
Abstract
The dynamics of a diffusive predator–prey model with time delay and Michaelis–Menten-type harvesting subject to Neumann boundary condition is considered. Turing instability and Hopf bifurcation at positive equilibrium for the system without delay are investigated. Time delay-induced instability and Hopf bifurcation are also discussed. By the theory of normal form and center manifold, conditions for determining the bifurcation direction and the stability of bifurcating periodic solution are derived. Some numerical simulations are carried out for illustrating the theoretical results.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Dynamics of predator–prey model is one of important subjects in ecology and mathematical ecology, and many researchers have studied it and derive some important results [1–9]. Leslie–Gower model [10, 11] is one of the classical predator–prey models. Chen et al. [12] discussed the stability/instability of the coexistence equilibrium and associated Hopf bifurcation in a diffusive Leslie–Gower predator–prey model. Aziz-Alaoui and Okiye [13] studied the boundedness and global stability in a modified Leslie–Gower predator–prey model with Holling type II functional response:
where x and y represent the population densities of prey and predator, respectively. All parameters are positive parameters. \(r_1\) and \(r_2\) are the growth rate of prey and predator. \(b_1\) represents the competition among individuals of prey. \(a_1\) and \(a_2\) are the maximum value which per capita reduction rate of prey and predator can attain. \(k_1\) is the average saturation rate. In this model, in the case of prey severe scarcity, predator can switch to other foods denoted as \(k_2\).
Considering time delay in the negative feedback of the predator’s density, Nindjina et al. [14] investigated the following model:
In [14], Nindjina et al. discussed the global stability of the positive equilibrium by constructing a Lyapunov function. In [15], Yafia et al. investigated the Hopf bifurcations at the positive equilibrium.
For economic reasons, human needs to exploit biological resources and harvest some biological species, such as in fishery, forestry and wildlife management. Therefore, it is necessary to study the suitable population model with harvesting. Many researchers have studied system (1.2) with different types of harvesting, constant harvesting [16], linear harvesting[17], non-selective harvesting [18] and so on. Among these types of harvesting, Yuan et al. [19] suggest that Michaelis–Menten-type prey harvesting is more realistic than other types of harvesting from biological and economic points of view. They studied the following model:
All parameters are positive. q represents the catch ability, E is the effort applied to harvest prey, and \(m_1\) and \(m_2\) are suitable constants. In [19], Yuan et al. assume that the environment provides the same protection to both the predator and prey (\(k_1=k_2\)), and discuss the stability of the equilibria and obtained the critical conditions for the saddle-node-Hopf bifurcation.
In the real world, predators and their preys distribute inhomogeneous in different spatial location at time t. And they will move or diffuse to areas with smaller population concentration or more food to get a good living environment. Hence, taking into account diffusion appears to be more reasonable. In mathematics, predator–prey with diffusion will exhibit complex dynamical properties. Many researchers have shown that the diffusion coefficients may induce Turing instability and spatially non-homogeneous bifurcating periodic solution [20–23]. Hence, taking into account diffusion appears to be more reasonable and interesting. In this manuscript, we suppose the region prey and predator lived is closed and no species (prey or predator) entering and leaving region at the boundary. Therefor, we choose Neumann boundary condition. On the other hand, time delay plays an important role in many biological dynamical systems, being particularly relevant into predator–prey models [24–27]. In predator–prey models, time delay exists in maturation time, capturing time, gestation time or others. Many scholars have devote to investigating delayed predator–prey models and suggest that time delay contributes critically to the stable or unstable outcome of prey and predator’s densities. Time delay may induce bifurcating periodic solution, and prey and predator’s densities exhibit oscillatory behavior. Different from works in [19], we introduce time delay in the resource limitation of the prey which is one of important aspects [25–27]. Based on these reasons, we investigate the following system:
For simplicity, we also assume \(k_1=k_2=k\). After the following nondimensionalization: \(u=\frac{r_1}{b_1}\widetilde{u}, v= \frac{r_1}{a_1b_1}\widetilde{v}, t= \frac{\widetilde{t}}{r_1}, d_1=\frac{D_1}{r_1}, d_2=\frac{D_2}{r_1}, \alpha = \frac{1}{r_1}, \beta = \frac{a_2}{r_2a_1}, m= \frac{kb_1}{r_1}, s= \frac{r_2}{r_1}, h = \frac{qEb_1}{r_1^2m_2}, c= \frac{m_1Eb_1}{m_2r_1}\) and drop the tilde, system (1.1) can be changed to
In this paper, we assume \(\Omega =(0,l\pi ), l> 0\).
