
Abstract
Considering a good pest control program should reduce the pest to levels acceptable to the public, we investigate the threshold harvesting policy on pests in two predator–prey models. Both models are nonsmooth and the aim of this paper is to provide how threshold harvesting affects the dynamics of the two systems. When the harvesting threshold is larger than some positive level, the harvesting does not affect the ecosystem; when the harvesting threshold is less than the level, the model has complex dynamics with multiple coexistence equilibria, limit cycle, bistability, homoclinic orbit, saddle-node bifurcation, transcritical bifurcation, subcritical and supercritical Hopf bifurcation, Bogdanov–Takens bifurcation, and discontinuous Hopf bifurcation. Firstly, we provide the complete stability analysis and bifurcation analysis for the two models. Furthermore, some numerical simulations are given to illustrate our results. Finally, it is found that harvesting lowers the level of both species for natural enemy–pest system while raises the densities of both species for the pest–crop system. It is seen that the threshold harvesting policy of the enemy system is more effective than the crop system.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Pest is an important constraint to crop production worldwide, such as codling moth on apples, or boll weevil on cotton, and cause serious losses in yield and quality of cultivated plants. Therefore, farmers have been evolving a wide range of strategies for combating the various pests suffered by crops, and growing understanding of the interactions between the pest and crops or the pest natural enemy has enabled us to develop a wide array of measures for the control of pests. Such experiences have led to the development of effective and economical pest management programs [1–6]. Headley [6] has defined the term economic Threshold (T) as measuring the pest population levels at which pest control strategies should be initiated. More generally, T is usually interpreted as the number of insect pests in field when control actions must be taken to prevent the economic injury level from being reached and exceeded, where the level is the lowest pest population density that will cause economic damage [7–9].
Harvest management, including caching, the mixture of sugar–acetic acid–ethanol, sticky insect glue, frequency vibrational lamp, which may trap and kill many kinds of pest, is a traditional no pollution method of pest control. Most research has focused attention on the analysis and modeling of biological systems with harvesting. A classical predator–prey model with harvest management is as follows:

where x,y denote the density of prey population and predator population, respectively. The parameter r is the intrinsic growth rate and k is the environmental carrying capacity of prey population. The constant β(>0) is the maximum uptake rate for prey species, β 1(>0) denotes the ratio of biomass conversion (satisfying the obvious restriction 0<β 1<β) and d(>0) is the natural death rate of prey species. The term \(\frac{\beta xy}{\alpha +x}\) represents the functional response for the eating of prey by predator and α is the half saturation constant for a Holling type II functional response [10, 11], which contributes toward the growth of prey species. H(y) is the harvesting function. Some authors [12–14] have studied the predator–prey model with nonzero constant harvesting, H(y)=h, while others [15–17] have discussed a class of predator–prey models under constant proportion harvesting function, H(y)=hy. From these literatures, we note that over exploitation would result in the extinction of the predator population. However, complete eradication of the pest species is generally not possible, nor is it biologically or economically desirable. Therefore, a good pest control program should reduce the size of the pest population to levels acceptable to the public. This implies that there is an economic threshold T above, which the financial damage is sufficient to justify using such harvesting measures [18, 19]. Besides, we also need to point out that the harvesting starts at t=0, independence of the population size, is not realistic either. For practical reasons, threshold harvesting policy considers starting harvesting only when a population has reached threshold T.
In this paper, we consider continuous threshold harvesting policy for two predator–prey models. It works as follows: When a population is above a certain level or threshold T, harvesting occurs; when the population falls below that level, harvesting stops. The policy was first studied by Collie and Spencer [20], and additional analysis has been done since then by [21–24] and referenced therein. In this regard, threshold harvesting policy considers starting harvesting only when the pest population has reached a certain threshold level T. Classically, such harvesting function is defined as

In this way, once the pest population passes the size T, then harvesting starts and increases smoothly to a limit value h. We believe that this harvesting function is more sound from the biological viewpoint. Thus, the threshold harvesting policy is acceptable in economic terms to people while keeping the pest species which is harvested from extinction.
In this paper, we shall consider two predator–prey systems. For the first system, prey (pest) is harvested while predator (natural enemy )is protected. In practice, we control the pest population size by harvesting when the amount of the prey (pest) reaches a threshold. We implement the harvesting threshold T and the harvesting function (1.2) on the following model:

In the second system, predator (pest) is harvested while prey (crop) is protected.

Here, we observe that the second Eqs. (1.3) and (1.4) are always negative if β 1<d. So, we assume that β 1−d>0.
The aim of this paper is to provide how harvesting threshold affect the dynamics of the ecosystem. When the harvesting threshold is larger than some positive level, the harvesting policy does not affect the ecosystem; when the harvesting threshold is less than the level, the model has far richer dynamics. We will show that model (1.4) has at most three equilibria and model (1.4) has at most four equilibria in \(R^{2}_{+}\), and can exhibit numerous kinds of bifurcation phenomena, including the bifurcation of cusp type of codimension 2 (i.e., Bogdanov–Takens bifurcation), the subcritical and supercritical Hopf bifurcations [25, 26]. In particular, prey and predator species in model cannot become extinct simultaneously (mutual extinction) for all values of parameters and initial values, i.e., positive harvesting rate h can prevent mutual extinction. Prey and predator species can coexist in a positive equilibrium (or a stable limit cycle, or a unstable homoclinic loop) for some values of parameters and initial values, respectively. These results reveal far richer dynamics compared to the model (1.1) with no harvesting.
The organization of this paper is as follows. Firstly, we give and briefly describe our models. Then we provide the complete stability analysis both from local and global point of view, a rigorous bifurcation analysis including discontinuous Hopf bifurcation and some numerical analysis (in order to illustrate our results) for the two models in the next two sections, respectively. Lastly, we end the paper with a conclusion in Sect. 4.
2 Stability analysis
In this section, we give a qualitative analysis of system (1.3). From the standpoint of biology, we are only interested in the dynamics of model (1.3) in the closed first quadrant \(R_{+}^{2}\). Thus, we consider the biologically meaningful initial condition x(0)=x 0>0,y(0)=y 0>0. Regarding the boundedness of the solution for the model system (1.3), we state the following theorem.
Theorem 2.1
All the solutions of system (1.3) with the positive initial conditions (x 0,y 0) are uniformly bounded.
Proof
Since T>0, it follows that the positive x-axis and y-axis are both invariant. Let M(t)=x+βy/β 1. Using the relations, derivative of M with respect to (1.3) takes the form,

Since 0≤H(x)≤h for x≥0, there exists L such that
The right side of the above inequality is bounded for all \((x, y)\in R_{+}^{2}\). By using the differential inequality, it follows that
thus as t→+∞, 0<M<L/d. That is, solutions stay in
By the definition of M(t), it is known that there exists a constant M 1>0 such that x(t)≤M 1 and y(t)≤M 1 for t large enough. The proof is completed. □
Next, we discuss steady states of system (1.3). Since the threshold T>0, the extinction equilibria E 0(0,0) always exists for any parametric value. For the purpose of avoiding total population declines to zero as time goes to infinity, the stability of the extinction equilibrium is firstly investigated, which is determined by the nature of eigenvalues of the Jacobian matrix

Clearly, the eigenvalues are r, −d. The extinction equilibrium E 0 is a saddle point with unstable manifold in x-direction and stable manifold in y-axis.
Theorem 2.2
The extinction equilibrium E 0 is saddle with unstable manifold in x-direction and stable manifold in y-direction.
Remark 2.1
As we know, over exploitation would result in the extinction of the population. However, since threshold harvesting policy is starting only when the prey population reaches the threshold T, the extinction equilibrium is always unstable. That is at least one of the two populations persists and is not extinct for sufficiently large time. So, threshold harvesting policy should ensure the sustainability of system (1.3).
In the following sections, we only discussed the two cases of boundary equilibrium and coexistence equilibrium. We now examine the nullclines of the system:

By the properties of the function H(x), the system (1.3) has only one predator-free equilibrium E 0(k,0) when T≥k while it is intricate when T<k. Let

When T<x<k, the predator-free equilibrium \((\overline{x}, 0)\) satisfies the equation

So, there are at most three predator-free equilibria: \(E_{1}^{0}(\overline{x}_{1}, 0)\), \(E_{2}^{0}(\overline{x}_{2}, 0)\), and \(E_{3}^{0}(\overline{x}_{3}, 0)\) where \(\overline{x}_{1}\), \(\overline{x}_{2}\), \(\overline{x}_{3}\) are larger than T. In fact, we show this case in Fig. 1. Suppose that \(\overline{x}_{1}>\overline{x}_{2}>\overline{x}_{3}>T\). Derivative of F(x) with respect to x takes the form
So, we have \(F'(\overline{x}_{1})<0\), \(F'(\overline{x}_{2})>0\) and \(F'(\overline{x}_{3})<0\). If there are only two predator-free equilibria \(E_{1}^{0}(\overline{x}_{1}, 0)\) and \(E_{2}^{0}(\overline{x}_{2}, 0)\), then \(F'(\overline{x}_{1})<0\) and \(F'(\overline{x}_{2})=0\). If there is only one predator-free equilibrium \(E_{1}^{0}(\overline{x}_{1}, 0)\), then \(F'(\overline{x}_{1})<0\).

The existence of the free-predator equilibrium
For the coexistence equilibrium E ∗=(x ∗,y ∗), it follows that x ∗=dα/(β 1−d) and y ∗ is always negative if x ∗>k, and the system (1.3) doesn’t exist coexistence equilibrium. So, we will consider two cases as below: dα/(β 1−d)<k and dα/(β 1−d)≥k.
2.1 The case dα/(β 1−d)<k
For dα/(β 1−d)<k, system (1.3) has one extinction equilibrium E 0=(0,0) and one coexistence equilibrium E ∗=(x ∗,y ∗), and has at most three predator-free equilibria \(E_{i}^{0}=(\overline{x}_{i},0),\ i=1,2,3\) or has at least one predator-free equilibrium \(E_{1}^{0}=(\overline{x}_{1},0)\). Next, we examine the stability of equilibria for three cases of the threshold T.
2.1.1 0<T≤dα/(β 1−d)
Some basic facts are given for the case which is simply showed in Fig. 2.

