
Abstract
For mass production of urease B subunit (UreB) and heat shock protein A subunit (HspA) of Helicobacter pylori with Bombyx mori nuclear polyhedrosis virus (BmNPV) baculovirus expression system (BES) and to determine whether they could be used as an oral vaccine against H. pylori, besides, to determine the time course of expressed recombinant protein and the optimum acquisition time directly through green fluorescence, HspA and enhanced green fluorescence protein (EGFP) genes were cloned into vector pFastBacDual to form donor vector pFastBacDual-(EGFP) (HspA), UreB gene was cloned into vector pFastBacDual to form donor vector pFastBacDual-UreB,then they were transformed into E. coli BmDH10Bac to obtain the recombinant Bacmid-(EGFP) (HspA) and Bacmid-UreB respectively. They were used to transfect BmN cells and generated the recombinant baculovirus BmNPV-(EGFP) (HspA) and BmNPV-UreB. Using these recombinant baculovirus BmNPV-(EGFP) (HspA) and BmNPV-UreB inoculated the silkworm pupae, a recombinant HspA and UreB protein were expressed in silkworm pupae, which were around 13 and 62 kDa in sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and western blot analysis. After oral immunization of mice, serum specific IgG antibodies against HspA and UreB in vaccine group were much higher than that in mock and native silkworm powder control groups. The results indicated that the expressed recombinant HspA and UreB in silkworm pupae would possess good immunogenicity. In addition, when EGFP and HspA proteins were expressed, a direct correlation between the increase in intensity of fluorescence and HspA concentration.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
H. pylori infection remains a significant global public health problem. This pathogen is a major cause of chronic gastritis, gastric and duodenal ulcers, and gastric carcinomas, mucosa-associated lymphoid tissue (MALT) lymphoma and primary gastric non-Hodgkin’s lymphoma, and is considered as a class I carcinogen by the World Health Organization [1–4]. Vaccine development against this infection appears to be a preferable strategy [5]. Many studies have demonstrated that H. pylori in infected mouse stomach can be cleared by oral immunization with H. pylori urease [6–9]. HspA can be used as a vaccine candidate because of its strong antigenic properties [10–12]. Vaccine has been applied successfully as a potential therapeutic strategy for preventing and treating. In recently, human H. pylori oral subunit vaccine (UreB + LTB) has been approved for use in China, its anti-H. pylori infection protective time is a year and protective rate is 72.1%. However, the establishment of long-term oral immunity to protect of H. pylori infection has not yet been reported. The ability to induce long-term immunity to H. pylori is necessary for the repeated administration of antigens. Therefore, the recombinant therapeutic protein production becomes a major bottleneck. The potential therapeutic applications in humans would be limited by the low levels of production (the available expression technologies such as prokaryotes, cultivated mammalian cells, and transgenic animals and plants) and the high cost (the use of expensive fermentation techniques and stringent purification protocols). The development of recombinant protein production in the silkworm baculovirus expression system (BES) may overcome these limitations and thus may facilitate the use of transgenic edible proteins. It is potential for not only low-cost but also high capacity production (up to 20% of total cell protein) [13]. These features make the silkworm as an ideal system of expression and delivery package for producing oral medical proteins.
In order to determine the time course of expressed recombinant proteins in silkworm pupae and the optimum acquisition time. The traditional way is that haemolymph from 10 silkworm larvae or pupae is harvested every 12 h beginning at 72 h post-infection and stored at −20°C. Subsequently, the haemolymph is analysed by SDS-PAGE or serial diluted by ELISA to detect the time phase of maximum expression products, which is a very time-consuming and labor-intensive procedure and takes up a lot of resources. This drawback has become a hard problem for practical and industrial utilization of silkworm bioreactor and academic research field.
