
Abstract
Radiation therapy is widely used in the treatment of primary malignant brain tumors and metastatic tumors of the brain with either curative or palliative intent. The limitation of cancer radiation therapy does not derive from the inability to ablate tumor, but rather to do so without excessively damaging the patient. Among the varieties of radiation-induced brain toxicities, it is the late delayed effects that lead to severe and irreversible neurological consequences. Following radiation exposure, late delayed effects within the CNS have been attributable to both parenchymal and vascular damage involving oligodendrocytes, neural progenitors, and endothelial cells. These reflect a dynamic process involving radiation-induced death of target cells and subsequent secondary reactive neuroinflammatory processes that are believed to lead to selective cell loss, tissue damage, and functional deficits. The progressive, late delayed damage to the brain after high-dose radiation is thought to be caused by radiation-induced long-lived free radicals, reactive oxygen species, and pro-inflammatory cytokines. Experimental studies suggest that radiation-induced brain injury can be successfully mitigated and treated with several well established drugs in wide clinical use which exert their effects by blocking pro-inflammatory cytokines and reactive oxygen species. This review highlights preclinical and early clinical data that are translatable for future clinical trials.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Radiation therapy is one of the most effective adjuvant treatments to surgery for the treatment of malignant central nervous system (CNS) tumors. Despite the use of sophisticated radiation delivery methods, the risk of inadvertent injury to normal, non-targeted critical brain structures limits the maximum dose used in radiation therapy. Consequently, three distinct strategies have been proposed to improve its utility. One strategy exploits recent advances in the delivery of radiation using image guided precision localization to permit delivery of higher doses of radiation to well-defined intracranial targets. Another strategy exploits advances in targeting radiation using radiosurgery and/or intensity modulated radiotherapy to minimize exposure and toxicity in the surrounding normal brain tissues. Other strategies employ novel neurobiological findings to prevent/decrease normal tissue toxicity or enhance the intrinsic radiosensitivity of tumor tissue to increase the therapeutic efficiency of radiation. Pharmacological strategies designed to reduce normal tissue damage would complement focused high-dose precision radiotherapy if the drug therapies do not inadvertently protect tumor from radiation therapy. Finding ways to prevent both acute and late delayed toxicity in normal brain tissue associated with brain tumor irradiation is of paramount importance, since the devastation resulting from this toxicity can sometimes be far worse than the initial lesion that was treated.
Clinical manifestation of radiation brain toxicity
Clinical responses of the brain to whole brain radiation may be grouped as acute, early delayed, and late delayed effects. Acute effects occur during and/or shortly after the radiation exposure and are characterized by symptoms of fatigue, dizziness, and signs of increased intracranial pressure. The acute effects are considered to be secondary to edema and disruption of the blood–brain barrier (BBB). Early delayed effects occur 6–12 weeks post-irradiation and usually show reversible symptoms generalized weakness and somnolence, in part resulting from a transient demyelination. It is, however, the late delayed effects that may lead to severe irreversible neurological consequences. Clinical effects of the late damage to the brain range from minor-to-severe cognitive deficits to focal or diffuse necrosis of the brain parenchyma. Late radiation necrosis may be associated with focal neurological signs such as seizures, dysfunction of the cranial nerves, and increased intracranial pressure caused by the persistent vasogenic edema resulting from BBB damage. Late radiation necrosis can be detected using magnetic resonance imaging where it manifests as diffuse non-specific changes in white matter.
Since the normal brain tissue tolerance is directly related to the volume and dose of tissues being irradiated, the smaller incremental exposure of the normal tissue would be routinely used in the clinical radiotherapy. The dose volume histogram would be a useful guide in estimating the volume of normal tissues being exposed. Nevertheless, long-term functional changes following whole brain irradiation can be more pronounced in young children, particularly when radiation is used during the developmental stages of neural tissues. Treatment-related late effects with cognitive development and learning capability as well as neuroendocrine functions have been shown among childhood cancer survivors [1]. Although the dose-response relationship is not well established in humans, higher radiation doses may be associated with increased incidence of long-term complications. Kim and Brown addressed the issue of response of tumor and late-responding normal tissue of critical intracranial structures to both single dose and fractionated radiosurgery [2]. As expected, single dose radiosurgery was found to be more effective in curing small intracranial tumors than large tumors. They further tested the validity of linear quadratic iso-effect formula for the normal tissue injury using the optic neuropathy as an end-point. The sparing effect of fractionated radiosurgery is greater for late-responding tissues, relative to the rapidly proliferating tumor tissues.