The organization of this paper is as follows. In Sect. 2, we study the dynamics of non-delay system, including stability, Turing instability and existence of Hopf bifurcation at positive equilibrium. In Sect. 3, we study the effect of delay on the model including stability and Hopf bifurcation at positive equilibrium. In Sect. 4, we give some numerical simulations. Finally, we end the paper with a brief conclusion in Sect. 5.
2 The effect of diffusion on the non-delay model
Without delay, system (1.5) becomes
In [19], Yuan et al. have discussed the existence of trivial and positive equilibria. For convenience, in this paper we assume system (2.1) has a positive equilibrium and denote as \(E_*(u_*,v_*)\).
2.1 Local stability analysis of the model without diffusion
For system (2.1) without diffusion, the Jacobian matrix at \(E_*(u_*,v_*)\) is
where
Obviously, \(a_2<0\). The characteristic equation corresponding to \(E_*(u_*,v_*)\) is
Makeing the following hypotheses:
Theorem 2.1
Suppose \(\mathbf {(H_1)}\) holds. Then for system (2.1) without diffusion, the following statements are true.
-
(i)
If \(a_1\le 0\), for \(s>0\) the equilibrium \(E_*(u_*,v_*)\) is local asymptotically stable;
-
(ii)
If \(a_1>0\), for \(s>a_1\) the equilibrium \(E_*(u_*,v_*)\) is local asymptotically stable;
-
(iii)
If \(a_1>0\), the system undergoes Hopf bifurcation at \(E_*(u_*,v_*)\) when \(s=a_1\).
Proof
Obviously, the roots of Eq. (2.3) are given by
Under condition (i) (or (ii)), \(a_1-s<0\) holds; then, the roots of Eq. (2.3) have negative real parts. Therefore, the equilibrium \(E_*(u_*,v_*)\) is local asymptotically stable.
When \(s=a_1\), Eq. (2.3) has a pair of pure imaginary roots \(\pm \sqrt{-4s(a_1 +a_2/\beta )}\). Meanwhile, when s near \(a_1\), Eq. (2.3) has a pair of complex eigenvalues \(\alpha (s)\pm i\omega (s)\), where
And hence, we have
Therefore, the system undergoes Hopf bifurcation at \(E_*(u_*,v_*)\) when \(s=a_1\).
2.2 Turing instability and Hopf bifurcation
For system (2.1), the characteristic equation at \(E_*(u_*,v_*)\) is
where
and the eigenvalues are given by
Obviously, if \(a_1-s<0\), then \(T_n(s)\le T_0(s)<0\) for \(n\in \mathbb {N}_0\). Suppose \(\mathbf{(H_1)}\) holds; then, \(D_0(s)=-s(a_1 +a_2/\beta )>0\), and if \(s\ge \frac{ d_2a_1}{d_1} \) also holds, then \(D_n(s)\ge D_0(s)>0\).
Denote
and
Remark 2.1
Under the hypotheses \(\mathbf{(H_1)}\), we can obtain the following relationship about \(a_1, \frac{ d_2a_1}{d_1}\) and \(s_{\pm }\):
Lemma 2.1
Suppose \(\mathbf{(H_1)}\) holds; then, the following statements are true.
-
(i)
If for \(s\in (0,s_-)\cup (s_+,\infty )\), there exists a \(k \in \mathbb {N}\) such that \(\frac{k^2}{l^2}\in (z_-, z_+)\), then \(D_k(s)<0\);
-
(ii)
If one of followings holds:
$$\begin{aligned} (1)~~s\in (s_-, s_+), \end{aligned}$$$$\begin{aligned}&(2)~~s\,{\in }\, (0,s_-)\cup (s_+,\infty ), \text {but there are no } k \in \mathbb {N}\\&\quad ~~ \text{ such } \text{ that }~~\frac{k^2}{l^2}\in (z_-, z_+), \end{aligned}$$then \(D_k(s)>0\) for \(n\in \mathbb {N}_0,\).
Proof
Define
If \((d_2a_1-sd_1)^2+4d_1d_2s(a_1 +a_2/\beta )>0\) that is \(s\in (0,s_-)\cup (s_+,\infty )\), then \(h(z)=0\) has two roots \(z_{\mp }\). And if there exists a \(k\in \mathbb {N}\) such that \(\frac{k^2}{l^2}\in (z_-, z_+)\), then \(D_k(c)=h\left( \frac{k^2}{l^2}\right) <0\). This completes the proof of (i).
From the discussion above, we know that \(D_n(s)>0\) for \(n=0,1,2,\ldots ,\) under the conditions of (ii). Hence, the conclusion of (ii) follows.
Theorem 2.2
Suppose \(\mathbf{(H_1)}\) holds, and \(\sigma \) and \(s_-\) are defined by (2.9) and (2.7), respectively. Then for system (2.1), the following statements are true.