Enhanced green fluorescence protein (EGFP) should be the most favorable biotechnological tools used to investigate the function of target gene by visualizing, monitoring, and quantifying in living cells [14–16]. We considered these properties of EGFP, in this study, we took advantages of EGFP to measure target protein HspA optimum expression time phase by a one-step visualization. However, in many cases, protein fusion can influence protein folding of each fusion partner, expression level, and function loses its antigen-binding activity, which shows a little fluorescence activity [17]. We tried to use a dual expression baculovirus to express the EGFP and target HspA gene under P10 and Pph promoters by a non-fusion way. The Pph and pl0 promoters have been used to drive expression of many foreign genes originating from prokaryotes or eukaryotes [18–20]. We tried to determine whether high-level expression of antigen HspA though Pph promoters and UreB though P10 promoters could be obtained in this expression system and whether its expression production possesses strong immunogenicity.
Materials and methods
Materials and cell line and silkworm
UreB and HspA genes (cloned from H. pylori ATCC 43504) and EGFP gene templates were preserved in our laboratory. pFastBacDual was purchased from Invitrogen (San Diego, CA) and FUGENE 6 transfection reagent was the product of Roche Applied Science (United States). The DH10Bac/BmNPV E. coli was supplied by Prof. W.B. Wang. The insect cell culture medium TC-100 was purchased from Sigma. B. mori cell line, BmN (BmN4), originating from the ovary of silkworms was preserved in our laboratory and cultured at 27°C with TC-100 medium with 10% fetal bovine serum (FBS). Hybrid strain larvae of silkworm (Qingsong × Haoyue) were reared on mulberry leaves in this study for recombinant protein expression.
Cloning of HspA, EGFP genes and UreB gene
A pair of primers termed FP and RP were designed as follows: according to EGFP gene and HspA of Helicobacter pylori gene (accession number. GeneBank GQ357182 and AY295084) EGFP: FP: 5′-GCCTCGAGATGGTGAGCAAGGGCGAGGAC-3′ RP: 5′-AG GGTACCTTACTTGTACAGCTCG TCCATG-3′ HspA: FP: 5′-GCGAATTCATGAAGTTTCTACCATTAGG-3′; RP: 5′-AGTCTAGATTAGTGTTTTTTGTGATCAT G-3′. EGFP: XhoI and KpnI; HspA: EcoRI and XbaI recognition sites were introduced into the 5′-terminal of FP (underlined) and RP (underlined), respectively; UreB of Helicobacter pylori gene (accession number. GeneBank AY714224) FP: 5′-GGCTCGAG ATGATTAGCAGAAAAGAATATG-3′; RP: 5′-TTGGTACCCTAGAAAATGCTAAAGAGTTG-3′. XhoI and KpnI recognition sites were introduced into the 5′-terminal of FP (underlined) and RP (underlined), respectively, to facilitate cloning. The polymerase chain reaction (PCR) was done under the following conditions: preheating at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min, and extension at 72°C for 1 min (EGFP and HspA genes) 1.5 min (UreB gene). The final extension was performed at 72°C for 10 min. The PCR products were analyzed by agarose gel electrophoresis and recovered using a TaKaRa agarose gel DNA purification kit (Japan). The fragments were then ligated into the TaKaRa pMD18-T cloning vector and sequenced using the dideoxychain termination method with ABI 3730 DNA Analyzer. We named the obtained plasmids pMD18-T-EGFP, pMD18-T-HspA and pMD18-T-UreB.