Pathogenesis of white matter necrosis
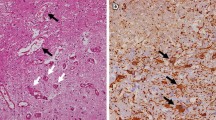
Histological features of the brain injury are diverse and not specific to radiation. Classically, however, the most prominent histopathological changes are (1) parenchymal cell loss that involves demyelination of white matter, encephalomalacea, gliosis, and neural cell loss, and (2) vascular endothelial damages that may cause altered permeability and BBB breakdown in the acute and subacute phases, and teleangiectasia, hyalinosis and fibrinoid deposits in the vessel walls in the late phase [3–5]. Since white matter necrosis is the dominant histopathological presentation following high dose radiation exposure, numerous radiobiological studies have been carried out to identify the primary target cell populations involved, which are postulated to be either endothelial or glial progenitors and/or stem cells [6, 7]. Since the white matter necrosis is consistently associated with demyelination and oligodendrocytes produce myelin for myelin sheaths, earlier radiobiological studies focused on determining the in vitro/in vivo radiosensitivity of glial cells including the progenitor cells of oligodendrocytes, known as O-2A cells [8]. Results clearly showed that oligodendrocytes are the most radiosensitive type of glial cell, with cell death occurring rather early after relatively low doses of irradiation [9]. Thus, the data seemed to be consistent with the white matter selectivity of radiation-induced brain injury. However, the time course of oligodendrocyte depletion after irradiation was not consistent with that of white matter necrosis during the development of late delayed effects [10].
In an attempt to determine whether it is the endothelial cell or the glial progenitor cell that is the primary target responsible for the white matter necrosis, a novel radiobiological study has been carried out using boron neutron capture therapy. The method is based on the short-ranged alpha particles released from the neutron capture reaction in the stable boron isotope. Selective irradiation of the microvasculature was achieved by intraperitoneal administration of a boron compound which remains within the endothelium, since BBB excludes the boronated compound from the brain parenchyma. The selective vascular damage from the boron neutron capture irradiation, with only a minimal dose to glial cells, resulted in significant demyelination and white matter necrosis indicating that endothelial cell loss plays a significant role in this pathogenesis [11]. It is, however, interesting to note that selective irradiation of the vascular endothelium has no effect on the survival of murine intestinal crypt stem cells and endothelial cell damage is not causative in the eventual development of the gastrointestinal syndrome [12].
In addition to the changes in tissue architecture, there are also changes in cellular composition such as increased numbers of reactive astrocytes (gliosis) and microglia at the zone of radionecrosis [13, 14]. These reactive cells have been shown to produce reactive oxygen species (ROS), cytokines and growth factors that may cause cell-to-cell interaction and progressive inflammatory radiation injury [15–17]. In particular, excessive generation of ROS from the injured and/or pro-inflammatory cells has been implicated in the development of late delayed effects. Hence, the damage response is now viewed as a continuous, dynamic and interactive process. The associated pathologies are thought to be a consequence of complex interplay among many different processes including vascular endothelial cell loss, excessive generation of ROS and inflammatory mediators [5, 14].
Excessive generation of ROS following radiation
Reactive oxygen species include oxygen ions, free radicals, and both inorganic and organic peroxides. They are highly reactive due to the presence of unpaired electrons in their shells. ROS are produced continually in most tissues and are part of normal cell signaling functions. Under normal conditions, numerous antioxidant systems such as superoxide dismutase (SOD), catalase, and glutathione peroxidase defend cells against oxidative damage. However, during times of environmental stress, ROS levels can increase dramatically, overwhelming antioxidant systems and resulting in significant damage to cell structures [18–21]. This scenario is referred to as oxidative stress. Damaging ROS might arise from several sources including infiltrating activated leukocytes, macrophages, and neurons (as a direct consequence of excitotoxic stimulation). Tissue hypoxia resulting from vascular damage is another continual source of ROS generation. Further, pro-inflammatory cytokines and growth factors greatly increase intracellular ROS generation [22, 23].
The brain is highly susceptible to oxidative stress. High rates of aerobic glycolysis generate a continual supply of ROS within mitochondria. Relative to other tissues, glial cells and neurons contain relatively low levels of antioxidant enzymes such as SOD, catalase, glutathione peroxidase, etc. [24]. Moreover, myelin membranes contain high levels of peroxidizable fatty acids, making them exceptionally vulnerable to ROS [25].
Given the fact that persistent oxidative stress and ROS derived from pro-inflammatory cytokines and growth factors are involved in the pathogenesis of central nervous system toxicity, strategies to block effector molecules or to reduce oxidative stress are attractive approaches for mitigating radiation-induced toxicity. Dietary supplements with selective antioxidants have been shown to prevent the reduction in the plasma level of total antioxidant status in rodents exposed to low and high linear energy transfer radiation [26]. Another approach to intercept reactive oxidants is to test drugs that induce systemic antioxidant biochemical properties.