-
(i)
If \(s>a_1\) and \(s\ge \frac{ d_2a_1}{d_1}\), then the equilibrium \(E_*(u_*,v_*)\) is asymptotically stable;
-
(ii)
If \(a_1< s <\frac{ d_2a_1}{d_1}\) and \(\frac{d_1}{d_2}>\sigma \), then the equilibrium \(E_*(u_*,v_*)\) is asymptotically stable;
-
(iii)
If \(a_1< s <\frac{ d_2a_1}{d_1}\) and \(\frac{d_1}{d_2}<\sigma \), then the equilibrium \(E_*(u_*,v_*)\) is asymptotically stable when one of the followings holds: (1) \(s \in (s_-,\frac{ d_2a_1}{d_1})\); (2) \(s\in (a_1,s_-)\), but there does not exist a \(k\in \mathbb {N}\) such that \(\frac{k^2}{l^2}\in (z_-, z_+)\);
-
(iv)
If \(a_1<s<s_-\) and \(\frac{d_1}{d_2}<\sigma \), and there exists a \(k\in \mathbb {N}\) such that \(\frac{k^2}{l^2}\in (z_-, z_+)\), then the equilibrium \(E_*(u_*,v_*)\) is Turing unstable;
-
(v)
If \(a_1>0\), the system (2.1) undergoes a Hopf bifurcation at \(E_*(u_*,v_*)\) when \(s=s_n\), for \(0\le n \le n^*-1\), where \(s_n\) and \(n^*\) are defined in the following proof. Moreover, the bifurcating periodic solution is spatially homogeneous when \(s=s_0\) and spatially non-homogeneous when \(s=s_n\) for \(1\le n \le n^*-1\).
Proof
Obviously, under conditions (i), (ii) or (iii), \(T_n(s)<0\) and \(D_n(s)>0\) for \(n\in \mathbb {N}_0\). Then, all roots of Eq. (2.4) have negative real parts. Therefore, the equilibrium \(E_*(u_*,v_*)\) is asymptotically stable. Under condition (iv), by Lemma (2.1, there exists a \(k\in \mathbb {N}\) such that \(D_k(s)<0\). Then, Eq. (2.4) has a root \(\lambda ^{(k)}(s)\) with positive real parts. Therefore, the equilibrium \(E_*(u_*,v_*)\) is Turing unstable.
Suppose \(\mathbf{(H_1)}\) holds, and \(a_1>0\), from (2.6), we know that (2.4) has purely imaginary roots if and only if
and \(D_n(s_n)>0\). From (2.11), we know that there exists a integer \(n_1^*\ge 1\) such that \(s_n>0\) for \(n=0,1,2,\ldots ,n_1^*-1\), and \(s_n \le 0\) for \(n=n_1^*,n_1^*+1,\ldots \). Substituting \(s_n\) into \(D_n(s)\) (see (2.5)) yields
By \(D_0(s_0)=-a_1\left( a_2/\beta +a_1\right) >0\), we know that there exists an integer \(n_2^*\ge 1\) such that \(D_n(s_n)>0\) when \(n=0,1,\ldots , n_2^*-1\), and \(D_n(s_n)\le 0\) when \(n\ge n_2^*\). Let \(n^*=\min \{ n_1^*, n_2^*\}\) and
be the roots of Eq. (2.4) satisfying
Then, when s is near \(s_n\)
and from the definition of \(T_n\) in (2.5), it follows that
This implies that the transversal condition is satisfied at each \(s_n, n=0,1,2, \ldots , n^*-1\). Therefore, the system (2.1) undergoes a Hopf bifurcation at \(E_*(u_*,v_*)\) when \(s=s_n\), for \(0\le n \le n^*-1\).
3 The effect of delay on the system
3.1 Stability analysis and existence of Hopf bifurcation
In the following, by analyzing the associated characteristic equation at \(E_*(u_*,v_*)\), we investigate stability of \(E_*(u_*,v_*)\) and existence of Hopf bifurcation for system (1.5). We always suppose \(\mathbf{(H_1)}\) and one of conditions (i-iii) in Theorem (2.2) hold.
Denote

Linearizing system (1.5) at \(E_*(u_*,v_*)\), we have
where

and \(L:\mathscr {C}_{\tau } \mapsto X\) is defined by
for \(\phi =(\phi _1, \phi _2)^T \in \mathscr {C}_{\tau }\) with
From Wu [28], we obtain that the characteristic equation for linear system (3.1) is
It is well known that the eigenvalue problem
has eigenvalues \(\mu _n=n^2/l^2~(n=0,1,\cdots )\) with corresponding eigenfunctions
Substituting
into the characteristic Eq. (3.2), it follows that
Therefore, the characteristic Eq. (3.2) is equivalent to
where
When \(\tau =0\), system (1.5) becomes (2.1); if one of conditions (i–iii) in Theorem (2.2) holds, then all the roots of Eq. (3.3) with \(\tau =0\) have negative real parts for \(n \in \mathbb {N}_0\) and \(\Delta _n(0,\tau )>0\).
