
Abstract
Variation in plant N resorption may change with stand development because plants tend to adjust their ecophysiological traits with aging. In addition, changes in soil nitrogen (N) pools associated with stand development may also affect plant N resorption. Here, we examined green- and senesced-leaf N concentrations and resorption of trembling aspen ( Populus tremuloides Michx.) in boreal forest stands of different ages (7, 25, 85 and 139 years, respectively). All sampled stands originated from wildfires and established on similar parent materials (glacial tills) and had similar climates. N concentrations in both green and senesced leaves increased between 27% and 54% along the stand age chronosequence. Resorption efficiency (percentage difference of N between green and senesced leaves) and proficiency (N concentration in senesced leaves) were higher for leaves in younger stands than in older stands. An analysis of covariance indicated that the patterns of leaf N concentration and resorption were affected significantly by stand age, but not by available soil N concentration. Our results indicate that at an intra-specific level, plants could adjust their N resorption efficiency and proficiency with stand development.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In boreal forests, stand-replacing wildfire is widespread and is considered as the principal natural disturbance factor for secondary succession (Nepstad et al. 1999; Lindenmayer et al. 2004; Dellasala et al. 2006; Bond-Lamberty et al. 2007). Previous studies showed that production, biodiversity, carbon storage, soil nutrients, and other processes of ecosystems change with stand age, or time since stand-replacing fire (Wardle et al. 1997; Reich et al. 2001; Deluca et al. 2008; Hart and Chen 2008). There is a large literature on the effects of time since fire on soil characteristics (see review by Ice et al. 2004). For example, several studies have documented that soil nitrogen (N) availability increases (Brais et al. 1995; Simard et al. 2001; Bond-Lamberty et al. 2006; Leduc and Rothstein 2007; Duran et al. 2008) or decreases (Van Cleve et al. 1983; Deluca et al. 2002) along secondary successional gradients in boreal forests.
Although still in debate, nutrient resorption from senescing leaves, an important mechanism of nutrient conservation, is often thought to change with soil nutrient availability (Kobe et al. 2005; Richardson et al. 2005; Yuan et al. 2005b; Huang et al. 2008; Yuan and Chen 2009). Because soil nutrient availability changes with stand development since stand-replacing fire, and many ecophysiological plant traits have been linked to plant aging (Callaghan 1980; Groom et al. 1997), plant nutrient resorption may also change with stand age. However, no study, to our knowledge, has addressed the effects of stand age on plant nutrient resorption.
In North American boreal forests, widely distributed trembling aspen (Populus tremuloides Michx.) is an ecologically and economically important tree species. This species regenerates profusely after fire and grows to form stands with different ages (Brassard and Chen 2008; Hart and Chen 2008). It remains part of the forest cover, although in diminishing importance, for more than 150 years while the late successional coniferous species, white spruce (Picea glauca Moench.), balsam fir (Abies balsamea Mill.), and white cedar (Thuja occidentalis Linn.) gradually attain stand dominance (Bergeron and Dubuc 1989; Chen and Popadiouk 2002). To date, many aspects of the nutrient ecology of P. tremuloides have been studied (e.g. Ruark and Bockheim 1988; Killingbeck et al. 1990; Madritch et al. 2006), but there are still many voids in our understanding, particularly in the areas of resorption variation for this species with stand development.
The aim of this study was to examine variability of N resorption efficiency and proficiency in P. tremuloides during stand development covering a time span of 140 years. Because plant ecophysological traits are associated with aging, we hypothesized that N resorption efficiency of P. tremuloides changes with stand development no matter plant N resorption efficiency is inversely related to soil N availability (Kobe et al. 2005; Yuan et al. 2005a; Yuan et al. 2007) or remains unchanged (Aerts 1996).
Materials and methods
Study sites
Coinciding in part with those examined by Brassard et al. (2008), and Hart and Chen (2008), our study area was located in a boreal mixed-wood forest region, ≈150 km north of Thunder Bay, Ontario, between 49°38′ N and 49°27′ N and from 89°54′ W to 89°29′ W. The climate is moderately dry, cool with mean annual temperature and mean annual precipitation being 2.6°C and 710 mm, respectively. The study area is dominated by P. tremuloides, jack pine (Pinus banksiana Lamb.), black spruce (Picea mariana Mill.), and white birch (Betula papyrifera Marsh.).