The nullclines and equilibria for 0<T≤dα/(β 1−d)
1. \(\overline{x}_{i}\) satisfies the following equation:

2. x ∗=dα/(β 1−d) and y ∗ satisfies

For the boundary equilibria \(E_{i}^{0}\), the characteristic equation is

(a) If the system has only one predator-free equilibrium \(E_{1}^{0}\), then one of the eigenvalues \(F'(\overline{x}_{1})<0\) and the other eigenvalues is positive if \(\overline{x}_{1}>d\alpha/(\beta_{1}-d)\). Thus, \(E_{1}^{0}\) is saddle with stable manifold in x-axis and unstable manifold in

If \(\overline{x}_{1}<d\alpha/(\beta_{1}-d)\), then both the eigenvalues are negative, hence \(E_{1}^{0}\) is asymptotically stable (A.S.). In fact, it is also globally asymptotically stable (G.A.S.). Otherwise, system (1.3) exists a closed orbit in B 1 from Theorem 2.1. Then there must exist an equilibrium in the closed orbit. This is impossible, since the coexistence equilibrium E ∗ does not exist (y ∗<0). Therefore, system (1.3) does not exist limit cycle. Certainly, if the coexistence equilibrium E ∗ exists, then the predator-free equilibrium \(E_{1}^{0}\) is saddle.
For the coexistence equilibrium E ∗=(x ∗,y ∗) where x ∗>T, the Jacobian matrix takes the form

Its determinant and trace are respectively

If \(\operatorname{Tr}J(E^{*})>0\), then the eigenvalues of J(E ∗) have positive real parts. Hence, E ∗ is unstable, and the existence of the limit cycle is guaranteed by the Poincaré–Bendixson theorem and Theorem 2.1.
If \(\operatorname{Tr}J(E^{*})<0\), then all the eigenvalues of J(E ∗) have negative real parts, hence the equilibrium E ∗ is A.S. Further, if \((\operatorname{Tr}J(E^{*}))^{2}-4\operatorname{Det}J(E^{*})\geq 0\) then the equilibrium E ∗ is stable node; if \((\operatorname{Tr}J(E^{*}))^{2}-4\operatorname{Det}J(E^{*})<0\), then the equilibrium E ∗ is stable focal point. Next, we can use the Lyapunov–LaSalle theorem to give a sufficient condition under which it is G.A.S. Consider

Its derivative along the solutions of (1.3) is

It follows for x≤T that

Clearly, if βy ∗/(α 2+αx ∗)−r/k<0 then dV/dt<0. Further, if x>T, then

So, it follows that dV/dt<0 if βy ∗/(α(α+x ∗))+1/(h+x ∗−T)−r/k<0. In summary, the inequalities βy ∗/(α(α+x ∗))+1/(h+x ∗−T)−r/k<0, βy ∗/(α(α+x ∗))−r/k<0 and \(\operatorname{Tr}J(E^{*})<0\) are equivalent to
Hence, the Lyapunov–LaSalle theorem implies that all solutions ultimately approach the equilibrium E ∗ if (2.6) holds.
If \(\operatorname{Tr}J(E^{*})=0\), then the Jacobian matrix J(E ∗) has a pair of pure imaginary eigenvalues and the coexistence equilibrium E ∗ is a center-type equilibrium. Further, we consider k as bifurcation parameter, i.e., \(\operatorname{Tr}J(E^{*})=0\) when k=k ∗. In this case, Hopf bifurcation occurs. In fact, the two pure imaginary eigenvalues are

Hence, all the conditions for Hopf-bifurcations are satisfied. Thus, there exist small amplitude periodic solutions near E ∗.
If \(x^{*}=\overline{x}_{1}\), i.e., \(\frac{d\alpha}{\beta_{1}-d}=\overline{x}_{1}\), transcritical bifurcation appears. When \(x^{*}<\overline{x}_{1}\) and \(\operatorname{Tr}J(E^{*})<0\), from the above analysis we node that the interior equilibrium E ∗ is A.S. Besides, we had shown that \(E_{1}^{0}\) is a saddle when \(x^{*}<\overline{x}_{1}\). As x ∗ increases, it satisfies (2.6), then E ∗ is G.A.S. If x ∗ tends to \(\overline{x}_{1}\) from the left side, then E ∗ toward to \(E_{1}^{0}\), and \(\overline{x}_{1}=x^{*}\) i.e., E ∗ collides \(E_{1}^{0}\). Thus, \(E_{1}^{0}\) is saddle-node, and transcritical bifurcation appears. Interestingly, all orbits tends to \(E_{1}^{0}\) because the unstable manifold direction does not belong to B 1. Also, we can obtain that y′<0 in B 1 and there are only two boundary equilibria where E 0 is saddle with unstable manifold in x-direction and stable manifold in y-direction.
Summarizing the above discussions, we obtain the following theorem.
Theorem 2.3
If \(\overline{x}_{1}<x^{*}\), the predator-free equilibrium \(E_{1}^{0}\) is saddle with stable manifold in x-direction; if \(\overline{x}_{1}>x^{*}\), \(E_{1}^{0}\) is G.A.S.; if \(x^{*}=\overline{x}_{1}\), transcritical bifurcation appears and all orbits tends to \(E_{1}^{0}\). If \(\operatorname{Tr}J(E^{*})<0\), the coexistence equilibrium E ∗ is A.S.; further, if the inequality (2.6) holds E ∗ is G.A.S.; if \(\operatorname{Tr}J(E^{*})>0\), E ∗ is unstable and system (1.3) has at least one limit cycle; if \(\operatorname{Tr}J(E^{*})=0\), Hopf bifurcation occurs.
(b) If the system has three predator-free equilibria \(E_{i}^{0}, i=1,2,3\), then \(F'(\overline{x}_{1})<0\), \(F'(\overline{x}_{2})>0\) and \(F'(\overline{x}_{3})<0\).
If \(x^{*}<\overline{x}_{3}\), at least one eigenvalues of \(J(E^{0}_{i})\) is always positive, thus \(E_{i}^{0}, i=1,3\) is saddle, \(E_{2}^{0}\) is an unstable node. In addition, since \(\operatorname{Tr}J(E^{*})<0\) always holds, the coexistence equilibrium E ∗ is A.S.
If \(\overline{x}_{3}<x^{*}<\overline{x}_{2}\), the eigenvalues of \(J(E^{0}_{3})\) are negative and system (1.3) has no coexistence equilibrium. So, it is impossible that system exists a closed orbit in B 1. Hence, the stable node \(E^{0}_{3}\) is G.A.S., \(E_{1}^{0}\) is saddles, \(E_{2}^{0}\) is unstable node.
If \(\overline{x}_{2}<x^{*}<\overline{x}_{1}\), the eigenvalues of \(J(E^{0}_{3})\) are negative and one eigenvalues of \(J(E^{0}_{i}),\,i=2,3\) is positive, thus the stable node \(E^{0}_{3}\) is G.A.S., \(E_{1}^{0}\) is saddles with the stable manifold in x-direction, \(E_{2}^{0}\) is saddle with the unstable manifold in x-direction. In addition, the stability of E ∗ is completely same as in above discussion of (a).
If \(\overline{x}_{1}<x^{*}<k\), the coexistence equilibrium does not exist; the eigenvalues of \(J(E^{0}_{i})\,(i=1,3)\) are negative, one eigenvalue of \(J(E^{0}_{2})\) is positive, and the other is negative. Thus, \(E^{0}_{i}\,(i=1,3)\) are a stable node and \(E_{2}^{0}\) is saddle with the stable manifold in x-direction. Besides, the first quadrant \(R_{+}^{2}\) is divided into two regions by the stable manifold of \(E_{2}^{0}\): in the first region, \(E^{0}_{1}\) is G.A.S and in the second region \(E^{0}_{3}\) is G.A.S (bistability).
If \(x^{*}=\overline{x}_{i}\) (i=1,2,3), transcritical bifurcation appears.
(c) If the system have only two predator-free equilibria \(E_{i}^{0}, i=1,2\), i.e., \(F'(\overline{x}_{1})<0\) and \(F'(\overline{x}_{2})=0\), then saddle-node bifurcation appears. When \(F'(\overline{x}_{2})>0\), there are three predator-free equilibria \(E_{2}^{0}\), \(E_{3}^{0}\), and \(E_{1}^{0}\). As the value of \(F'(\overline{x}_{2})\) decreases, the equilibrium \(E_{3}^{0}\) tends to the equilibrium \(E_{2}^{0}\). It is equal to zero, i.e., \(F'(\overline{x}_{2})=0\) implies that \(E_{3}^{0}\) collides \(E_{2}^{0}\). In particular, system (1.3) has only one predator-free equilibrium \(E_{1}^{0}\) for F′(x)<0. Therefore, the system undergoes a saddle-node bifurcation.
Finally, we discuss stability of the coexistence equilibrium E ∗ for the case T=dα/(β 1−d)=x ∗. For convenience, we give the following two systems with harvesting and without harvesting, respectively,

When T=dα/(β 1−d)=x ∗, then the coexistence equilibrium \(E^{*}_{a}\) of (2.7(a)), is equal to the coexistence equilibrium \(E^{*}_{b}\) of (2.7(b)), \(E^{*}_{a}=E^{*}_{b}\). Accordingly, the determinant and trace of the Jacobian matrix \(J(E^{*}_{i})\) are \(\operatorname{Tr}_{i}\) and \(\operatorname{Det}_{i}\) (i=a,b), respectively. Based on the above analysis, we can obtain

We summarize the dynamical behavior of system (1.3) at the coexistence equilibrium E ∗ when T=x ∗ as follows, which is shown in Fig. 3.

The dynamical properties of (1.3) at the coexistence equilibrium E ∗ when T=x ∗
(1) If Tr b >0, then Tr a >0, \(E^{*}_{a}\) and \(E^{*}_{b}\) are unstable. Furthermore,
(i) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}\geq0\), then both \(E^{*}_{a}\) and \(E^{*}_{b}\) are unstable node; hence, E ∗ is unstable node and the existence of the limit cycle is guaranteed by Poincaré–Bendixson theorem (see Fig. 3(a)).
(ii) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}<0\) and \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}\geq 0\), then \(E^{*}_{a}\) is an unstable node and \(E^{*}_{b}\) is unstable focus; hence, E ∗ is unstable and the existence of the limit cycle is guaranteed by the Poincaré–Bendixson theorem (see Fig. 3(b)).
(iii) if \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}<0\), then both \(E^{*}_{a}\) and \(E^{*}_{b}\) are unstable focus; in this case, it is intricate, that is, the stability of E ∗ may be stable or unstable or system (1.3) has periodic solutions (see Fig. 3(c)).
(2) If Tr b <0 and Tr a >0, then \(E^{*}_{a}\) is unstable and \(E^{*}_{b}\) is stable. Furthermore,
(i) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}\geq 0\) and \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}\geq 0\), then \(E^{*}_{a}\) is an unstable node and \(E^{*}_{b}\) is a stable node, system (1.3) has homoclinic loops (see Fig. 3(d)).
(ii) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}\geq 0\) and \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}<0\), then \(E^{*}_{a}\) is unstable focus and \(E^{*}_{b}\) is a stable node; hence, E ∗ is unstable and the existence of the limit cycle is guaranteed by the Poincaré–Bendixson theorem.
(iii) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}<0\) and \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}\geq 0\), then \(E^{*}_{a}\) is an unstable node and \(E^{*}_{b}\) is stable focus; hence, E ∗ is stable.
(iv) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}<0\) and \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}<0\), then \(E^{*}_{a}\) is unstable focus and \(E^{*}_{b}\) is stable focus. In this case, it is intricate, that is, the stability of E ∗ may be stable or unstable or system (1.3) has periodic solutions.
(3) If Tr a <0, then \(E^{*}_{a}\) and \(E^{*}_{b}\) are stable equilibria. Furthermore,
(i) if \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}\geq 0\), then both \(E^{*}_{a}\) and \(E^{*}_{b}\) are stable node; hence, E ∗ is stable node (see Fig. 3(e)).
(ii) if \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}<0\) and \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}\geq 0\), then \(E^{*}_{a}\) is stable focus and \(E^{*}_{b}\) is stable node, hence, E ∗ is stable (see Fig. 3(f)).
(iii) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}<0\), then both \(E^{*}_{a}\) and \(E^{*}_{b}\) are stable focus; in this case, it is intricate, that is, the stability of E ∗ may be stable or unstable or system (1.3) has periodic solutions.
Furthermore, we assume one of the coexistence equilibria \(E^{*}_{a}\) and \(E^{*}_{b}\) is stable focus and the other is unstable focus, that is the case of (2.(iv)). Let J −(E ∗) and J +(E ∗) denote the left and right Jacobian matrix of system (1.3) at the coexistence equilibrium E ∗ when T=x ∗, respectively,

where \(H'_{+}(x)\) is the right derivative and \(H'_{-}(x)\) is the left derivative, and \(H'_{\mp}(x)\) denotes \(H'_{+}(x)\) or \(H'_{-}(x)\). It is obvious that \(H'_{-}(x)=H'_{+}(x)\) when x ∗≠T. Without loss of generality, the real parts of eigenvalues of J −(E ∗) are all negative and there exists at least one eigenvalue λ of J +(E ∗) such that the real part of λ is positive. Then there exists a jump from J −(E ∗) to J +(E ∗). Therefore, it is difficult to study the stability of equilibrium E ∗ when T=x ∗. Here, we introduce the generalized Jacobian matrix of Clarke [27–29]. In our case, the generalized Jacobian matrix J at E ∗ when T=x ∗ is