We shall seek critical values of \(\tau \) such that there exists a pair of simple purely imaginary eigenvalues. \(i\omega \) (\(\omega >0\)) is a root of Eq. (3.3) if and only if \(\omega \) satisfies
Then, we have
which lead to
Let \(z = \omega ^2\), then (3.4) can be rewritten into the following form
Denote
Then, the roots of (3.5) are given by \(z_{\pm }=\frac{P \pm \sqrt{R}}{2}.\) We discuss the existence of positive roots for Eq. (3.5) under these three cases:
-
Case 1. (i) \(R<0\); (ii) \(R>0, Q>0, P<0\); (ii) \(R=0, P\le 0\).
-
Case 2. (i) \(Q<0\); (ii) \(R=0, P>0\).
-
Case 3. (i) \(P>0, Q>0, R>0\).
Obviously, in Case 1, Eq. (3.5) has no positive root; then, Eq. (3.3) has no root with purely imaginary. In Case 2, Eq. (3.5) has one positive root; then, Eq. (3.3) has a pair of purely imaginary roots \(\pm i\omega ^+_n\) at \(\tau ^{j,+}_n,~j=0,1,2,\ldots \). In Case 3, Eq. (3.5) has two positive roots; then, Eq. (3.3) has two pair of purely imaginary roots \(\pm i\omega ^\pm _n\) at \(\tau ^{j,\pm }_n,~j\in \mathbb {N}_0\) where
Fix parameters \(\alpha ,~h,~m,~c,~\beta ,~s,~d_1,~d_2,~l\), define
Lemma 3.1
Suppose one of conditions \((i-iii)\) in Theorem (2.2) and \(\mathbf{{(H_1)}}\) hold.
-
(i)
If \(R=0\), then \(\text {Re}\left( \frac{\mathrm{d} \lambda }{\mathrm{d} \tau }\right) |_{\tau =\tau ^{j,\pm }_n}=0\);
-
(ii)
If \(R>0\), then \(\text {Re}\left( \frac{\mathrm{d} \lambda }{\mathrm{d} \tau }\right) |_{\tau =\tau ^{j,+}_n}>0, \text {Re} ( \frac{\mathrm{d} \lambda }{\mathrm{d} \tau })|_{\tau =\tau ^{j,-}_n}<0\) for \(\tau \in \mathcal {D}\) and \(j \in \mathbb {N}_0\).
Proof
Differentiating two sides of (3.3) with respect \(\tau \), we have
Then
where \(\Lambda =(\omega ^{j,\pm }_n)^4 u_*^2 +C^2_n u_*^2 (\omega ^{j,\pm }_n)^2>0\). Therefore, \(\alpha '_n(\tau ^{j,n}_n)>0(<0)\).
From (3.6), we have \(\tau ^{0,\pm }_n<\tau ^{j,\pm }_n\) \((j\in \mathbb {N})\). For \(k\in \mathcal {D}\), define the smallest \(\tau \) so that the stability will change, \(\tau _*=\text{ min }\{\tau ^{0,\pm }_k ~\text{ or }~ \tau ^{0,+}_k \mid k \in \mathcal {D} \}\). According to the above analysis, we have the following theorem.
Theorem 3.1
Suppose \(\mathbf{(H_1)}\) and one of conditions (i–iii) in Theorem (2.2) hold; for system (1.5), the following statements are true.
-
(i)
In Case 1, the equilibrium \(E_*(u_*,v_*)\) is local asymptotically stable for all \(\tau \ge 0\);
-
(ii)
In Case 2 or Case 3, the equilibrium \(E_*(u_*,v_*)\) is local asymptotically stable for \(\tau \in [0,\tau _*)\) and unstable for \(\tau \in [\tau _*,\tau _*+\epsilon )\) with some \(\epsilon \);
-
(iii)
In Case 2 or Case 3, system (1.5) undergoes a Hopf bifurcation at the equilibrium \(E_*(u_*,v_*)\) when \(\tau =\tau ^{j,+}_n\) \((\tau =\tau ^{j,-}_n), j\in \mathbb {N}_0, n \in \mathcal {D}\).
