
Abstract
A descriptive temporal model is considered to be the best available estimator for accretion, resorption and proportional nutrient resorption. However, ecological studies rarely collect sufficient data for applying such a model. A less-demanding and commonly used estimator for proportional resorption (PR) calculates PR as the percentage of the nutrient pool that is withdrawn from mature foliage before leaf abscission. Data from an intensive sampling campaign of the aboveground nutrient pools and fluxes of two Betula pendula Roth. stands were used. We showed that the commonly used estimator is not an accurate estimator for accretion, resorption and proportional resorption. The commonly used estimator underestimated the proportional resorption of N on the average by 3–10%, and the proportional resorption of P by 20–25%. The low accuracy of the estimations was shown to be caused by a lack of selectiveness of the commonly used estimator. In other words, the commonly used estimator does not measure the underlying processes in specific nutrient accretion and resorption at the stand level. However, when a sufficiently high sampling density with several samples at a given point in time is used, then the commonly used estimator preserves the ranking relationship between the PR of different sites for N in 97% of the cases and for P in 71%. The commonly used estimator can thus be used in comparative studies as an index for proportional nutrient resorption only. The quantitative results should not be taken literally, as they are based on only two sets of observations. However, the results show that the commonly used estimator should no longer be used as a measure for accretion, resorption or PR whenever the plant accretes nutrients in the foliage as a compensation for nutrient losses due to foliar leaching and litterfall during the growing season.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Resorption is a nutrient conservation mechanism that affects key processes such as nutrient uptake (Chapin 1980), competition (Killingbeck 1985), carbon cycling (Asner et al. 2001) and fitness in terms of growth and reproduction (May and Killingbeck 1992). Nitrogen and phosphorus, especially, are highly resorbed as a reaction to self-shading as the canopy grows (Saur et al. 2000; Franklin and Agren 2002) or from senescing leaves before abscission (Staaf 1982; Chapin and Kedrowski 1983). On the average, around 50% of the N and P are resorbed from the tree foliage (Aerts 1996). Nutrients that are resorbed and stored internally are a readily available pool of nutrients for the plant. This pool allows a higher rate of growth than would be possible based on nutrient uptake from the soil alone. Nutrients that are not resorbed will be circulated via litterfall. However, nutrient circulation via litterfall has the disadvantage that neighbouring plants compete for the nutrients, and that nutrients can be lost through leaching or incorporated in stable soil pools. There is still some controversy about whether resorption is an adaptive strategy of plants to their growing environment. As arguments against the theory of an adaptive strategy, resorption was reported to be: (1) similar under ambient or elevated CO2 concentrations (Norby and Cotrufo 1998; Norby et al. 2000), (2) lower in evergreens than in deciduous trees for N, (3) similar in evergreens and deciduous trees for P, and (4) higher, lower or similar in fertile soils compared to infertile soils (Aerts 1996; Knops et al. 1997; Wright and Westoby 2004). The ecological theory was revised on the basis of these unexpected or indisputable observations: plants growing in infertile environments can conserve nutrients mainly by extending the life span of their plant parts and/or by minimizing the nutrient content of those parts that are abscissed (Eckstein et al. 1999; Oleksyn et al. 2003).
The resorption process is commonly quantified by means of the parameter nutrient resorption efficiency (van Heerwaarden et al. 2003), which gives the proportion of the nutrient pool that was withdrawn from the foliage before leaf abscission. The parameter of nutrient resorption efficiency is not more than a proportion (see Eqs. 4, 5). Following on Wright and Westoby (2004), we therefore used the wording proportional nutrient resorption. A reliable estimate of proportional nutrient resorption should be derived from its underlying physiological processes, i.e. accretion and resorption. The cumulative accretion or uptake of nutrients is commonly estimated by quantifying the nutrient pool in mature leaves. The cumulative resorption of nutrients is estimated as the difference between the nutrient pools of mature and abscissed leaves. Finally, the commonly used estimator of proportional nutrient resorption assumes that the canopy nutrient pools are not affected by canopy exchange.
Although a lot of ecological theory is based on, or revised on the basis of, observation and comparison of the commonly used estimator of proportional nutrient resorption, one important issue has not yet been addressed: is this commonly used estimator a reliable parameter? Therefore, the aims of this study are to test whether the commonly used estimator: (1) is an accurate and precise estimator for proportional resorption, (2) measures the underlying physiological processes of resorption and accretion, and (3) preserves differences in resorption, accretion and proportional resorption (PR) between sites.
2 Materials and methods
2.1 Experimental set-up
We made a comparative study in two 30×40 m experimental plots of even-aged silver birch (Betula pendula Roth.) (Table 1) situated in southwestern Finland in the southern boreal coniferous zone (Ahti et al. 1968). The Harjavalta plot (61°19′N, 22°07′E) is situated about 30 km from the coast, and the area was subjected to a heavy pollution load since the 1940s, mainly from a smelter producing copper and nickel. The pollution load and the effects of pollution on the ecosystems in the vicinity of the plant were well documented in the literature and summarized by Kiikkilä (2003). The Hämeenkyrö plot (61°42′N, 23°10′E) is situated about 80 km northeast of Harjavalta. The plot is located more than 40 km from industry and is not affected by known local sources of pollution. The sandy soil underlying both plots is a Dystric Cambisol (FAO classification).