Let J(q)=(1−q)J −(E ∗)+qJ +(E ∗) and suppose the characteristic equation of J(q) is as follows:
When q=0, the real parts of roots of (2.9) are negative, and when q=1 there exists a negative real parts root and a positive real parts root. Directive computing shows that a 0(q)≠0 which implies 0 is not the root of (2.9). Hence, there is a 0<q ∗<1 such that Eq. (2.9) has a pair of conjugate eigenvalues ±ωi where ω>0 when q=q ∗. Therefore, the discontinuous bifurcation [28] occurs; this implies there exists an asymptotically stable limit cycle.
In the following subsections, similarly discussion will be done for the case T=x ∗. So, we shall omit it.
2.1.2 dα/(β 1−d)<T<k
Some basic facts are given for this case which is simply showed in Fig. 4.

The nullclines and equilibria for dα/(β 1−d)<T<k
1. \(\overline{x}_{i}, i=1,2,3\) satisfies the following equation:
2. \(y^{\ast}=\frac{r}{\beta}(1-\frac{x^{\ast}}{k})(\alpha+x^{\ast})\), and \(x^{\ast}=\frac{d\alpha}{\beta_{1}-d}<T\).
(a) If there is only one the predator-free equilibrium \(E_{1}^{0}\), the stability of which is completely same as in Sect. 2.1.1; we omit the details.
For the coexistence equilibrium E ∗=(x ∗,y ∗) where x ∗>T, the determinant and trace of the Jacobian matrix J(E ∗) are respectively

If k>2x ∗−α, then the equilibrium E ∗ is unstable. The existence of the limit cycle is guaranteed by the Poincaré–Bendixson theorem and Theorem 2.1.
If k<2x ∗+α, then the equilibrium E ∗ is A.S. It is easy to compute that

Further, if \((\operatorname{Tr}J(E^{*}))^{2}-4\operatorname{Det}J(E^{*})\geq 0\), then the equilibrium E ∗ is a stable node; if \((\operatorname{Tr}J(E^{*}))^{2}-4\operatorname{Det}J(E^{*})<0\) then the equilibrium E ∗ is stable focal point. Next, we can use the Lyapunov–LaSalle theorem to give a sufficient condition under which it is G.A.S. Consider

Its derivative along the solutions of (1.3) is
For the two cases of x≤T and x>T>x ∗, it follows that
Since the inequalities k<2x ∗+α and k<x ∗+α are equivalent to k<x ∗+α. Hence, the Lyapunov–LaSalle theorem implies that E ∗ is G.A.S. if k<x ∗+α.
If k=2x ∗+α, then the Jacobian matrix J(E ∗) has a pair of pure imaginary eigenvalues \(\lambda =\pm\frac{\beta_{1}\alpha \beta x^{\ast}y^{\ast}}{(\alpha+x^{\ast})^{3}}i\), and the coexistence equilibrium E ∗ is a center-type equilibrium. Further, we have
Hence, Hopf bifurcation occurs, that is, there exist small amplitude periodic solutions near E ∗. We will summarize the discussions as the following theorem.
Theorem 2.4
The predator-free equilibrium \(E_{1}^{0}\) is saddle with stable manifold in x-direction. If k<2x ∗+α, the coexistence equilibrium E ∗ is A.S; further, if k<x ∗+α, E ∗ is G.A.S. If k>2x ∗+α, E ∗ is unstable and system (1.3) has at least one limit cycle. If \(\operatorname{Tr}J(E^{*})=0\), Hopf bifurcation occurs.
(b) If the system has three predator-free equilibria \(E_{i}^{0},\ i=1,2,3\), then \(F'(\overline{x}_{1})<0\), \(F'(\overline{x}_{2})>0\), and \(F'(\overline{x}_{3})<0\). It follows from \(x^{*}<\overline{x}_{3}\) that one eigenvalues of \(J(E^{0}_{i})\) is always positive, thus \(E_{i}^{0},\ i=1,3\) is saddle, \(E_{2}^{0}\) is unstable node. In addition, the stability of E ∗ is completely same as the above discussion, we omit the details.
(c) If the system has only two predator-free equilibria \(E_{i}^{0},\ i=1,2\), i.e., \(F'(\overline{x}_{1})<0\) and \(F'(\overline{x}_{2})=0\), then saddle-node bifurcation appears. When \(F'(\overline{x}_{2})>0\), there are three predator-free equilibria \(E_{1}^{0}\), \(E_{2}^{0}\), and \(E_{3}^{0}\). As the value of \(F'(\overline{x}_{1})\) decreases, the equilibrium \(E_{3}^{0}\) tends to the equilibrium \(E_{2}^{0}\), and it is equal to zero, i.e., \(F'(\overline{x}_{2})=0\) implies that \(E_{1}^{0}\) collides \(E_{2}^{0}\). In particular, system (1.3) has only one equilibrium \(E_{1}^{0}\) for \(F'(\overline{x}_{2})<0\). Therefore, the system undergoes a saddle-node bifurcation.
2.1.3 dα/(β 1−d)<k≤T
Some basic facts are given for this case, which are simply shown in Fig. 5.

The nullclines and equilibria for dα/(β 1−d)<k≤T
1. There is only one predator-free equilibrium E 0(k,0).
2. \(y^{\ast}=\frac{r}{\beta}(1-\frac{x^{\ast}}{k})(\alpha +x^{\ast})\), and \(x^{\ast}=\frac{d\alpha}{\beta_{1}-d}<k\).
By Theorem 2.1, we proved that any solution of system (1.3) starting outside B 1 either enters into B 1 at some finite time, say t 0>0, and then it remains in its interior B 1 for all t>t 0 or tends to the boundary equilibrium E 0∈∂Ω. According to the theory of a classical predator–prey system

we arrive at the following result.
Theorem 2.5
The two boundary equilibria E 0, E 0 are saddles. If k<2x ∗+α, the coexistence equilibrium E ∗ is G.A.S. If k>2x ∗+α, E ∗ is unstable and system (1.3) has at least one limit cycle. If k=2x ∗+α, Hopf bifurcation occurs.
A detailed analysis in this direction can be found in [30]. If k>2x ∗+α, the coexistence equilibrium E ∗ is unstable and system has at least one limit cycle. Indeed, one can show that this limit cycle is unique and G.A.S.
Theorem 2.6
For x≠dα/(β 1−d), system (1.3) has a unique globally asymptotically stable limit cycle on the positive quadrant if k>2x ∗+α.
Proof
Let g(x)=r(1−x/k), p(x)=βx/(α+x), q(x)=β 1 x/(α+x). Then

Using a uniqueness theorem in [31], we conclude that there is a unique G.A.S limit cycle if k≥2x ∗+α. □
2.2 The case k≤dα/(β 1−d)
For k≤dα/(β 1−d), system (1.3) has no coexistence equilibrium E ∗, has one extinction equilibrium E 0(0,0), and has at least one predator-free equilibrium \(E_{1}^{0}(\overline{x}_{1},0)\) or has at most three predator-free equilibria \(E_{i}^{0}(\overline{x}_{i},0);\ i=1,2,3\).
2.2.1 k≤T
When k≤T, basic facts in this case are as follows:
1. The extinction equilibrium E 0 is saddle with unstable manifold in the x-direction and stable manifold in the y-direction.
2. There is no coexistence equilibrium E ∗, one predator-free equilibrium \(E_{1}^{0}\).
If k<dα/(β 1−d), the predator-free equilibrium \(E_{1}^{0}\) is G.A.S. Since the eigenvalues of \(J(E_{1}^{0})\) are both negative, the system has no coexistence equilibrium in B 1. Again, if k=dα/(β 1−d), transcritical bifurcation appears. If dα/(β 1−d)<k<(β 1+d)α/(β 1−d), E ∗ is G.A.S and \(E_{1}^{0}\) is saddle, the unstable manifold of \(E_{1}^{0}\) directs inward B 1. Thus, there exists a heteroclinic orbit connecting E ∗ and \(E_{1}^{0}\).
Theorem 2.7
The extinction equilibrium E 0 is saddle. If k<dα/(β 1−d), the predator-free equilibrium \(E_{1}^{0}\) is G.A.S; if k=dα/(β 1−d), \(E_{1}^{0}\) is a saddle-node and transcritical bifurcation appears.
2.2.2 T<k≤dα/(β 1−d)
When T<k, basic facts in this case are as follows:
1. There is no coexistence equilibrium. For the predator-free equilibrium \(E_{i}^{0}(\overline{x}_{i}, 0)\), we have \(r\overline{x}_{i}(1-\frac{\overline{x}_{i}}{k}) - \frac{h(\overline{x}_{i}-T)}{h+\overline{x}_{i}-T}=0\), and \(T<\overline{x}_{i}<k\), \(\overline{x}_{i}<\frac{d\alpha}{\beta_{1}-d}\).
3. The extinction equilibrium E 0 is saddle with unstable manifold in x-direction and stable manifold in y-direction.
As mentioned before, the eigenvalues of \(J(E_{i}^{0})\) are