Construction and proliferation of recombinant Bacmid Baculoviruses
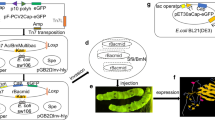
pMD18-T-EGFP was digested by XhoI and KpnI. pMD18-T-HspA was digested by EcoRI and XbaI. The EGFP and HspA fragments were inserted into multiple cloning site (MCS) of the donor plasmid pFastBacDual and transformed into E. coli DH10Bac/BmNPV. Through inside transposition in bacteria, The EGFP and HspA genes (target genes) were transferred to a baculovirus shuttle vector (bacmid) within a mini-att Tn7 target site and formed a recombinant bacmid baculovirus containing target genes, i.e., the EGFP gene and HspA gene. The E. coli DH10Bac containing recombinant bacmid baculoviruses was propagated on Luria–Bertani (LB) medium which contained antibiotics (50 μg/ml kanamycin, 7 μg/ml gentamicin, and 10 μg/ml tetracycline), 100 μg/ml X-gal, and the inducer, 40 μg/ml IPTG. The white (Lac−) colonies were selected for further amplification. The large recombinant bacmid baculoviruses rBacmid/BmNPV/(EGFP) (HspA) were isolated from the DH10Bac/BmNPV and analyzed by PCR with the M13 forward and M13 reverse primers. The isolated recombinant bacmid baculoviruses rBacmid/BmNPV/(EGFP) (HspA) were transfected into BmN cells using FuGENE 6 transfection reagent under the ratio 3:2 (3 μl of FUGENE 6 and 2 μg of bacmid DNA, according to the instruction of Roche). The P1 viral stock was gathered from the cell medium containing virus 120 h post-transfection. We amplified the viral stock through further infection of BmN cells using the P1 stock until the P3 stock was obtained. The P3 stock was stored at 4°C and preserved from light. pMD18-T-UreB was digested by XhoI and KpnI. The UreB fragment was inserted into multiple cloning site (MCS) of the donor plasmid pFastBacDual and transformed into E. coli DH10Bac/BmNPV. We obtained P3 rBacmid/BmNPV/UreB high titer viral stock according to above method.
Expression of EGFP, HspA genes and UreB gene in silkworm pupae
P3 stock recombinant virus (wild baculoviruses as control) was introduced into the silkworm pupae by means of subcutaneous injections delivering 2 μl of P3 stock at about 2 × 105 particles per pupa. Express of EGFP in infected silkworm pupae was visualized and photographed under fluorescence microscope in complete darkness every 12 h starting at 72 h post-infection. The rBacmid/BmNPV/(EGFP) (HspA) infected silkworm pupae haemolymph was collected every 12 h starting at 72 h post-infection for determining the expression course of HspA protein. The hemolymph was centrifuged for 5 min at 8,000 rpm and 4°C. The supernatant was preserved at −20°C for further use. The hemolymph of rBacmid/BmNPV/UreB infected silkworm pupae was collected using the same method for further use.
Quantification of HspA protein levels at different time point in silkworm pupae
The HspA protein levels in silkworm pupae was determined by quantitative ELISA assay. A 96-well microtiter plate was loaded with serial dilutions of the hemolymph in bicarbonate buffer, pH 9.6 (15 mM Na2CO3, 35 mM NaHCO3), and incubated overnight at 4°C. The plate was washed three times in PBS containing 0.05% Tween-20 (PBST). The plate was blocked by incubation in 1% bovine serum albumin (BSA) in PBS (100 μl/well) for 2 h at 37°C, followed by three washes with PBST. The plate was incubated in a 1:8000 dilution of rabbit anti-HspA polyclone antibody (100 μl/well) at 37°C for 2 h, followed by three washes with PBST. The plate was then incubated with a 1:2000 dilution of anti-rabbit IgG conjugated with horseradish peroxidase (100 μl/well) for 2 h at 37°C and washed three times with PBST. Finally, the chromogenic substrate o-phenylenediamine (100 μl/well) was added to the wells, and the plate was incubated for 20 min at 37°C to develop color, followed by the addition of 2 M H2SO4 (50 μl/well) to stop the reaction. The plate was cooled to room temperature. The optical density of each well was measured at 492 nm.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and western blot analysis
Both the hemolymph and the lysed cells were subjected to 12% SDS-PAGE after boiling with the same volume of loading buffer (100 mM Tris Cl, pH 6.8, 200 mM DTT, 4% SDS, 0.2% bromophenol blue, and 20% glycerol) for 5 min. The gel was stained with Coomassie Brilliant Blue R-250. The proteins were electrophoretically transferred to a PVDF membrane after SDS-PAGE. The membrane was firstly blocked in PBST (137 mM NaCl, 2.68 mM KCl, 8.06 mM Na2HPO4, 1.47 mM KH2PO4, and 0.1% Tween 20) plus 1% BSA for 2 h at 25°C and washed three times (each for 10 min) with PBST. The film was then incubated with PBST containing 1% BSA and diluted rabbit anti-HspA of H. pylori and rabbit anti-UreB of H. pylori antibody (1:300) for 2 h at 25°C and subsequently washed three times (each for 10 min) with PBST. After incubation for 1 h at 25°C in the same PBST with peroxidase-labeled goat anti-rabbit IgG (1:500) and being washed with PBS (137 mM NaCl, 2.68 mM KCl, 8.06 mM Na2HPO4 and 1.47 mM KH2PO4), the immunoactive protein was detected by adding DAB chromogenic substrate (2 mg DAB, 20 μl of 30% H2O2, and 10 ml of PBS).
