
Abstract
The mating type (−)-specific gene of the alpha-box and the mating type (+)-specific gene of the high-mobility group (HMG) DNA-binding domain were confirmed in zoophilic dematophytes of Arthroderma simii and A. vanbreuseghemii. The sequence of the alpha-box gene was 1,375 bp, containing 2 exons (from 172 to 463 bp and from 513 to 1,375 bp) in the A. simii (−) mating type strain and 1,380 bp, containing 2 exons (from 177 to 468 bp and from 518 to 1,380 bp) in the A. vanbreuseghemii (−) mating type strain. The sequence of the HMG gene was 1,871 bp, containing 2 exons (from 181 to 362 bp and from 426 to 1,440 bp, coding a protein of 398 amino acids) in the A. simii (+) mating type strain and 1,811 bp containing 2 exons (from 158 to 339 bp and from 403 to 1,381 bp, coding a protein of 386 amino acids) in the A. vanbreuseghemii (+) mating type strain. Of 15 animal isolates and 72 human isolates examined, the alpha-box gene was detected in five of the animal isolates and in none of the human isolates, while the HMG gene was detected in the other 10 of the animal isolates and in all of the human isolates. Phylogenetic analysis of the alpha-box and HMG genes of Trichophyton mentagrophytes complex strains and the Microsporum gypseum strain revealed that these strains were divided into 4 clusters; the first cluster consisting of A. vanbreuseghemii and the isolates from animals and humans, the second cluster consisting of A. simii, the third cluster consisting of A. benhamiae and the fourth cluster consisting of M. gypseum. These results indicate that anthropophilic T. mentagrophytes evolved from the A. vanbreuseghemii (+) mating strain.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Trichophyton mentagrophytes is epidemiologically divided into two distinct forms, zoophilic and anthropophilic [1]. The zoophilic isolates of T. mentagrophytes have been generally identified by morphological and biochemical examination as well as through mating experiments. The confirmed teleomorphs of the zoophilic isolates of the T. mentagrophytes complex are Arthroderma benhamiae, A. simii and A. vanbreuseghemii [1–4]. On the other hand, no teleomorph has been identified in an anthropophilic isolate of T. mentagrophytes, such as T. mentagrophytes var. interdigitale (T. interdigitale), or in the other anthropophilic strains, such as T. rubrum. However, T. interdigitale generally reacts as a (+) type by producing pseudoascocarps when paired with tester strains of A. benhamiae, A. vanbreuseghemii or A. simii [1–4], suggesting evolution from geophilic strains to the zoophilic strains and eventually to the anthropophilic strains, including T. interdigitale strains [1]. Heidemann et al. [5] re-classified the T. mentagrophytes complex into 4 groups and established the groups of zoophilic and anthropophilic strains of T. interdigitale (related to A. vanbreuseghemii) by internal transcribed spacer (ITS) region analysis. However, the mechanism by which T. interdigitale lost the ability to mate in the process of evolving to being anthropophilic is unknown.
In a previous study, we identified the mating type (−)-specific gene of the alpha-box and the mating type (+)-specific gene of the high-mobility group (HMG) DNA-binding domain in zoophilic dematophytes of A. benhamiae [6]. In the present study, these genes were examined in the clinical isolates of zoophilic and anthropophilic T. mentagrophytes as well as in zoophilic dematophytes of A. simii and A. vanbreuseghemii by PCR analysis.
Materials and Methods
Strains
The (+) and (−) mating type strains of A. simii and of A. vanbreuseghemii (+) and (−) were used in this study. The clinical isolates of T. mentagrophytes, 15 from animals and 72 from human tinea pedis, used are listed in Table 1.