In this case \(\overline{x}_{i}<d\alpha/(\beta_{1}-d)\), then λ 2<0.
(a) If there is only one predator-free equilibrium \(E_{1}^{0}\), then \(F'(\overline{x}_{1})<0\), i.e., λ 1<0. Thus, \(E_{1}^{0}\) is G.A.S. Since there is no coexistence equilibrium E ∗ in B 1 and E 0 is saddle.
(b) If there is three predator-free equilibria \(E_{i}^{0},\ i=1,2,3\), then \(F'(\overline{x}_{1})<0\), \(F'(\overline{x}_{2})>0\) and \(F'(\overline{x}_{3})<0\). Hence, \(E_{i}^{0}\) (i=1,3) is a A.S node and \(E_{2}^{0}\) is saddle. In this case, the first quadrant \(R_{+}^{2}\) is divided into two regions by the stable manifold of \(E_{2}^{0}\): in the first region \(E^{0}_{1}\) is G.A.S and in the second region \(E^{0}_{3}\) is G.A.S (bistability).
(c) If there is two predator-free equilibria \(E_{i}^{0},\ i=1,2\), then \(F'(\overline{x}_{1})<0\) and \(F'(\overline{x}_{2})=0\), hence \(E_{1}^{0}\) is a G.A.S node and system (1.3) exhibits saddle-node bifurcations. Let \(s=r-2r\overline{x}_{2}/k+\sqrt{r-2r\overline{x}_{2}/k}\) with \(\overline{x}_{2}<k/2\), and \(h^{*}=ks(\overline{x}_{2}-T)/(k+2r\overline{x}_{2}-kr)\). It is easy to compute that \(F'(\overline{x}_{2})=0\) is equivalent to h=h ∗. If h=h ∗, then λ 1=0. On the other hand, one can write \(\lambda_{2}=\beta_{1}\overline{x}_{2}/(\alpha+\overline{x}_{2})-d\), so that λ 2≠0 (so long as \(\overline{x}_{2}\neq d\alpha/(\beta_{1}-d)\)). We shift the equilibrium \((h^{*}, \overline{x}_{2}, 0)\) to the origin: ξ=h−h ∗, \(u=x-\overline{x}_{2}\), v=y, and the new system can be written as
Then we have

To make sure ∂ 2 F 1/∂u 2 (0,0,0)≠0, we need to require that \({h^{*}}^{2}\neq r(h^{*}+\overline{x}_{2}-T)^{3}/k\), which is true if \(3r(\overline{x}_{2}-T)/k>1\). Then we can conclude that there exists a saddle-node bifurcation [34] if h=h ∗, \(\overline{x}_{2}<k/2\) and \(3r(\overline{x}_{2}-T)/k>1\). In fact, the system has only one predator-free equilibrium \(E_{1}^{0}\) for h>h ∗, there is two equilibria \(E_{1}^{0}\) and \(E_{2}^{0}\) for h=h ∗, and there are three equilibria \(E_{1}^{0}\), \(E_{2}^{0}\) and \(E_{3}^{0}\) for h<h ∗.
Theorem 2.8
The system (1.3) exhibits saddle-node bifurcations if h=h ∗ with \(k/3r+T<\overline{x}_{2}<k/2\).
We continue to discuss the case T<dα/(β 1−d). From above discussion, we know that if there are three predator-free equilibria, then \(F'(\overline{x}_{1})<0\), \(F'(\overline{x}_{2})>0\) and \(F'(\overline{x}_{3})<0\). Besides, if \(\overline{x}_{i}<d\alpha/(\beta_{1}-d)\) (i=1,2,3) then \(E_{i}^{0}\,(i=1,3)\) are A.S and \(E_{2}^{0}\) is saddle. In this case, the first quadrant \(R_{+}^{2}\) is divided into two regions by the stable manifold of \(E_{1}^{0}\): in the first region limit cycle is G.A.S. and in the second region \(E^{0}_{3}\) is G.A.S. (bistability).
Further, observe that \(J(E_{i}^{0})\) has one zero eigenvalue precisely when \(\overline{x}_{i}=d\alpha/(\beta_{1}-d)\) and  , where a
12≠0 and a
11≠0 as long as \(F'(\overline{x}_{i})\neq 0\). The eigenvalue λ=0 of \(J(E_{i}^{0})\) has an eigenvector \(v^{T}=(1\ -\frac{a_{11}}{a_{12}})\), and a left eigenvector w
T=(0 1). If we write the right-hand side of system (1.3) as f(x,y), then one gets \(w^{T} f_{d}(\overline{x}, 0)=0\) and
, where a
12≠0 and a
11≠0 as long as \(F'(\overline{x}_{i})\neq 0\). The eigenvalue λ=0 of \(J(E_{i}^{0})\) has an eigenvector \(v^{T}=(1\ -\frac{a_{11}}{a_{12}})\), and a left eigenvector w
T=(0 1). If we write the right-hand side of system (1.3) as f(x,y), then one gets \(w^{T} f_{d}(\overline{x}, 0)=0\) and

Therefore, by Sotomayor’s theorem, one concludes that under the conditions established above, system (1.3) experiences a transcritical bifurcation. Thus, under some condition relating \(\overline{x}_{i}\) to h and T, we have an explicit value of the predator death rate d which is such that \(\overline{x}_{i}=d\alpha/(\beta_{1}-d)\). When such bifurcation happens, and an exchange of stability between equilibria is possible.
Theorem 2.9
The system (1.3) exhibits transcritical bifurcations if \(\overline{x}_{i}=d\alpha/(\beta_{1}-d)\).
2.3 Numerical results
Our focus so far has been on the dynamics of the system (1.3). To facilitate the interpretation of our mathematical results in model (1.3), we proceed to investigate the complex case of system (1.3) has three predator-free equilibria by numerical simulations.
Let r=0.6, k=10, β=1.2, α=1, h=2, T=0.1, β=0.9. In this case, we consider x ∗=dα/(β 1−d) as bifurcation parameter. In fact, x ∗ is an increasing function of d, which has the range: 0–0.9. It is easy to verify that there is three predator-free equilibria \(E_{3}^{0}(0.273, 0)\), \(E_{2}^{0}(2.146, 0)\), \(E_{1}^{0}(5.688, 0)\). The extinction equilibrium E 0 is saddle.
(1) If d=0.1, then x ∗=0.125 and \(T<x^{*}<\overline{x}_{3}\). Thus, the coexistence equilibrium E ∗ is G.A.S., the predator-free equilibria: \(E_{i}^{0}\) (i=1,3) are saddle and \(E_{2}^{0}\) is unstable node, and there exist heteroclinic orbit connecting E ∗ and \(E_{i}^{0}\) (i=1,2,3), which is shown in Fig. 6(a).

(a) Let d=0.1. E ∗ is G.A.S, \(E_{i}^{0}\) (i=1,3) are saddle and \(E_{2}^{0}\) is unstable node, and there exist heteroclinic orbit connecting E ∗ and \(E_{i}^{0}\) (i=1,2,3). (b) When d=0.3, E ∗ does not exist, \(E_{3}^{0}\) is G.A.S., \(E_{2}^{0}\) is an unstable node and \(E_{3}^{0}\) is saddle, and there exist heteroclinic orbit connecting \(E^{0}_{3}\) and \(E_{i}^{0}\) (i=1,2)
(2) As d increase and d=0.3, it follows that x ∗=0.5 and \(\overline{x}_{3}<x^{*}<\overline{x}_{2}\), hence there is no E ∗, and \(E_{3}^{0}\) is G.A.S., \(E_{2}^{0}\) is unstable node and \(E_{1}^{0}\) is saddle and there exist heteroclinic orbit connecting \(E^{0}_{3}\) and \(E_{i}^{0}\) (i=1,2), which is shown in Fig. 6(b).
(3) When d=0.7, then x ∗=3.5 and \(\overline{x}_{2}<x^{*}<\overline{x}_{1}\) and \(\operatorname{Tr}J(E^{*})>0\). Thus, E ∗ is unstable node, \(E_{3}^{0}\) is G.A.S., \(E_{i}^{0}\) (i=2,3) are saddle, and there exist two heteroclinic orbit connecting E ∗ and \(E_{2}^{0}\), \(E^{0}_{3}\) and \(E_{1}^{0}\), which is shown in Fig. 7(a).

(a) When d=0.7, E ∗ is unstable node, \(E_{1}^{0}\) and \(E_{2}^{0}\) are saddle, \(E_{3}^{0}\) is G.A.S, and there exist two heteroclinic orbit connecting E ∗ and \(E_{2}^{0}\), \(E^{0}_{3}\) and \(E_{1}^{0}\). (b) When d=0.715, E ∗ is unstable focus and \(E_{1}^{0}\) and \(E_{2}^{0}\) are saddle. Thus, \(R_{+}^{2}\) is divided into two regions by unstable manifold of \(E_{2}^{0}\), which ultimately approach the stable limit cycle: in the first region limit cycle is G.A.S. and in the second region \(E^{0}_{3}\) is G.A.S. (bistability)
(4) When d=0.715, then x ∗=3.86 and \(\overline{x}_{2}<x^{*}<\overline{x}_{1}\) and \(\operatorname{Tr}J(E^{*})>0\). Thus, E ∗ is unstable focus, \(E_{3}^{0}\) is A.S., \(E_{i}^{0},\,(i=1,2)\) are saddle. Thus, the first quadrant \(R_{+}^{2}\) is divided into two regions by the stable manifold of \(E_{2}^{0}\): in the first region limit cycle is G.A.S. and in the second region \(E^{0}_{3}\) is G.A.S (bistability), which is shown in Fig. 7(b).
(5) Further increase d=0.72 will lead to the limit cycle is broken. It is easily to verify that x ∗=4 and \(\overline{x}_{2}<x^{*}<\overline{x}_{1}\) and \(\operatorname{Tr}J(E^{*})<0\). The equilibria \(E_{1}^{0}\) and \(E_{2}^{0}\) are saddle. Thus, \(R_{+}^{2}\) is divided into two regions by the stable manifold of \(E_{2}^{0}\): in the first region E ∗ is G.A.S. and in the second region \(E^{0}_{3}\) is G.A.S., which is shown in Fig. 8(a) (bistability).