The experimental design on both plots was the same. Thirty-six stand throughfall collectors and 18 litterfall traps were distributed systematically over the plot. The throughfall collectors consisted of plastic funnels with a 0.03 m2 collection surface, which drained into a 3-l plastic bag. The litterfall traps were funnel-shaped with a collection surface of 0.5 m2 . A cotton bag with a mesh bottom for improved drainage was fitted to the neck of each funnel. Following measurement of the tree stand, 45 neighbouring trees were chosen on each 30×40 m plot. The selected trees were of the same height and diameter class, and had a dominant or co-dominant position in the stand. The 45 trees were divided into three different sets of 15 trees to avoid excessive leaf removal. Only one set of 15 trees was sampled each sampling period, that way less than 4% of the leaf mass of the sampling trees was lost due to sampling. Foliar samples were taken from the crown at a height of 10, 16 and 22 m above ground level. At each of the three canopy positions, a sample of at least 200 leaves was taken per individual tree. Data from the three different crown positions were used for a more accurate estimate of average nutrient concentrations. In each set of 15 trees, two trees were equipped with a stemflow collector. Stemflow was therefore measured on six trees on each plot. The stemflow collectors consisted of circular collars connected to a 40-l barrel by plastic tubing. Six bulk precipitation collectors were placed in a straight line on an open field located less than 500 m from the plots. The construction of the bulk precipitation collectors was similar to that of the throughfall collectors. Rainwater and litter samples were collected weekly from bud break (week 18) to abscission (week 45) in 2002. The mass of each throughfall (MTFΔt , g m−2), stemflow (MSFΔt , g m−2) and bulk precipitation (MBPΔt , g m−2) sample was recorded during sampling. Foliage was sampled 18 times, i.e. in weeks 20, 21, 22, 24, 26, 28, 31, 33, 35, 37, 38, 39, 40, 41, 42, 43, 44 and 45. The water, litter and foliage samples were transported to the laboratory within 10 h of sampling.
2.2 Sample preparation and chemical analysis
Before analysis, the samples from different weeks were bulked so that rainwater and litter had the same sampling frequency as foliage (18 times). Subsequently, the 36 throughfall and six bulk precipitation samples were pair-wise bulked; samples from neighbouring sampling locations were bulked according to their relative weights. Consequently, 18 throughfall ([TFx,Δt], g l−1), three bulk precipitation ([BPx,Δt], g l−1) and six stemflow (not bulked) ([SFx,Δt], g l−1) samples were analysed each sampling period for each plot. The pH was measured on unfiltered samples. The samples were then filtered through a 0.45 μm membrane filter. Part of the filtrate was preserved with concentrated HNO3 prior to determination of Na+ by inductively coupled plasma atomic emission spectrometry (ICP-AES). The unpreserved part of the filtrate was frozen prior to determination of Ntot by flow injection analysis, and NH +4 , SO 2−4 , NO −3 and PO 3−4 by ion chromatography. Foliar samples were dried at 60°C, the number of leaves was counted and the sample mass weighed. The mass per leaf for fresh leaves was calculated (LEAFSMΔt , g leaf−1) for each plot, for all 18 sampling periods, and for all 45 foliage samples separately. Litterfall samples were dried at 60°C and leaves were separated from twigs, buds and seeds. Leaves were counted and weighed (LITMΔt , g m−2) so that the mass per leaf of litter could be calculated (LITSMΔt , g leaf−1). Samples from the 18 litterfall traps were then bulked in threes: six samples were therefore analysed per plot. Unwashed leaves and litterfall were milled and sieved before analysis. The N concentration of the leaves ([LEAFx,t], g g−1) and litterfall ([LITx,Δt], g g−1) was determined without further pre-treatment on a CHN analyser. The P and Na concentrations in the leaves were determined by ICP-AES following wet digestion in HNO3/H2O2. The leaf samples were digested by a Closed Wet digestion method in a microwave. The results were expressed per 105°C dry mass.
The quality of the analytical methods was checked by means of method blanks, repeated measurements of internal reference samples, repeated measurement of certified reference samples and participation in inter-laboratory tests. When episodes of high NH +4 or Na+ deposition were observed, their timing and magnitude was compared to respectively, NH +4 and Na+ measurements on the EMEP plots, which are managed by the Finnish Meteorological Institute (unpublished data). Episodes of high NH +4 or Na+ deposition often coincided with air masses transported over the Baltic Sea and/or high precipitation amounts.
2.3 Resorption, accretion and proportional resorption
A descriptive temporal model that uses data from intensive sampling of the aboveground nutrient pools and fluxes was developed (Duchesne et al. 2001) for estimating cumulative accretion and resorption at the whole stand level throughout the growing season. Nutrient losses from the foliage pool can arise from four principal pathways: litterfall, leaf leaching, resorption and herbivory. As no signs of pests were observed, nutrient losses through herbivory were neglected in this study. Consequently, net resorption at the stand level is the sum of temporal changes in nutrient pools that cannot be allocated to litterfall and leaching:
where LEAFx,Δt (g m−2) is the change in the foliar pool of nutrient x in the period Δt, LITx,Δt (g m−2) is the loss of nutrient x due to litterfall in period Δt, and LEACHx,Δt (g m−2) is the change of the nutrient pool due to interactions with incident precipitation in period Δt. The cumulative accretion (cACC x ) for the growing season was calculated as \(\sum_{\Delta t= 1}^{18}{\rm nRES}_{{x,\Delta t}}\) for nRESx,Δt>0. The cumulative resorption (cRES x ) was calculated as \(\sum_{\Delta t= 1}^{18} {\rm nRES}_{{x,\Delta t}} \) for nRESx,Δt<0.
