
Abstract
Strigolactones have recently been identified as the long sought-after signal required to inhibit shoot branching (Gomez-Roldan et al. 2008; Umehara et al. 2008; reviewed in Dun et al. 2009). Here we briefly describe the evidence for strigolactone inhibition of shoot branching and, more extensively, the broader context of this action. We address the central question of why strigolactone mutants exhibit a varied branching phenotype across a wide range of experimental conditions. Where knowledge is available, we highlight the role of other hormones in dictating these phenotypes and describe those instances where our knowledge of known plant hormones and their interactions falls considerably short of explaining the phenotypes. This review will focus on bud outgrowth in herbaceous species because knowledge on the role of strigolactones in shoot branching to date barely extends beyond this group of plants.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Regulation of bud outgrowth
In different species, and also different positions along the plant stem, axillary buds exhibit differing degrees of development and outgrowth during the vegetative phase. Along the main shoot axis in pea (Pisum sativum), axillary meristems develop into buds with immature leaves at most nodes, and the outgrowth of these buds typically remains arrested (Stafstrom and Sussex 1988, 1992; Stafstrom 1993). In Arabidopsis (Arabidopsis thaliana), axillary buds form late during vegetative growth and are generally suppressed until the switch to flowering (Grbić and Bleecker 2000). In monocots such as rice (Oryza sativa), the characteristic growth habit comprises the release of many axillary buds to form tillers, while in other monocot species such as corn (Zea mays) the outgrowth of axillary buds remains relatively restricted (McSteen 2009).
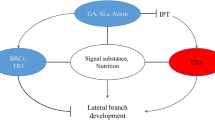
Despite the diversity in growth habits, bud outgrowth among herbaceous species is a multifactorial process determined largely by the same factors. In addition to the classical hormones auxin and cytokinin, and the recently identified strigolactone compound (Gomez-Roldan et al. 2008; Umehara et al. 2008), other external and internal cues also contribute to spatial and temporal regulation of bud outgrowth. Together, these are thought to impose three distinct states of dormancy: (1) endodormancy, by factors acting within the bud, (2) paradormancy, by signals within the plant but external to the bud, and (3) ecodormancy, by environmental factors external to the plant (Lang et al. 1987). Interpretation of multiple signals within buds may be accomplished, at least in part, by the existence of distinct stages at which different signals might act (Fig. 1; Dun et al. 2006). For example, numerous studies demonstrate that the transition of axillary buds from dormancy to growth is likely to comprise an intermediate phase of altered metabolic activity. This transition phase is characterised by increased cellular activity compared to cells within dormant buds, as evidenced by gene and protein expression levels and even small increases in bud size (Stafstrom and Sussex 1988, 1992; Madoka and Mori 2000; Shimizu-Sato and Mori 2001; Morris et al. 2005; Shimizu-Sato et al. 2008). Importantly, buds undergoing this transition are capable of cycling back and forth between dormancy and growth (Stafstrom and Sussex 1988; Devitt and Stafstrom 1995; Shimizu and Mori 1998; Napoli et al. 1999; Shimizu-Sato and Mori 2001). This may provide a convenient framework in which to consider multiple branching inputs, where factors such as light, nutrient availability, the age and position of the bud and the plant’s developmental stage may determine the ability of individual buds to respond to other long-distance signals such as auxin, cytokinin and strigolactone.

Stages of bud outgrowth. Buds may progress (bold arrows) through different growth states depending on different regulatory factors (standard arrows). Various cues including positional and developmental information can cause dormant buds (1) to enter a stage of transition, characterised by altered metabolic activity. (Stafstrom and Sussex 1988; Devitt and Stafstrom 1995; Shimizu and Mori 1998; Napoli et al. 1999; Shimizu-Sato and Mori 2001). Buds may then become responsive (2) to regulatory factors such as stem auxin level, which may be influenced by florigen signalling (Shalit et al. 2009). Auxin acts to independently up-regulate strigolactones and down-regulate cytokinin level. Auxin and strigolactones inhibit the progression to growth while cytokinins promote this progression. If a bud is induced to grow (3), auxin transported from the bud into the PATS acts to feedback up-regulate strigolactone synthesis and attract nutrient import. Depending on nutritional status and hormonal interactions a growing bud will enter into sustained growth (4) and can inhibit the growth of additional buds by correlative inhibition (Morris 1977; Li and Bangerth 1999)
Hormone interactions and functions
Strigolactones are carotenoid-derived terpenoid lactones first characterised as germination stimulants of the parasitic plant genera Striga and Orobanche, and as hyphal branching regulators in arbuscular mycorrhizal associations (Cook et al. 1972; Matusova et al. 2005; Akiyama et al. 2005; Humphrey and Beale 2006; López-Ráez et al. 2008). Strigolactones, or a derivative thereof, have recently been identified as the long-distance branching inhibitor regulated by the RMS (RAMOSUS) pathway in pea, MAX (MORE AXILLARY GROWTH) pathway in Arabidopsis, and D/HTD (DWARF/HIGH TILLERING DWARF) pathway in rice (Gomez-Roldan et al. 2008; Umehara et al. 2008). Mutants for these pathways that are defective in the production, but not the perception, of the branching inhibitor have reduced strigolactone levels. Furthermore, the increased branching in the synthesis mutants, but not the perception mutants, can be repressed by addition of the strigolactone analogue, GR24 (Gomez-Roldan et al. 2008; Umehara et al. 2008). As small, potent molecules required for communication within the plant body and between plants and the environment, strigolactones therefore fit the bill as a new class of phytohormone (Klee 2008; Leyser 2008; Dun et al. 2009).
The hormones auxin and cytokinin are also essential components of the network controlling branching in plants. Auxin transported downwards from the shoot tip in the polar auxin transport stream (PATS) indirectly represses bud outgrowth, while cytokinin promotes cell division and differentiation and bud outgrowth (reviewed in Kyozuka 2007; Shimizu-Sato et al. 2009). A number of studies have begun to elucidate how these hormones and strigolactones may specifically interact to regulate bud dormancy.