(a) When d=0.72, the limit cycle is broken, \(E_{1}^{0}\) and \(E_{2}^{0}\) are saddle. Thus, \(R_{+}^{2}\) is divided into two regions by the unstable manifold of \(E_{2}^{0}\): in the first region E ∗ is G.A.S. and in the second region \(E^{0}_{3}\) is G.A.S (bistability). (b) When d=0.78, E ∗ does not exist and \(E_{2}^{0}\) is saddle. Thus, \(R_{+}^{2}\) is divided into two regions by the unstable manifold of \(E_{2}^{0}\): in the first region \(E_{1}^{0}\) is G.A.S. and in the second region \(E^{0}_{3}\) is G.A.S (bistability)
(6) If d=0.78, then x ∗=6.5 and \(\overline{x}_{1}<x^{*}<k\), hence there is E ∗, and the equilibrium \(E_{2}^{0}\) is saddle. Thus, the first quadrant \(R_{+}^{2}\) is divided into two regions by the stable manifold of \(E_{2}^{0}\): in the first region \(E_{1}^{0}\) is G.A.S. and in the second region \(E^{0}_{3}\) is G.A.S., which is shown in Fig. 8(b) (bistability).
We made the following observations.
(1) When the harvesting threshold is larger than the carrying capacity of the prey, T>k, the harvesting does not affect the ecosystem, that is, it is not necessary to control the prey, which cannot be viewed as the pest. By Theorem 2.1, all solutions of system (1.3) ultimately approach the region B 1 in which the dynamics of system (1.3) is equivalent to the classical predator–prey system (2.10). From the biological view point, the natural enemy has enough food for predation even if the prey is harvested, which is in line with reality.
(2) When the harvesting threshold for the pest is below the carrying capacity, T<k, the presence of threshold harvesting makes the dynamic behavior more complex with multiple equilibria, limit cycle, heteroclinic orbit, bistability, saddle-node bifurcation, transcritical bifurcation and Hopf bifurcation, which is shown in Figs. 5, 6, 7.
(3) Besides, when the harvesting threshold is below the carrying capacity, T<k, the system has one predator-free equilibrium E 0(k,0) in the absence of harvesting; the system has predator-free equilibria \(E^{0}(\overline{x},0)\) in the presence of harvesting, and \(\overline{x}<k\). Consequently, the harvesting decrease the density of pest species. Further, when the harvesting threshold \(\frac{d\alpha}{\beta_{1}-d}<T<k\), the asymptotic state of a solution can either be the coexistence equilibrium or a limit cycle around the equilibrium, which implies the equilibrium level is always below the threshold, that is the aim of the control is attained.
(4) Since harvesting starts only the prey population is above the threshold T, it is easily to compute
The continuous threshold harvesting decrease the density of predator species and this is happen due to the loss of food for predator species. Thus, harvesting also has impact on the natural enemy species.
(5) It is observed that the system has the bistability. So, the asymptotic state of a solution of the system can either be an equilibrium or a limit cycle which is shown in example 4. So, the extinction or survival of the predator (natural enemy) is possible depending on the initial size of the both species populations. In the example presented in this article, the coexistence equilibrium E ∗ and the predator-free equilibrium \(E^{0}_{3}\) are G.A.S in example 5, predator-free equilibria \(E^{0}_{3}\) and \(E^{0}_{1}\) are G.A.S in example 6. The consequence of global stability is that exploitation will not irreversibly change the system. As long as the predator are not made extinct by excessive exploitation of their food supply, the system is able to recover. Once harvesting is stopped, the system will asymptotically approach its equilibrium level.
3 Stability analysis of system (1.4)
In this section, we give a qualitative analysis of system (1.4). Using the same approach of Theorem 2.1, the positivity and boundedness of the solution of system (1.4) are stated as follows. Positivity implies that the system persists, i.e., the populations survive. Boundedness may be interpreted as a natural restriction to growth as a consequence of limited resources.
Theorem 3.1
All solutions of system (1.4) with the positive initial conditions (x 0,y 0) are uniformly bounded in B 2, which is positively invariant, where
Next, we discuss steady states for (1.4). Since threshold T>0, the extinction equilibrium E 0(0,0) and prey-free equilibrium E 0(k,0) always exist for any parametric value. For the coexistence equilibrium E ∗(x ∗,y ∗), it follows that
From the first Eq. (3.2), the coexistence equilibrium E ∗ exists only when x ∗<k. In the absence of harvesting, H(y)=0, system (1.4) has only one coexistence equilibrium \(E^{*} =(\frac{d\alpha}{\beta_{1}-d},\,y^{\ast})\) where

Since the threshold harvesting policy starts only when the size of pest population is above the threshold T, we will consider two cases in the next sections: 0<T<T 0 and 0<T 0≤T.
Assume 0<T<T 0. Then there is no coexistence equilibrium E ∗ when y<T. Note that \(\frac{\beta_{1}x}{\alpha+x}\) is a increasing function with respect to x. When y>T, if the coexistence equilibrium E ∗=(x ∗, y ∗) exists then dα/(β 1−d)<x ∗<k and y ∗>T. Substituting \(y=\frac{r}{\beta}(1-\frac{x}{k})(\alpha +x)\) into the expression x, i.e., the second Eq. (3.2), then x ∗ is a root of the following quartic equation:

where (β 1−d)/(β 2 k 2)>0. By T<y ∗, it is easy to verify that
In such case, the equation p(x)=0 at most has three positive roots in interval (dα/(β 1−d),k). That is, system (1.4) have at most three coexistence equilibria. In fact, we show this case in Fig. 9(a). Denote the three coexistence equilibria by \(E_{1}^{*}(x_{1}^{*},\,y_{1}^{*})\), \(E_{2}^{*}(x_{2}^{*},\,y_{2}^{*})\) and \(E_{3}^{*}(x_{3}^{*},\,y_{3}^{*})\) and suppose that \(x_{1}^{*}>x_{2}^{*}>x_{3}^{*}\). Let the right side of system (1.4) be
By the formula for the implicit function derivation, the slopes of x-nullcline and y-nullcline are \(-\frac{{f_{1}}_{x}}{{f_{1}}_{y}}\) and \(-\frac{{f_{2}}_{x}}{{f_{2}}_{y}}\). Thus, if there is one E ∗ such that
then system (1.4) have two coexistence equilibria, \(E_{1}^{*}\) and \(E_{2}^{*}\) or \(E_{3}^{*}\) and \(E_{2}^{*}\). If there is one E ∗ such that
then system (1.4) have three equilibria, \(E_{1}^{*}\), \(E_{2}^{*}\), and \(E_{3}^{*}\). Under other conditions, there is only one equilibrium.

The nullclines and equilibria
Assume 0<T 0≤T, it follows from (3.4) that p(dα/(β 1−d))=−h(T 0−T)≥0. So, there is at most four coexistence equilibria. There is only one coexistence equilibrium E ∗(x ∗,y ∗) when y≤T, which is given by

For the case y>T, based on the above analysis, the coexistence equilibrium E ∗ will be unique, provided that (3.7) holds for every such E ∗, since if there are more than one E ∗, the equality (3.7) must not hold for alternate equilibria; system (1.4) have two coexistence equilibria \(E_{1}^{*}\) and \(E_{2}^{*}\), provided that there is only one equilibrium such that (3.8) holds and (3.7) does not hold; system (1.4) have three coexistence equilibria \(E_{1}^{*}\), \(E_{2}^{*}\), and \(E_{3}^{*}\), provided that there are one equilibrium such that (3.7) and (3.8) holds, respectively; system (1.4) have four coexistence equilibria \(E_{1}^{*}\), \(E_{2}^{*}\), \(E_{3}^{*}\), and \(E_{4}^{*}\), provided that there are two equilibria such that (3.7) holds; there is no equilibrium under other conditions, which is shown in Fig. 9(b).
3.1 The case 0<T<T 0
When 0<T<T 0, basic facts in this case are as follows.
1. The extinction equilibrium E 0(0, 0) and pest-free equilibrium E 0(k, 0) exist.
2. When y<T, there is no coexistence equilibrium; when y>T, there is at least one coexistence equilibrium \(E_{1}^{*}=(x^{*}_{1},\,y^{*}_{1})\) or is at most three coexistence equilibria \(E_{i}^{*}=(x^{*}_{i},\,y^{*}_{i}),\ i=1,2,3\) where \(\frac{d\alpha}{\beta_{1}-d}<x_{i}^{\ast}<k\).
3.1.1 Stability analysis
Let us now consider the stability of the system (1.4) governing the extinction equilibrium E 0(0,0), the pest-free equilibrium E 0(k,0) and the coexistence equilibrium \(E^{*} (x_{i}^{\ast},y_{i}^{\ast}),\ i=1,2,3\). The stability of the equilibrium E(x, y) is determined by the nature of eigenvalues of the Jacobian matrix

For the extinction equilibrium E 0, we can get the same results for system (1.3) as Theorem 2.2 and Remark 2.1.
Theorem 3.2
The extinction equilibrium E 0 is saddle with unstable manifold in x-direction and stable manifold in y-direction.
Remark 3.1
Threshold harvesting should ensure the sustainability of system.
For the pest-free equilibrium E 0, the eigenvalues are −r and \(\frac{\beta_{1}k}{\alpha+k}-d\). So, if \(k>\frac{d\alpha}{\beta_{1}-d}\), it is saddle with stable manifold in x-axis and unstable manifold direction in \(y=-\frac{\beta_{1} k+(r-d)(\alpha+k)}{\beta k}(x-k)\).
If \(k<\frac{d\alpha}{\beta_{1}-d}\), the pest-free equilibrium is stable. In fact, the pest-free equilibrium is G.A.S. In this case, the coexistence equilibrium E ∗ does not exist. It is impossible that system (1.4) exists a closed orbit in B 2. Again, if the coexistence equilibrium E ∗ exists, then the pest-free equilibrium E 0 is saddle.
For the coexistence equilibrium E ∗(x ∗,y ∗) where y ∗>T, the Jacobian matrix takes the form

Its determinant and trace are respectively

If D<0, one of the eigenvalues has positive real parts and the other one has negative real parts. Hence, E ∗ is an unstable saddle, and the existence of the limit cycle is guaranteed by Poincaré–Bendixson theorem and Theorem 3.1.
If D>0, Tr>0, then eigenvalues of J(E ∗) have positive real parts. Hence, E ∗ is an unstable node, and the existence of limit cycle is also guaranteed.
If D>0, Tr<0, then all eigenvalues of J(E ∗) have negative real parts, hence E ∗ is stable. Further, if \((\operatorname{Tr}J(E^{*}))^{2}-4\operatorname{Det}J(E^{*})\geq 0\) then E ∗ is stable node; if \((\operatorname{Tr}J(E^{*}))^{2}-4\operatorname{Det}J(E^{*})<0\) then E ∗ is stable focal point. Next, we can use the Lyapunov–LaSalle theorem to give a sufficient condition under which it is G.A.S. Consider

Its derivative along the solutions of (1.4) is

It follows for y≤T that

Clearly, if \(\frac{\beta y^{*}}{\alpha (\alpha +x^{*})}-\frac{r}{k}<0\) then \(\frac{dV}{dt}< 0\). Further, if y>T, then