VEGF, hypoxia and blood–brain barrier
Vascular endothelial growth factor (VEGF) is among the first growth factors upregulated during the pathogenesis associated with late delayed effects and is intimately involved in the development of vascular pathologies and white matter necrosis [27, 28]. The upregulation of VEGF evolves gradually following brain radiation and typically occurs several weeks prior to the development of overt tissue pathology [12]. The cascade of events leading to white matter necrosis is initiated by a gradual depletion of vascular endothelial cells forming the BBB. The reduction in endothelial cell number is not accompanied by a substantial decrease in vascular density, rather the endothelial cell density in the existing vasculature is reduced which gradually diminishes the integrity of the BBB [27]. This gives rise to vasogenic edema, inflammation, and tissue hypoxia which eventually result in the nearly simultaneous induction of HIF-1α and VEGF [15, 27, 29]. The potent effect of VEGF on vascular permeability exacerbates disruption of BBB, worsening vasogenic edema, inflammation, and tissue hypoxia and resulting in further increases in VEGF concentration. VEGF concentrations eventually reach levels sufficient to induce endothelial proliferation (i.e., angiogenesis) resulting in a dramatic increase in endothelial cells referred to as “conditional renewal” [28]. This angiogenic response persists for approximately 20 weeks; however, it ultimately fails to restore BBB integrity, resulting in a precipitous decline in the number of endothelial cells that eventually leads to the development of white mater necrosis.
Recent reports suggesting that anti-VEGF therapy can normalize BBB function in microvessels damaged during radiosurgery in the human brain suggest that manipulating the VEGF signaling cascade may be a useful strategy for mitigating late delayed injuries resulting from brain radiation as well [30, 31]. Applied at the appropriate times, anti-VEGF therapies may act to normalize the perivascular microenvironment, thereby preventing the exacerbation of vasogenic edema and/or facilitating a more successful restoration of BBB integrity via angiogenesis.
Neurogenesis and cognitive dysfunction
Whereas high doses of radiation produce overt histopathological changes such as demyelination and vasculopathies within the brain parenchyma, lower dose exposures produce cognitive dysfunction without inducing obvious morphological changes. Although the pathogenesis of radiation-induced cognitive dysfunction is unknown, recent studies suggest that it may involve impaired neurogenesis within the subgranular zone (SGZ) of the dentate gyrus [32–34]. Whole brain radiation doses as low as 2 Gy are sufficient to reduce the rate of proliferation among neuronal progenitors within the SGZ [35]. At higher radiation doses proliferation among these progenitors is effectively blocked, accompanied by an impairment of contextual fear conditioning and synaptic plasticity within the dentate gyrus [36]. It is presently unknown whether impaired neurogenesis within the SGZ contributes directly to the varied forms of radiation-induced cognitive impairment. Although the neuronal progenitors within the SGZ are highly radiosensitive, direct effects of radiation on synaptic plasticity in the hippocampus or in other limbic or paralimbic structures may also be involved. It is, therefore, important to consider the functional status of synaptic plasticity, as measured by long term potentiation (LTP), along with the status of N-methyl-D-aspartate (NMDA) receptors in multiple brain structures in this context [37–39]. The volume of irradiated tissue may also be relevant in this regard. Though this has yet to be systematically investigated in this context, several clinical observations suggest that the limbic system, and in particular the hippocampus, functions as a parallel circuit between hemispheres, requiring bilateral irradiation of these limbic structures to produce cognitive impairment [40].
Recent studies by Monje et al. have shown that radiation-induced impairment of neurogenesis within the rat dentate gyrus results from a disruption of a microvascular “niche” within the SGZ [41]. They postulated that neurogenesis is coordinated within this microenvironment by signals derived both from within the brain parenchyma and from circulating blood–plasma. Subsequent studies by Monje and her colleagues showed that neurogenesis within the SGZ is inversely correlated with the activated microglial load after radiation, and can be partially restored by administering the anti-inflammatory drug, indomethacin [42]. These data suggest that reducing the inflammatory status associated with the dysfunction of the vascular endothelium may represent a successful strategy for preserving neurogenesis and perhaps mitigating cognitive impairment following brain irradiation.
Therapeutic strategies
Based on the foregoing discussion, progressive damage to the CNS after irradiation is increasingly thought to be caused by radiation-induced and long-lived free radicals, ROS, and pro-inflammatory cytokines, resulting in a deterioration of neurological function. Strategies aimed at blocking effector molecules or otherwise reducing oxidative stress are attractive for preventing or mitigating radiation-induced CNS toxicity. One approach to impeding the actions of reactive oxidants is to administer drugs that induce endogenous antioxidant biochemical processes. Several drugs which block pro-inflammatory cytokines and ROS are currently available and in wide clinical use.
Angiotensin-converting enzyme (ACE) inhibitors
Angiotensin converting enzyme inhibitors (ACEi) are widely used as antihypertensive agents. In cardiac patients, ACEi improve cardiac function and reduce the risk of re-infarction, morbidity, and mortality [43, 44]. ACE converts angiotensin I to angiotensin II. Angiotensin II is a potent vasopressor which acts by binding to G protein-coupled receptors, angiotensin II type 1 and type 2 (AT1R and AT2R, respectively). Binding to AT1R produces vasoconstrictive effects, whereas binding to AT2R produces vasodialating effects [45]. These receptors are widely distributed in various tissues including brain, where they are found on both neurons and glial cells. After tissue injury, AT2R are commonly upregulated resulting in greater production of angiotensin II [46]. In this context, angiotensin II is a pro-inflammatory and pro-fibrogenic mediator which, in combination with other cytokines and growth factors, may participate in the development of long-term tissue and organ injury [46].