Animal and breeding conditions
Forty Specific-pathogen (H. pylori)-free (SPF) female BALB/c mice purchased from animal experiment center of Medical College, Soochow University,China were used between the ages of 8 and 12 weeks. The animals were housed in polycarbonate cages and fed on a commercial pellet diet with water ad libitum. All procedures were conducted in accordance with the P. R. China legislation under No. 8910M047 on the use and care of laboratory animals and with the guidelines established by the Institute for Experimental Animals of Soochow University and were approved by the university committee for animal experiments.
Immunogen preparation and immunization of mice and antigen-specific antibody assays
The pupa expressed recombinant HspA (rHspA) and recombinant UreB (rUreB) 96 h post-inoculation with recombinant virus rBacmid/BmNPV/(EGFP) (HspA) and rBacmid/BmNPV/UreB were collected and dried with a vacuum dryer. The dried pupae were homogenized to powder and stored at −20°C up to use. Mice were immunized orally via feeding needles with a priming dose on day 0 and booster doses on days 10, 20 and 30, as follows: Immunogen groups: 200 mg silkworm powder (including rHspA or rUreB) and 10 μg cholera toxin B subunit (CTB) which could be used as a mucosal immune adjuvant, suspended in 0.5 ml of 0.2 M sodium bicarbonate, administered by gavage, using a ball-end feeding needle. Control groups: 0.5 ml 0.2 M sodium bicarbonate and 0.5 ml 0.2 M sodium bicarbonate with 200 mg silkworm powder (without including rHspA and rUreB) and 10 μg CTB. Serum samples, collected from the tail veins of mice at 10th day post the each time inoculation, were used to titrate humoral responses against H. pylori HspA or UreB. Serial dilutions of individual sera were tested in enzyme-linked immunosorbent assays (ELISAs), The determination was performed on 96-well ELISA plates coated with 10 μg of HspA/well or UreB/well. and incubated for 90 min at 37°C or overnight at 4°C. After incubation the wells were washed with PBS-0.3% Tween 20. Then blocked with PBS-1% bovine serum albumin (BSA) for 1 h at 37°C. Serum samples were added at an initial dilution of 1:200 in duplicate, with 1:3 serial dilutions performed in PBS-1% BSA Plates were incubated for 1 h at 37°C and then washed in PBS-0.3% Tween 20. A 1:5,000 dilution of rabbit anti-mouse IgG -horseradish peroxidase conjugates were added to the plates for 1 h at 37°C. Plates were washed again, developed with o-phenylenediamine as the substrate for 10 min, and stopped with 2 M H2SO4. The optical density of each well was measured at 492 nm.
Statistical analysis
The primary analysis was a comparison between immunized groups and mock controls. All data were presented as mean ± SEM. With all statistical analyses by t test, differences between groups were considered statistically significant for P < 0.05.