Changes in the foliar nutrient pool in period Δt (LEAFx,Δt) were calculated as:
where LEAFx,t (g m−2) is the nutrient pool at time t, which was calculated as the product of the nutrient concentration ([LEAFx,t], g g−1) and the foliar mass at time t (g m−2). The relative spring-time build-up of the foliar mass was modelled using the effective temperature sum (Kellomäki et al. 2000). Parameter settings for southwestern Finland were chosen according to Kellomäki et al. (2000, 2001). The relative spring-time build-up was then multiplied by the total biomass of leaves in litterfall ( \(\sum_{\Delta t= 1}^{18}{\rm LITM}_{\Delta{t}} \), g m−2) and corrected with the mass per leaf for litter (LITSM t , g leaf−1) and mass per leaf for fresh leaves (LEAFSM t , g leaf−1) to obtain the leaf mass at sampling time t (LEAFM t , g m−2). The loss of nutrient x due to litterfall in period Δt (LITx,Δt, g m−2) was calculated as the product of the nutrient concentration in litterfall ([LITx,Δt], g g−1) and the biomass of leaves in litter (LITMΔt , g m−2) in period Δt. The change of the foliar nutrient pool due to interactions between the canopy, and the precipitation and interception deposition (LEACHx,Δt, g m−2) was calculated as follows:
where TFx,Δt is the nutrient flux in throughfall (g m−2), SFx,Δt is the nutrient flux in stemflow (g m−2), BPx,Δt is the nutrient flux in bulk precipitation (g m−2), and DDx,Δt is the nutrient flux in dry deposition (g m−2) in period Δt. Except for DDx,Δt, all fluxes are calculated from the product of the measured nutrient concentration ([TFx,Δt], [SFx,Δt], [BPx,Δt], g g−1) and the measured mass of the flux (MTFΔt , MSFΔt , MBPΔt , g m−2). Consequently, the DDx,Δt flux has to be estimated before LEACHx,Δt and thus nRESx,Δt (g m−2) can be calculated. nRESx,Δt was calculated on the basis of three different estimates for the DDx,Δt flux. In the most conservative approach, the DDx,Δt flux was absent; in a worst case approach, the DDx,Δt flux was set to equal BPx,Δt; and in the last approach, the DDx,Δt flux was estimated using a seasonal dry deposition factor (DDF) for Na+ (Ulrich 1983). The last method assumes that leaching of Na+ is negligible compared to the Na+ deposition flux (LEACHNa≈0). This assumption allows for the calculation of the DD x flux on the basis of the BP x , TF x and SF x flux. The DDF, which is calculated as the ratio between the estimated DD x and the BP x flux of Na+, is then assumed to be a model for dry deposition of particles. Despite the fact that this assumption is not valid when the deposition processes are principally different, which is true for N (Beier 1991), the DDFs were calculated for both plots from the seasonal data and then used to calculate the DDx,Δt flux of N and P for every sampling period.
The descriptive temporal model was considered to be the best available estimator of the real foliar accretion and resorption because it estimates accretion and resorption on the basis of 18 independent time steps, and each time step uses direct measurements of all nutrient pools and fluxes, except for dry deposition. The proportional nutrient resorption (PR, %) at the stand level was then calculated as:
However, ecological studies rarely collect sufficient data for applying a descriptive temporal model and calculating the PR by means of Eq. 4. A less-demanding approach is commonly used, in which PR at the stand level is calculated as follows (Helmisaari 1992):
Where PR is the proportional resorption (%), t is the time before senescence, i.e. the last weeks of July, and Δt is the period of litterfall from August until November. Many authors suggest using leaf area as a basis for expressing the PR (Oland 1963; Nordell and Karlsson 1995). Using leaf area as a basis assumes that the leaf area is constant during the course of the observations. Although this assumption might be valid for the commonly used estimator, it is not valid for the best available estimator as this estimator uses data from bud break to the end of litterfall. Therefore, accretion, resorption and PR were expressed on a unit stand area basis because the stand area is constant during the growing season. Due to the fact that both leaf area and stand area are constant during the course of observations for the commonly used estimator, simple dimensional analysis shows (not given) that the commonly used estimator gives the same proportional nutrient resorption irrespective of whether the nutrient pools are expressed on a unit leaf area or a unit stand area basis.