Auxin has been shown to promote the expression of strigolactone biosynthesis genes encoded by MAX3 and MAX4 in Arabidopsis, RMS5 and RMS1 in pea, and HTD1 and D10 in rice (Foo et al. 2005; Bainbridge et al. 2005; Johnson et al. 2006; Zou et al. 2006; Arite et al. 2007; Hayward et al. 2009). In Arabidopsis, this regulation is dependent on AXR1 (AUXIN RESISTANT1) (Bainbridge et al. 2005; Hayward et al. 2009). AXR1 is required for the optimal activity of SCF (Skp1-Cul1/Cdc53-F-box) E3 ubiquitin ligase complexes including the auxin receptor SCFTIR1 (TRANSPORT INHIBITOR1), and auxin binding to SCFTIR1 induces the degradation of Aux/IAA transcriptional repressors (reviewed in Mockaitis and Estelle 2008). In Arabidopsis, recent data has provided evidence that this transcriptional interaction between auxin and strigolactone synthesis genes is required, at least in part, for auxin-mediated branching inhibition. The auxin response mutant bodenlos, which has a lesion in domain II of the Aux/IAA transcriptional repressor IAA12, has very low MAX3 and MAX4 expression and increased branching that is dependent on strigolactone supply (Hayward et al. 2009). The increased branching in additional auxin response mutants can also be suppressed by strigolactone application (Brewer et al. 2009).
While the above studies demonstrate auxin may regulate strigolactone levels, this interaction is not sufficient to completely explain auxin-mediated branching inhibition (Bennett et al. 2006; Hayward et al. 2009), which also likely involves auxin-mediated repression of cytokinin levels (reviewed in Shimizu-Sato et al. 2009). In pea stems, auxin inhibits the expression of ISOPENTENYL TRANFERASE (IPT) cytokinin biosynthesis genes and promotes the expression of a CYTOKININ OXIDASE (CKX) gene involved in cytokinin breakdown (Tanaka et al. 2006; Shimizu-Sato et al. 2009). In Arabidopsis, cytokinin levels are negatively regulated by auxin in an AXR1-dependent manner (Nordström et al. 2004). Thus a transcriptional interaction via the SCFTIR E3 ubiquitin ligase signalling cascade may control both strigolactone and cytokinin levels. On the other hand, a functional interaction between strigolactones and cytokinin is yet to be clarified and evidence so far suggests they may act partly independently, as discussed further below (Bennett et al. 2006; Ongaro et al. 2008; Ferguson and Beveridge 2009; Leyser 2009). Nonetheless, it is known that low strigolactone signalling in the shoot leads to decreased cytokinin content in the root xylem sap (Foo et al. 2007) and, in Arabidopsis, exogenous cytokinin can prevent the up-regulation of strigolactone synthesis gene expression by auxin in the root (Bainbridge et al. 2005).
Currently, the mechanism of strigolactone action in branching inhibition is uncertain. Strigolactone perception or signal transduction requires localised action of an F-box protein within axillary buds or the adjacent nodal stem, encoded by MAX2 in Arabidopsis, RMS4 in pea and D3 in rice (Stirnberg et al. 2002, 2007; Ishikawa et al. 2005; Johnson et al. 2006; Gomez-Roldan et al. 2008; Umehara et al. 2008). Therefore, strigolactone perception via an SCF complex, similarly to auxin and SCFTIR1, may target additional downstream branching regulators for ubiquitination and degradation by the 26S proteasome (Stirnberg et al. 2007). In Arabidopsis, a gene thought to act downstream of the MAX pathway is BRANCHED1 or TEOSINTE BRANCHED-LIKE1 (BRC1 or TBL1), identified based on its similarity to TEOSINTE BRANCHED1 (TB1) in maize and its orthologue, OsTB1/FC1 (FINE CULM1), in rice (Doebley et al. 1997; Hu et al. 2003; Takeda et al. 2003; Aguilar-Martínez et al. 2007). TB1 encodes a member of the TCP (TB1, CYCLOIDEA, PCF domain) transcription factor family, and TB1 is required in buds to inhibit outgrowth. In Arabidopsis, BRC1 expression is down-regulated in max mutant buds, and double mutants for brc1 and the max genes show similar branching phenotypes as the single mutants, suggesting they act on the same pathway (Aguilar-Martínez et al. 2007; Finlayson 2007). BRC1 is also thought to act partly independently of cytokinin to control branching (Aguilar-Martínez et al. 2007), consistent with some distinct aspects of strigolactone- and cytokinin-mediated branching control (Leyser 2009). In rice, however, evidence so far suggests that FC1 may act independently from strigolactone, as FC1 expression is not altered in strigolactone pathway mutants (Arite et al. 2007).
Two hypotheses have been presented for the ultimate output of strigolactone signalling. The first is that strigolactones modulate the ability for buds to establish an auxin flow into the PATS of the primary stem (Bennett et al. 2006; reviewed in Ongaro and Leyser 2008). This is based on the premise that axillary meristems require auxin-transporting vascular connections to the main stem vasculature, canalised by auxin flow and driven by source-sink relationships, in order to function and grow (reviewed in Ongaro and Leyser 2008; Leyser 2009). Accordingly, auxin transport and/or level is increased in strigolactone pathway mutants of Arabidopsis and rice, and where tested, branching in these mutants can be reduced by applying polar auxin transport inhibitors (Bennett et al. 2006; Arite et al. 2007; Lin et al. 2009). The second hypothesis for strigolactone action builds upon the auxin regulation of strigolactone synthesis and suggests that strigolactones function downstream of auxin to inhibit branching as second messengers (reviewed in Dun et al. 2009). In accordance with this hypothesis, GR24 applied to buds can inhibit bud outgrowth in auxin-depleted stems of decapitated pea plants (Brewer et al. 2009). Here, auxin transport from buds is proposed to be important for continued growth, rather than the initial release of buds, and the increased auxin content often observed in the mutants is suggested to be a result of bud outgrowth and/or feedback up-regulation (Dun et al. 2006, 2009).
Auxin and strigolactone may also interact to control branching as part of a feedback process. Recently it was demonstrated in Arabidopsis shoots that the increased auxin content of strigolactone pathway mutants acts to feedback up-regulate the expression levels of strigolactone synthesis genes (Hayward et al. 2009). Auxin and strigolactone are therefore likely to be linked in a feedback loop, where low strigolactone leads to increased auxin content, which in turn up-regulates strigolactone synthesis (Hayward et al. 2009; Fig. 1).
Developmental and positional effects
The position or age of a bud along the stem can determine its ability to grow out. During vegetative development, bud outgrowth tends to occur in an acropetal wave before shifting to a basipetal wave during the reproductive phase (McSteen and Leyser 2005). This pattern of branch development broadly defines three distinct zones in the main stem: (1) an upper zone, where increased bud outgrowth accompanies flowering, (2) a middle zone directly beneath the upper zone where bud outgrowth is typically reduced, and (3) a basal zone (Weberling 1989; Napoli et al. 1999). In pea, the bud at node 2 within the basal zone is particularly responsive to outgrowth following the release of apical dominance by decapitation or girdling (discussed below; Ferguson and Beveridge 2009). Buds at these basal nodes are also very responsive to photoperiod, having a greater tendency for outgrowth under short days (Beveridge et al. 2003). Cytokinin application to buds at the first four nodes only promotes branching in nodes 1 and 2 in intact plants; decapitation is additionally required to trigger the outgrowth at the upper nodes (King and Van Staden 1988). Buds at lower nodes, particularly node 2 may possess particularly well-established vascular connections (Ferguson and Beveridge 2009).