3.2 Stability and direction of Hopf bifurcation
In this section, we shall study the direction of Hopf bifurcation and stability of the bifurcating periodic solution by applying center manifold theorem and normal form theorem of partial functional differential equations [28, 29]. Let \(\tilde{u}(x,t)=u(x,\tau t)-u_*\) and \(\tilde{v}(x,t)=v(x,\tau t)-v_*\). For convenience, we drop the tilde. Then, the system (1.5) can be transformed into
for \(x \in (0, l\pi )\), and \(t>0\). Let
When \(\mu = 0\), system (1.5) undergoes a Hopf bifurcation at the equilibrium (0, 0). Then, (3.8) can be rewritten in an abstract form in the phase space \(\mathscr {C}_1:=C([-1,0],X)\)
where \(L_{\mu }(\phi )\) and \(F(\phi ,\mu )\) are defined by
with
respectively, for \(\phi =(\phi _1, \phi _2)^T \in \mathscr {C}_1\).
Consider the linear equation
According to the results in Sect. 2, we know that \(\Lambda _n:=\{i \omega _n \tilde{\tau },-i \omega _n \tilde{\tau }\}\) are characteristic values of system (3.12) and the linear functional differential equation
By Riesz representation theorem, there exists \(2\times 2\) matrix function \(\eta ^n(\sigma , \tilde{\tau })-1\le \sigma \le 0\), whose elements are of bounded variation functions such that
for  .
.
In fact, we can choose
where
Let \(A(\tilde{\tau })\) denote the infinitesimal generators of semigroup included by the solutions of Eq. (3.13) and \(A^*\) be the formal adjoint of \(A(\tilde{\tau })\) under the bilinear paring
for  has a pair of simple purely imaginary eigenvalues \(\pm i \omega _n \tilde{\tau }\), and they are also eigenvalues of \(A^*\). Let P and \(P^*\) be the center subspace, that is, the generalized eigenspace of \(A(\tilde{\tau })\) and \(A^*\) associated with \(\Lambda _n\), respectively. Then, \(P^*\) is the adjoint space of P and \(\text{ dim }P=\text{ dim }P^*=2\).
has a pair of simple purely imaginary eigenvalues \(\pm i \omega _n \tilde{\tau }\), and they are also eigenvalues of \(A^*\). Let P and \(P^*\) be the center subspace, that is, the generalized eigenspace of \(A(\tilde{\tau })\) and \(A^*\) associated with \(\Lambda _n\), respectively. Then, \(P^*\) is the adjoint space of P and \(\text{ dim }P=\text{ dim }P^*=2\).
It can be verified that \( p_1(\theta )=(1,\xi )^Te^{i\omega _n \tilde{\tau } \sigma }~~ (\sigma \in [-1,0]),~~p_2(\sigma )=\overline{p_1(\sigma )} \) is a basis of \(A(\tilde{\tau })\) with \(\Lambda _n\) and \( q_1(r)=(1,\eta )e^{-i\omega _n \tilde{\tau } r}~~ (r \in [0,1]), ~~q_2(r)=\overline{q_1(r)} \) is a basis of \(A^*\) with \(\Lambda _n\), where
Let \(\Phi =(\Phi _1,\Phi _2)\) and \(\Psi ^*=(\Psi ^*_1,\Psi ^*_2)^T\) with
for \(\theta \in [-1,0]\), and
for \(r \in [0,1]\). Then, we can compute by (3.16)
Define \((\Psi ^*,\Phi )=(\Psi ^* _j,\Phi _k)=\left( \begin{array}{c} D^*_1~~ D^*_2\\ D^*_3~~ D^*_4 \end{array} \right) \) and construct a new basis \(\Psi \) for \(P^*\) by
Then, \((\Psi ,\Phi )=I_2\). In addition, define \(f_n:=(\beta ^1_n,\beta ^2_n)\), where
We also define
Thus, the center subspace of linear Eq. (3.12) is given by \(P_{CN} \mathscr {C}_1 \oplus P_{S} \mathscr {C}_1\) and \(P_{S} \mathscr {C}_1\) denotes the complement subspace of \(P_{CN} \mathscr {C}_1\) in \(\mathscr {C}_1\),
for \(u=(u_1,u_2), v=(v_1,v_2), u,v\in X\) and \(<\phi ,f_0>=(<\phi ,f^1_0>, <\phi ,f^2_0>)^T\).
Let \(A_{\tilde{\tau }}\) denote the infinitesimal generator of an analytic semigroup induced by the linear system Eqs. (3.12), and (3.8) can be rewritten as the following abstract form
where
By the decomposition of \(\mathscr {C}_1\), the solution above can be written as
where
and
In particular, the solution of (3.9) on the center manifold is given by
Let \(z=x_1-i x_2\), and notice that \(p_1=\Phi _1+i\Phi _2\). Then, we have
and
Hence, Eq. (3.20) can be transformed into
where
From [28], z satisfies
where
Let
from Eqs. (3.21) and (3.24), we have
and
with
Hence,
with
and
Denote
Notice that
and we have
Then, by (3.23), (3.25) and (3.33), we have \(g_{20}=g_{11}=g_{02}=0\), for \(n=1,2,3,\cdots \). If \(n=0\), we have the following quantities:
And for \(n\in \mathbb {N}_0, g_{21}=\tilde{\tau }( \gamma _1 \kappa _1 +\gamma _2 \kappa _2)\).