Results
Construction and proliferation of recombinant baculovirus rBacmid/BmNPV/(EGFP) (HspA) and rBacmid/BmNPV/UreB
The target EGFP and HspA genes were inserted into the MCS of the donor plasmid pFastBacDual under P10 promoter within XhoI and KpnI sites and polyhedron promoters within EcoRI and XbaI sites by T4 DNA ligation and transformed to E. coli DH10Bac/BmNPV. Through inside transposition in bacteria, the EGFP and HspA genes (target genes) were transferred to a baculovirus shuttle vector (bacmid) within a mini-att Tn7 target site and formed a recombinant bacmid baculovirus containing target gene. This transposition reaction occurred in the presence of transposase supplied by the helper plasmid. The white colonies (Lac−) were selected from the plate containing 50 μg/ml kanamycin, 10 μg/ml tetracycline, 7 μg/ml gentamicin, 100 μg/ml X-gal, and 40 μg/ml IPTG. The large recombinant plasmids Bacmid/BmNPV (EGFP) (HspA) were isolated from the white colonies for analysis. PCR was done by using the M13 forward and M13 reverse as primers and recombinant Bacmid DNA as template, which was too large to perform a restriction analysis. The result indicated the successful generation of recombinant Bacmid/BmNPV/(EGFP) (HspA). Then the purified recombinant baculoviruses were used to transfect the BmN cells of B. mori by liposomes. The transfected cells typically increased their cell diameter and nuclei, stopped growing, and exhibited lysis and mass (Fig. 1) 120 h post-transfection. After three transfection cycles, a high viral titer of P3 viral stock was collected and protected from light at 4°C. The target UreB gene was inserted into the MCS of the donor plasmid pFastBac Dual under P10 promoter within XhoI and KpnI sites by T4 DNA ligation and transformed to E. coli DH10Bac/BmNPV. We obtained high titer rBacmid/BmNPV/UreB as above methods.

Phenotype of BmN cells infected with recombinant virus Bacmid-UreB a normal cells b infected cells
Expression of EGFP, HspA genes and UreB gene in pupae
The P3 stock (wild virus as control). Virus were injected to silkworm pupae by a syringe. After being infected with this recombinant virus, the silkworm pupae presented typical symptoms of BmNPV infection (Fig. 2). The silkworm pupae was inoculated with rBacmid/BmNPV/(EGFP) (HspA),which typical kinetic intensity of green fluorescence was shown in (Fig. 3a) for an infection performed. The intensity of green fluorescence increased after infection until 72 h post-infection, when the maximum was reached at 96 h post-infection, the intensity of green fluorescence remained at its almost constant for up to 24 h and thereafter it declined (consistent with the result of quantification of EGFP protein ELISA, date not shown). The time of maximum intensity of green fluorescence coincided with the onset of rHspA protein production, which occurred at 72 h post-infection and continued to increase until about 108 h post-infection. During 96–120 h, the concentration of rHspA remained almost constant by ELISA and SDS-PAGE analysis (Fig. 3b, c). A direct correlation with the time phase of maximum intensity of green fluorescence and the time phase of maximum recombinant protein concentration can be seen. Therefore, EGFP can be used as a tool to estimate the optimum harvest time by a non-fusion protein way in a dual promoter BES. Meanwhile, the hemolymph was harvested 96 h post-inoculation and centrifuged for 5 min at 12,000 rpm and 4°C The supernatant were examined using 12% SDS-PAGE and Western blotting. A band around 13 kD was visualized and positively identified using Western blot analysis (Fig. 4). This band indicated the successful expression of rHspA in the silkworm pupa and rHspA possessed immunogenicity. The same method was used to assess the expression of UreB in the infected pupae. We found a band around 62 kDa (Fig. 5). This band indicated the successful expression of rUreB in the silkworm pupa and rUreB possessed immunogenicity.

Symptom of silkworm pupa infected with recombinant virus BmNPV-UreB a normal pupa b infected pupa

a The time courses of EGFP expressed in Silkworm pupae by the intensity of green fluorescence visualization. The photograph of the pupae was taken every 12 h from 72 h post-infection using fluorescent illuminator in complete darkness respectively. b The time courses of HspA expressed in Silkworm pupae by the SDS-PAGE analysis. M. Protein marker, the pupae haemolymph was collected on ice every 12 h. From 72 h post-infection. c The time courses of HspA expressed in Silkworm pupae by the sandwich-ELISA. The pupae haemolymph was collected on ice every 12 h from 72 h post-infection. The hemolymph was diluted with 1,000 folds dilution

Analysis of recombinant HspA expression in silkworm pupae with SDS-PAGE and Western blotting. (1) Protein marker; (2) silkworm pupa at 96 h post-infection with wild virus; (3) silkworm pupa at 96 h post-infection with recombinant virus; (4) western blotting of control; (5) western blotting band of HspA

Analysis of recombinant UreB expression in silkworm pupae with SDS-PAGE and Western blotting. (1) Protein marker; (2) silkworm pupa at 96 h post-infection with wild virus; (3) silkworm pupa at 96 h post-infection with recombinant virus; (4) western blotting of control; (5) western blotting band of UreB
H. pylori HspA or UreB-specific serum IgG response
Serum IgG anti-H. pylori HspA or UreB-specific antibodies were detected at 10 days post the each time inoculation. There were significantly higher levels of specific HspA or UreB antibodies IgG in the group treated with silkworm powder including rHspA or rUreB than in the control mice vaccinated with only sodium bicarbonate group and silkworm powder (without including rHspA or rUreB) group (P < 0.01, n = 10) (Fig. 6). The result suggested that vaccination with silkworm powder including rHspA or rUreB enhanced the immune response.