We selected four stand-age classes, each with three replications for this nutrient resorption study. All selected stands regenerated naturally after wildfire. Every effort was made to intersperse sampled stands to avoid sampling stands of the same age in close proximity to one another to minimize the impact of spatial structure. For each stand, time since last stand-replacing fire was estimated using tree rings. During field sampling in 2006, the stands were aged 7, 25, 72, and 139 years old since fire, respectively (hereafter referred to as 7-, 25-, 72-, and 139-year-old stands) (Table 1). Other than stand age differences, all soils on the selected sites were classified as the Brunisolic order on relatively deep glacial tills parent material, and located in the same vegetation zone as those studied by Hart and Chen (2008).
Plant sampling
In each stand, a plot of ≈0.2 ha, uniform in stand composition, structure, understory vegetation, and soil characteristics, was established, within which all leaf samples were taken. Within the plot, we sampled 5 healthy, well-spaced dominant trees of P. tremuloides. Both green and senesced leaves were sampled from the same individuals. Because foliar N and P concentrations of P. tremuloides are relatively stable from the time of full leaf expansion to the onset of senescence (Alban 1985), we sampled fully green leaves in August 2006. Green leaves were collected from upper, middle, and lower crown positions. Branches with green leaves were sampled with the aid of a shotgun. At least 25 leaves were collected from each sample tree.
Recently fallen senesced leaves were collected in October 2006 when leaves fully senesced. Also, at least 25 leaves were sampled from each of these selected trees, and were combined into one sample per stand.
Leaf samples were placed in damp plastic bags and brought back to the laboratory. Leaf area was determined within 24 h using a leaf area meter (LI-3100; Li-Cor, USA). The leaf samples were then oven-dried at 60°C to constant mass, weighed to determine leaf mass per unit area (LMA), and ground in a Wiley mill (No. 40 sieve). Green- and senesced-leaf samples were digested using a persulfate–peroxide procedure in a block digestor and analyzed colorimetrically for N with a Technicon Auto Analyzer II (Technicon Industrial Systems, Tarrytown, New York).
Soil sampling
To determine soil conditions, five soil samples were taken within each sample plot underneath these same five selected trees in August 2006 with a metallic tube (20 cm high and 10 cm in diameter) at the 0–15 cm soil layer and analyzed for total N and available N (NH +4 -N and NO −3 -N). The soil was sampled randomly 1–5 m away from the trunk of each of five individuals. Soil pH was measured in a suspension of soil in 10 mM CaCl2 (3:5 v:v). The fresh soils were sieved (<2 mm) and extracted in 1M KC1 (1:10 w/v) for 45 min. Total N was determined using Kjeldahl acid-digestion method (Bremner and Mulvaney 1982) and soil available nitrogen (NH +4 -N and NO −3 -N) was analyzed colorimetrically using a continuous-flow ion auto-analyzer as mentioned above.
Definitions and calculations
Area-based N resorption efficiency (NRE) was calculated as N mass per unit of leaf area (grams per square meter) of green (Ng) and senesced (Ns) leaves: NRE = [(Ng−Ns)/Ng]×100%, where Ng was N concentration in green leaves and Ns was N concentration in senesced leaves. For the calculation, leaf data were pooled together to derive the mean of each stand. The N concentrations in senesced leaves were considered a direct indicator of N resorption proficiency (NRP), which is defined as the absolute level to which N is reduced in senesced leaves (Killingbeck 1996). Plants with a lower N concentration in their freshly fallen leaves are considered more proficient.
Statistical analyses
Statistical tests were performed with SYSTAT 12 for Windows (Systat Inc., Chicago, IL). Prior to analysis, whenever needed, LMA and N data were transformed logarithmically to meet the assumptions of normality and homogeneity. NRE data were arcsin-transformed. One-way analysis of variance was used to test the effect of stand age on leaf N and soil N. When the difference was significant, post-hoc multiple comparisons were subsequently made using Tukey–Kramer test. Regression analysis was used to determine the relationships between natural-log stand age and green- and senesced-leaf N concentrations. Analysis of covariance (ANCOVA) was used to assess the effects of stand age and the covariant soil N on plant variables. Soil available N, which reflects N availability for plant uptake, was used in the ANCOVA.