In addition to the role of angiotensin II as an inflammatory effector molecule, there is an evolving body of evidence supporting the idea that angiotensin II-induced oxidative stress plays an important role in angiotensin II-mediated cellular responses. Griendling et al. first documented that angiotensin II increases NADPH oxidase activity in macrophages, an abundant source of ROS [47]. Tojo et al. have shown that ACEi significantly reduces NADPH oxidation products and lipid peroxidation products in rats with heart failure [48]. This is further supported by the data that PPAR-γ ligands, which are potent anti-oxidants, may attenuate angiotensin-induced ROS generation and monocyte adhesion to endothelial cells [49]. Taken together, these results suggest that angiotensin II-induced oxidative stress may be an important mechanism by which this neurohormone induces many of its tissue pathologic effects.
The role of ACE and its metabolites in the pathogenesis of radiation-induced tissue and organ injury was reviewed by Robbins and Diz [50]. In the mid-1980s Ward and his colleagues first showed a modification of radiation-induced pulmonary dysfunction in rats with the use of captopril, one of the first ACEi used in humans [50]. However, enthusiasm for the use of ACEi to ameliorate radiation-induced lung fibrosis was dampened when the investigators failed to demonstrate that the radioprotection was permanent [51]. On the other hand, Moulder and his colleagues showed that, in a rat model of radiation nephropathy, the severity of both functional and histopathological injury could be reduced by captopril therapy [52]. Importantly, the protective effect of ACEi in radiation-induced renal injury was maintained, even when the treatment was non-continuous or if ACEi were used at non-hemodynamic doses [53–55]. These radioprotective effects were notably absent for radiation-induced pneumonitis. Recently, using ramipril, a lipophilic ACEi currently being used in humans, Kim and his colleagues were the first to demonstrate neuroprotection against late delayed brain injury [56]. It is significant to note that functional and histopathological protection was observed even when ramipril was administered weeks after the radiation exposure (up to 2 weeks after high single dose of radiation) [57]. More recently, they demonstrated that rampril reduced the inhibition of neurogenesis following whole brain radiation in rats [58].
Statins
Stains, HMG-Co-A reductase inhibitors, block the rate-limiting step of cholestrol biosynthesis. Clinically, statins are widely used to treat hypercholesterolemia and atherosclerosis. In addition to the primary mode of action, statins possess pleitropic effects that are under active investigation. The inhibition of cellular proliferation, the restoration of endothelial activity, the inhibition of platelet reactivity, and an antioxidant potential are only a few examples of effects attributed to statins on various vascular tissues. Statins have also been shown to possess potent anti-inflammatory properties [59]. Recent in vitro and in vivo studies demonstrated that statins reduce the generation of intracellular ROS in smooth muscle cells after stimulation of the NADPH oxidase with growth factors or angiotensin [60]. Haendeler et al. showed additional novel antioxidant mechanism by which statins reduce ROS through s-nitrosylation of thioredoxin in endothelial cells [61]. There are increasing experimental data available demonstrating the neuroprotective effects of statins after ischemic and/or traumatic brain injury [62]. Treatment of stroke in the rat with statins induced angiogenesis, neurogenesis and synaptogenesis in the ischemic brain, and the subsequent brain remodeling contributed to functional recovery [63]. Molecular mechanisms underlying the role of statins in the induction of brain plasticity and subsequent improvement of neurologic outcome after treatment of stroke are thought in part to be mediated by increased expression of brain-derived neurotrophic factor and VEGF. Statins also provided a significant neuroprotection in rats subjected to acute traumatic brain injury [64].
Using statins, Williams and her colleagues have shown a significant mitigation of radiation-induced pneumonitis in rats [65]. Statin-treated animals showed a substantial reduction in both macrophage and lymphocyte populations in the irradiated lung compared to radiation alone. More recently, statins were shown to be effective in mitigating delayed intestinal radiation injury in rats [66, 67]. We have obtained data showing significant protection against the inhibitory effects of radiation on hippocampal neurogenesis after whole brain irradiation in rats [68]. Taken together, emerging biological data at the molecular and cellular levels are disclosing an increasing number of relevant properties of statins that seem strongly connected to vascular endothelial protection and both anti-inflammatory and anti-fibrotic conditions. In addition, statins have been shown to reduce the susceptibility of lipid peroxidation [69]. These data provide indirect evidence supporting an antioxidant effect of statins, independent of their primary lipid-lowering action.