Now, a complete description for \(g_{21}\) depends on the algorithm for \(W_{20}(0)\) and \(W_{11}(0)\) which we shall compute.
From [28], we have
and \(W(z,\overline{z})\) satisfies
where
Hence, we have
that is
By (3.33), we have that for \(\theta \in [-1,0)\),
Therefore, by (3.34), for \(\theta \in [-1,0)\),
and
where
By the definition of \(A_{\tilde{\tau }}\) and (3.35), we have
That is
where
Using the definition of \(A_{\tilde{\tau }}\) and (3.35), we have that for \(-1\le \theta <0\)
As
and
we have
That is
where

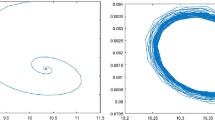
Phase portraits of system (1.5) without delay and diffusion. Left \(s =0.2\) and initial condition (0.3, 0.9). Right \(s =0.09\) and initial condition (0.3, 0.9)

For system (1.5) without delay, \(s=0.2\), and initial condition is (0.3, 0.9). Left component u(x, t) (stable). Right component v(x, t) (stable)
Similarly, from (3.36), we have
That is
Similar to the procedure of computing \(W_{20}\), we have
where
Thus, we can compute the following quantities which determine the direction and stability of bifurcating periodic orbits:
Then, we have the following theorem.

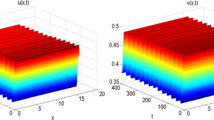
For system (1.5) without delay, \(s=0.2, l=2\), and initial condition is (0.3, 0.9). Left component u(x, t) (Turing unstable). Right component v(x, t) (Turing unstable)

For system (1.5) without delay, \(s=0.2, l=0.5\), and initial condition is (0.3, 0.9). Left component u(x, t) (stable). Right component v(x, t) (stable)
Theorem 3.2
For any critical value \(\tau ^j_n\), we have
-
(i)
\(\mu _2\) determines the directions of the Hopf bifurcation: if \(\mu _2>0\) (resp.<0), then the Hopf bifurcation is forward (resp. backward), that is, the bifurcating periodic solutions exist for \(\mu >0\) (resp. \(\mu <0\)) ;
-
(ii)
\(\beta _2\) determines the stability of the bifurcating periodic solutions on the center manifold: if \(\beta _2<0\) (resp. >0), then the bifurcating periodic solutions are orbitally asymptotically stable (resp. unstable) .
-
(iii)
\(T_2\) determines the period of bifurcating periodic solutions: if \(T_2>0\) (resp. \(T_2<0\)), then the period increases (resp. decreases).
4 Numerical simulations
Fix parameters
Hence, \(E_*(0.3772,0.9544)\) is the unique positive equilibrium, and \(a_1\approx 0.1069, a_2\approx -0.2371, a_1+a_2/\beta \approx -0.3674<0\); then, \(\mathbf{(H_1)}\) holds.
For system (1.5) without delay and diffusion, by Theorem (2.1), if \(s>a_1\), then equilibrium \(E_*(u_*,v_*)\) is locally asymptotically stable, and the system undergoes Hopf bifurcation at \(E_*(u_*,v_*)\) when \(s=a_1\) (shown in Fig. 1).

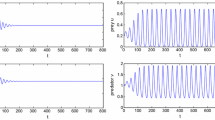
For system (1.5) without delay, \(s=0.09\) and initial condition is (0.3, 0.9). Left component u(x, t) (periodic solution). Right component v(x, t) (periodic solution)

For system (1.5), \(\tau =1.5\) and initial condition is (0.3, 0.9). Left component u(x, t) (stable). Right component v(x, t) (stable)
For system (1.5) without delay, we have \(\sigma \approx 0.0637\). Set \(d_1=0.05, d_2=0.5\) and \(l=2\), then \(d_1/d_2>\sigma \). By Theorem (2.2) (i) and (ii), \(s>a_1\), then equilibrium \(E_*(u_*,v_*)\) is locally asymptotically stable and Turing instability will not occur; this is shown in Fig. 2.