Auxin is one candidate for mediating some of these positional effects (reviewed in McSteen and Leyser 2005). As described above, it has been proposed that vascular connectivity and the ability of buds to plumb into the PATS is critical for outgrowth (Ongaro and Leyser 2008; Ongaro et al. 2008). The maintenance of bud outgrowth also appears to correlate with the ability of buds to synthesise auxin de novo and export it into the stem, even in cases where buds already have well-developed vascular connections (Morris 1977; Morris and Johnson 1990; Li and Bangerth 1999). Furthermore, auxin movement may also determine the ability of a bud to attract nutrients to support sustained outgrowth (Davies and Wareing 1965; Phillips 1968; Jiang et al. 2001; Yang et al. 2007; Fig. 1).
Three-dimensional positioning can also influence axillary branching. When plants are placed horizontally, bud outgrowth occurs along the upper stem, and when plants are completely inverted, the most basal bud grows out (Cline and Riley 1984; Prasad and Cline 1985). Likewise, when only the upper part of the main stem is bent downwards, the axillary bud at the node closest to the bend point is induced to grow. This response is likely to be mediated by gravity because the agravitropic weeping (we) mutants in Japanese morning glory (Pharbitis nil) exhibit a reduced lateral outgrowth response of buds to stem bending (Kitazawa et al. 2008). This gravity-induced release of axillary buds appears to act independently of auxin and cytokinin, and possibly independently of strigolactone, as auxin and cytokinin levels do not change in accordance with shoot bending. The we mutants also possess a normal decapitation response that can be rescued by auxin application. A gravity-mediated mechanism for bud outgrowth further highlights the ability of plants to adapt their growth pattern in order to cope with changing conditions.
Correlative inhibition
For branching to be optimised in the context of a growing, healthy shoot system, it must be carefully modulated. In branching terms, correlative inhibition refers to the ability of one branch to suppress the growth of other axillary shoots, and this has been associated with the active transport of auxin from the growing branch (Morris 1977; Li and Bangerth 1999). In pea, where more than one dormant bud may exist at each node, the largest bud will generally be induced to grow, suppressing the growth of the accessory buds (reviewed in Shimizu-Sato and Mori 2001). In rms mutants, growing basal axillary buds can inhibit the outgrowth of aerial buds and accessory buds in the same node, suggesting correlative inhibition acts at least partly independently from the strigolactone pathway (Morris et al. 2005; Ferguson and Beveridge 2009). Using a two-branched system in Arabidopsis, correlative inhibition was found to comprise both MAX-pathway dependent and independent components that can be transmitted unilaterally across the stem (Ongaro et al. 2008). Interestingly, double mutants for the aforementioned rms genes and rms2, which is not yet cloned but causes increased xylem cytokinin content, display outgrowth of previously inhibited accessory buds compared to single rms mutants (Beveridge et al. 1997; Murfet and Symons 2000a, b), suggesting the RMS2 gene product and/or cytokinin may also be involved in correlative inhibition independent from strigolactones.
Decapitation
Activities which cause loss of the shoot apex, such as pruning and herbivory, can lead to the activation of dormant axillary buds, enabling continued shoot growth and reproduction. This is particularly important in species that show a strong apical dominance phenotype (where lateral shoot growth is normally suppressed). Loss of auxin produced by these apices is central to this response, resulting in increased sink strength of the main stem PATS for bud-derived auxin, reduced strigolactone production and increased cytokinin biosynthesis and levels (Sachs and Thimann 1964, 1967; Bangerth 1994; Nordström et al. 2004; Tanaka et al. 2006; Brewer et al. 2009; Hayward et al. 2009). Nonetheless, in pea, the outgrowth of the bud at node 2 is rapidly induced by decapitation in a manner that does not correlate with the reductions in auxin level and strigolactone synthesis gene expression in the stem (Morris et al. 2005; Ferguson and Beveridge 2009). Decapitation itself may trigger a physical signal (such as an electrochemical pulse), which is transmitted along the main stem to buds at every node to induce an initial growth response (Morris et al. 2005; Ferguson and Beveridge 2009). The enhanced ability of the bud at node 2 to respond to this signal and subsequently grow may be due its well-developed vascular connectivity. Such connections are essential for meristems to export auxin and receive nutrients, as previously described. Indeed, physical re-directions in assimilates, minerals, endogenous cytokinins and water flow from the shoot apex to axillary buds occur rapidly after loss of the shoot apex, as well as alterations in photosynthate supply and source-sink relations (Husain and Linck 1966; Stahlberg and Cosgrove 1992; Turnbull et al. 1997). In addition, cytokinin is metabolised in buds soon after decapitation (Mader et al. 2003).
Further evidence for an auxin-independent effect of decapitation on bud outgrowth is that branching at certain nodes of pea is not always induced when stem auxin levels are depleted by girdling or inhibiting polar auxin transport, unless plants are simultaneously decapitated (Morris et al. 2005; Ferguson and Beveridge 2009). For example, buds at node 2 of pea remain dormant even when the plant is girdled in the internode directly above (Ferguson and Beveridge 2009). In these girdled plants, reduced branching responses are associated with low expression of IPT cytokinin synthesis genes in the vicinity of the bud, suggesting that the regulation of cytokinin by factors independent from polar auxin transport could be an additional determining factor for bud outgrowth (Ferguson and Beveridge 2009). Accordingly, cytokinin application to these buds restores outgrowth (Ferguson and Beveridge 2009). In pea rms mutants and in the rice d10 mutant, treatments that disrupt the flow of auxin from the shoot tip cause an additive branching response (Beveridge 2000; Arite et al. 2007; Ferguson and Beveridge 2009). Again this suggests that auxin inhibits branching via strigolactone-dependent and independent pathways. Buds in different locations may also be controlled by partially distinct genetic pathways, as shown in the monocot, foxtail millet (Setaria italica), where the branches produced in the basal nodes are under separate control to those produced in the upper nodes (Doust et al. 2004; Doust 2007).