Preparation of Genomic DNA
The mycelia were obtained by culturing isolates in Sabouraud’s dextrose broth (1% peptone and 4% dextrose) at 27°C for 5 days. The mycelial cells of dermatophytes were collected by centrifugation at 1,500×g for 5 min and then frozen in liquid nitrogen and homogenized. They were then lysed in a lysis buffer containing 1 mg of zymolyase-100T/ml (Takara Bio Company, Kyoto, Japan), 0.1 mM EDTA, 0.1% sodium dodecyl sulfate (SDS), 10 mM Tris hydrochloride (pH 8.0) and 0.3% 2-mercaptoethanol, at 37°C for 16 h. High molecular weight DNA was extracted from the mycelial cells by the phenol/chloroform method. DNA samples dissolved in TE buffer (10 mM Tris–HCl, pH 8.0 and 1 mM EDTA) were used for further analysis.
Alpha-Box Gene Sequencing
To clone the alpha-box gene from (−) mating type strains (VUT-77008 and VUT-77010), primers were prepared based on the conserved sequence of the alpha-box gene of the A. benhamiae (GenBank accession number, AB570254) [6]. The primers are listed in Table 2.
Thirty-five cycles of PCR amplification were performed under the following conditions: denaturation for 1 min at 94°C, primer annealing for 1 min at 55°C and polymerization for 2 min at 72°C in a total reaction volume of 30 μl of amplification mixture containing 10 mM Tris–HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.001% gelatin, 200 mM each deoxynucleoside triphosphate, 1.0 U Taq polymerase (Takara, Kyoto, Japan) and 0.5 μg of each primer.
The PCR products were purified by gel electrophoresis and cloned into the pCRII vector (Invitrogen, San Diego, CA, USA). The plasmid DNA from more than three clones of each species was extracted using the QIAGEN plasmid kit (QIAGEN, Vallencia, CA, USA) and separately sequenced by the dideoxy chain termination method using an ABI PRISM 310 Genetic Analyzer (PerkinElmer, Inc., Foster City, CA, USA).
HMG Gene Sequencing
To clone the HMG gene 2 (+) mating type strains (VUT-77007 and VUT-77009), primers were prepared based on the conserved sequence of the HMG gene of the A. benhamiae (GenBank accession number AB542198) [6]. The primers are listed in Table 2. The PCR products obtained were sequenced using the techniques mentioned above.
PCR Analysis of the Alpha-Box and HMG Genes Specific to the Genomic DNA of Clinical Isolates of T. mentagrophytes
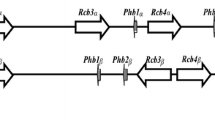
The primers TmMATa1S and TmMATa1R amplified a 471-bp fragment of the A. vanbreuseghemii alpha-box gene fragment. The primers TmHMG1S and TmHMG1R amplified a 524-bp fragment of the A. vanbreuseghemii HMG fragment.
Genomic DNA samples (100 ng) from the clinical isolates were amplified by PCR in a volume of 30 μl, using a reaction mixture containing 10 mM Tris–HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.001% gelatin, 200 mM each deoxynucleoside triphosphate, 1.0 unit of Taq polymerase (Takara) and 0.5 μg of a pair of primers. Amplification was carried out over 35 cycles consisting of template denaturation (1 min, 94°C), primer annealing (1 min, 55°C) and polymerization (2 min, 72°C). The PCR products obtained were sequenced using the techniques mentioned above.
Phylogenetic Analysis of the Alpha-Box and HMG Genes
The DNA sequences were compared using Clustal W multiple sequence alignment programs [7], and a phylogenetic tree was constructed by the TREEVIEW displaying phylogenies program [8]. Bootstrap analysis was performed on 1,000 random samples and analyzed by the Clustal W programs [9].
Results
Alpha-Box Genes of A. simii and A. vanbreuseghemii
The sequence of the alpha-box gene of the A. simii (−) mating type was determined to be 1,375 bp, which contained 2 exons from 172 to 463 bp and from 513 to 1,375 bp, coding a protein of 384 amino acids, beginning with a putative initiating methionine (ATG).