Titers of serum IgG specific anti-H. pylori HspA or UreB in mice. a Vaccinated with sodium bicarbonate b Vaccinated with silkworm powder (without including rHspA or rUreB) c Vaccinated with silkworm powder (including rHspA) d Vaccinated with silkworm powder (including UreB) (P < 0.01, n = 10)
Discussion
With the development of biotechnology, B. mori has been used as an important bioreactor for the production of recombinant proteins through baculovirus expression system (BES) [21, 22]. The BES is an inherently safe platform for the production of vaccines and other biologics. Baculoviruses have a narrow host range, restricted primarily to Lepidopteran species, which are non-infectious to vertebral animals [23, 24]. An oral vaccine expressed in silkworm pupae using BES offers several advantages over other existing oral vaccine technologies: (i) Expression of recombinant proteins in silkworm pupae using BES is a highly efficient protein production platform, which can large-scale production of foreign proteins; (ii) As an edible medicine, the purification of expression of heterologous proteins in the silkworm pupae is not necessary. The reports have indicated that the protein of silkworm pupae has multiple physiologic and pharmacologic activities, and can be used as a pharmaceutical intermediate, a dietary supplement, or a food additive [25, 26]; (iii)The cost of producing recombinant protein in silkworm is much lower than the cost of producing the same amount by E. coli or S. cerevisiae fermentation, because silkworms can be conveniently reared with mulberry leaves at much lower cost; (iv)The proteinase inhibitors and biocapsule-like fat in silkworm bodies may increase the stability of the recombinant proteins so that gastrointestinal degradation would not be a problem. The heterologous proteins would be protected from gastric enzymes, acid and other harmful aspects of the environment.
In this study, we utilized a non-fusion co-expression system technology for expressing EGFP and HspA foreign proteins in silkworm pupae simultaneously. This method was initially developed utilizing EGFP expression under the control of the P10 promoter and target protein HspA expression under the control of the Pph promoter. The pl0 gene was activated a few hours earlier than the polyhedrin gene [27]. Therefore, the expression level of EGFP was higher and fluorescence appears brighter due to its earlier expression in silkworm. Thus, EGFP fluorescence could be used as an estimate of target gene expression without harvesting haemolymph and SDS-PAGE or ELISA analysis. In our experiment, we found that strong EGFP fluorescence of the silkworm pupae correlated significantly with the highest expression levels of target protein HspA, as determined by SDS-PAGE and ELISA analysis (Fig. 3a, b, c). This co-expression system allowed for an easier determination of the time course of expression, leading to a simplification of the optimum acquisition time determination step. Expression of EGFP under P10 promoter had no detectable effect on the expression yield of HspA gene under Pph promoter, comparison of the expression levels of HspA expressed alone or together with EGFP, as determined by ELISA analysis (date not shown).