Despite an initial growth response following decapitation, the sustained growth of buds in pea and many species is reliably inhibited by exogenous auxin applied to the decapitation site (Thimann and Skoog 1933, 1934; Cline 1996; Cline et al. 2001). In flowering Arabidopsis, however, auxin cannot prevent the outgrowth of buds following decapitation except in the axr3 increased auxin-response mutant and in isolated cauline nodes lacking a root system (Cline 1996, 2000; Cline et al. 2001; Ouellet et al. 2001). In these isolated nodes, basally supplied cytokinin can counteract the inhibitory effect of apically supplied auxin on bud outgrowth (Chatfield et al. 2000). Perhaps in different species, differences in the relative levels or influence (response) of branching hormones in the context of diverse developmental programs and environmental signals may be a source of varied branching responses. In species with short life-cycles such as Arabidopsis, a strong response to decapitation after the transition to reproductive growth may be particularly important for the completion of the life-cycle.
Light
The length and quality of light that plants are exposed to can alter their branching patterns. As mentioned above, under short day conditions, branching is increased at basal nodes in pea (Beveridge et al. 2003). Similar branching patterns are also observed in Arabidopsis and petunia (Petunia hybrida) (Grbić and Bleecker 2000; Stirnberg et al. 2002; Snowden and Napoli 2003). Unlike WT plants, intact die neutralis (dne) mutants in pea lack the ability to produce basal branches under short days, suggesting that the DNE gene product is probably required to mediate photoperiod responsiveness in basal axillary buds. The branching pattern of rms mutants is dependent on photoperiod, as the basal branching habit of rms1 and rms2 mutants under short days is shifted towards a relatively more aerial pattern of branching under long days (Beveridge et al. 2003). Reduced outgrowth of buds at basal nodes under long days is also observed in decapitated WT plants, suggesting a similar pathway mediates this photoperiod response (Beveridge et al. 2003).
The ratio of red to far-red light (R:FR) is critical in the light reception response. The relative amount of FR light is greatly enhanced when light conditions are low (i.e. canopy shading), or when light is reflected (i.e. crowding or increased plant density). High FR levels alter phytochrome activity and initiate light responses, particularly phytochrome B, which acts in the shade response (Robson et al. 1993). As a consequence, the plant’s resources are invested into growth of the main shoot axis, including the hypocotyl, at the expense of bud outgrowth and leaf growth (Cerdán and Chory 2003; Casal et al. 2004; Doust 2007; Kebrom and Brutnell 2007; Carabelli et al. 2007).
Recent work has demonstrated interactions and overlap between strigolactone- and light-mediated branching control. In Arabidopsis, the MAX2 F-box gene has been identified independently in three mutant screens for delayed leaf senescence, increased branching and photomorphogenesis regulation, respectively (Woo et al. 2001; Stirnberg et al. 2002; Shen et al. 2007). In rice, the D3 F-box is also involved in leaf senescence (Yan et al. 2007). The pleiotropic phenotypes of max2 mutants are indicative of multiple roles for MAX2 in plant development and consistent with a possible function for the nuclear-localised F-box in regulating multiple signalling cascades (Stirnberg et al. 2007). MAX2 expression is induced by light in WT seedlings, and max2 mutant seedlings are hyposensitive to R and FR light in a phytochrome-dependent manner (Shen et al. 2007). However, MAX2 and phyB appear to have antagonistic roles in older plants in branching regulation, as phyB mutants display small reductions in branch number and MAX2 is epistatic to phyB in double mutants which possess a max2 increased-branching phenotype (Reed et al. 1993; Shen et al. 2007). These findings suggest MAX2 could play different roles in the plant depending on developmental stage.
In Sorghum bicolor, increased apical dominance in a phyB mutant is accompanied by increased expression of the TB1 orthologue SbTB1 (Kebrom et al. 2006). Thus, low R:FR ratios leading to inactive phyB (or non-functional phyB in a mutant), results in maintenance of SbTB1 expression and bud outgrowth is inhibited. Further to this, expression of BRC1 in Arabidopsis is increased under crowded conditions, though it is not known for certain if the crowding effect is specifically related to reduced R:FR ratios (Aguilar-Martínez et al. 2007). Together, these data suggest TB1 could be an important integrator of light and strigolactone to control bud dormancy in some species.
Another point of cross-talk between light and branching could potentially occur via cytokinin catabolism. As part of shade avoidance response, the arrest of leaf growth is also known to rely on the up-regulation of certain CKX genes under low R:FR ratios (Carabelli et al. 2007). It is possible that a low R:FR ratio might also restrict branching in a similar way by maintaining levels of CKX enzymes. As reviewed in Leyser (2009), much of this relationship between light quality and branching may be mediated by affects of light on auxin fluxes and auxin response.
Flowering
In most annual/model species, the amount of vegetative branching is typically decreased under environmental conditions conducive to flowering, while delayed flowering promotes branching (e.g., Beveridge et al. 2003). In Arabidopsis, while growth under long days promotes flowering and results in less overall rosette branching, rosette bud outgrowth is usually not induced until the switch to flowering. This is possibly due to reduced auxin production by, and increased distance from, the floral apex (McSteen and Leyser 2005).
In monocots and dicots, overexpression or high expression of TERMINAL FLOWER1 (TFL1) usually leads to delayed flowering and increased lateral branching, while tfl1 mutants exhibit early flowering and little to no branching (Ratcliffe et al. 1998; Jensen et al. 2001; Nakagawa et al. 2002; Foucher et al. 2003). In tomato, TFL and FLOWERING LOCUS T (FT) homologues are considered important regulators of vegetative development in their own right (Shalit et al. 2009). The aforementioned circadian clock photoperiod response gene DNE in pea may act via regulation of FT to enhance basal branching under short days (Liew et al. 2009). In sweet pea, where floral induction occurs vary late in development (even in early day-neutral flowering lines) assimilate partitioning and basal branching is regulated by the photoperiod response pathway well before flowering has been induced at the shoot apical meristem (Beveridge et al. 1992). In perennial plants, FT has been strongly implicated in growth cessation and bud endodormancy (reviewed by Horvath 2009). Consequently, evidence from a broad range of species supports the notion that florigen, mediated by FT, can be considered as a general plant hormone affecting vegetative and reproductive development.