Results
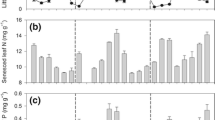
Green-leaf mass per unit area (LMA) ranged from 77.2 to 99.8 g m−2 and increased with stand age (Fig. 1A). Mass-based N in green leaves ranged from 20.2 to 25.7 mg g−1. Also, both mass- and area-based N concentrations increased significantly with stand age (Figs. 1B and C). Along the stand age chronosequence, mass-based N concentration increased by 27%. There were strong effects of stand age on green-leaf LMA and N concentration (Table 1), both of which were highest in the 139-yr stands and lowest in the 7-yr stands.

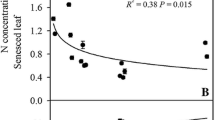
Variations in green-and senesced-leaf mass per unit area (LMA), mass-based and area-based N concentrations from the four study stand-age classes in northwestern Ontario, Canada. LMA=leaf mass per unit area. Vertical bars represent mean± 1 SE. ♦=7-yr stands, ■=25-yr stands, ▲=85-yr stands, ●=139-yr stands. Filled and open symbols represent green and senesced leaves respectively. (A) LMA vs. stand age (Green-leaf: ln y= −46.1+11.1ln x, r 2 = 0.91, P < 0.001. Senesced-leaf: ln y= −40.2+10.3ln x, r 2 = 0.78, P < 0.001). (B) [N]mass vs. stand age (Green-leaf: ln y= −31.7+11.3ln x, r 2 = 0.86, P < 0.001. Senesced-leaf: ln y= −8.4+5.7 ln x, r 2 = 0.92, P < 0.001). (C) [N]area vs. stand age (Green-leaf: ln y= −0.7+5.9ln x, r 2 = 0.93, P < 0.001. Senesced-leaf: ln y= −5.7+3.9ln x, r 2 = 0.92, P < 0.001)
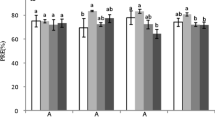
N resorption efficiency (NRE) presented mean values ranging from 58.4% to 68.5% based on leaf mass and from 67.1% to 75.3% based on leaf area. NRE differed significantly among stands, with the highest values in 7-yr stands and lowest in 139-yr stands (Table 1).
Mass-based N resorption proficiency (NRP), i.e., N concentrations in senesced leaves, ranged from 6.4 to 10.7 mg g−1. NRP decreased with stand age (i.e., the absolute N values increased with stand age). Along the stand age chronosequence, mass-based NRP decreased by 40% (Fig. 1).
Soil total N differed among four stand-age classes (Table 1). There was a significant increase in soil total N with stand age. However, soil available N did not differ significantly with stand age. Most plant variables were closely correlated to each other (Table 2). A significant positive correlation was found between green-leaf N and soil total and available N. NRE based on leaf mass was negatively correlated with soil total and available N (Table 2). ANCOVA showed that plant response variables were mostly affected by stand age while soil available N was insignificant (Table 3).
Discussion
There were significant shifts in green- and senesced-leaf N concentrations within an individual species, P. tremuloides along the stand age chronosequence (Table 1), suggesting that senesced N concentration (i.e., resorption proficiency) could change with stand development. N resorption based on both leaf area and mass showed similar patterns with stand development, with the highest values in 7-year stands and the lowest values in 139-year stands (Table 1). These results supported our hypothesis that stand age has an impact on plant N conservation strategies.
Our results showed an increase in soil total N status with stand development in P. tremuloides dominated stands (Table 1). Similar findings were reported in Mountain Ash stands (Polglase et al. 1992) and boreal forests (Brais et al. 1995). However, there was no significant difference in soil available N among four stand-age classes, consistent with the results by Rundel and Parsons (1980) who found that nutrient levels were similar among the sites along an age gradient surface soil. It is not surprising that there were different patterns of soil total and available N in our study since total amount of N does not reflect plant-available N, which mainly comes from mineralization. Often high humus content is even negatively correlated with N availability (Christian Körner, personal communication). In this study, soil available N was not significantly related to soil total N (Table 1). In fact, there is no agreement on how N transformations change with secondary succession. In boreal forests, N mineralization rates have been reported to increase (Polglase et al. 1992; Brais et al. 1995) or decrease (Van Cleve et al. 1983; Wardle et al. 1997; Deluca et al. 2002) with secondary succession. This divergence in findings may be dependent upon the methodologies used or the severity and characteristics of the disturbance prior to secondary succession (Vitousek et al. 1989). Wildfire has a net negative effect on C and N mineralization through consumption of organic substrates (White et al. 2004), and therefore N mineralization rate would increase through stand development as soil organic matter stocks increase. P. tremuloides is restricted to soils with little organic matter after wildfire. Thus temporal patterns of N availability largely parallel the loss and accumulation of organic matter as stands develop. Our results suggested that the N availability in our ecosystem reflected the balance of the loss of soil organic matter during fire and its accumulation after fire. Increasing N in soil with stand development could also be related to biological inputs of N fixation (Zackrisson et al. 2004; Deluca et al. 2008) and accumulative atmospheric N deposition (Magnani et al. 2007).