For system (1.5) without delay, set \(d_1=0.05, d_2=3\), then \(d_1/d_2<\sigma \) and \(s_-\approx 0.4088\). If set \(s=0.2\), then \(s\in (a_1,s_-), z_-\approx 0.4683\) and \(z_+\approx 1.5691\). If set \(l=2\), then there exists a \(k=2\) such that \(k^2/l^2 \in (z_-,z_+)\); by Theorem (2.2) (iv), \(E_*(u_*,v_*)\) is Turing unstable, and this is shown in Fig. 3. If set \(l=0.5\), then there doesn’t exist \(k\in \mathbb {N}\) such that \(k^2/l^2 \in (z_-,z_+)\); by Theorem (2.2) (iii), \(E_*(u_*,v_*)\) is locally asymptotically stable, and this is shown in Fig. 4. Set \(s=0.09\), by Theorem (2.2) (v), Hopf bifurcation occurs, this is shown in Fig. 5.

For system (1.5), \(\tau =1.7\) and initial condition is (0.3, 0.9). Left component u(x, t) (periodic solution). Right component v(x, t) (periodic solution)
For system (1.5), set \(d_1=0.05, d_2=3, l=2\) and \(s=0.5\). By direct computation, we have \(\mathcal {D}=[0,1,2,3,4,5,6,7,8]\) and \(\tau _*=\tau ^0_0 \approx 1.6118\). By Theorem (3.1), we know that if \(\tau \in [0,\tau _*)\), then the equilibrium \(E_*(u_*,v_*)\) is locally asymptotically stable. This is shown in Fig. 6. By Theorem (3.1), system (1.5) undergoes a Hopf bifurcation at the equilibrium \(E_*(u_*,v_*)\) when \(\tau =\tau _*\). By Theorem (3.2), we have
Hence, the direction of the bifurcation is forward, and the bifurcating period solutions are locally asymptotically stable. In addition, the period of bifurcating periodic solutions decreases. This is shown in Fig. 7.
5 Conclusion
In this paper, we have considered a diffusive modified Leslie–Gower predator–prey model with Michaelis–Menten-type harvesting in prey. The model shows rich and varied dynamics.
For the model without delay, we study the effect of diffusion, including stability and Turing instability of positive equilibrium. When \(s>a_1\) and \(s\ge \frac{ d_2a_1}{d_1}\), the equilibrium \(E_*(u_*,v_*)\) is asymptotically stable and the diffusion has no effect on the system. When \(d_1/d_2>\sigma \), then for \(s>a_1\) equilibrium \(E_*(u_*,v_*)\) is locally asymptotically stable and Turing instability will not occur. When \(\frac{d_1}{d_2}<\sigma \), for \(a_1<s<s_-\), choose a suitable l (represents the region \(\Omega =(0,l\pi )\)) such that there exist a \(k\in \mathbb {N}\) such that \(\frac{k^2}{l^2}\in (z_-, z_+)\), then Turing instability occurs. But when we change l such that there doesn’t exist \(k\in \mathbb {N}\) satisfying \(k^2/l^2 \in (z_-,z_+), E_*(u_*,v_*)\) is locally asymptotically stable. These results suggest that diffusion coefficients and the region’s size all affect the stability of equilibrium \(E_*(u_*,v_*)\).
In addition, the time delay in the resource limitation of the prey plays an important role in coexistence of predator and prey. We obtained that when \(\tau \) crosses the critical value \(\tau _*\), the stability of the positive equilibrium \(P(u_*,v_*)\) changes and Hopf bifurcation occurs. That means the predator and prey coexist and converge to the coexisting equilibrium point when time delay is smaller than the critical value, and the predator and the prey species may coexist in an oscillatory mode when time delay crosses the critical value.
References
Yi, F., Wei, J., Shi, J.: Bifurcation and spatiotemporal patterns in a homogeneous diffusive predator-prey system. J. Differ. Equ. 246(5), 1944–1977 (2009)
Tang, X., Song, Y.: Stability, Hopf bifurcations and spatial patterns in a delayed diffusive predator-prey model with herd behavior. Appl. Math. Comput. 254(C), 375–391 (2015)
Yang, R., Wei, J.: Stability and bifurcation analysis of a diffusive prey-predator system in Holling type III with a prey refuge. Nonlinear Dyn. 79(1), 631–646 (2015)
Soresina, C., Groppi, M., Buffoni, G.: Dynamics of predator-prey models with a strong Allee effect on the prey and predator-dependent trophic functions. Nonlinear Anal. Real World Appl. 30, 143–169 (2016)
Jana, S., Guria, S., Das, U., et al.: Effect of harvesting and infection on predator in a prey-predator system. Nonlinear Dyn. 81(1–2), 1–14 (2015)
Wang, J., Shi, J., Wei, J.: Dynamics and pattern formation in a difusive predator-prey system with strong Allee efect in prey. J. Differ. Equ. 251(4–5), 1276–1304 (2011)