2.4 Bootstrap simulations
Bootstrap simulations can be used as repetitions for hypothesis testing and for calculating confidence intervals of the estimated parameters when real repetitions are not available (Efron and Tibshirani 1993). Two hundred and fifty bootstrap simulations were made for each plot and each element. Within each simulation, three foliar (one from the bottom, one from the middle and one from the top of the crown), one throughfall, one stemflow, one bulk precipitation and one leaf litterfall sample was randomly selected for each sampling period from week 18 until week 45 from the observations of LEAFM t , LEAFMx,t-1, LITSM t , LEAFSM t , LITMΔt , [LITx,Δt], [TFx,Δt], [SFx,Δt], [BPx,Δt], MTFΔt , MSFΔt , MBPΔt and DDx,Δt. Consequently, each bootstrap simulation had its own real foliar accretion, resorption and proportional nutrient resorption. All the bootstrap simulations were re-run with a higher sampling density at a given point in time. The sampling density was increased and 30 foliar (for ten trees one from the bottom, one from the middle and one from the top of the crown), three throughfall, three stemflow, three bulk precipitation and six observations for leaf litterfall were used for each of the 18 time steps.
2.5 Hypothesis testing
2.5.1 Is the commonly used estimator for PR accurate and precise?
The descriptive temporal model was used to calculate the best available estimator of the real cumulative accretion, cumulative resorption, leaching and PR for each bootstrap. The proportional nutrient resorption was then calculated by Eq. 4. The 95% confidence limit for the cumulative accretion, resorption, and PR were calculated from the 250 bootstrap simulations. In addition, each bootstrap simulation contained the required data for calculating the PR by means of the commonly used estimator (Eq. 5). For each bootstrap simulation, the best available estimator for proportional nutrient resorption (Eq. 4) was compared to the commonly used estimator for PR (Eq. 5). If PR calculated with Eq. 5 is an accurate and precise estimator for PR, it should equal PR calculated with Eq. 4.
2.5.2 Is the commonly used estimator for PR selective?
A selective estimator provides only responses for the processes being studied. Consequently, a selective estimator for PR responds to changes in the cumulative resorption and accretion. Equation 4 thus provides a selective estimator for PR. Therefore, the commonly used estimator (Eq. 5) will be selective if the following statements are valid: (a) cACCx,Δt ≈ ([LEAFx,Δt]×LEAFMΔt ) or the cumulative accretion or uptake of nutrients is related to the nutrient pool in mature leaves, (b) \({\rm cRES}_{x,\Delta{t}}\approx \left({\left[{\rm LEAF}_{x,\Delta{t}}\right]} \times{\rm LEAFM}_{\Delta{t}} \right) - \left({\left[{\rm LIT}_{x,\Delta{t}} \right]} \times{\rm LITM}_{\Delta{t}} \right)\) or the cumulative resorption of nutrients is related to the difference between nutrient pools of mature and abscised leaves, and (c) LEACHx,Δt=0 or the nutrient pools are not affected by canopy exchange. Statements (a), (b) and (c) were derived by substituting Eq. 5 in 4. Substitution is justified because each forest plot has only one real value for PR. PR calculated with the commonly used estimator must therefore equal PR calculated with Eq. 4. Statements (a) and (b) were evaluated by calculating Pearson correlation coefficients between both terms in the statements. Correlation coefficients were calculated for the low, as well as for the high sampling density simulations. Statement (c) was evaluated by comparing the average accretion, resorption, and PR calculated with and without leaching for the low sampling density simulations.
2.5.3 Does the commonly used estimator for PR preserve differences between the plots?
Hundred of the 250 bootstrap simulations for nutrient cycling at Hämeenkyrö were randomly selected. Each of these randomly selected simulations was matched with a randomly selected bootstrap simulation for Harjavalta. For each of the 100 pairs, the best available estimator for accretion, resorption and PR, as well as the commonly used estimator for accretion, resorption and proportional resorption, were available for both plots. The ranking relationship of the best available estimator for accretion, resorption and proportional resorption for Hämeenkyrö and Harjavalta was determined and compared to the relationship for the commonly used estimator. The preservation of the relationship was tested for the simulations that were based on both the low and high sampling density.
3 Results
3.1 N and P concentrations and biomass
Seasonal variation of the N and P concentrations, and foliar and litterfall biomass, were similar in Hämeenkyrö and Harjavalta (not shown). Buds began to break in week 18 and foliar biomass reached its maximum around week 31 (Fig. 1). Bud break was followed by a rapid increase of leaf N and P concentrations up until week 20 (Fig. 1). From then on, N showed a different dynamic pattern than P. The N concentration decreased almost continuously until week 38 and decreased rapidly afterwards. The P concentrations deceased between week 20 and 25. After this point of time, the P concentration remained relatively constant during the growing season. Litterfall started in week 25, and therefore occurred before the foliar biomass reached its maximum. A sharp increase in litterfall occurred in week 35 (Fig. 1). The dynamics for N and P concentration in litterfall were similar to the dynamics for N and P in foliage; the N concentrations decreased during the growing season, whereas the P concentrations remained relatively constant (Fig. 1).