Strigolactone mutants do not possess major pleiotropic effects on flowering time or flower number. However, the dad1-1 mutant in petunia flowers later than WT, dad2-1 and dad3 mutants (Napoli 1996; Snowden and Napoli 2003). Nevertheless, interactions between flowering and axillary branching involving florigen, strigolactone and other chemical hormones are beginning to emerge. Overexpression of a gene promoting the vegetative-to-flowering transition, TaVRN1, encoding a MADS-box transcription factor, has been shown to increase axillary bud outgrowth, as well as MAX4 expression (Adam et al. 2007). This raises the possibility that a feedback mechanism(s) may act to coordinate the strigolactone branching pathway with inflorescence development. Recently it was suggested that interactions between auxin and florigen may coordinate sympodial branching, axillary bud release and flowering in tomato (Solanum lycopersicon L.). Mutations in the tomato TFL1 homologue SELF-PRUNING (SP) gene can be mimicked by the PATS inhibitor TIBA (2,3,5-triiodobenzoic acid) and sp mutants possess reduced apical dominance (Pnueli et al. 2001; Shalit et al. 2009). A possible model for the regulated release of axillary buds to form sympodial branches is that a particular ratio between the gene products of SP (an inhibitor of florigen) and SINGLE FLOWER TRUSS (SFT) (florigen) arrests the PATS in localised manner, leading to outgrowth of an adjacent axillary meristem (discussed in Shalit et al. 2009).
Depleted gibberellin and auxin content underlie the reduced internode lengths and increased branching at upper nodes of gigas-1 (gi-1) late flowering pea plants under long days compared to short days (Beveridge et al. 2001). These gi-1 plants likely carry a mutation in one of the FLOWERING LOCUS T-LIKE (FTL) genes in pea (Weller et al. 2009) and exhibit photoperiod-dependent delayed flowering which under long-days is accompanied by a reiterative reduction in vegetative growth commencing at about the time where flower development occurs in WT (Taylor and Murfet 1994; Beveridge et al. 2001; Hecht et al. 2005). In contrast with the gibberellin flowering pathway in Arabidopsis, gibberellin application to pea can only restore the vegetative phenotype back to WT and has no affect on flowering (Beveridge and Murfet 1996).
Nutrients
Nutrient status can undoubtedly affect the ability of shoots to produce lateral branches. As strigolactone acts as a shoot branching inhibitor and a promoter of hyphal branching in mycorrhizal associations, it is possible strigolactone could provide the plant with a means for balancing nutrient levels, particularly phosphorus, with shoot growth (Akiyama et al. 2005; Gomez-Roldan et al. 2008; Umehara et al. 2008; Leyser 2009). Indeed, in rice, tomato and red clover, phosphorous starvation can increase the production of strigolactones (Yoneyama et al. 2007; López-Ráez et al. 2009; Umehara et al. 2008). Finer regulation of branching might also be achieved in combination with auxin and cytokinin, which exhibit nutrient dependent responses either in movement, accumulation or levels (for review, see Rubio et al. 2009). As discussed above, auxin flow in the PATS may influence the movement of nutrients within the plant. Auxin transport can act as a sink for phosphorus (Davies and Wareing 1965). Nitrogen availability is critical for cytokinin synthesis and nitrate has been shown to up-regulate cytokinin levels and IPT gene expression (Takei et al. 2004; Miyawaki et al. 2004). Boron deficiency in pea causes bud outgrowth, probably due to reduced auxin transport out of the apex (Wang et al. 2006). Defoliation of plants cannot inhibit branching in plants triggered by decapitation, though branch length is reduced (Ferguson and Beveridge 2009). This suggests nutrients, specifically carbon sources, could limit the sustained outgrowth of buds but not the initial release of bud dormancy.
Conclusions and perspectives
Like many developmental processes, shoot branching is under multifactorial regulatory control. Even with the discovery of strigolactones, the complexities and number of interactions means we are still far from a solid understanding of branching control in plants. Moreover, in some cases, the hormone interactions are in themselves multiple. For example, auxin may affect strigolactone levels, and strigolactones may affect auxin levels in a feedback role, or more directly, as strigolactone function may involve localised auxin transport. In addition to strigolactone and auxin, cytokinin, nutrient supply and the interaction with light, photoperiod and the induction of flowering are all important. It is likely that branching is induced by changes in different pathways under different conditions and at different developmental stages and that the relative importance of each may vary in different species. Elucidation of the strigolactone signal transduction pathway and the mechanisms of crosstalk in biosynthesis and signalling are likely to yield major progress in understanding branching control in the future. However, the discovery of strigolactones already provides new avenues to consider agricultural and horticultural opportunities such as suppressing an undesirable vegetative flush after pruning.
References
Adam H, Ouellet F, Kane NA, Agharbaoui Z, Major G, Tominaga Y et al (2007) Overexpression of TaVRN1 in Arabidopsis promotes early flowering and alters development. Plant Cell Physiol 48:1192–1206
Aguilar-Martínez JA, Poza-Carrión C, Cubas P (2007) Arabidopsis BRANCHED1 acts as an integrator of branching signals within axillary buds. Plant Cell 19:458–472
Akiyama K, Matsuzaki K, Hayashi H (2005) Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 435:824–827
Arite T, Iwata H, Ohshima K, Maekawa M, Nakajima M, Kojima M et al (2007) DWARF10, an RMS1/MAX4/DAD1 ortholog, controls lateral bud outgrowth in rice. Plant J 51:1019–1029
Bainbridge K, Sorefan K, Ward S, Leyser O (2005) Hormonally controlled expression of the Arabidopsis MAX4 shoot branching regulatory gene. Plant J 44:569–580
Bangerth F (1994) Response of cytokinin concentration in the xylem exudate of bean (Phaseolus vulgaris L.) plants to decapitation and auxin treatment and relationship to apical dominance. Planta 194:439–442