In our study, N concentration in green leaves increased with stand age (Table 1). Increasing N concentration with stand age could be a direct consequence of increasing soil N supply because green-leaf N concentration, in some instances, reflects site fertility (Lambers et al. 2008). In the present study, soil N availability, however, did not change with stand age (Table 1), suggesting factors other than soil N could have affected leaf-level N variation in P. tremuloides. The changes in green-leaf N with stand development might be related to the dilution effects because the greater leaf biomass production rates in younger stands could result in reduced N concentrations even if absolute uptake rates are high. Compared to old stands, young stands have rapid biomass production and thus higher N demands (Miller 1995), which could be a possible driver of higher N resorption efficiency in young stands. Other factors such as an altered light regime with stand development (Ilisson and Chen, unpublished data) could also influence green-leaf N concentrations because plants growing at different irradiances can have widely different traits such as photosynthetic capacity, growth rates and N economy (Chen and Klinka 1997; Hikosaka 2005).
All the stands we sampled were even-aged, dominated by Populus. Therefore, plant age would correspond to stand age. Because trees generally become old as stand ages, it is difficult to separate the effects of tree age from the effects of stand age on growth (Bond 2000; Mencuccini et al. 2007). For example, we could not sample 139-yr-old trees in the youngest stands and vice versa. In fact, chronosequence approach is still disputed because the space for time substitution assumption may not be met in sampling design (Johnson and Miyanishi 2008). However, this type of approach is useful for studying long-term changes in ecosystem structure and function over the life of a stand (Piccolo et al. 1994; Walker and del Moral 2003; Bloom and Mallik 2006).
In this study, P. tremuloides generally resorbs 50%–80% of its foliar N prior to leaf senescence, consistent with woody plants elsewhere (Yuan and Chen 2009). Earlier studies on tropical soil chronosequence in Hawaii suggested that the proportion of N and P resorbed by a native tree species, Metrosideros polymorpha, varies inversely with nutrient availability, from about 40% in intermediate-aged, fertile sites to nearly 70% in youngest and oldest low-N sites (Vitousek 1998). However, we did not find a significant effect of soil N on resorption efficiency in P. tremuloides (Table 3). No nutritional controls on N resorption efficiency were also found in other studies (see review by Aerts 1996).
It should be noted that calculations based solely on the difference in the N concentration between mature green and senescent leaves ignore possible changes in specific leaf mass due to resorption of carbon compounds during senescence. Thus, estimates of proportional N resorption made on the basis of leaf area may be more accurate than those made on the basis of leaf mass (van Heerwaarden et al. 2003; Luyssaert et al. 2005). In this study, we calculated the resorption of N from senesced leaves based on N content per unit leaf mass and per unit leaf area (Table 1). The results showed that both mass- and area-based N resorption responded to stand age in a similar pattern (Tables 1 and 2).
Our study supports the hypothesis that green-leaf N concentration increase, but N resorption efficiency decreases with stand development (Table 1). High N resorption in young stands in comparison with old stands suggests better conservation of N in more recently disturbed stands. Our results for N concentrations in senesced leaves of P. tremuloides ranged from 6.4 to 10.7 mg g−1, depending on stand age. Killingbeck (1996) suggested that any values less than 7 mg g−1 for N could be considered “complete resorption,” and values greater than 10 mg g−1 for N could be considered “incomplete resorption.” By these criteria, our results suggested that N resorption in our study shifted from complete to incomplete with stand development.
Previous studies have shown that the efficiency and proficiency of autumn N remobilization from senescing leaves exhibits genetic variation in Populus and other tree species (Aerts 1996; Killingbeck 1996; Harvey and van den Driessche 1999; Weih and Nordh 2002). If N economy differs among ecotypes such as mountain birch (Weih and Karlsson 1999) and among genotypes such as Salix (Weih and Nordh 2002), the N resorption in Populus would be expected to be affected by both genotype and environment (Cooke and Weih 2005). Unfortunately, we have no information about either ecotype or genotype of our studied species. According to our sampling approach, the studied species of P. tremuloides, however, were anticipated to come from the same ecotype and genotype, suggesting all differences was only dependent upon the effects of stand age.