Seasonal variation of N (first panel) and P (second panel) concentration in foliage and litterfall in Hämeenkyrö. The average nutrient concentration in foliage is shown by the solid line. The average nutrient concentration in litterfall is shown by the dotted line. The third panel shows the average evolution of the foliar biomass (solid line). The average evolution of the litterfall biomass is shown by the dotted line. The error bars illustrate the 95% CI
The level to which nutrient concentrations are reduced in senesced leaves or litter is called the resorption proficiency and is a different but complementary index of nutrient conservation from the proportional nutrient resorption (Killingbeck 1996). According to Killingbeck’s (1996) benchmarks of resorption proficiency for deciduous species, N and P resorption were incomplete in Hämeenkyrö and Harjavalta in the year 2002.
3.2 Best available estimator of accretion and resorption
The differences between the N and P dynamics were reflected in the pattern of the cumulative N and P accretion and resorption. Patterns in the cumulative accretion and resorption were similar in Hämeenkyrö and Harjavalta (Fig. 2). At the start of the growing season, N was accreted in the leaves. N accretion in the leaves ceased almost completely in week 31 (Fig. 2) when the canopy was fully formed (Fig. 1). Phosphorus, however, continued to be accreted in the leaves until the end of the growing season. Resorption of both N and P began at the start of the growing season, but remained low during canopy formation. From week 31 onward, resorption increased rapidly (Fig. 2) and, by the end of the growing season, the best available estimator for proportional resorption (±95% confidence limit) was 68.30 (±9.99)% and 62.46 (±7.69)% of the total N pool in Hämeenkyrö and Harjavalta, respectively; and 55.18 (±12.51)% and 59.38 (±10.84)% of the total P pool in Hämeenkyrö and Harjavalta, respectively.

Best available estimator of the cumulative foliar accretion and resorption for N (first and second panel) and P (third and fourth panel) between week 18 and 45 in year 2002 for the plots in Hämeenkyrö and in Harjavalta. Cumulative accretion and resorption were calculated at the stand level according to Eq. 1. The positive line shows the average cumulative accretion of N or P, and the negative line shows the average cumulative resorption of N or P. The error bars show the 95% CI. All calculations are based on a low sampling density. The difference between the cumulative accretion and resorption is a measure for the amounts of nutrients lost through litterfall and leaching
All the data needed for the model can be easily obtained from direct measurements except for the dry deposition flux (Anderson and Hovmand 1999). As dry deposition was not measured, it had to be estimated. Although dry deposition can affect accretion, resorption and thus PR through foliar leaching, there was no evidence to show that the three tested scenarios for dry deposition resulted in different estimations for foliar N accretion (ANOVA; P=0.423 in Hämeenkyrö, P=0.617 in Harjavalta), N resorption (P=0.309, P=0.414), P accretion (P=0.969, P=0.989), and P resorption (P=0.779, P=0.857). Consequently, all three scenarios were equally suitable for estimating foliar accretion and resorption. However, we preferred to set the dry deposition equal to the bulk deposition, as the absence of dry deposition is an unrealistic assumption and calculating the DDF according to Ulrich would have been based on several untested assumptions.
3.3 Is the commonly used estimator for PR accurate and precise?
The commonly used estimator was, on the average, neither accurate nor precise for estimating the N PR in Harjavalta or the P PR in Hämeenkyrö and Harjavalta (Fig. 3). As an exception, the commonly used estimator for PR resulted in an accurate and precise estimate of the proportional resorption for N in Hämeenkyrö (Fig. 3). The good result for PR for N in Hämeenkyrö was coincidently caused by averaging the results of the individual bootstrap simulations: the low accuracy of the commonly used estimator was confirmed by low Pearson correlation coefficients (r) between the commonly used estimator and best available estimator for PR for the individual simulations, i.e. r=0.210 for N in Hämeenkyrö, r=0.080 for N in Harjavalta, r=0.268 for P in Hämeenkyrö and r=0.156 for P in Harjavalta. An inaccurate estimator has a correlation coefficient which strongly deviates from 1. The precision, expressed as the 95% confidence interval (95% CI), ranged from 7.7% to 10% for N and from 11% to 13% for P.

As the low accuracy and precision of the commonly used estimator may have been caused by the variability introduced by the low sampling density of the experiment, increasing the sampling density at all 18 sampling times should increase the accuracy and precision. This was tested by re-running all the bootstrap simulations with a higher sampling density at all 18 sampling times. On the average, the higher sampling density resulted in an increased precision for the commonly used estimator. The PR values were estimated with a precision, expressed as the 95% CI, ranging from 2.6% to 6.3% for N and from 2.8% to 5.7% for P. Although the precision increased considerably, the accuracy of the commonly used estimator did not change very much as indicated by the weak correlations between commonly used estimator and best available estimator for the individual simulations (r=0.205 for N in Hämeenkyrö, r=0.308 for N in Harjavalta, r=0.02 for P in Hämeenkyrö, and r=0.072 for P in Harjavalta.). Consequently, the low sampling density at all 18 sampling times caused the low precision, but was not the overriding cause for the lack of accuracy.
3.4 Is the commonly used estimator for PR selective?
The bootstrap simulations contained the data required for quantifying the validity of statements (a) through (c) on the plots in Hämeenkyrö and Harjavalta. However, the evidence which was found in support of these statements was weak (Table 2). The relationship between the cumulative accretion and the nutrient pool in mature leaves was weak [statement (a)]. Furthermore, only a weak relationship was found between the cumulative resorption of nutrients and the difference between the nutrient pools of mature and abscissed leaves [statement (b)]. We can argue that the weak correlations are a reflection of a high variability introduced by the low sampling density and, therefore, do not reflect the validity of statements (a) and (b). Although a higher sampling density at all 18 sampling times resulted in a reduction of the variability, the evidence supporting statements (a) and (b) remained weak (Table 2).