Superoxide dismutase (SOD) mimetics
Under normal physiological conditions, numerous cellular anti-oxidant systems exist to defend against oxidative stress and maintain the redox balance within the cell. ROS are cleared from the cell by enzymatic systems including SOD, catalase, and glutathione peroxidase, or non-enzymatic systems including antioxidant vitamins such as alpha tocopherol and ascorbic acid. An early clinical study using synthetic Cu/Zn SOD has shown a promising result in the treatment of radiation-induced skin fibrosis [70, 71]. Others have used different formulations of SOD and synthetic SOD mimetics and demonstrated neuroprotective effects following ischemic brain injury in the rat [72, 73]. Salen-manganese complexes are low-molecular weight synthetic compounds that exhibit both SOD and catalase activities, catalytically destroying both superoxide and hydrogen peroxide, respectively [74, 75]. The synthetic SOD mimetic (e.g., Eukarion) showed reversal of age-related learning deficits and brain oxidative stress in mice [76]. More recently, use of viral vectors (e.g., adenovirus or adeno-associated virus) carrying SOD gene is being investigated in mitigating the effect of radiation-induced skin and lung injury [77, 78].
VEGF inhibitors
VEGF is mitogenic, angiogenic, and a potent mediator of vascular permeability, being 50 times more effective than histamine [79]. VEGF causes extravasation of plasma proteins in skin bioassays and increases hydraulic conductivity in perfused microvessels [80]. Hypoxia triggers VEGF expression [81]. Although VEGF acts as a survival factor for endothelial cells and its depletion can cause vascular damage [82], excessive production of VEGF arising from activated astrocytes following the endothelial damage can cause vasogenic edema, resulting in tissue hypoxia, breakdown of the blood–brain barrier, and white matter necrosis.
Bevacizumab is a humanized monoclonal antibody that binds to VEGF and was approved by FDA for use in combination with standard chemotherapy in the treatment of metastatic colon and non-small cell lung cancer [83, 84]. In addition, several small molecule VEGF inhibitors as well as other anti-VEGF monoclonal antibodies are in various stages of clinical trials for solid tumors. Small-molecule inhibitors of VEGF receptor tyrosine kinase activity include sorafenib, sunitinib, and vandetanib. Another class of VEGF inhibitor includes VEGF Trap. VEGF trap is a fusion protein consisting of key domains from human VEGF receptors 1 and 2 with human IgG1 Fc. As a decoy receptor, it binds VEGF with high affinity and blocks all VEGF-A isoforms as well as placental growth factor [85].
Using a VEGF antagonist, Ferrara et al. was the first to show a significant reduction in the ischemia/reperfusion-related brain edema and injury, implicating VEGF in the pathogenesis of stroke and related disorders [80]. Anti-VEGF therapy can also “normalize” oxygen delivery and BBB function in tumor vasculature, improving the delivery of chemotherapeutic drugs and/or the efficacy of radiosurgery to destroy the tumor [31]. The involvement of VEGF in the pathogenesis leading to white matter necrosis following brain radiation suggests that anti-VEGF therapy may be useful for mitigating late delayed injuries. Indeed, Gonzales et al. recently treated a series of 8 patients with radiation necrosis, assessed on the basis of MRI and biopsy, with bevacizumab for 2–3 weeks. Treatment with the drug alone produced an over 50% reduction in both MRI fluid-attenuated inversion recovery abnormalities and T1-weighted Gd-contrast abnormalities [30]. It was possible to achieve an average reduction in daily dexamethasone requirements of 8.6 mg. Although long-term follow-up data are needed to establish whether brain necrosis is reduced, the preliminary data are encouraging. Once a reliable pre-clinical animal model of the radiation-induced white matter necrosis in the brain is established, more detailed molecular and cellular mechanistic studies should be carried out to determine the role of VEGF inhibitors in preventing and mitigating white matter necrosis of the brain.
References
Leung W, Hudson MM, Strickland DK et al (2000) Late effects of treatment in survivors of childhood acute myeloid leukemia. J Clin Oncol 18:3273–3279
Kim JH, Khil MS, Kolozsvary A et al (1999) Fractionated radiosurgery for 9L gliosarcoma in the rat brain. Radiat Res 45:1035–1040
van der Kogel AJ (1991) Central nervous system injury in small animal models. In: Gutin RH, Leibel SA, Sheline GE (eds) Radiation injury to the nervous system. Raven Press, New York, pp 91–111
Hopewell JW (1998) Radiation injury to the central nervous system. Med Ped Oncol 1(Suppl):1–9