Yang, R., Wei, J.: Bifurcation analysis of a diffusive predator-prey system with nonconstant death rate and Holling III functional response. Chaos Solitons Fractals 70(8), 1–13 (2015)
Yan, X.P., Zhang, C.H.: Stability and turing instability in a diffusive predator-prey system with Beddington–DeAngelis functional response. Nonlinear Anal. Real World Appl. 20(20), 1–13 (2014)
Tang, X., Song, Y.: Stability, Hopf bifurcations and spatial patterns in a delayed diffusive predator-prey model with herd behavior. Appl. Math. Comput. 254, 375–391 (2015)
Leslie, P.H.: Some further notes on the use of matrices in population mathematics. Biometrika 35(3–4), 1024–8 (1947)
Leslie, P.H., Gower, J.C.: The properties of a Stochastic model for the predator-prey type of interaction between two species. Biometrika 47(3–4), 219–234 (1960)
Chen, S., Shi, J., Wei, J.: Global stability and Hopf bifurcation in a delayed diffusive Leslie–Gower predator-prey system. Int. J. Bifurc. Chaos 22(3), 379–397 (2012)
Aziz-Alaoui, M.A., Okiye, M.D.: Boundedness and global stability for a predator-prey model with modified Leslie–Gower and Holling-type II schemes. Appl. Math. Lett. 16(7), 1069–1075 (2003)
Nindjin, A.F., Aziz-Alaoui, M.A., Cadivel, M.: Analysis of a predator-prey model with modified Leslie–Gower and Holling-type II schemes with time delay. Nonlinear Anal. Real World Appl. 7(5), 1104–1118 (2006)
Yafia, R., Adnani, F.E., Alaoui, H.T.: Limit cycle and numerical similations for small and large delays in a predator-prey model with modified Leslie-Gower and Holling-type II schemes. Nonlinear Anal. Real World Appl. 9(5), 2055–2067 (2008)
Dai, G., Tang, M.: Coexistence region and global dynamics of a harvested predator-prey system. Siam J. Appl. Math. 58(1), 193–210 (1998)
Kar, T.K., Ghorai, A.: Dynamic behaviour of a delayed predator-prey model with harvesting. Appl. Math. Comput. 217(22), 9085–9104 (2011)
Das, T., Mukherjee, R.N.C.K.S.: Bioeconomic harvesting of a prey-predator fishery. J. Biol. Dyn. 3(5), 447–62 (2009)
Yuan, R., Jiang, W., Wang, Y.: Saddle-node-Hopf bifurcation in a modified Leslie–Gower predator-prey model with time-delay and prey harvesting. J. Math. Anal. Appl. 422(2), 1072–1090 (2015)
Zhao, H., Huang, X., Zhang, X.: Turing instability and pattern formation of neural networks with reaction-diffusion terms. Nonlinear Dyn. 76(1), 115–124 (2014)
Yang, R., Song, Y.: Spatial resonance and Turing–Hopf bifurcations in the Gierer–Meinhardt model. Nonlinear Anal. Real World Appl. 31, 356–387 (2016)
Zheng, Q., Shen, J.: Turing instability in a gene network with cross-diffusion. Nonlinear Dyn. 78(2), 1301–1310 (2014)
Shen, J.: Canard limit cycles and global dynamics in a singularly perturbed predator–prey system with non-monotonic functional response. Nonlinear Anal. Real World Appl. 31, 146–165 (2016)
Adak, D., Bairagi, N.: Complexity in a predator-prey-parasite model with nonlinear incidence rate and incubation delay. Chaos Solitons Fractals 81, 271–289 (2015)
May, Robert M.: Time-delay versus stability in population models with two and three trophic levels. Ecology 54(2), 315–325 (1973)
Song, Yongli, Wei, Junjie: Local Hopf bifurcation and global periodic solutions in a delayed predator-prey system. J. Math. Anal. Appl. 301(1), 1–21 (2005)
Pal, P.J., Mandal, P.K., Lahiri, K.K.: A delayed ratio-dependent predator-prey model of interacting populations with Holling type III functional response. Nonlinear Dyn. 76(1), 201–220 (2014)
Wu, J.: Theory and Applications of Partial Functional-Differential Equations. Springer, New York (1996)
Hassard, B.D., Kazarinoff, N.D., Wan, Y.H.: Theory and Applications of Hopf Bifurcation. Cambridge University Press, Cambridge (1981)
Acknowledgments
The authors wish to express their gratitude to the editors and the reviewers for the helpful comments. This research is supported by the Fundamental Research Funds for the Central Universities, National Nature Science Foundation of China (No.11601070) and Heilongjiang Provincial Natural Science Foundation (No.A2015016).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Yang, R., Zhang, C. Dynamics in a diffusive modified Leslie–Gower predator–prey model with time delay and prey harvesting. Nonlinear Dyn 87, 863–878 (2017). https://doi.org/10.1007/s11071-016-3084-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11071-016-3084-7