The inverse of the N concentration in leaf litter has been used as an index of leaf-level N use efficiency (NUE) (Vitousek 1982; Aerts and Chapin 2000; Yuan and Li 2007). Following this approach, N resorption proficiency increased and leaf-level NUE decreased with stand age (Fig. 1). Because leaf-level NUE was often found to increase with decreasing soil N (Vitousek 1982; Yuan et al. 2006; Yuan et al. 2008), the higher leaf-level NUE in younger stands suggested that P. tremuloides used N more efficiently than in old stands, although these stands were similar in soil N availability (Table 1).
In summary, the extent of N resorption seems to change as stand ages: younger stands were more efficient and also more proficient in resorbing N than old stands. The significant effect of stand age on green- and senesced-leaf N concentrations supports the hypothesis that this species could adjust its resorption efficiency and proficiency in response to stand aging.
References
Aerts R (1996) Nutrient resorption from senescing leaves of perennials: are there general patterns? J Ecol 84:597–608
Aerts R, Chapin FS (2000) The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Adv Ecol Res 30:1–67
Alban DH (1985) Seasonal changes in nutrient concentration and content of aspen suckers in Minnesota. For Sci 31:785–794
Bergeron Y, Dubuc M (1989) Succession in the southern part of the Canadian boreal forest. Vegetatio 79:51–63
Bloom RG, Mallik AU (2006) Relationships between ericaceous vegetation and soil nutrient status in a post-fire Kalmia angustifolia-black spruce chronosequence. Plant Soil 289:211–226
Bond BJ (2000) Age-related changes in photosynthesis of woody plants. Trends Plant Sci 5:349–353
Bond-Lamberty B, Gower ST, Wang C, Cyr P, Veldhuis H (2006) Nitrogen dynamics of a boreal black spruce wildfire chronosequence. Biogeochemistry 81:1–16
Bond-Lamberty B, Peckham SD, Ahl DE, Gower ST (2007) Fire as the dominant driver of central Canadian boreal forest carbon balance. Nature 450:89–92
Brais S, Camire C, Bergeron Y, Pare D (1995) Changes in nutrient availability and forest floor characteristics in relation to stand age and forest composition in the southern part of the boreal forest of northwestern Quebec. For Ecol Manage 76:181–189
Brassard BW, Chen HYH (2008) Effects of forest type and disturbance on diversity of coarse woody debris in boreal forest. Ecosystems 11:1078–1090
Brassard BW, Chen HYH, Wang JR, Duinker PN (2008) Effects of time since stand-replacing fire and overstory composition on live-tree structural diversity in the boreal forest of central Canada. Can J For Res 38:52–62
Bremner JM, Mulvaney CS (1982) Nitrogen-total. In: Page AL, Miller RH, Keeney DR (eds) Methods of Soil Analysis, American Society of Agronomy, Soil Science Society of America, Madison, WI, pp. 595–624
Callaghan TV (1980) Age-related patterns of nutrient allocation in Lycopodium annotinum from Swedish Lapland—strategies of growth and population-dynamics of tundra plants. Oikos 35:373–386
Chen HYH, Klinka K (1997) Light availability and photosynthesis of Pseudotsuga menziesii seedlings grown in the open and in the forest understory. Tree Physiol 17:23–29
Chen HYH, Popadiouk RV (2002) Dynamics of North American boreal mixedwoods. Envir Rev 10:137–166
Cooke JEK, Weih M (2005) Nitrogen storage and seasonal nitrogen cycling in Populus: bridging molecular physiology and ecophysiology. New Phytol 167:19–30
Dellasala DA, Karr JR, Schoennagel T, Perry D, Noss RF, Lindenmayer D, Beschta R, Hutto RL, Swanson ME, Evans J (2006) Post-fire logging debate ignores many issues. Science 314:51–52
Deluca TH, Nilsson MC, Zackrisson O (2002) Nitrogen mineralization and phenol accumulation along a fire chronosequence in northern Sweden. Oecologia 133:206–214
Deluca TH, Zackrisson O, Gundale MJ, Nilsson MC (2008) Ecosystem feedbacks and nitrogen fixation in boreal forests. Science 320:1181