Tofilon PJ, Fike JR (2000) The radioresponse of the central nervous system: a dynamic process. Radiat Res 153:357–370
Hopewell JW, van der Kogel AJ (1999) Pathophysiological mechanisms leading to the development of late radiation-induced damage to the central nervous system. Front Radiat Ther Oncol 33:265–275
Kamiryo T, Kassell NF, Thai QA et al (1996) Histological changes in the normal rat brain after gamma irradiation. Acta Neurochir 138:451–459
French-Constant C, Raff MC (1986) The oligodendrocyte type-2 astrocyte cell lineage is specialized for myelination. Nature 323:335–338
Vrdoljak E, Bill CA, van der Kogel AJ et al (1992) Radiation-induced apoptosis of oligodendrocytes in vitro. Int J Radiat Biol 62:475–480
Calvo W, Hopewell JW, Reinhold HS et al (1988) Time-and dose-related changes in the white matter of the rat brain after single doses of X-rays. Br J Radiol 61:1043–1052
Coderre JA, Morris GM, Micca PL et al (2006) Late effects of radiation on the central nervous system: role of vascular endothelial damage and glial stem cell survival. Radiat Res 166:495–503
Schuller BW, Binns PJ, Riley KJ et al (2006) Selective irradiation of the vascular endothelium has no effect on the survival of murine intestinal crypt stem cells. Proc Natl Acad Sci USA 103:3787–3792
Raju U, Gumin GJ, Tofilon PJ (2000) Radiation-induced transcription factor activation in rat cerebral cortex. Int J Radiat Biol 76:1045–1053
Belka C, Budach W, Kortmann RD et al (2001) Radiation-induced CNS toxicity-molecular and cellular mechanisms. Brit J Cancer 85:1233–1239
Tsao MN, Li YQ, Lu G et al (1999) Upregulation of vascular endothelial growth factor is associated with radiation-induced blood–spinal cord barrier disruption. J Neuropath Exp Neurol 58:1051–1060
Logan A, Berry M (1993) Transforming growth factor β1 and basic fibroblast growth factor in the injured CNS. Trends Pharmacol Sci 14:337–343
Chiang CS, Hong JH, Stadler A et al (1997) Delayed molecular responses to brain irradiation. Int J Radiat Biol 72:45–53
Schnackenberg CS (2002) Oxygen radicals in cardiovascular-renal disease. Curr Opin Pharmacol 2:121–125
Simonian NA, Coyle JT (1996) Oxidative stress in neurodegenerative diseases. Ann Rev Pharmacol 36:83–106
Ehara S, Ueda M, Naruko T et al (2001) Elevated levels of oxidized low density lipoprotein show a positive relationship with the severity of acute coronary syndromes. Circulation 103:1955–1960
Robbins MEC, Zhao W (2004) Chronic oxidative stress and radiation-induced late normal tissue injury: a review. Int J Radiat Biol 80:251–259
Lo YY, Wong JM, Cruz TF (1996) Reactive oxygen species mediate cytokine activation of c-Jun NH2-terminal kinases. J Biol Chem 271:15703–15707
Watanabe Y, Suzuki O, Haruyama T et al (2003) Interferon-gamma induces reactive oxygen species and endoplasmic reticulum stress at the hepatic apoptosis. J Cell Biochem 89:244–253
Dringen R, Gutterer JM, Hirrlinger J (2000) Glutathione metabolism in brain. Eur J Biochem 267:4912–4916
Smith KJ, Kapoor R, Felts PA (1999) Demyelination: the role of reactive oxygen and nitrogen species. Brain Pathol 9:69–92
Guan J, Stewart J, Ware JJ et al (2006) Effects of dietary supplements on the space radiation-induced reduction in total antioxidant status in CBA mice. Radiat Res 165:373–378
Li Y-Q, Ballinger JR, Nordal RA et al (2001) Hypoxia in radiation-induced blood–spinal cord barrier breakdown. Cancer Res 61:3348–3354
Lyubimova N, Hopewell JW (2004) Experimental evidence to support the hypothesis that damage to vascular endothelium plays the primary role in the development of late radiation-induced CNS injury. Br J Radiol 77:488–492
Nordal RA, Nagy A, Pintilie M et al (2004) Hypoxia and hypoxia-inducible factor-1 target genes in central nervous system radiation injury: a role for vascular endothelial growth factor. Clin Cancer Res 10:3342–3353
Gonzalez J, Kumar AJ, Conrad CA et al (2007) Effect of Bevacizumab on radiation necrosis of the brain. Int J Radiat Oncol Biol Phys 67:323–326
Winkler F, Kozin SV, Tong RT et al (2004) Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response to radiation: role of oxygenation, angiopoietin-1, and matrix metalloproteinases. Cancer Cell 6:553–563
Raber J, Rola R, LeFevour A et al (2004) Radiation-induced cognitive impairments are associated with changes in indicators of hippocampal neurogenesis. Radiat Res 162:39–47
Madsen TM, Kristjansen PE, Bolwig TG et al (2003) Arrested neuronal proliferation and impaired hippocampal function following fractionated brain irradiation in the adult rat. Neurosci 119:635–642
Fike JR, Rola R, Limoli CL (2007) Radiation response of neural precursor cells. Neurosurg Clin N Am 18:115–127