Bennett T, Sieberer T, Willett B, Booker J, Luschnig C, Leyser O (2006) The Arabidopsis MAX pathway controls shoot branching by regulating auxin transport. Curr Biol 16:553–563
Beveridge CA (2000) Long-distance signalling and a mutational analysis of branching in pea. Plant Growth Regul 32:193–203
Beveridge CA, Murfet IC (1996) The gigas mutant in pea is deficient in the floral stimulus. Physiol Plant 96:637–645
Beveridge CA, Ross JJ, Murfet IC (1992) Mutant dn influences dry matter distribution, assimilate partitioning and flowering in Lathyrus odoratus L. J Exp Bot 43:55–62
Beveridge CA, Symons GM, Murfet IC, Ross JJ, Rameau C (1997) The rms1 mutant of pea has elevated indole-3-acetic acid levels and reduced root-sap zeatin riboside content but increased branching controlled by graft-transmissible signal(s). Plant Physiol 115:1251–1258
Beveridge CA, Batge SL, Ross JJ, Murfet IC (2001) Hormone physiology of pea mutants prevented from flowering by mutations gi or veg1. Physiol Plant 113:285–291
Beveridge CA, Weller JL, Singer SR, Hofer JM (2003) Axillary meristem development. Budding relationships between networks controlling flowering, branching and photoperiod responsiveness. Plant Physiol 131:927–934
Brewer PB, Dun EA, Fergusion BJ, Rameau CA, Beveridge CA (2009) Strigolactone acts downstream of auxin to regulate bud outgrowth in pea and Arabidopsis. Plant Physiol 150:482–493
Carabelli M, Possenti M, Sessa G, Ciolfi A, Sassi M, Morelli G, Ruberti I (2007) Canopy shade causes a rapid and transient arrest in leaf development through auxin-induced cytokinin oxidase activity. Genes Dev 21:1863–1868
Casal JJ, Fankhauser C, Coupland G, Blázquez MA (2004) Signalling for developmental plasticity. Trends Plant Sci 9:309–314
Cerdán PD, Chory J (2003) Regulation of flowering time by light quality. Nature 423:881–885
Chatfield SP, Stirnberg P, Forde BG, Leyser O (2000) The hormonal regulation of axillary bud growth in Arabidopsis. Plant J 24:159–169
Cline M (1996) Exogenous auxin effects on lateral bud outgrowth in decapitated shoots. Ann Bot 78:255–266
Cline M (2000) Execution of the auxin replacement apical dominance experiment in temperate woody species. Am J Bot 87:182–190
Cline MG, Riley L (1984) The presentation time for shoot inversion release of apical dominance in Pharbitis nil. Ann Bot 53:897–900
Cline MG, Chatfield SP, Leyser O (2001) NAA restores apical dominance in the axr3-1 mutant of Arabidopsis thaliana. Ann Bot 87:61–65
Cook CE, Whichard LP, Wall ME, Egley GH, Coggon P, Luhan PA et al (1972) Germination stimulants. II. The structure of strigol—a potent seed germination stimulant for witchweed (Striga lutea Lour.). J Am Chem Soc 94:6198–6199
Davies CR, Wareing PF (1965) Auxin-directed transport of radiophosphorus in stems. Planta 65:139–156
Devitt ML, Stafstrom JP (1995) Cell cycle regulation during growth-dormancy cycles in pea axillary buds. Plant Mol Biol 29:255–265
Doebley J, Stec A, Hubbard L (1997) The evolution of apical dominance in maize. Nature 386:485–488
Doust AN (2007) Grass architecture: genetic and environmental control of branching. Curr Opin Plant Biol 10:21–25
Doust AN, Devos KM, Gadberry MD, Gale MD, Kellogg EA (2004) Genetic control of branching in foxtail millet. Proc Natl Acad Sci USA 101:9045–9050
Dun EA, Ferguson BJ, Beveridge CA (2006) Apical dominance and shoot branching. Divergent opinions or divergent mechanisms? Plant Physiol 142:812–819
Dun EA, Brewer PB, Beveridge CA (2009) Strigolactones: discovery of the elusive shoot branching hormone. Trends Plant Sci 14:364–372
Ferguson BJ, Beveridge CA (2009) Roles for auxin, cytokinin and strigolactone in regulating shoot branching. Plant Physiol 149:1929–1944
Finlayson S (2007) Arabidopsis TEOSINTE BRANCHED1-LIKE1 regulates axillary bud outgrowth and is homologous to monocot TEOSINTE BRANCHED1. Plant Cell Physiol 48:667–677
Foo E, Bullier E, Goussot M, Foucher F, Rameau C, Beveridge CA (2005) The branching gene RAMOSUS1 mediates interactions among two novel signals and auxin in pea. Plant Cell 17:464–474
Foo E, Morris SE, Parmenter K, Young N, Wang H, Jones A et al (2007) Feedback regulation of xylem cytokinin content is conserved in pea and Arabidopsis. Plant Physiol 143:1418–1428
Foucher F, Morin J, Courtiade J, Cadioux S, Ellis N, Banfield MJ, Rameau C (2003) DETERMINATE and LATE FLOWERING are two TERMINAL FLOWER1/CENTRORADIALIS homologs that control two distinct phases of flowering initiation and development in pea. Plant Cell 15:2742–2754
Gomez-Roldan V, Fermas S, Brewer PB, Peuch-Pagès V, Dun EA, Pillot J-P et al (2008) Strigolactone inhibition of shoot branching. Nature 455:189–194
Grbić B, Bleecker AB (2000) Axillary meristem development in Arabidopsis thaliana. Plant J 21:215–223
Hayward A, Stirnberg P, Beveridge CA, Leyser O (2009) Interactions between auxin and strigolactone in shoot branching control. Plant Physiol 151:400–412
Hecht V, Foucher F, Ferrándiz C, Macknight R, Navarro C, Morin J et al (2005) Conservation of Arabidopsis flowering genes in model legumes. Plant Physiol 137:1420–1434
Horvath D (2009) Common mechanisms regulate flowering and dormancy. Plant Sci 177:523–531
Hu W, Zhang S, Zhao Z, Sun C, Zhao Y, Luo D (2003) The analysis of the structure and expression of OsTB1 gene in rice. J Plant Physiol Mol Biol 29:507–514
Humphrey AJ, Beale MH (2006) Strigol: biogenesis and physiological activity. Phytochemistry 67:636–640
Husain SM, Linck AJ (1966) Relationship of apical dominance to the nutrient accumulation in Pisum sativum var. Alaska. Physiol Plant 19:992–1010
Ishikawa S, Maekawa M, Arite T, Onishi K, Takamure I, Kyozuka J (2005) Suppression of tiller bud activity in tillering dwarf mutants of rice. Plant Cell Physiol 46:79–86
Jensen CS, Salchert K, Nielsen KK (2001) A TERMINAL FLOWER1-like gene from perennial ryegrass involved in floral transition and axillary meristem identity. Plant Physiol 125:1517–1528
Jiang F, Li C, Jeschke WD, Zhang F (2001) Effect of top excision and replacement by 1-naphthylacetic acid on partition and flow of potassium in tobacco plants. J Exp Bot 52:2143–2150