Duran J, Rodriguez A, Fernandez-Palacios JM, Gallardo A (2008) Changes in soil N and P availability in a Pinus canariensis fire chronosequence. For Ecol Manage 256:384–387
Groom PK, Lamont BB, Markey AS (1997) Influence of leaf type and plant age on leaf structure and sclerophylly in Hakea (Proteaceae). Aust J Bot 45:827–838
Hart SA, Chen HYH (2008) Fire, logging, and overstory affect understory abundance, diversity, and composition in boreal forest. Ecol Monogr 78:123–140
Harvey HP, van den Driessche R (1999) Poplar nutrient resorption in fall or drought: influence of nutrient status and clone. Can J For Res 29:1916–1925
Hikosaka K (2005) Leaf canopy as a dynamic system: ecophysiology and optimality in leaf turnover. Ann Bot 95:521–533
Huang JY, Zhu XG, Yuan ZY, Song SH, Li X, Li LH (2008) Changes in nitrogen resorption traits of six temperate grassland species along a multi-level N addition gradient. Plant Soil 306:149–158
Ice GG, Neary DG, Adams PW (2004) Effects of wildfire on soils and watershed processes. J For 102:16–20
Johnson EA, Miyanishi K (2008) Testing the assumptions of chronosequences in succession. Ecol Lett 11:419–431
Killingbeck KT (1996) Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology 77:1716–1727
Killingbeck KT, May JD, Nyman S (1990) Foliar senescence in an aspen (Populus tremuloides) clone - the response of element resorption to interramet variation and timing of abscission. Can J For Res 20:1156–1164
Kobe RK, Lepczyk CA, Iyer M (2005) Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology 86:2780–2792
Lambers H, Chapin FSI, Pons TL (2008) Plant physiological ecology. New York, Springer, p 610
Leduc SD, Rothstein DE (2007) Initial recovery of soil carbon and nitrogen pools and dynamics following disturbance in jack pine forests: a comparison of wildfire and clearcut harvesting. Soil Biol Bioch 39:2865–2876
Lindenmayer DB, Foster DR, Franklin JF, Hunter ML, Noss RF, Schmiegelow FA, Perry D (2004) Ecology—Salvage harvesting policies after natural disturbance. Science 303:1303
Luyssaert S, Staelens J, De Schrijver A (2005) Does the commonly used estimator of nutrient resorption in tree foliage actually measure what it claims to? Oecologia 144:177–186
Madritch M, Donaldson JR, Lindroth RL (2006) Genetic identity of Populus tremuloides litter influences decomposition and nutrient release in a mixed forest stand. Ecosystems 9:528–537
Magnani F, Mencuccini M, Borghetti M, Berbigier P, Berninger F, Delzon S, Grelle A, Hari P, Jarvis PG, Kolari P, Kowalski AS, Lankreijer H, Law BE, Lindroth A, Loustau D, Manca G, Moncrieff JB, Rayment M, Tedeschi V, Valentini R, Grace J (2007) The human footprint in the carbon cycle of temperate and boreal forests. Nature 447:848–850
Mencuccini M, Martinez-villalta J, Hamid HA, Korakaki E, Vanderklein D (2007) Evidence for age- and size-mediated controls of tree growth from grafting studies. Tree Physiol 27:463–473
Miller HG (1995) The Influence of stand development on nutrient demand, growth and allocation. Plant Soil 169:225–232
Nepstad DC, Verissimo A, Alencar A, Nobre C, Lima E, Lefebvre P, Schlesinger P, Potter C, Moutinho P, Mendoza E, Cochrane M, Brooks V (1999) Large-scale impoverishment of Amazonian forests by logging and fire. Nature 398:505–508
Piccolo MC, Neill C, Cerri CC (1994) Net nitrogen mineralization and net nitrification along a tropical forest-to-pasture chronosequence. Plant Soil 162:61–70
Polglase PJ, Attiwill PM, Adams MA (1992) Nitrogen and phosphorus cycling in relation to stand age of Eucalyptus regnans F. Muell.II. N-mineralization and nitrification. Plant Soil 142:167–176
Reich PB, Bakken P, Carlson D, Frelich LE, Friedman SK, Grigal DF (2001) Influence of logging, fire, and forest type on biodiversity and productivity in southern boreal forests. Ecology 82:2731–2748
Richardson SJ, Peltzer DA, Allen RB, McGlone MS (2005) Resorption proficiency along a chronosequence: responses among communities and within species. Ecology 86:20–25