The seasonal flux for leaching [statement (c)] indicated nutrient loss from the foliage for 70% of the simulations for N and 100% of simulations for P in Hämeenkyrö, and 80% of the simulations for N and 100% of the simulations for P in Harjavalta. Consequently, leaching was not symmetrically centred around zero and there was strong evidence to reject the statement that leaching equalled zero. Leaching (Table 3) was low compared to foliar accretion and resorption in Hämeenkyrö and Harjavalta (Table 4). Not taking leaching into account in the estimation of accretion and resorption resulted in both under- and overestimations of foliar accretion, resorption and proportional resorption. The difference in PR was significant (P<0.05) for P in Hämeenkyrö, and for N in Harjavalta (Table 4).
3.5 Does the commonly used estimator for PR preserve differences between plots?
The commonly used estimator is often applied in comparative ecological studies for testing ecological theory, i.e. is the proportional resorption lower on fertile plots than on infertile plots? Despite its shortcomings, the commonly used estimator might be a valuable parameter in comparative ecological studies if it preserves the ranking between the plots. Two plots with a different (t-test, P=0.000 for N and P=0.000 for P) PR for N and P were investigated in this study. The ranking was preserved for 81% of the comparisons for N and for 53% of the comparisons for P. When the precision of the PR was increased by calculating the PR from a higher sampling density at all 18 sampling times, the ranking was preserved for 97% and 72% of the comparisons for N and P, respectively. However, preservation of the ranking relationship was not necessarily caused by a correct ranking of the underlying processes. Between 60% and 88% of the cases preserved the correct ranking for the accretion, while 47–59% of the cases preserved the ranking for resorption.
4 Discussion
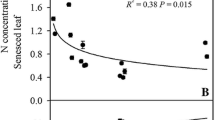
The commonly used estimator was found to be inaccurate, as the PR for N was, on the average, underestimated by 3–10% and the P resorption by 20–25%. Consequently, care should be taken when interpreting accretion, resorption and proportional resorption based on the commonly used estimator. Some authors, for example Côté et al. (2002), further simplify the commonly used estimator and do not account for seasonal changes in the foliar and leaf litter biomass which leads to a further underestimation of the PR by 20% (van Heerwaarden et al. 2003). Inaccuracy of the commonly used estimator is caused by a lack of selectiveness, since the commonly used estimator does not respond to the processes, it is expected to respond to, i.e. accretion and resorption. The lack of selectiveness came as a surprise, because the assumptions that make the calculation of commonly used estimator less demanding than that of best available estimator are, at first sight, reasonable. Using the commonly used estimator assumes that the cumulative accretion relates to the maximum nutrient pool in mature leaves, and that the cumulative resorption relates to the differences in the nutrient pool between abscissed and mature leaves. Only weak evidence was found in support of the assumed relationships, which illustrates that the cumulative accretion and resorption cannot be approximated by the nutrient pools before and after abscission. First, the nutrient pool in mature leaves does not quantify those nutrient losses which were compensated for by nutrient accretion. For example, the canopy P pool decreased between t and t+1 due to litterfall. As a means for understanding the shortcoming of the commonly used estimator, we assumed that this loss was completely compensated for by an increased P concentration in the foliage such that the P pool in mature leaves remained unchanged between t and t+1. However, the cumulative accretion at t+1 increased with an amount similar to the P losses due to litterfall. Therefore, the nutrient pool in mature leaves underestimates the cumulative accretion (Fig. 4; similar graphs for Harjavalta, not given). Although the results in the present study are based on only two plots, the shortcoming of the nutrient pool as an estimator of the cumulative accretion is valid whenever the plant compensates for nutrient losses due to foliar leaching and litterfall. Our data of even-aged B. pendula stands in the southern boreal coniferous zone (Fig. 2) and data for an uneven-aged stand dominated by sugar maple (Acer saccharum Marsh.) in association with yellow birch (B. alleghaniensis Britt.) and American beech (Fagus grandifolia Ehrh.) in the northern hardwood zone (Duchesne et al. 2001), show that losses due to foliar leaching and litterfall are compensated for, by accretion during the whole growing season. Second, the commonly used estimator estimates the cumulative resorption as the difference in nutrient pools between mature and abscissed leaves. The nutrient pool in abscissed leaves can be correctly estimated by multiplying the weighted average nutrient concentration in leaf litter with the total biomass of leaf litter. However, the commonly used estimator of resorption is based on the commonly used estimator for accretion. As the commonly used estimator for accretion is flawed, the commonly used estimator for resorption must also be flawed. Finally, the commonly used estimator does not use deposition and throughfall data, and consequently assumes that there is no nutrient leaching from the foliage. Although leaching occurs and its flux differs from zero, the flux is low compared to the accretion and resorption fluxes and, in general, its omission did not change the estimation of cumulative accretion and resorption. Nevertheless, omission of the leaching flux was shown to have a potential for changing the ratio between resorption and accretion and thus the estimation of the PR.