Johnson X, Brcich T, Dun EA, Goussot M, Haurogné K, Beveridge CA et al (2006) Branching genes are conserved across species. Genes controlling a novel signal in pea are co-regulated by other long-distance signals. Plant Physiol 142:1014–1026
Kebrom TH, Brutnell TP (2007) The molecular analysis of the shade avoidance syndrome in the grasses has begun. J Exp Bot 58:3079–3089
Kebrom TH, Burson BL, Finlayson SA (2006) Phytochrome B represses Teosinte Branched1 expression and induces sorghum axillary bud outgrowth in response to light signals. Plant Physiol 140:1109–1117
King RA, Van Staden J (1988) Differential responses of buds along the shoot of Pisum sativum to isopentyladenine and zeatin application. Plant Physiol Biochem 26:253–259
Kitazawa D, Miyazawa Y, Fujii N, Hoshino A, Iida S, Nitasaka E et al (2008) The gravity-regulated growth of axillary buds is mediated by a mechanism different from decapitation-induced release. Plant Cell Physiol 49:891–900
Klee H (2008) Plant biology: hormones branch out. Nature 455:176–177
Kyozuka J (2007) Control of shoot and root meristem function by cytokinin. Curr Opin Plant Biol 10:442–446
Lang GA, Early JD, Martin GC, Darnell RL (1987) Endo-, para-, and eco-dormancy: physiological terminology and classification for dormancy research. Hortic Sci 22:371–377
Leyser O (2008) Strigolactones and shoot branching: a new trick for a young dog. Dev Cell 15:337–338
Leyser O (2009) The control of shoot branching: an example of plant information processing. Plant Cell Environ 32:694–703
Li CJ, Bangerth F (1999) Autoinhibition of indoleacetic acid transport in the shoots of two-branched pea (Pisum sativum) plants and its relationship to correlative dominance. Physiol Plant 106:415–420
Liew LC, Hecht V, Laurie RE, Knowles CL, Vander Schoor JK, Macknight RC, Weller JL (2009) DIE NEUTRALIS and LATE BLOOMER 1 contribute to regulation of the pea circadian clock. Plant Cell 21:3198–3211
Lin H, Wang R, Qian Q, Yan M, Meng X, Fu Z et al (2009) DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth. Plant Cell 21:1512–1525
López-Ráez JA, Charnikhova T, Gómez-Roldán V, Matusova R, Kohlen W, De Vos R, Verstappen F, Puech-Pages V, Bécard G, Mulder P, Bouwmeester H (2008) Tomato strigolactones are derived from carotenoids and their biosynthesis is promoted by phosphate starvation. New Phytol 178:863–874
López-Ráez JA, Matusova R, Cardoso C, Jamil M, Charnikhova T, Kohlen W et al (2009) Strigolactones: ecological significance and use as a target for parasitic plant control. Pest Manag Sci 65:471–477
Mader JC, Turnbull CGN, Emery RJN (2003) Transport and metabolism of xylem cytokinins during lateral bud release in decapitated chickpea (Cicer arietinum) seedlings. Physiol Plant 117:118–129
Madoka Y, Mori H (2000) Acropetal disappearance of PsAD1 Protein in pea axillary buds after the release of apical dominance. Plant Cell Physiol 41:556–564
Matusova R, Rani K, Verstappen FWA, Franssen MCR, Beale MH, Bouwmeester HJ (2005) The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche spp. are derived from the carotenoid pathway. Plant Physiol 139:920–934
McSteen P (2009) Hormonal regulation of branching in grasses. Plant Physiol 149:46–55
McSteen P, Leyser O (2005) Shoot branching. Annu Rev Plant Biol 56:353–374
Miyawaki K, Matsumoto-Kitano M, Kakimoto T (2004) Expression of cytokinin biosynthetic isopentenyltransferase genes in Arabidopsis: tissue specificity and regulation by auxin, cytokinin, and nitrate. Plant J 37:128–138
Mockaitis K, Estelle M (2008) Auxin receptors and plant development: a new signaling paradigm. Annu Rev Cell Dev Biol 24:55–80
Morris DA (1977) Transport of exogenous auxin in two-branched dwarf pea seedlings (Pisum sativum L.): some implication for polarity and apical dominance. Planta 136:91–96
Morris DA, Johnson CF (1990) The role of auxin efflux carriers in the reversible loss of polar auxin transport in the pea (Pisum sativum L.) stem. Planta 181:117–124
Morris SE, Cox MCH, Ross JJ, Krisantini S, Beveridge CA (2005) Auxin dynamics after decapitation are not correlated with the initial growth of axillary buds. Plant Physiol 138:1665–1672
Murfet IC, Symons GC (2000a) Double mutant rms2 rms5 expresses a transgressive, profuse branching phenotype. Pisum Genet 32:33–38
Murfet IC, Symons GC (2000b) The pea rms2-1 rms4-1 double-mutant phenotype is transgressive. Pisum Genet 32:59–60
Nakagawa M, Shimamoto K, Kyozuka J (2002) Overexpression of RCN1 and RCN2, rice TERMINAL FLOWER 1/CENTRORADIALIS homologs, confers delay of phase transition and altered panicle morphology in rice. Plant J 29:743–750
Napoli C (1996) Highly branched phenotype of the petunia dad1-1 mutant is reversed by grafting. Plant Physiol 111:27–37
Napoli CA, Beveridge CA, Snowden KC (1999) Re-evaluating concepts of apical dominance and the control of axillary bud outgrowth. Curr Top Dev Biol 44:127–169
Nordström A, Tarkowski P, Tarkowska D, Norbaek R, Åstot C, Dolezal K, Sandberg G (2004) Auxin regulation of cytokinin biosynthesis in Arabidopsis thaliana: a factor of potential importance for auxin-cytokinin-regulated development. Proc Nat Acad Sci 101(21):8039–8044
Ongaro V, Leyser O (2008) Hormonal control of shoot branching. J Exp Bot 59:67–74
Ongaro V, Bainbridge K, Williamson L, Leyser O (2008) Interactions between axillary branches of Arabidopsis. Mol Plant 1:388–400
Ouellet F, Overoorde PJ, Theologis A (2001) IAA17/AXR3: biochemical insight into an auxin mutant phenotype. Plant Cell 13:829–841
Phillips IDJ (1968) Nitrogen, phosphorus and potassium distribution in relation to apical dominance in dwarf bean (Phaseolus vulgaris, c.v. Canadian Wonder). J Exp Bot 19:617–627
Pnueli L, Gutfinger T, Hareven D, Ben-Nairn O, Ron N, Adir N et al (2001) Tomato SP-interacting proteins define a conserved signalling system that regulates shoot architecture and flowering. Plant Cell 13:2687–2702
Prasad TK, Cline MG (1985) Shoot inversion-induced ethylene in Pharbitis nil induces the release of apical dominance by restricting shoot elongation. Plant Sci 38:163–172
Ratcliffe OJ, Amaya I, Vincent CA, Rothstein S, Carpenter R et al (1998) A common mechanism controls the life cycle and architecture of plants. Development 125:1609–1615