Ruark GA, Bockheim JG (1988) Biomass, net primary production, and nutrient distribution for an age sequence of Populus tremuloides ecosystems. Can J For Res 18:435–443
Rundel PW, Parsons DJ (1980) Nutrient changes in 2 chaparral shrubs along a fire-induced age gradient. Am J Bot 67:51–58
Simard DG, Fyles JW, Pare D, Nguyen T (2001) Impacts of clearcut harvesting and wildfire on soil nutrient status in the Quebec boreal forest. Can J Soil Sci 81:229–237
Van Cleve K, Oliver L, Schlentner R, Viereck LA, Dyrness CT (1983) Productivity and nutrient cycling in taiga forest ecosystems. Can J For Res 13:747–766
van Heerwaarden LM, Toet S, Aerts R (2003) Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: facts and solutions. Oikos 101:664–669
Vitousek PM (1982) Nutrient cycling and nutrient use efficiency. Am Nat 119:553–572
Vitousek PM (1998) Foliar and litter nutrients, nutrient resorption, and decomposition in Hawaiian Metrosideros polymorpha. Ecosystems 1:401–407
Vitousek PM, Matson PA, VanCleve K (1989) Nitrogen availability and nitrification during succession - Primary, secondary, and old-field seres. Plant Soil 115:229–239
Walker LR, del Moral R (2003) Primary succession and ecosystem rehabilitation. Cambridge University Press
Wardle DA, Zackrisson O, Hornberg G, Gallet C (1997) The influence of island area on ecosystem properties. Science 277:1296–1299
Weih M, Karlsson PS (1999) Growth response of altitudinal ecotypes of mountain birch to temperature and fertilisation. Oecologia 119:16–23
Weih M, Nordh NE (2002) Characterising willows for biomass and phytoremediation: growth, nitrogen and water use of 14 willow clones under different irrigation and fertilisation regimes. Biomass Bioenergy 23:397–413
White LL, Zak DR, Barnes BV (2004) Biomass accumulation and soil nitrogen availability in an 87-year-old Populus grandidentata chronosequence. For Ecol Manage 191:121–127
Yuan ZY, Chen HYH (2009) Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Global Ecol Biogeogr 18:11–18
Yuan ZY, Chen HYH, Li LH (2008) Nitrogen use efficiency: does a trade-off exist between the N productivity and the mean residence time within species? Aust J Bot 56:272–277
Yuan ZY, Li LH (2007) Soil water status influences plant nitrogen use: a case study. Plant Soil 301:303–313
Yuan ZY, Li LH, Han XG, Chen SP, Wang ZW, Chen QS, Bai WM (2006) Nitrogen response efficiency increased monotonically with decreasing soil resource availability: a case study from a semiarid grassland in northern China. Oecologia 148:564–572
Yuan ZY, Li LH, Han XG, Huang JH, Jiang GM, Wan SQ (2005a) Soil characteristics and nitrogen resorption in Stipa krylovii native to northern China. Plant Soil 273:257–268
Yuan ZY, Li LH, Han XG, Huang JH, Wan SQ (2005b) Foliar nitrogen dynamics and nitrogen resorption of a sandy shrub Salix gordejevii in northern China. Plant Soil 278:183–193
Yuan ZY, Liu WX, Niu SL, Wan SQ (2007) Plant nitrogen dynamics and nitrogen-use strategies under altered nitrogen seasonality and competition. Ann Bot 100:821–830
Zackrisson O, Deluca TH, Nilsson MC, Sellstedt A, Berglund LM (2004) Nitrogen fixation increases with successional age in boreal forests. Ecology 85:3327–3334
Acknowledgements
We thank Brian Brassard and Gareth Davis for their assistance in field work. This work was supported by the Natural Science and Engineering Research Council of Canada (283336-04 & STPGP 322297), the National Centre of Excellence Network of Sustainable Forest Management, and the Natural Science Foundation of China (30600076).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Martin Weih.
Rights and permissions
About this article
Cite this article
Yuan, Z.Y., Chen, H.Y.H. Changes in nitrogen resorption of trembling aspen (Populus tremuloides) with stand development. Plant Soil 327, 121–129 (2010). https://doi.org/10.1007/s11104-009-0036-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-009-0036-8