So, if T<y ∗<T+c then \(\frac{dV}{dt}< 0\). In summary, \(\frac{dV}{dt}< 0\) always holds if
Hence, the Lyapunov–LaSalle theorem implies that all solutions ultimately approach the coexistence equilibrium E ∗ if (3.9) holds.
If \(k=\frac{d\alpha}{\beta_{1}-d}\), transcritical bifurcation appears. When \(k>\frac{d\alpha}{\beta_{1}-d}\), D>0 and Tr<0, the coexistence equilibrium E ∗ is stable. Besides, we had shown that E 0 is saddle when \(k>\frac{d\alpha}{\beta_{1}-d}\). In this case, as k decreases when it is below some level such that the inequality (3.9) holds, then E ∗ is G.A.S. When \(k=\frac{d\alpha}{\beta_{1}-d}\), then E ∗ collides E 0. Thus, E 0 is saddle-node, and transcritical bifurcation appears. Interestingly, all orbits will eventually tend to E 0, since \(\frac{dy}{dt}<0\) always holds in B 2 and E 0 is saddle with unstable manifold in x-direction and stable manifold in y-direction.
Summarizing the above discussion, we obtain the following theorem.
Theorem 3.3
If \(k>\frac{d\alpha}{\beta_{1}-d}\), the pest-free equilibrium E 0 is saddle with stable manifold in x-direction and unstable manifold in \(y=-\frac{\beta_{1} k+(r-d)(\alpha+k)}{\beta k}(x-k)\); if \(k<\frac{d\alpha}{\beta_{1}-d}\), E 0 is G.A.S; if \(k=\frac{d\alpha}{\beta_{1}-d}\), transcritical bifurcation appears and all orbits will eventually tend to E 0. Assume that D>0 and Tr<0, the coexistence equilibrium E ∗ is stable; furthermore, if the inequality (3.9) holds, E ∗ is G.A.S; if D<0, E ∗ is saddle; if D>0 and Tr>0, E ∗ is an unstable node, and there is at least one limit cycle near E ∗.
3.1.2 Bifurcations analysis
From above discussion, system (1.4) has a degenerate positive equilibrium when Tr=0 and D=0 hold. By a standard arguments of bifurcation theorem, we conclude that some bifurcation may occur for system (1.4). It is interesting that what kinds of bifurcation system (1.4) can undergo when the original parameters of system vary. Firstly, we will discuss the Hopf bifurcation.
Assume that D>0 and Tr<0, the coexistence equilibrium E ∗ is stable; if D>0 and Tr>0, E ∗ is an unstable node. The existence of Hopf bifurcation can be guaranteed. In other words, there are parameter values so that E ∗ satisfies \(\operatorname{Tr}J(E^{*})=0\). If Tr=0 and D>0, then the Jacobian matrix J(E ∗) has a pair of pure imaginary eigenvalues. In fact, the two pure imaginary eigenvalues are \(\lambda =\pm\sqrt{D}i\) and Re(λ)=Tr/2. Also, we have
Hence, the conditions for Hopf-bifurcations are satisfied. Thus, there exist small amplitude periodic solutions near E ∗.
Note that in parameter space (r,k,β,α,β 1,d,h,T,c), there is a surface
such that the coexistence equilibrium E ∗ is a center. Next, we discuss conditions under which E ∗ is center-type and the system undergo a Hopf bifurcation, and to determine the direction of such bifurcation we explicitly the corresponding Lyapunov number [32]. We first shift E ∗(x ∗, y ∗) to the origin using a change of coordinates given by u=x−x ∗ and v=y−y ∗, then we obtain the corresponding power series expansions, and noticing that some of the coefficients vanish, we get
Here, O i (|(u,v)|4), i=1,2 is the same order infinity, and
It is clear that a 10 b 01−a 01 b 10=D>0, a 10+b 01=0 because all parameters belongs to H. Since \(a_{01}=\frac{-\beta x^{*}}{\alpha +x^{*}}\), using the formula of the first Lyapunov number at the origin of (3.11) in [32], which is
where x ∗ satisfies Eq. (3.4) and

Therefore, the sign of σ is determined by R. If R≠0, then the origin of (3.11) is a weak focus of multiplicity one, it is stable if R<0 and unstable if R>0. Thus, system (1.4) experiments subcritical (σ>0) and supercritical (σ<0) Hopf bifurcations. With the aid of numerical calculation, we can find that the sign of σ is not determined. For instance, r=1,k=1,β=0.12,α=0.59,β 1=0.41,d=0.1,h=0.1,c=0.1,T=0.1, for which we have σ=−49.83<0. If r=1,k=1,β=0.47,α=1.56,β 1=0.78,d=0.2,h=0.15,c=0.15,T=0.1, it is easy to compute σ=51.676>0. Therefore, in the surface H there exists a curve
such that σ=0 where σ is a continuous function of all parameters. When all parameters is at the curve l, the origin of (3.11) is a weak focus of multiplicity at least two or a center. Hence, the surface of H is divided into two parts H sup and H sub by the curve l. That is,

The surface H sup is the supercritical Hopf-bifurcation surface and the surface H sub is the subcritical Hopf-bifurcation surface of system (1.4). Summarizing the above discussion, we have the following theorem.
Theorem 3.4
When \(\operatorname{Tr}J(E^{*})=0\), the system (1.4) exhibits subcritical if σ>0 and supercritical Hopf bifurcations if σ<0.
Notice that D=f 1 x f 2 y −f 1 y f 2 x at E ∗, system has only two coexistence equilibria when there are parameters so that E ∗ satisfies D=0; system has three coexistence equilibria when D>0; system has only one coexistence equilibrium under other conditions. Thus, if system has only two coexistence equilibria \(E_{i}^{*},\ i=1,2\) i.e., the case of D=0 at \(E_{2}^{*}\), then saddle-node bifurcation appears. Then there exists a surface in parameter space
such that for all parameters on the surface SN, system (1.4) has only two equilibria \(E_{1}^{*}\) and \(E_{2}^{*}\), \(E_{2}^{*}\) is a saddle-node. When the parameters pass from one side of the surface to the other side, the number of equilibria changes from two to three. This implies that system (1.4) undergoes a saddle-node bifurcation of codimension 1. The surface SN is called a saddle-node bifurcation surface. In this case, we consider h as bifurcation parameter, and D=0 when h=h ∗.
The Jacobian of (1.4) at \(E_{2}^{*}\) is given in Sect. 3.1.1. The eigenvalues are \(\lambda_{1,2}=\frac{\mathrm{Tr}\pm \sqrt{\mathrm{Tr}^{2}-4D}}{2}\). When the parameters belongs to SN, one can readily see that λ 1=0 and λ 2=Tr≠0. We shift the equilibrium \((h^{*}, x_{2}^{*}, y^{*}_{2})\) to the origin: ξ=h−h ∗, \(u=x-x_{2}^{*}\), \(v=y-y_{2}^{*}\), and the new system can be given
Then, we have

It is east to verify that \(\frac{\partial^{2} F_{2}}{\partial v^{2}}(0,0,0)\neq 0\). Then we can conclude that there exists [34] a saddle-node bifurcation when h=h ∗>0. In particular, this means that the system has one coexistence equilibria \(E_{1}^{*}\) for h>h ∗, there is exactly two equilibria \(E_{1}^{*}\) and \(E_{2}^{*}\) for h=h ∗, and there are three equilibria \(E_{1}^{*}\), \(E_{2}^{*}\) and \(E_{3}^{*}\) for h<h ∗.
Theorem 3.5
The system (1.4) exhibits saddle-node bifurcations if h=h ∗.
From Theorem 3.5, saddle-node bifurcation appears when D=0. That is, there are three coexistence equilibria \(E_{1}^{*}\), \(E_{2}^{*}\) and \(E_{3}^{*}\) when D>0. As the value of D decrease, the equilibrium \(E_{3}^{*}\) tends to the equilibrium \(E_{2}^{*}\), when it is equal to zero, i.e., D=0, \(E_{3}^{*}\) collides \(E_{2}^{*}\). Thus, the root \(x_{2}^{*}\) is a double root of equation (3.4) for D=0.
Next, we discuss the case of D=0 and Tr=0 at \(E_{2}^{*}\). In such case, both eigenvalues of the Jacobian matrix at \(E_{2}^{*}\) are zero. The Jacobian matrix is not zero matrix since \(x_{2}^{*}>0\). Thus, system (3.14) can be transformed to
where P 1(u,v) and Q 1(u,v) are smooth functions with at least the second order with respect to (u, v). Because \(x_{2}^{*}\) is a double root of Eq. (3.4), it follows that \(Q_{1}(u, 0)=k_{2}u^{2}+\overline{Q}(u)\), where k 2 is a nonzero constant depending on parameter (r,k,β,α,β 1,d,h,T,c) and \(\overline{Q}(u)\) is a smooth function with at least the third order with respect to u. By a series of nonsingular transformations in [33], system (3.15) becomes
where h(u), g(u) and Q 2(u,v) are smooth functions in all variables, h(0)=g(0)=Q 2(0,0)=0, k 2 and k 3 are constants depending on parameters (r, k,β,α,β 1,d,,h,T,c) and k 2≠0, m≥1 is an integer and. From ([33], Theorem 7.3, Chap. 2), the equilibrium (0,0) of system (3.16) is a cusp. This implies equilibrium \((x_{2}^{*}, y_{2}^{*})\) of system (1.4) is a cusp.
There are parameter values so that E ∗ satisfies D=0 and Tr=0, i.e., there exists a curve C={(r,k,β,α,β 1,d,h,T,c): D=0, Tr=0} such that there are only two equilibria \(E_{1}^{*}\) and \(E_{2}^{*}\), and \(E_{2}^{*}\) is a cusp for all parameters on the curve C.
Next we only transform system (1.4) to the canonical normal form of cusp of codimension two as in [34]. Let \(u=x-x_{2}^{*}\) and \(v=y-y_{2}^{*}\), and expand system (1.4) in a power series around the origin, then system (1.4) becomes
where O i (|(u, v)|3), i=1,2 is the same order infinity, a 10,a 01,a 20,a 11, b 10, b 01, b 20,b 11, b 02 see (3.12). Make the affine transformation u 1=u, v 1=a 10 u+a 01 v, then system (3.17) becomes
Here, O i (|(u 1,v 1)|3), i=3,4 is the same order infinity. Make further some the C ∞ changes of variables in a small neighborhood of (0,0)

then
where O 5(|(u 2,v 2)|3) is the same order infinity and

Since D=a 10 b 01−a 01 b 10=0 and Tr=a 10+b 01=0, this leads that the origin of (3.19) is a cusp of codimension 2 if η 1 η 2≠0. Hence, we present the \(E_{2}^{*}\) is a cusp of codimension two.
Theorem 3.6
If D=0, Tr=0 and η 1 η 2≠0, system (1.4) has two coexistence equilibria \(E_{i}^{*}(x_{i}^{*}, y_{i}^{*})\) i=1,2, and \(E_{2}^{*}\) is a cusp of codimension two.
In the following, we will find the versal unfolding of \(E_{2}^{*}(x_{2}^{*}, y_{2}^{*})\) depending on the original parameters of system (1.4), where d and h can be chosen as bifurcation parameters, Bogdanov–Takens bifurcation will occur.
Let d=d ∗−κ 1, h=h ∗−κ 2, where parameters d=d ∗ and h=h ∗ satisfies Tr=0 and D=0 respectively, and κ 1 and κ 2 are very small parameters. Consider the following system:
When κ 1=κ 2=0, system (3.20) has only two positive equilibrium \(E_{1}^{*}\), \(E_{2}^{*}\), and \(E_{2}^{*}\) is a cusp of codimension 2.
Next, we reduce system (3.20) to the normal form in successive steps. These steps are reminiscent of those performed in the proof of Theorem 3.6. For simplicity, we omit the laborious steps and write down the normal form directly:

where

In addition, G 1(u 3) is power series in u 3 of powers \(u_{3}^{i}\) satisfying i≥3, G 2(u 3) is power series in u 3 of powers \(u_{3}^{i}\) satisfying i≥2, G 3(u 3,v 3) is power series in (u 3,v 3) of powers \(u_{3}^{i}v_{3}^{j}\) satisfying i+j≥1, and G 1(u 3), G 2(u 3), G 3(u 3, v 3) are all functions depending on κ 1, κ 2. When D=0, Tr=0, 0<|κ 1|≪1, 0<|κ 2|≪1 and η 1 η 2≠0, it follows that η 3 η 4≠0. Applying the Malgrange preparation theorem, we have

where Q(u 3,κ 1,κ 2) is a power series in u 3 and Q(0,κ 1,κ 2)=η 3. For simplicity of computation, introducing the new time by

and further make the affine transformation

then system (3.21) becomes
where ξ i (0,κ 2)=0, i=1,2,3, \(\frac{\partial \xi_{j}(0, \kappa_{2})}{\partial\kappa_{1}}=0,\ j=1,3\) and R 2(z 3,z 4,0,0) is a power series in (z 3,z 4) of powers \(z_{3}^{i}z_{4}^{j}\) satisfying i+j≥3 and j≥2, and
Computing the Jacobian of (3.23) shows that the above parameter transformation from (κ 1,κ 2) to (μ 1,μ 2) is not singular in a small neighborhood of (κ 1,κ 2)=(0,0). Then system (3.22) can be rewritten as