Comparison of the best available estimator of the cumulative accretion (solid line) and the commonly used estimator of the cumulative accretion which is the nutrient pool (dotted line). The error bars show the 95% CI. Graphs are based on the low sampling density. At the end of the growing season, the nutrient pool drops to zero because the foliar biomass drops to zero
The precision of an estimator depends on the heterogeneity of the processes under study and on the sampling density, which is the ratio of the total size of the entity under study to the product of the sampling density and sample support (Luyssaert et al. 2003). The precision for the commonly used estimator was especially low for P (11–13%) when the sampling density was low, i.e. the sampling density was limited to one replicate sample at each of the 18 sampling times with a sample support as specified in the methods and material section. It is difficult to judge whether this low sampling density represented a commonly used sampling density, because very few studies report the number of replicate samples taken, the support of the samples and the total size of the entity under study. Compared to the sampling density used among others, Killingbeck (1985), and Norby et al. (2000), our settings for the low sampling density could be considered to be realistic. Consequently, the high variability for the commonly used estimator for P, as reported in literature reviews (Aerts 1996; Aerts and Chapin 2000) and the high variation in the difference between the measured resorption and the potential resorption (Killingbeck et al. 1990; Killingbeck 1996) could be an artefact of the used estimator and sampling density instead of a reflection of a highly variable physiological process. However, we have no direct evidence for this. By using a higher sampling density, i.e. 30 foliar (for ten trees one from the bottom, one from the middle and one from the top of the crown), three throughfall, three stemflow, three bulk precipitation and six observations for leaf litterfall for each of the 18 time steps, the precision of the commonly used estimator can be highly improved, which is an advantage when comparing the PR between plots and between years. A high precision ranging between 2.6% and 6.3% for N and P was obtained for the high sampling density.
Although the commonly used estimator is unselective, inaccurate and in general imprecise, it was found to be a useful parameter for comparing the PR between plots, as it preserved the ranking of the best available estimator of the PR for different plots well. Without paying special attention to the sampling program, there is a chance of 20% that the order is lost for N and of 50% that it will happen for P. We hypothesize that some of the conflicting results in relation to PR as an adaptive strategy of plants to their growing environment could have been caused by the loss of the order between plots in different growing environments. However, when the sampling density is increased, there is only a 3% chance that the order is lost for N and a 29% chance that this will happen for P. These favourable results were not accompanied by order preservation of the accretion and resorption. Nevertheless, as the best available estimator for both plots is rather similar and the order is well preserved, we suggest that order preservation is a more general characteristic of the commonly used estimator.
4.1 Conclusive remarks
As the calculations for both N and P are based on the same sampling, the more important underestimation of the proportional P resorption obtained using the commonly used estimator is due to different P dynamics during the course of the vegetation period compared to N. Although leaching occurs, the flux is low compared to the accretion and resorption fluxes and its omission did not change the estimation of cumulative accretion and resorption. The weakness of the commonly used estimator is caused by the use of the nutrient pool in mature leaves as an approximation of the cumulative accretion (Fig. 4). The nutrient pool in mature leaves would be a good approximation when accretion and resorption follow each other in time. Our data of even-aged silver birch stands in the southern boreal coniferous zone and data of uneven-aged stand dominated by sugar maple in the northern hardwood zone (Duchesne et al. 2001) show that accretion and resorption are not separated in time, but occur both throughout the growing season (Fig. 2). We do not recommend that our quantitative results be taken literally, as they are based on only two sets of observations. However, the results show that the commonly used estimator should no longer be used as a measure for accretion, resorption or PR whenever accretion and resorption are not separated in time. Nevertheless, when a sufficiently high sampling density is ensured, the commonly used estimator can be used in comparative studies as an index for PR only.