The sequence of the alpha-box gene of the A. vanbreuseghemii (−) mating type was determined to be 1,380 bp, which contained 2 exons from 177 to 468 bp and from 518 to 1,380 bp, coding a protein of 384 amino acids, beginning with a putative initiating methionine (ATG).
The amino acid sequence of the A. simii alpha-box gene (GenBank accession number: AB605768) shared approximately 86.4, 92.6 and 78.1% sequence similarity with the A. benhamiae, A. vanbreuseghemii and Microsporum gypseum alpha-box amino acid sequences (GenBank accession number: AB570254, AB605769 and FJ798800) (Fig. 1), respectively, in the conserved region.

Comparison of the homologous regions of the predicted protein sequences of the (−) mating type strains of the A. simii alpha-box (VUT-77008) (GenBank AB605768), A. benhamiae alpha-box (AB570254), A. vanbreuseghemii alpha-box (AB605769) and M. gypseum alpha-box (FJ798800). An asterisk indicates identity with the amino acid found in the A. simii alpha-box sequence
HMG Genes of A. simii and A. vanbreuseghemii
The sequence of the HMG gene of the A. simii (+) mating type was determined to be 1,871 bp, which contained 2 exons, from 181 to 362 bp and from 426 to 1,440 bp, coding a protein of 398 amino acids, beginning with a putative initiating methionine (ATG).
The sequence of the HMG gene of the A. vanbreuseghemii (+) mating type was determined to be 1,811 bp, which contained 2 exons, from 158 to 339 bp and from 403 to 1,381 bp, coding a protein of 386 amino acids, beginning with a putative initiating methionine (ATG). We determine the exons and introns of HMG gene A. simii and A. vanbreuseghemii by comparing with HMG gene A. benhamiae and Microsporum gypseum HMG (GenBank accession number: AB542198 and FJ798798).
The amino acid sequence of the A. simii HMG gene (GenBank accession number: AB605766) shared approximately 88.5, 88.1 and 65.3% sequence similarity with the A. benhamiae, A. vanbreuseghemii and M. gypseum HMG amino acid sequences (GenBank accession number: AB542198, AB605767 and FJ798798) (Fig. 2), respectively, in the conserved region.

Comparison of the homologous regions of the predicted protein sequences of the (+) mating type strains of A. simii HMG (VUT-77009) (GenBank AB605766), A. benhamiae HMG (AB542198), A. vanbreuseghemii HMG (AB605767) and M. gypseum HMG (FJ798798) (GenBank AB605768). An asterisk indicates identity with the amino acid found in the A. simii HMG sequence. The underlined amino acids indicate the highly conserved HMG signature sequence “PRPPNAFILYR” and “PRKPSEKKRR” of filamentous ascomycetes (2)
PCR Detection of Alpha-Box and HMG Genes Specific to the Clinical Isolates of T. mentagrophytes
Of 15 animal isolates and 72 human isolates examined, the alpha-box gene was detected in 5 of the animal isolates and in none of the human isolates, while the HMG gene was detected in the other 10 of the animal isolates and in all of the human isolates.
Sequence similarity among clinical isolates of T. mentagrophytes and the type strains
The nucleotide sequence similarity was 99–100% among the alpha-box gene fragments from the 5 animal isolates and the regions of the type strain of A. vanbreuseghemii (−) mating type deposited in the database (DDBJ accession no. AB605769).
Similarly, the nucleotide sequence similarity was 99–100% among the HMG gene fragments from the 10 animal isolates and the regions of the type strain of A. vanbreuseghemii (+) mating type deposited in the database (DDBJ accession no. AB605769). Moreover, the nucleotide sequence similarity was 100% among the HMG gene fragments from the 72 human isolates and the A. vanbreuseghemii (+) mating type strain deposited in the database (DDBJ accession no. AB605769).
Phylogenetic Tree for the Alpha-Box and HMG Genes
Phylogenetic analysis of the alpha-box gene of T. mentagrophytes complex strains revealed that they were divided into four clusters; the first cluster consisting of A. vanbreuseghemii isolates and the five animal isolates, the second cluster consisting of A. simii, the third cluster consisting of A. benhamiae and the forth cluster consisting of M. gypseum (Fig. 3).