To our knowledge, this is the first report on the production of HspA and UreB using silkworm. In the infected silkworm pupae, the recombinant HspA and UreB proteins were efficiently released into the pupae hemolymph. On average, 0.4 ml of hemolymph was obtained per pupa. The ELISA results showed that the maximum amount of HspA protein in the hemolymph reached 0.52 mg/ml at the 108 h or so post-infection (Fig. 3b, c) and the maximum amount of UreB protein in the hemolymph reached 0.49 mg/ml by ELISA. These results demonstrated that HspA protein expressed under Pph promoter and UreB protein expressed under P10 promoter are all high level. Our results also revealed that expression products of baculovirus expressing system were well immunogenic. In our experiment for HspA and UreB expression, Western blotting study indicated that BES expressing rHspA and rUreB of H. pylori were strongly immunogenic and were specifically recognized by rabbit anti-HspA polyclonal antibody sera (Fig. 4) and rabbit anti-UreB polyclonal antibody sera (Fig. 5). In animal model experiment, detecting the results of specific serum IgG against HspA and UreB of H. pylori revealed that rHspA and rUreB induced a marked immune response (Fig. 6). In conclusion, our results have demonstrated that a high-level expression of the HspA and UreB genes can be obtained in the silkworm pupae though dual promoter system, and the expressed recombinant HspA and UreB in silkworm pupae possess good immunogenicity. The availability of large quantities of HspA and UreB that the silkworm provides should greatly facilitate the future research and testing of these proteins for potential application in medicine. It is hoped that silkworm will provide a convenient, feasible and effective means of treating clinical diseases, such as an oral vaccine against H. pylori infection.
References
Nakamura S, Matsumoto T, Suekane H, Takeshita M, Hizawa K, Kawasaki M, Yao T, Tsuneyoshi M, Iida M, Fujishima M (2001) Predictive value of endoscopic ultrasonography for regression of gastric low grade and high grade MALT lymphomas after eradication of Helicobacter pylori. Gut 48:454–460
Uemura N, Okamoto S, Yamamoto S, Matsumura N, Yamaguchi S, Yamakido M, Taniyama K, Sasaki N, Schlemper RJ (2001) Helicobacter pylori infection and the development of gastric cancer. N Engl J Med 345:784–789
Casella G, Buda CA, Maisano R, Schiavo M, Perego D, Baldini V (2001) Complete regression of primary gastric MALT-lymphoma after double eradication Helicobacter pylori therapy: role and importance of endoscopic ultrasonography. Anticancer Res 21(2B):1499–1502
Graham DY (2000) Helicobacter pylori infection is the primary cause of gastric cancer. J Gastroenterol 35(Suppl 12):90–97
Raghavan S, Hjulstrom M, Holmgren J, Svennerholm AM (2002) Protection against experimental Helicobacter pylori infection after immunization with inactivated H. pylori whole-cell vaccines. Infect Immun 70:6383–6388
Kleanthous H, Myers GA, Georgakopoulos KM, Tibbitts TJ, Ingrassia JW, Gray HL (1998) Rectal and intranasal immunizations with recombinant urease induce distinct local and serum immune responses in mice and protect against Helicobacter pylori infection. Infect Immun 66(6):2879–2886
Guy B, Hessler C, Fourage S, Haensler J, Vialon-Lafay E, Rokbi B (1998) Systemic immunization with urease protects mice against Helicobacter pylori infection. Vaccine 16(8):850–856
Corthesy-Theulaz I, Porta N, Glauser M, Saraga E, Vaney AC, Haas R (1995) Oral immunization with Helicobacter pylori urease B subunit as a treatment against Helicobacter infection in mice. Gastroenterology 109(1):115–121
Ermak TH, Giannasca PJ, Nichols R, Myers GA, Nedrud J, Weltzin R (1998) Immunization of mice with urease vaccine affords protection against Helicobacter pylori infection in the absence of antibodies and is mediated by MHC Class II-restricted responses. J Exp Med 188(12):2277–2288