Reed JW, Nagpal P, Poole DS, Furuya M, Chory J (1993) Mutations in the gene for the red/far-red light receptor phytochrome B alter cell elongation and physiological responses throughout Arabidopsis development. Plant Cell 5:147–157
Robson PRH, Whitelam GC, Smith H (1993) Selected components of the shade-avoidance syndrome are displayed in a normal manner in mutants of Arabidopsis thaliana and Brassica rapa deficient in phytochrome B. Plant Physiol 102:1179–1184
Rubio V, Bustos R, Irigoyen ML, Cardona-López X, Rojas-Triana M, Paz-Ares J (2009) Plant hormones and nutrient signaling. Plant Mol Biol 69:361–373
Sachs T, Thimann K (1964) Release of lateral buds from apical dominance. Nature 201:939–940
Sachs T, Thimann KV (1967) The role of auxins and cytokinins in the release of buds from dominance. Am J Bot 54:136–144
Shalit A, Rozman A, Goldshmidt A, Alvarez JP, Bowman JL, Eshed Y et al (2009) The flowering hormone florigen functions as a general systemic regulator of growth and termination. Proc Natl Acad Sci USA 106:8392–8397
Shen H, Luong P, Huq E (2007) The F-box protein MAX2 functions as a positive regulator of photomorphogenesis in Arabidopsis. Plant Physiol 145:1471–1483
Shimizu S, Mori H (1998) Analysis of cycles of dormancy and growth in pea axillary buds based on mRNA accumulation patterns of cell cycle-related genes. Plant Cell Physiol 39:255–262
Shimizu-Sato S, Mori H (2001) Control of outgrowth and dormancy in axillary buds. Plant Physiol 127:1405–1413
Shimizu-Sato S, Ike Y, Mori H (2008) PsRBR1 encodes a pea retinoblastoma-related protein that is phosphorylated in axillary buds during dormancy-to-growth transition. Plant Mol Biol 66:125–135
Shimizu-Sato S, Tanaka M, Mori H (2009) Auxin-cytokinin interactions in the control of shoot branching. Plant Mol Biol 69:429–435
Snowden KC, Napoli CA (2003) A quantitative study of lateral branching in petunia. Funct Plant Biol 30:987–994
Stafstrom JP (1993) Axillary bud development in pea: apical dominance, growth cycles, hormonal regulation and plant architecture. In: Amasino RM (ed) Cellular communication in plants. Plenum Press, New York, pp 75–86
Stafstrom JP, Sussex IM (1988) Patterns of protein synthesis in dormant and growing vegetative buds of pea. Planta 176:497–505
Stafstrom JP, Sussex IM (1992) Expression of a ribosomal protein gene in axillary buds of pea seedlings. Plant Physiol 100:1494–1502
Stahlberg R, Cosgrove DJ (1992) Rapid alteration in growth rate and electric potentials upon stem excision in pea seedlings. Planta 187:523–531
Stirnberg P, van de Sande K, Leyser HMO (2002) MAX1 and MAX2 control shoot lateral branching in Arabidopsis. Development 129:1131–1141
Stirnberg P, Furner IJ, Leyser HMO (2007) MAX2 participates in an SCF complex which acts locally at the node to suppress shoot branching. Plant J 50:80–94
Takeda T, Suwa Y, Suzuki M, Kitano H, Ueguchi-Tanaka M, Ashikari M et al (2003) The OsTB1 gene negatively regulates lateral branching in rice. Plant J 33:513–520
Takei K, Ueda N, Aoki K, Kuromori T, Hirayama T, Shinozaki K et al (2004) AtIPT3 is a key determinant of nitrate-dependent cytokinin biosynthesis in Arabidopsis. Plant Cell Physiol 45:1053–1062
Tanaka M, Takei K, Kojima M, Sakakibara H, Mori H (2006) Auxin controls local cytokinin biosynthesis in the nodal stem in apical dominance. Plant J 45:1028–1036
Taylor SA, Murfet IC (1994) A short day mutant in pea is deficient in the floral stimulus. Flower Newsl 18:39–43
Thimann KV, Skoog F (1933) Studies on the growth hormone of plants III. The inhibiting action of the growth substance on bud development. Proc Natl Acad Sci USA 19:714–716
Thimann KV, Skoog F (1934) On the inhibition of bud development and other functions of growth substances in Vicia faba. Proc R Soc Lond B Biol Sci 114:317–339
Turnbull CGN, Raymond MAA, Dodd IC, Morris SE (1997) Rapid increases in cytokinin concentration in lateral buds of chickpea (Cicer arietinum L.) during the release of apical dominance. Planta 202:271–276
Umehara M, Hanada A, Yoshida S, Akiyama K, Arite T, Takeda-Kamiya N et al (2008) Inhibition of shoot branching by new terpenoid plant hormones. Nature 455:195–200
Wang G, Römheld V, Li C, Bangerth F (2006) Involvement of auxin and CKs in boron deficiency induced changes in apical dominance of pea plants (Pisum sativum L.). J Plant Physiol 163:591–600
Weberling F (1989) Morphology of flowers and inflorescences. Cambridge University Press, Cambridge
Weller JL, Hecht V, Liew LC, Sussmilch FC, Wenden B, Knowles CL et al (2009) Update on the genetic control of flowering in garden pea. J Exp Bot 60:2493–2499
Woo HE, Chung KM, Park JH, Oh SA, Ahn T, Hong SH et al (2001) ORE9, an F-box protein that regulates leaf senescence in Arabidopsis. Plant Cell 13:1779–1790
Yan H, Saika H, Maekawa M, Takamure I, Tsutsumi N, Kyozuka J, Nakazono M (2007) Rice tillering dwarf mutant dwarf3 has increased leaf longevity during darkness-induced senescence or hydrogen peroxide-induced cell death. Genes Genet Syst 82:361–366
Yang HY, Li CJ, Zhang FS (2007) Shoot apex demand determines assimilate and nutrients partitioning and nutrient-uptake rate in tobacco plants. J Integr Plant Biol 49:1654–1661
Yoneyama K, Yoneyama K, Takeuchi Y, Sekimoto H (2007) Phosphorus deficiency in red clover promotes exudation of orobanchol, the signal for mycorrhizal symbionts and germination stimulant for root parasites. Planta 225(4):1031–1038
Zou J, Zhang S, Zhang W, Li G, Chen Z, Zhai W et al (2006) The rice HIGH-TILLERING DWARF1 encoding an orthologue of Arabidopsis MAX3 is required for negative regulation of the outgrowth of axillary buds. Plant J 48:687–696
Acknowledgments
We thank the Australian Research Council for grant funding and the Australian Postgraduate Award scheme for funding to AH and TW.
Author information
Authors and Affiliations
Corresponding author
Additional information
Tanya Waldie and Alice Hayward contributed equally to this work.
Rights and permissions
About this article
Cite this article
Waldie, T., Hayward, A. & Beveridge, C.A. Axillary bud outgrowth in herbaceous shoots: how do strigolactones fit into the picture?. Plant Mol Biol 73, 27–36 (2010). https://doi.org/10.1007/s11103-010-9599-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11103-010-9599-2