos-Rodriguez O, Campbell JF, Christen JM, Shapiro-Ilan DI, Lewis EE, Ramaswamy SB (2007) Attraction behaviour of three entomopathogenic nematode species towards infected and uninfected hosts. Parasitology 134:729–738
Rasmann S, Turlings TCJ (2008) First insights into specificity of below ground tritrophic interactions. Oikos 117:362–369
Rasmann S, Köllner TG, Degenhardt J, Hiltpold I, Toepfer S, Kuhlmann U, Gershenzon J, Turlings TCJ (2005) Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 434(7034):732–737
Rasmann S, Erwin AC, Halitschke R, Agrawal AA (2011) Direct and indirect root defences of milkweed (Asclepias syriaca): trophic cascades, trade-offs and novel methods for studying subterranean herbivory. J Ecol 99(1):16–25
Rasmann S, Ali J, Helder J, van der Putten W (2012) Ecology and evolution of soil nematode chemotaxis. J Chem Ecol. doi:10.1007/s10886-012-0118-6
Schnee C, Köllner TG, Held M, Turlings TCJ, Gershenzon J, Degenhardt J (2006) The products of a single maize sesquiterpene synthase form a volatile defense signal that attracts natural enemies of maize herbivores. Proc Natl Acad Sci USA 103(4):1129–1134
Shapiro-Ilan DI, Gouge DH, Piggott SJ, Fife JP (2006) Application technology and environmental considerations for use of entomopathogenic nematodes in biological control. Biol Control 38(1):124–133
Szalai M, Komáromi JP, Bažok R, Barčic JI, Kiss J, Toepfer S (2011) Generational growth rate estimates of Diabrotica virgifera virgifera populations (Coleoptera: Chrysomelidae). J Pest Sci 84(1):133–142
Tamo C, Ricard I, Held M, Davison AC, Turlings TCJ (2006) A comparison of naive and conditioned responses of three generalist endoparasitoids of lepidopteran larvae to host-induced plant odours. Anim Biol 56(2):205–220
Toepfer S, Haye T, Erlandson M, Goettel M, Lundgren JG, Kleespies RG, Weber DC, Walsh GC, Peters A, Ehlers RU, Strasser H, Moore D, Keller S, Vidal S, Kuhlmann U (2009) A review of the natural enemies of beetles in the subtribe Diabroticina (Coleoptera: Chrysomelidae): implications for sustainable pest management. Biocontrol Sci Technol 19:1–65
Torayama I, Ishihara T, Katsura I (2007) Caenorhabditis elegans integrates the signals of butanone and food to enhance chemotaxis to butanone. J Neurosci 27(4):741–750
Torr P, Heritage S, Wilson MJ (2004) Vibrations as a novel signal for host location by parasitic nematodes. Int J Parasitol 34(9):997–999
Turlings TCJ, Ton J (2006) Exploiting scents of distress: the prospect of manipulating herbivore-induced plant odours to enhance the control of agricultural pests. Curr Opinion Plant Biol 9(4):421–427
Turlings TCJ, Tumlinson JH, Lewis WJ (1990) Exploitation of herbivore-induced plant odors by host-seeking parasitic wasps. Science 250(4985):1251–1253
Turlings TCJ, Lengwiler UB, Bernasconi ML, Wechsler D (1998) Timing of induced volatile emissions in maize seedlings. Planta 207(1):146–152
Turlings TCJ, Davison AC, Tamò C (2004) A six-arm olfactometer permitting simultaneous observation of insect attraction and odour trapping. Physiol Entomol 29:45–55
Turner SL, Li N, Guda T, Githure J, Cardé RT, Ray A (2011) Ultra-prolonged activation of CO2-sensing neurons disorients mosquitoes. Nature 474(7349):87–91
van Tol RWHM, van der Sommen ATC, Boff MIC, van Bezooijen J, Sabelis MW, Smits PH (2001) Plants protect their roots by alerting the enemies of grubs. Ecol Lett 4(4):292–294
Vet LEM, Dicke M (1992) Ecology of infochemical use by natural enemies in a tritrophic context. Annu Rev Entomol 37:141–172
Vidal S, Kuhlmann U, Edwards R (2005) Western corn rootworm: ecology and management. CABI Publishers, Wallingford
Wang Y, Gaugler R (1998) Host and penetration site location by entomopathogenic nematodes against Japanese beetle larvae. J Invertebr Pathol 72(3):313–318
Wesseler J, Fall EH (2010) Potential damage costs of Diabrotica virgifera virgifera infestation in Europe - the ‘no control’ scenario. J Appl Entomol 134(5):385–394
Wilson MJ, Ehlers RU, Glazer I (2012) Entomopathogenic nematode foraging strategies – is Steinernema carpocapsae really an ambush forager? Nematology in press. doi:10.1163/156854111X617428
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Philippe Hinsinger.
Rights and permissions
About this article
Cite this article
Turlings, T.C.J., Hiltpold, I. & Rasmann, S. The importance of root-produced volatiles as foraging cues for entomopathogenic nematodes. Plant Soil 358, 51–60 (2012). https://doi.org/10.1007/s11104-012-1295-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-012-1295-3