A tree showing phylogenetic relationships among the obtained alpha-box gene sequences from dermatophyte species. Numbers at branches were determined by bootstrap analysis, indicating the times in 1,000 repeat sub-samples in monophylogenic grouping. Bracket indicates the DDBJ session number of alpha-box gene analysis of dermatophyte species
Phylogenetic analysis of the HMG gene of the T. mentagrophytes complex strains revealed that they were also divided into four clusters; the first cluster consisting of A. vanbreuseghemii and the 10 animal isolates and all 72 human isolates, the second cluster consisting of A. benhamiae, the third cluster consisting of A. simii and the forth cluster consisting of M. gypseum (Fig. 4).

A tree showing phylogenetic relationships among obtained HMG gene sequences of dermatophyte species. Numbers at branches were determined by bootstrap analysis, indicating the times in 1,000 repeat sub-samples in monophylogenic grouping. Bracket indicates the DDBJ session number of HMG gene analysis of dermatophyte species
Discussion
In this study, we identified the mating type (−)-specific gene of the alpha-box and the mating type (+)-specific gene of the HMG DNA-binding domain in A. simii and A. vanbreuseghemii. Kawasaki et al. [10] reported that these teleomorphs, A. benhamiae, A. simii and A. vanbreuseghemii, could mate among themselves and might be cospecific, although these species are from distant lineages. The amino acid sequence of the alpha-box gene in A. simii shared approximately 86.4, 92.6 and 78.1% similarity with those from A. benhamiae, A. vanbreuseghemii and M. gypseum, respectively, while the amino acid sequence of the HMG gene in A. simii shared approximately 88.5, 88.1 and 65.3% similarity with those in A. benhamiae, A. vanbreuseghemii and M. gypseum, respectively. These results indicate that more than 85% homology of alpha-box or HMG genes could be cospecific in dermatophytes.
PCR detected the HMG gene in all human isolates examined but did not detect the alpha-box gene in any human isolate. The HMG gene of anthropophilic isolates of T. interdigitale (T. mentagropytes from tinea pedis), with no mating ability, showed 100% homology with that of A. vanbreuseghemii. This result supports previous findings that T. interdigitale reacts as a (+) type by producing pseudoascocarps when paired with tester strains of A. benhamiae, A. vanbreuseghemii or A. simii [1].
Heidemann et al. [5] reported that the internal transcribed spacer (ITS) region of T. interdigitale is also genetically close to that of T. mentagrophytes from human tinea pedis and distinct from that of T. mentagrophytes from animals and human tinea corporis. Moreover, Symoens et al. [11] have shown that all the isolates from tinea pedis and tinea unguium identified as T. interdigitale based on ITS sequences mated with A. vanbreuseghemii tester strains, but had lost their ability to give fertile cleistothecia. Therefore, they suggested that T. interdigitale has to be considered as a humanized species derived from the sexual relative A. vanbreuseghemii [5, 11].
In the present study, the nucleotide sequences of the HMG gene fragments from 10 animal isolates with mating ability and 72 human isolates showed 99% (3–4 nucleotide changes in 420 bp) and 100% similarity, respectively, with those of the (+) mating type strain of A. vanbreuseghemii deposited in the database. The phylogenetic tree of the HMG gene indicates that the Japanese isolates of T. mentagrophytes from human tinea pedis are genetically close to animal isolates (Fig. 4). Therefore, these anthropophilic isolates of T. mentagrophytes may have derived from a strain of the A. vanbreuseghemii (+) mating type and then widely developed.