References
Aerts R (1996) Nutrient resorption from senescing leaves of perennials: are there general patterns? J Ecol 84:597–608
Aerts R, Chapin FS (2000) The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Adv Ecol Res 30:1–67
Ahti T, Hämet-Ahti L, Jalas J (1968) Vegetation zones and their sections in northwestern Europe. Ann Bot Fenn 5:169–211
Anderson HV, Hovmand MF (1999) Review of dry deposition measurements of ammonia and nitric acid to forest. Forest Ecol Manag. DOI 10.1016/S0378-1127(98)00378-8
Asner GP, Townsend AR, Riley WJ, Matson PA, Neff JC, Cleveland CC (2001) Physical and biogeochemical controls over terrestrial ecosystem responses to nitrogen deposition. Biogeochemistry. DOI 10.1023/A:1010653913530
Beier C (1991) Separation of gaseous and particulate dry deposition of sulfur at a forest edge in Denmark. J Environ Qual 20:460–466
Côté B, Fyles JW, Djalivand H (2002) Increasing N and P resorption efficiency and proficiency in northern deciduous hardwoods with decreasing foliar N and P concentrations. Ann For Sci. DOI 10.1051/forest:2002023
Chapin FS (1980) The mineral nutrition of wild plants. Annu Rev Ecol Syst 11:233–260
Chapin FS, Kedrowski RA (1983) Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees. Ecology 64:376–391
Duchesne L, Ouimet R, Camiré C, Houle D (2001) Seasonal nutrient transfer by foliar resorption, leaching, and litterfall in a northern hardwood forest at Lake Clair Watershed, Qubec, Canada. Can J For Res 31:333–344
Eckstein RL, Karlsson PS, Weih M (1999) Leaf life span and nutrient resorption as determinants of plant nutrition conservation in temperate-artic regions. New Phytol. DOI 10.1046/j.1469–8137.1999.00429.x
Efron B, Tibshirani RJ (1993) An introduction to the bootstrap. Chapman & Hall, New York
Franklin O, Agren GI (2002) Leaf senescence and resorption as mechanisms of maximizing photosynthetic production during canopy development at N limitation. Funct Ecol 16:727–733
van Heerwaarden LM, Toet S, Aerts R (2003) Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: facts and solutions. Oikos 101:664–669
Helmisaari HS (1992) Nutrient retransloctaion within foliage of pinus sylvestris. Tree Physiol 10:45–58
Kellomäki S, Rouvinen I, Peltola H, Strandman H (2000) Density of foliage mass and area in the boreal forest cover in Finland, with applications to the estimation of monoterpene and isoprene emissions. Atmos Environ. DOI 10.1016/S1352-2310(00)00309-5
Kellomäki S, Rouvinen I, Peltola H, Strandman H, Steinbrecher R (2001) Impact of global warming on the tree species composition of boreal forests in Finland and effects on emissions of isoprenoids. Global Change Biol. DOI 10.1046/j.1365-2486.2001.00414.x
Kiikkilä O (2003) Heavy-metal pollution and remediation of forest soil around the Harjavalta Cu–Ni Smelter in SW Finland. Silva Fenn 37:399–415
Killingbeck KT (1985) Autumnal resorption and accretion of trace metals in gallery forest trees. Ecology 66:283–286
Killingbeck KT (1996) Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency. Ecology 77:1716–1727
Killingbeck KT, May JD, Nyman S (1990) Foliar senescence in aspen (Populus tremuloides) clone: the response of element resorption to interramet variation and timing of abscission. Can J For Res 20:1156–1164
Knops JMH, Koenig WD, Nash III TH (1997) On the relationship between nutrient use efficiency and fertility in forest ecosystems. Oecologia 10:550–556. DOI 10.1007/s004420050194
Luyssaert S, Mertens J, Raitio H (2003) Support, shape and number of replicate samples for tree foliage analysis. J Environ Monit. DOI 10.1039/b210727a
May JD, Killingbeck KT (1992) Effects of preventing nutrient resorption on plant fitness and foliar nutrient dynamics. Ecology 73:1868–1878
Norby RJ, Cotrufo MF (1998) A question of litter quality. Nature. DOI 10.1038/23812
Norby RJ, Long TM, Hartz-Rubin JS, O’Neill EG (2000) Nitrogen resorption in senescing tree leaves in a warmer, CO2-enriched atmosphere. Plant Soil. DOI 10.1023/A:1004629231766
Nordell KO, Karlsson PS (1995) Resorption of nitrogen and dry-matter prior to leaf abscission—variation among individuals, sites and years in the Mountain Birch. Funct Ecol 9:326–333
Oland K (1963) Changes in the content of dry matter and major nutrient elements of apple foliage during senescence and abscission. Physiol Plantarum 16:682–694
Oleksyn J, Reich PB, Zytkowiak R, Karolewski P, Tjoelker MG (2003) Nutrient conservation increases with latitude of origin in European Pinus sylvestris populations. Oecologia. DOI 10.1007/s00442-003-1265-9
Saur E, Nambiar E, Fife D (2000) Foliar nutrient retranslocation in Eucalyptus globus. Tree Physiol 20:1105–1112
Staaf H (1982) Plant nutrient changes in beech leaves during senescence as influenced by site characteristics. Acta Oecol 3:161–170
Ulrich B (1983) Interaction of forest canopies with atmospheric constituents: SO2, alkali and earth alkali cations and chloride. In: Ulrich B, Pankrath J (eds) Effects of accumulation of air pollutants in forest ecosystems. Reidel, Dordrecht, pp 33–45
Wright IJ, Westoby M (2004) Nutrient concentration, resorption and lifespan: leaf traits of Australian sclerophyll species. Funct Ecol 17:10–19
Acknowledgements
Sebastiaan Luyssaert was funded by a Marie Curie Individual Fellowship (QLK-CT-2001–51941), Jeroen Staelens was funded as a Research Assistant of the Fund for Scientific Research (F.W.O.), Flanders, Belgium. The authors wish to thank Timo Salmi from the Finnish Meteorological Institute for providing unpublished deposition data. The laboratory and field staff, Ghislaine Dossou and Toni Tunturi are thanked for their help with many practical issues. The experiments comply with the current laws of Finland.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Christian Koerner
Rights and permissions
About this article
Cite this article
Luyssaert, S., Staelens, J. & De Schrijver, A. Does the commonly used estimator of nutrient resorption in tree foliage actually measure what it claims to?. Oecologia 144, 177–186 (2005). https://doi.org/10.1007/s00442-005-0085-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-005-0085-5