By the theorem of Bogdanov and Takens in [34], we obtain the following theorem.
Theorem 3.7
When 0<|h−h ∗|≪1, 0<|d−d ∗|≪1 and η 1 η 2≠0, system (1.4) undergoes the cusp bifurcation of codimension 2 (i.e., the B–T bifurcation). Hence, there exists values of the parameters (r,k,β,α,β 1,d,h,T,c) such that system (1.4) has a unique stable limit cycle for some parameter values, and system (1.4) has a stable homoclinic loop for other parameter values.
3.2 The case 0<T 0≤T
When 0<T 0≤T, basic facts in this case are as follows.
1. The extinction equilibrium E 0(0, 0) and pest-free equilibrium E 0(k, 0) exist.
2. When y>T, there is at most four coexistence equilibria or there is no coexistence equilibrium; when y≤T, there is only one coexistence equilibrium \(E_{1}^{*}(x^{*}_{1}, y^{*}_{1})\) where \(x^{*}_{1}=\frac{d\alpha}{\beta_{1}-d}\) and \(y^{*}_{1}=T_{0}\).
Firstly, for the case y>T, the discussion is completely same as in above section, we omit the details.
When y<T, according to the theory of a classical predator–prey system
we obtain the following theorem.
Theorem 3.8
The extinction equilibrium E 0 is saddle. If \(k<\frac{d\alpha}{\beta_{1}-d}\), then the pest-free equilibrium E 0 is saddle; if \(k>\frac{d\alpha}{\beta_{1}-d}\), E 0 is G.A.S; if \(k=\frac{d\alpha}{\beta_{1}-d}\), transcritical bifurcation appears and all orbits will eventually tend to E 0. If k<α+2x ∗, the coexistence equilibrium E ∗ is stable; further, if k<α+x ∗, E ∗ is G.A.S; if k>α+2x ∗, E ∗ is unstable and system has at least one limit cycle; if k=α+2x ∗, Hopf bifurcation occurs.
Finally, we show the stability of the coexistence equilibrium E ∗ for the case \(x^{*}=\frac{d\alpha}{\beta_{1}-d}\) and T=y ∗=T 0. For convenience, we discuss the following two systems with harvesting and without harvesting, respectively,

When T=T 0, then the coexistence equilibrium \(E^{*}_{a}\) of (3.25(a)) is equal to the coexistence equilibrium \(E^{*}_{b}\) of (3.25(b)), \(E^{*}_{a}=E^{*}_{b}\). Accordingly, the determinant and trace of the Jacobian matrix \(J(E^{*}_{i})\) are Tr i and Det i (i=a,b), respectively. Based on the above analysis, we can obtain

We summarize the dynamics of system (1.4) at the coexistence equilibrium E ∗ when T=y ∗ as follows, which is shown in Fig. 10.

The dynamics of system (1.4) at the coexistence equilibrium E ∗ when T=y ∗
Assume Det b =f 1x f 2y −f 1y f 2x >0.
(1) If Tr b >0, then Tr a >0, \(E^{*}_{a}\) and \(E^{*}_{b}\) are unstable. Furthermore,
(i) if \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}\geq0\), both \(E^{*}_{a}\) and \(E^{*}_{b}\) are unstable node; hence, E ∗ is unstable node and the existence of limit cycle is guaranteed by the Poincaré–Bendixson theorem (see Fig. 10(a)).
(ii) if \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}<0\) and \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}\geq 0\), \(E^{*}_{a}\) is unstable focus and \(E^{*}_{b}\) is an unstable node; hence, E ∗ is unstable and the existence of the limit cycle is also guaranteed (see Fig. 10(b)).
(iii) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}<0\), both \(E^{*}_{a}\) and \(E^{*}_{b}\) are unstable focus; in this case, it is intricate, the stability of E ∗ may be stable or unstable or system (1.4) has periodic solutions (see Fig. 10(c)).
(2) If Tr b <0 and Tr a >0, then \(E^{*}_{a}\) is unstable and \(E^{*}_{b}\) is stable. Furthermore,
(i) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}\geq0\), then \(E^{*}_{a}\) is unstable node and \(E^{*}_{b}\) is stable node; in this case, system (1.4) has homoclinic loops (see Fig. 10(d)).
(ii) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}<0\) and \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}\geq 0\), then \(E^{*}_{a}\) is an unstable node and \(E^{*}_{b}\) is stable focus; hence, E ∗ is unstable.
(iii) if \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}<0\) and \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}<0\), then \(E^{*}_{a}\) is unstable focus and \(E^{*}_{b}\) is stable focus. In this case, it is intricate, the stability of E ∗ may be stable or unstable or system (1.4) has periodic solutions.
(3) If Tr a <0, then \(E^{*}_{a}\) and \(E^{*}_{b}\) are stable equilibria. Furthermore,
(i) if \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}\geq 0\) and \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}\geq 0\), then both \(E^{*}_{a}\) and \(E^{*}_{b}\) are stable node; hence, E ∗ is a stable node (see Fig. 10(e)).
(ii) if \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}\geq 0\) and \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}<0\), then \(E^{*}_{a}\) is a stable node and \(E^{*}_{b}\) is stable focus, hence, E ∗ is stable (see Fig. 10(f)).
(iii) if \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}<0\) and \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}\geq0\), then \(E^{*}_{a}\) is stable focus and \(E^{*}_{b}\) is stable node, hence, E ∗ is stable.
(iv) if \(\mathrm{Tr}_{a}^{2}-4\mathrm{Det}_{a}<0\) and \(\mathrm{Tr}_{b}^{2}-4\mathrm{Det}_{b}<0\), then both \(E^{*}_{a}\) and \(E^{*}_{b}\) are stable focus; in this case, it is intricate, the stability of E ∗ may be stable or unstable or system (1.4) has periodic solutions.
Assume Det b =f 1x f 2y −f 1y f 2x <0, then Tr a >0, hence \(E^{*}_{b}\) is a saddle and \(E^{*}_{a}\) is an unstable focus (see Fig. 10(g)) or node (see Fig. 10(h)).
Furthermore, for the case of (3(iii)), the discontinuous Hopf bifurcation of (1.4) at T=y ∗ occurs, this implies there exists an asymptotically stable limit cycle as required (using the same approach as system (1.3) at T=x ∗ in Sect. 2.2.1).
To facilitate the interpretation of our mathematical results, we summarize the dynamics of system (1.4) at the coexistence equilibrium E ∗ in Table 1 (without loss of generality, we only give the results of the case 0<T<T 0).
3.3 Numerical results
Our focus so far has been on the dynamics of system (1.4). To facilitate the interpretation of our mathematical results in model (1.4), we proceed to investigate it by numerical simulations. Since system (1.4) cannot be solved explicitly, it is difficult to study them analytically.
Let r=2.7,k=8.5,β=1.2,α=1,β 1=1,d=0.64,c=1,T=1. In this case, we consider h as bifurcation parameter. It easy to compute that T 0=4.943. Therefore, there is at most three coexistence equilibria caused by continuous threshold harvesting when T<T 0. The two boundary equilibria E 0, E 0 are unstable saddles.
If h=0.97, system (1.4) has only one coexistence equilibrium \(E_{3}^{*}\), which is unstable, hence there exists a unique G.A.S. cycle (see Fig. 11(a)). As soon as h=0.9755, there is two coexistence equilibria \(E_{i}^{*},\ i=2,3\): \(E_{3}^{*}\) is unstable, \(E_{2}^{*}\) is saddle-node and there exists homoclinic orbit which lies in the intersection of the stable manifold and the unstable manifold of the equilibrium \(E_{2}^{*}\) (see Fig. 11(b)). As the harvesting rate increases when h=1.005, there is three coexistence equilibria \(E_{i}^{*}\) (i=1,2,3): \(E_{3}^{*}\) is unstable, \(E_{2}^{*}\) is saddle and \(E_{1}^{*}\) is stable. System exists homoclinic orbit which joins the saddle to itself (see Fig. 12(a)). In this case, Figs. 3(b) and 4(a) show the system (1.4) exhibits saddle-node bifurcations. When the harvesting rate becomes slightly larger, i.e., h=1.1, the homoclinic orbit is broken and there is a stable limit cycle near equilibrium \(E_{3}^{*}\). Besides, the equilibrium \(E_{2}^{*}\) is saddle and \(E_{1}^{*}\) is stable (see Fig. 12(b)). If the harvesting rate is equal to the level, h=1.26, the limit cycle is broken and reach the stable equilibrium \(E_{3}^{*}\). The equilibrium \(E_{2}^{*}\) is saddle and \(E_{1}^{*}\) is stable (see Fig. 13(a)). If h=1.32815, the equilibrium \(E_{3}^{*}\) collides \(E_{2}^{*}\), and \(E_{2}^{*}\) is saddle-node (see Fig. 13(b)). From Figs. 12(a)–13(b), the stable equilibrium \(E_{3}^{*}\) becomes unstable, which implies that Hopf bifurcation occurs. Further increasing the harvesting rate h=1.4 will lead to the saddle-node bifurcations. Besides, the equilibrium \(E_{1}^{*}\) is G.A.S (see Fig. 14). Thus, our numerical simulations shows that system (1.4) undergoes the B–T bifurcation.