Suerbaum S, Thiberge JM, Kansau I, Ferrero RL, Labigne A (1994) Helicobacter pylori hspA-hspB heat-shock gene cluster: nucleotide sequence, expression, putative function and immunogenicity. Mol Microbiol 14:959–974
Ferrero RL, Thiberge JM, Kansau I, Wuscher N, Huerre M, Labigne A (1995) The GroES homolog of Helicobacter pylori confers protective immunity against mucosal infection in mice. Proc Natl Acad Sci USA 92:6499–6503
Kansau I, Guillain F, Thiberge JM, Labigne A (1996) Nickel binding and immunological properties of the C-terminal domain of the Helicobacter pylori GroES homologue (HspA). Mol Microbiol 22:1013–1023
Massotte D (2003) G protein-coupled receptor overexpression with the baculovirus insect cell system: a tool for structural and functional studies. Biochim Biophys Acta 1610:77–89
Van Roessel P, van Roessel AH (2002) Imaging into the future: visualizing gene expression and protein interactions with fluorescent proteins. Nat Cell Biol 4:E15–E20
Lippincott-Schwartz J, Snapp E, Kenworthy A (2001) Studying protein dynamics in living cells. Nat Rev Mol Cell Biol 2:444–456
Katagiri Y, Ingham KC (2002) Enhanced production of green fluorescent fusion proteins in a baculovirus expression system by addition of secretion signal. Biotechniques 33:24–26
Kye SY, Junho C, Kwang HP, Kisu K, Shin YI, Chayong C, Miejae I, Uhhyun K (2004) Expression system for enhanced green fluorescence protein conjugated recombinant antibody fragment. Hybrid Hybridomics 23:279–286
Jianmei L, Wan CD, Denis TMI, Xinghua L, Guangli L, Xiaofeng W, Wanfu Y, Chuanxi Z, Yungen M (2009) High-level expression of orange fluorescent protein in the silkworm larvae by the Bac-to-Bac system. Mol Biol Rep 36(2):329–335
Liyun W, Yuri STM, Shogo MYO (2008) Expression, purification and characterization of yeast protein disulfide isomerase produced by a recombinant baculovirus-mediated silkworm, Bombyx mori, pupae expression system. Biotechnol Lett 30:625–630
Li ZY, Yi YZ, Yin XP, Zhang ZF, Liu JX (2008) Expression of foot-and-mouth disease virus capsid proteins in silkworm-baculovirus its utilization as a subunit vaccine. PLoS ONE 3(5):1–7
Miyajima A, Schreurs J, Otus K, Kondo A, Arai K, Maeda S (1987) Use of the silkworm, Bombyx mori, and an insect baculovirus vector for high-level expression and secretion of biologically active mouse interleukin-3. Gene 58:273–281
Maeda S (1994) Expression of foreign genes in insect cells using baculovirus vectors. In: Maramorosch K, McIntosh AH (eds) Insect cell biotechnology. CRC Press, Boca Raton, pp 1–31
Carbonell LF, Miller LK (1987) Baculovirus interaction with nontarget organisms: a virus-borne reporter gene is not expressed in two mammalian cell lines. Appl Environ Microbiol 53:1412–1417
Herrington DA, Losonsdy GA, Smith G, Vovovitz F, Cochran M, Jackson K, Hoffman SL, Gordon DM, Levine MM, Edelman R (1992) Safety and immunogenicity in volunteers of a recombinant plasmodium falciparum circumsporozoite protein malaria vaccine produced in Lepidopteran cells. Vaccine 10:841–846
Wang BG, Sun XX, Zhao LY, Zhang GY, Mei SJ, An Y, Wang C (2002) Study on nutritional value of silkworm pupa powder. China Public Health 18(5):575–576
Yang HX, Zhu XR, Lu HS (2002) Research progress on application of silkworm pupa in medical science. Bull Sci Technol 18(4):318–322
Roelvink PW, van Meer MM, de Kort CAD, Possee RD, Hammock BD, Vlak JM (1992) Dissimilar expression of Autographa californica multiple nucleocapsid nuclear polyhedrosis virus polyhedrin and p l0 genes. J Gen Virol 73:1481–1489
Acknowledgments
This work was supported by research grants from the National Basic Research Program (973 Program) (No. 2005CB121000) and Key Fostering Project for Application Research of Soochow University.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhang, X., Shen, W., Lu, Y. et al. Expression of UreB and HspA of Helicobacter pylori in silkworm pupae and identification of its immunogenicity. Mol Biol Rep 38, 3173–3180 (2011). https://doi.org/10.1007/s11033-010-9988-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-010-9988-2