Mizumastu S, Monje M, Morhardt R et al (2003) Extreme sensitivity of adult neurogenesis to low doses of x-irradiation. Cancer Res 63:4021–4027
Saxe MD, Battaglia F, Wang JW et al (2006) Ablation of hippocampal neurogenesis impairs contextual fear conditioning and synaptic plasticity in the dentate gyrus. Proc Natl Acad Sci USA 14:17501–17506
Lynch MA (2004) Long term potentiation and memory. Physiol Rev 84:87–136
Bruel-Jungerman E, Davis S, Rampon C et al (2006) Long-term potentiation enhances neurogenesis in the adult dentate gyrus. J Neurosci 26:5888–5893
Shi L, Adams MM, Long A et al (2006) Spatial learning and memory deficits after whole-brain irradiation are associated with changes in NMDA receptor subunits in the hippocampus. Radiat Res 166:892–899
Armstrong CL, Gyato K, Awadalla AW et al (2004) A critical review of the clinical effects of therapeutic irradiation damage to the brain: the roots of controversy. Neuropsych Rev 14:65–86
Monje M, Palmer T (2003) Radiation injury and neurogenesis. Curr Opin Neurol 16:129–134
Monje ML, Mizumatsu S, Fike JR et al (2003) Irradiation induces neural precursor-cell dysfunction. Nat Med 8:955–962
Lonn EM, Yusuf S, Jha P et al (1994) Emerging role of angiotensin-converting enzyme inhibitors in cardiac and vascular protection. Circulation 90:2056–2069
Crawford DC, Chobanian AV, Brecher P (1994) Angiotensin II induces fibronectin expression associated with cardiac fibrosis in the rat. Cir Res 74:727–739
Nakajima M, Hutchinson HG, Fujinaga M et al (1995) The angiotensin II type 2 (AT2) receptor antagonizes the growth effects of the AT1 receptor: gain-of-function study using gene transfer. Proc Natl Acad Sci USA 92:10663–10667
Liu YH, Yang XP, Sharov VG et al (1997) Effects of angiotensin-converting enzyme inhibitors and angiotensin II type 1 receptor antagonists in rats with heart failure: role of kinins and angiotensin II type 2 receptors. J Clin Invest 99:1926–1935
Griendling KK, Minieri CA, Ollerenshaw JD et al (1994) Angiotensin II stimulates NADPH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ Res 74:1141–1148
Tojo A, Onozato ML, Kobayashi N et al (2002) Angiotensin II and oxidative stress in Dahl salt-sensitive rat with heart failure. Hypertension 40:834–839
Mehta JL, Hu B, Chen J et al (2003) Pioglitazone inhibits LOX-1 expression in human coronary artery endothelial cells by reducing intracellular superoxide radical generation. Arterioscl Thromb Vasc Biol 23:2203–2208
Robbins MEC, Diz DI (2006) Pathogenic role of the rennin-angiotensin system (RAS) in modulating radiation-induced late effects. Int J Radiat Oncol Biol Phys 64:6–12
Ward WF, Solliday HH, Moteni A et al (1983) Radiation injury in a rat lung. Angiotensin converting enzyme activity. Radiat Res 96:294–300
Ward WF, Kim YT, Molteni A et al (1988) Radiation-induced pulmonary endothelial dysfunction in rats: modification by an inhibitor of angiotensin converting enzyme. Int J Rad Oncol Biol Phys 15:135–140
Moulder JE, Fish BL, Cohen EP (1993) Treatment of radiation nephropathy with ACE inhibitors. Int J Radiat Oncol Biol Phys 27:93–99
Moulder JE, Fish BL, Cohen EP (1998) Radiation nephropathy is treatable with an angiotensinII type 1 (AT1) receptor antagonist. Radiother Oncol 46:307–315
Moulder JE, Fish BL, Regner KR et al (2002) Angiotensin II blockade reduces radiation-induced proliferation in experimental radiation nephropathy. Radiat Res 157:500–505
Kim JH, Brown SL, Kolozsvary A et al (2004) Modification of radiation injury by Ramipril, inhibitor of angiotensin-converting enzyme, on optic neuropathy in the rat. Radiat Res 161:137–142
Ryu S, Kolozsvary A, Jenrow KA et al (2007) Mitigation of radiation-induced optic neuropathy in rats by ACE inhibitor ramipril: importance of rampril dose and treatment. J Neuro-Oncol 82:119–124
Jenrow K, Liu J, Kolozsvary A et al (2007) Ramipril mitigates radiation-induced impairment of dentate gyrus neurogenesis. Abstract 4132. 13th International congress of radiation research
Diomede L, Albani D, Sottocorno M et al (2001) In vivo anti-inflammatory effect of statins is mediated by non-sterol mevalonate products. Arterioscler Throm Vasc Biol 21:1327–1332
Shishehbor MH, Bremen ML et al (2003) Statins promote potent systemic anti-oxidant effects through specific inflammatory pathways. Circulation 108:426–431
Haendeler J, Hoffman J, Zeiher A et al (2004) Antioxidant effects of statins via S-nitrosylation and activation of thioredoxin in endothelial cells. Circulation 110:856–861
Chen J, Zhang ZG, Li Y et al (2003/2005) Statins induce angiogenesis, neurogenesis, and synaptogenesis after stroke. Ann Neurol 53:743–751 Erratum in: Ann Neurol. 58:818