(a) The unique coexistence equilibrium \(E_{3}^{*}\) is unstable and the stable limit cycle occur. (b) There are two coexistence equilibria: one is an unstable focus and the other one is saddle-node. Hence, there exists a homoclinic orbit, which lies in the intersection of the stable manifold and unstable manifold of the equilibrium \(E_{2}^{*}\)

(a) There are three coexistence equilibria: \(E_{3}^{*}\) is unstable, \(E_{2}^{*}\) is saddle, and \(E_{1}^{*}\) is stable. The homoclinic orbit which joins the saddle to itself occur. (b) The homoclinic orbit is broken and system has stable limit cycle near \(E_{3}^{*}\). In addition, the equilibrium \(E_{2}^{*}\) is saddle and \(E_{1}^{*}\) is stable

(a) The limit cycle is broken and reach stable equilibrium \(E_{3}^{*}\), the equilibrium \(E_{2}^{*}\) is saddle, and \(E_{1}^{*}\) is stable. (b) The equilibrium \(E_{2}^{*}\) is a cusp of codimension 2, and \(E_{1}^{*}\) is G.A.S

When h=1.4, the unique coexistence equilibrium is G.A.S
As we know, if the coexistence equilibrium \(E_{2}^{*}\) is saddle, then the first quadrant \(R_{+}^{2}\) is divided into two regions by the stable manifold of \(E_{2}^{*}\). The results can be applied in the context of biological pest control. In the example of Figs. 12(b) and 13(a), if the aim is to lower the size of pest population in a finite time then we require the state of the ecosystem in the second region by harvesting strategy. If the objective is to maintain the state at this point (biological conservation), then we require the state of the ecosystem in the first region by harvesting strategy.
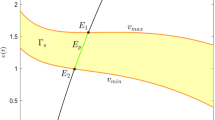
From above discussions, there is an important threshold X 0:=dα/(β 1−d) which is the equilibrium level of the components x in the absence harvesting. We now give some biological explanations of our threshold harvesting policy using the threshold X 0. When the threshold is above the environmental carrying capacity of the prey, X 0≥k, the system (1.4) has no coexistence equilibrium and the pest-free equilibrium E 0(k, 0) is G.A.S; this implies the pest species will eventually extinct. However, complete eradication of the pest species is generally not possible, nor is it biologically or economically desirable. Therefore, it is necessary to consider the coexistence case, which is in line with reality. When the threshold X 0 is in the interval ((k−α)/2, k), the unique coexistence equilibrium is G.A.S. whether there is harvesting or not; this implies the harvesting policy does not affect the ecosystem. From the biological view point, there is enough food for predation of the pest. As long as the pest are not made extinct by harvesting policy, the system is able to recover. Once harvesting is stopped, the system will asymptotically approach its natural equilibrium. Here, our control policy for the pest stop only when the population is above the threshold T. Consequently, the project of control pest cannot be reached. In this case, we need to take into account others integrated control strategies including available host resistance, chemical, and biological control measures such as pesticides or introducing natural enemy (this can be further discussed by state-dependent impulsive differential equation). When the threshold X 0 is below the level (k−α)/2, the presence of continuous threshold harvesting makes the dynamic behavior more complex with multiple coexistence equilibria (see Fig. 9), limit cycle, homoclinic orbit, saddle-node bifurcation, transcritical bifurcation, subcritical and supercritical Hopf bifurcation, and Bogdanov–Takens bifurcation, which is shown in Figs. 11, 12, 13, 14, and discontinuous Hopf bifurcation (see Fig. 10). In such cases, the equilibrium x ∗ is always above the threshold X 0 and the equilibrium y ∗ is always above the threshold T 0; the results imply harvesting for a pest raises the level of both species. This happens due to the harvesting of the pest lowers the level of the pest raises the level of prey, in turn, the increase of the prey make the pests have enough food for predation and raise the level of the pests. Thus, harvesting has impact on the total populations. In conclusion, in comparison with the predator–prey model without harvesting, the threshold harvesting makes the dynamics more complex and raises the equilibrium level of both species.
4 Discussion
In this paper, we have studied two Holling type II predator–prey models with continuous threshold harvesting, which represents situations when the harvesting policy needs to be applied only when the harvest population is above the threshold T. The two models are nonsmooth and the aim of this paper is to provide how the harvesting threshold affects the dynamics of both models. When the harvesting threshold is larger than some positive level, the harvesting policy does not affect the ecosystem; when the harvesting threshold is less than the level, the model has complex dynamics with multiple coexistence equilibria, limit cycle, bistability, homoclinic orbit, saddle-node bifurcation, transcritical bifurcation, subcritical and supercritical Hopf bifurcation, Bogdanov–Takens bifurcation, and discontinuous Hopf bifurcation. We provide the complete stability analysis for both models and carry out bifurcation analysis by choosing the death rate and the harvesting rate of the predator as the bifurcation parameters. Also, the figures of all degenerate structures are given.
The objective of threshold harvesting policy is to achieve a level of pest control that is acceptable in economic terms to farmers while causing minimal disturbance to the environments of nontarget individuals. Note that complete eradication of the pest species is generally not possible, nor is it biologically or economically desirable. Therefore, a good pest control program should reduce the pest to levels acceptable to the public. This implies that there is an economic harvesting threshold T above which the financial damage is sufficient to justify using such harvesting. Our results shows that harvesting firstly decrease the density of the pest species, and further the loss of food lowers the density of the natural enemy for natural enemy-pest system (1.3). While for the crop-pest system (1.4), the harvesting lowers the level of pests and raises the level of prey, in turn, the increase of the prey make the pests have enough food for predation and raise the level of the pests. It is seen that the threshold harvesting policy of system (1.3) is more effective than the system (1.4).
References
Hall, D., Norgaard, R.: On the time and application of pesticides. Am. J. Agric. Econ. 55, 198–201 (1973)
Sunding, D., Zivin, J.: Insect population dynamics, pesticide use and farmworker health. Am. J. Agric. Econ. 82, 527–540 (2000)
Liang, J., Tang, S.: Optimal dosage and economic threshold of multiple pesticide applications for pest control. Math. Comput. Model. 51, 487–503 (2010)
Shoemaker, C.: Optimal timing of multiple application of pesticides with residual toxicity. Biometrics 36, 803–812 (1979)
Talpaz, H., Curry, G., Sharpe, P., DeMichele, D., Frisbie, R.: Optimal pesticide application for controlling the boll weevil in cotton. Am. J. Agric. Econ. 60, 469–475 (1978)
Headley, J.: Defining the economic threshold, presented at the National Academy of Sciences. In: Symposium on Pest Control Strategies for the Future, Washington, DC, 15 April 1971, pp. 100–108 (1972)
Tang, S., Chen, L.: Modelling and analysis of integrated pest management strategy. Discrete Contin. Dyn. Syst. 4, 759–768 (2004)
Tang, S., Cheke, R.: State-dependent impulsive models of integrated pest management (IPM) strategies and their dynamic consequences. J. Math. Biol. 50, 257–292 (2005)
Tang, S., Xiao, Y., Chen, L., Cheke, R.: Integrated pest management models and their dynamical behavior. Bull. Math. Biol. 67, 115–135 (2005)
Pei, Y., Chen, L., Zhang, Q., Li, C.: Extinction and permanence of one-prey multi-predators of Holling type II function response system with impulsive biological control. J. Theor. Biol. 235, 495–503 (2005)
Pei, Y., Zeng, G., Chen, L.: Species extinction and permanence in a prey-predator model with two-type functional responses and impulsive biological control. Nonlinear Dyn. 52, 71–81 (2008)
Ji, L., Wu, C.: Qualitative analysis of a predator–prey model with constant-rate prey harvesting incorporating a constant prey refuge. Nonlinear Anal., Real World Appl. 11, 2285–2295 (2010)
Xiao, D., Ruan, S.: Bogdanov–Takens bifurcations in predator–prey systems with constant rate harvesting. Fields Inst. Commun. 21, 493–506 (1999)
Xiao, D., Jennings, L.: Bifurcations of a ratio-dependent predator–prey system with constant rate harvesting. SIAM J. Appl. Math. 65, 737–753 (2005)
Huang, Y., Chen, F., Li, Z.: Stability analysis of a prey–predator model with Holling type III response function incorporating a prey refuge. Appl. Math. Comput. 182, 672–683 (2006)
Pei, Y., Lv, Y., Li, C.: Evolutionary consequences of harvesting for a two-zooplankton one-phytoplankton system. Appl. Math. Model. 36, 1752–1765 (2012)
Tao, Y., Wang, X., Song, X.: Effect of prey refuge on a harvested predator–prey model with generalized functional response. Commun. Nonlinear Sci. Numer. Simul. 16, 1052–1059 (2011)
Tang, S., Xiao, Y., Cheke, R.: Multiple attractors of host–parasitoid models with integrated pest management strategies: eradication, persistence and outbreak. Theor. Popul. Biol. 73, 181–197 (2008)
Tang, S., Cheke, R.: Models for integrated pest control and their biological implications. Math. Biosci. 215, 115–125 (2008)
Collie, J., Spencer, P.: Management Strategies for fish populations subject to long term environmental variability and depensatory predation, Technical report 93-02, Alaska Sea Grant College, 629–650 (1993)
Aanes, S., Engen, S., Saether, B., Willebrand, T., Marcstrom, V.: Sustainable harvesting strategies of willow ptarmigan in a fluctuating environment. Ecol. Appl. 12, 281–290 (2002)
Leard, B., Rebaza, J., Saether, B.: Analysis of predator–prey models with continuous threshold harvesting. Appl. Math. Comput. 217, 5265–5278 (2011)
Lande, R., Saether, B., Engen, S.: Threshold harvesting for sustainability of fluctuating resources. Ecology 78, 1341–1350 (1997)
Rebaza, J.: Dynamics of prey threshold harvesting and refuge. J. Comput. Appl. Math. 236, 1743–1752 (2012)
Liu, X., Liu, S.: Codimension-two bifurcation analysis in two-dimensional Hindmarsh–Rose model. Nonlineat Dyn. 67, 847–857
Tian, R., Cao, Q., Yang, S.: The codimension-two bifurcation for the recent proposed SD oscillator. Nonlinear Dyn. 59, 19–27 (2010)
Clarke, F., Ledyaev, Y., Stern, R., Wolenski, P.: Nonsmooth Analysis and Control Theory. Springer, New York (1998)
Leine, R.: Bifurcations of equilibria in non-smooth continuous systems. Physica D 223, 121–137 (2006)
Leine, R., van Campen, D.: Bifurcation phenomena in non-smooth dynamical systems. Eur. J. Mech. A, Solids 25, 595–616 (2006)
Chattopadhyay, J., Sarkar, R., Mandal, S.: Toxin producing plankton may act as a biological control for planktonic blooms: a field study and mathematical modelling. J. Theor. Biol. 215, 333–344 (2002)
Kuang, N., Freedman, H.: Uniqueness of limit cycles in Gause-type models of predator–prey systems. Math. Biosci. 88, 76–84 (1988)
Perko, L.: Differential Equations and Dynamical Systems. Texts in Applied Mathematics, vol. 7. Springer, Berlin (2006)
Zhang, Z., Ding, T., Huang, W., Dong, Z.: Qualitative Theory of Differential Equations. Science Press, Beijing (1992) (in Chinese). English edition. Transl. Math. Monogr., vol. 101. Am. Math. Soc., Providence (1992)
Hale, J.: Dynamics and Bifurcations. Springer, Berlin (1991)
Acknowledgements
We are extremely grateful to the critical comments and invaluable suggestions made by anonymous honorable reviewers. This work is supported by the Natural Science Foundation of China, the Doctoral Program of Higher Education of China and National Natural Science Foundation of China (11101305).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lv, Y., Yuan, R. & Pei, Y. Dynamics in two nonsmooth predator–prey models with threshold harvesting. Nonlinear Dyn 74, 107–132 (2013). https://doi.org/10.1007/s11071-013-0952-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11071-013-0952-2