A lack of sexual ability of fungi could be caused by (1) degeneration of sex genes, (2) increased heterozygosity among alleles, (3) a lack of transportable elements and (4) linkage disequilibria [11]. Moreover, genes at the mating locus and in the pheromone response pathway are highly conserved, even in apparently asexual fungi [11]. The present results regarding the anthropophilic isolates of T. mentagrophytes (T. interdigitale) from tinea pedis suggest that the sex genes might be degenerated and that heterozygosity among alleles might be increased, since the mating locus was highly conserved.
Evolutionary theory suggests that sex is expensive and inefficient, as it requires the formation of non-identical haploid gametes, which must fuse to regenerate a diploid [12]. Even in fungi, the need to maintain two mating partners reduces fitness [12]. However, sex emerged early in evolution, and most eukaryotes undergo a sexual cycle [12].
After Homo sapiens appeared on the earth around 200,000 years ago, the anthropophilic T. mentagrophytes would have evolved from the A. vanbreuseghemii (+) mating strain, losing its mating ability and proliferating on the host, while allowing recombination in stressful conditions [12]. Therefore, clinical isolates of anthropophilic T. interdigitale may be genetically close to the A. vanbreuseghemii (+) mating strain due to the lack of MAT.
References
Kwon-Chung KJ, Bennett EJ. Dermatophytosis: medical mycology. Philadelphia: Lea & Febiger; 1992. p. 136–61.
Ajello L, Cheng S-L. The perfect state of Trichophyton mentagrophytes. Sabouraudia. 1967;4:230–4.
Stockdale MP, Mackenzie WRD, Austwick KCP. Arthroderma simii sp. nov., the perfect state of Trichophyton simii (Pinoy) comb. nov. Sabouraudia. 1967;4:112–23.
Takashio M. The Trichophyton mentagrophytes complex. In: Iwata K, editor. Recent advances in medical and veterinary mycology. Tokyo: University of Tokyo Press; 1977. p. 271–6.
Heidemann S, Monod M, Gräser Y. Signature polymorphisms in the internal transcribed spacer region relevant for the differentiation of zoophilic and anthropophilic strains of Trichophyton interdigitale and other species of T. mentagrophytes sensu lato. Br J Dermatol. 2010;62:282–95.
Kano R, Yamada T, Makimura K, Kawasaki M, Mochizuki T, Kamata H, Hasegawa A. Arthroderma benhamiae (The Teleomorph of Trichophyton mentagrophytes) mating type-specific genes. Mycopathologia. 2011;171:333–7.
Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl Acid Res. 1994;22:4673–80.
Page RDM. TREEVIEW: an application to display phylogenetic trees on personal computers. Comput Appl Biosci. 1996;12:357–8.
Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39:783–91.
Kawasaki M, Anzawa K, Wakasa A, Takeda K, Mochizuki T, Ishizaki H, Hemashettar B. Matings among three teleomorphs of Trichopyton mentagrophytes. Nippon Ishinkin Gakkai Zasshi. 2010;51:143–52.
Symoens F, Jousson O, Planard C, Fratti M, Staib P, Mignon B, Monod M. Molecular analysis and mating behaviour of the Trichophyton mentagrophytes species complex. Int J Med Microbiol. 2011;301:260–6.
Butler G. Fungal sex and pathogenesis. Clin Microbiol Rev. 2010;23:140–59.
Acknowledgments
This study was supported by grants-in-aid from the Academic Frontier Project of the Japanese Ministry of Education, Culture, Sports, Science and Technology (MEXT) and grants-in-aid from Nihon University. We thank Dr. T. Kasai (Kasai Dermatological Hospital), Dr. T. Kusunoki (Kusunoki Dermatological Hospital), Dr. M. Fujihiro, (Dept. of Dermatology, Ibi General Hospital) and Dr. H. Noguchi (Noguchi Dermatological Hospital) for providing the clinical isolates used in this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kano, R., Kawasaki, M., Mochizuki, T. et al. Mating Genes of the Trichophyton mentagrophytes Complex. Mycopathologia 173, 103–112 (2012). https://doi.org/10.1007/s11046-011-9487-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11046-011-9487-6