Lu D, Goussev A, Chen J et al (2004) Atorvastatin reduces neurological deficit and increases synaptogenesis, angiogenesis, and neuronal survival in rats subjected to traumatic brain injury. J Neurotrauma 21:21–32
Lu D, Qu C, Goussev A et al (2007) Statins increase neurogenesis in the dentate gyrus, reduce delayed neuronal death in the hippocampal CA3 region, and improve spatial learning in rat after traumatic brain injury. J Neurotrauma 24:1132–1146
Williams JP, Hernady E, Johnston CJ et al (2004) Effect of administration of lovastatin on the development of late pulmonary effects after whole-lung irradiation in a murine model. Radiat Res 161:560–567
Haydont V, Gilliot O, Rivera S et al (2007) Successful mitigation of delayed intestinal radiation injury uring pravastatin is not associated with acute injury improvement or tumor protection. Int J Radiat Oncol Biol Phys 68:1471–1482
Wang J, Boerma M, Fu Q et al (2007) Simvastatin ameliorates radiation enteropathy development after localized fractionated irradiation by a protein C-independent mechanism. Int J Radiat Oncol Biol Phys 68:1483–1490
Jenrow K, Liu J, Kolozsvary A et al Atorvastatin reduces impairment of dentate gyrus neurogenesis after whole brain radiation in rats (In preparation)
Li D, Chen H, Romeo F et al (2002) Statins modulate oxidized low density lipoprotein-mediated adhesion molecule expression in human coronary artery endothelial cells: role of LOX-1. J Pharmacol Exp Ther 13:601–605
Lefaix JL, Delanian S, Vozenin MC et al (1999) Striking regression of subcutaneous fibrosis induced by high doses of gamma rays using a combination of pentoxyfylline and alpha-tocopherol : an experimental study. Int J Radiat Oncol Biol Phys 43:839–847
Delanian S, Lefaix JL (2004) The radiation-induced fibroatrophic process: therapeutic perspective via the antioxidant pathway. Radiother Oncol 73:119–131
He YY, Hsu CY, Ezrin AM et al (1993) Polyethylene glycol-conjugated superoxide dismutase in focal cerebral ischemia-reperfusion. Am J Physiol 265:H252-H256
Imaizumi S, Woolworth V, Fishman RA et al (1990) Liposome-entrapped superoxide dismutase reduces cerebral infarction in cerebral ischemia in rats. Stroke 21:1312–1317
Doctrow SR, Huffman K, Marcus CB et al (1996) Salen manganese complexes: combined superoxide dismutase/catalase mimics with broad pharmacological efficacy. In: Sies H (ed) Antioxidants in “disease mechanisms and therapeutic strategies”, advances in pharmacology. Academic Press, NY, pp. 247–269
Baker K, Marcus CB, Huffman K et al (1998) Synthetic combined superoxide dismutase/catalase mimetics are protective as a delayed treatment in a rat stroke model: a key role for reactive oxygen species in ischemic brain injury. J Pharmacol Exper Therapeut 284:215–221
Liu R, Liu IY, Bi X et al (2003) Reversal of age-related learning deficits and brain oxidative stress in mice with superoxide dismutase/catalase mimetics. Proc Natl Acad Sci USA 100:8526–8531
Greenberger JS, Epperly MW, Gretton J et al (2003) Radioprotective gene therapy. Curr Gene Ther 3:183–195
Yan S, Brown SL, Kolozsvary A et al (2007) Mitigation of radiation-induced skin injury by AAV mediated MnSOD gene therapy (Submitted)
Hicklin DJ, Ellis LM (2005) Role of the VEGF pathway in tumor growth and angiogenesis. J Clin Oncol 23:1011–1057
van Bruggen N, Thigbodeaux H, Palmer JT et al (1999) VEGF antagonism reduces edema formation and tissue damage after ischemia/reperfusion injury in the mouse brain. J Clin Invest 104:1613–1620
Dibbens JA, Miller DL, Damert A et al (1999) Hypoxic regulation of vascular endothelial growth factor mRNA stability requires the cooperation of multiple RNA elements. Mol Biol Cell 10:907–919
Liu W, Ahmad SA, Reinmuth N et al (2000) Endothelial cell survival and apoptosis in tumor vasculature. Apoptosis 5:323–328
Yang JC, Haworth L, Sherry RM et al (2003) A randomized trial of bevacizumab, an anti-vascular endothelial growth factor antibody, for metastatic renal caner. N Engl J Med 349:2335–2345
Ferrara N, Hillan KJ, Gerber HP et al (2004) Discovery and development of bevacizumab, an anti-VEGF antibody for treating cancer. Nat Rev Drug Discov 3:391–400
Holash J, Davis S, Papadopoulos N et al (2002) VEGF-Trap: a VEGF blocker with potent antitumor effects. Proc Natl Acad Sci USA 99:11393–11398
Acknowledgement
The study was in part supported by NIH NIAID 5U19AI067734-020005 (JHK).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kim, J.H., Brown, S.L., Jenrow, K.A. et al. Mechanisms of radiation-induced brain toxicity and implications for future clinical trials. J Neurooncol 87, 279–286 (2008). https://doi.org/10.1007/s11060-008-9520-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11060-008-9520-x