
Abstract
Context
Fire regimes in many dry forests of western North America are substantially different from historical conditions, and there is concern about the ability of these forests to recover following severe wildfire. Fire refugia, unburned or low-severity burned patches where trees survived fire, may serve as essential propagule sources that enable forest regeneration.
Objectives
To quantify the influence of fire refugia spatial pattern and other biophysical factors on the process of post-fire tree regeneration; in particular examining both the proximity and density of surrounding refugia to characterize the landscape of refugial seed sources.
Methods
We surveyed regeneration at 135 sites in stand-replacement patches across a gradient of fire refugia density in eastern Oregon, USA. We characterized the influence of refugial seed source pattern and other environmental factors on the abundance of regenerating seedlings, and examined the relationship between post-fire climate and the temporal pattern of ponderosa pine seedling establishment.
Results
Tree seedlings were present in 83% of plots 12–17 years post-fire, and densities varied substantially (0–67800 stems ha−1, median = 1100). Variation in seedling abundance was driven by the spatial patterns of refugial seed sources. Despite widespread post-fire shrub cover, high-severity burned forests have not undergone a persistent type conversion to shrublands. Ponderosa pine seedling establishment peaked 5–11 years after fire and was not closely associated with post-fire climate.
Conclusions
Fire refugia and the seed sources they contain fostered tree regeneration in severely burned patches. Management practices that reduce refugia within post-fire landscapes may negatively influence essential forest recovery processes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The ability of forest ecosystems to maintain composition and function is currently challenged by changing climate and fire regimes. In much of the American west, fire regimes in dry mixed-conifer forests are substantially different from historical reference conditions (Allen et al. 2002; Hessburg et al. 2005), and the combined effects of fire suppression, logging, and grazing have significantly altered dry mixed-conifer forest structure and composition (Hagmann et al. 2013; Merschel et al. 2014). Increases in stand densities and fuel continuity are resulting in uncharacteristically severe fire effects, particularly when fires escape initial fire suppression efforts due to extreme fire weather conditions (Miller and Safford 2012; Stephens et al. 2013). Large patches of high-severity, stand-replacement fire effects in dry forest ecosystems may be slow or unable to regenerate due to seed source limitations (Haire and McGarigal 2010; Chambers et al. 2016), while increasingly hot and dry climatic conditions may further limit post-fire forest regeneration (Donato et al. 2016; Tepley et al. 2017; Stevens-Rumann et al. 2018). Projected increases in fire activity and climate warming are heightening concerns that high-severity fire will precipitate transformations of forests to alternative stable states dominated by shrub or herbaceous vegetation (Savage and Mast 2005; Stephens et al. 2013; Coop et al. 2016). Understanding the patterns and drivers of forest regeneration following high-severity fire is essential for determining the long-term resilience of forest ecosystems in the context of accelerating global change.
Fires create burn severity mosaics that include unburned and low-severity areas where the dominant vegetation survives, referred to here as fire refugia. As the locations least disturbed by a fire event, fire refugia in some ecosystems may host fire-sensitive plant communities (Wood et al. 2011; Adie et al. 2017), late-successional forest structure (Camp et al. 1997; Ouarmim et al. 2016), and legacy seed sources important to post-fire forest recovery (Landesmann and Morales 2018). Unlike species that are capable of resprouting following fire (e.g., quaking aspen) or producing aerial seed banks (e.g., lodgepole pine), obligate seeding conifer species rely on seeds dispersed from surviving trees in fire refugia or from seed sources outside the fire perimeter to regenerate in stand-replacement patches where fire has killed overstory forest. Average dispersal distances for these species are primarily a function of seed weight (Vander Wall 2003; Safford 2013). Numerous studies have reported that distance to nearest refugial seed source is a key control on post-fire conifer regeneration (e.g., Donato et al. 2009; Harvey et al. 2016; Kemp et al. 2016), illustrating how spatial patterns of fire effects influence ecological process such as forest regeneration (Turner et al. 1997). However, these patterns must be quantified in meaningful ways in order to understand their interactions with ecological function (Turner 1989). One-dimensional measures of distance to nearest seed source, which commonly are used in post-fire regeneration studies, may be inadequate to capture the spatial complexity of burn mosaics where multiple seed sources are likely to influence a site’s capacity to regenerate forest. For example, seed availability adjacent to a small refugium composed of one or two mature conifers may be very different than the seed availability adjacent to multiple, large refugia patches composed of many pre-fire trees. Quantifying the pattern of fire refugia in a way that accounts for the additive effect of seed sources in multiple dimensions may provide a more ecologically relevant metric for understanding variability in ecological processes like post-fire forest regeneration (Haire and McGarigal 2010).
Abiotic and biotic factors interact with seed source availability to mediate the pace and trajectory of post-fire forest reestablishment. Conifer seedlings are vulnerable to heat and drought stress (Rother et al. 2015; Petrie et al. 2016), and heat from insolation may be an important factor limiting post-fire seedling establishment in stand-replacement patches, especially during hot, dry years (Burns and Honkala 1990). North-facing aspects, higher elevations, and shade from surviving trees can buffer seedlings from heat and drought stress, promoting higher rates of establishment and survival compared to more exposed landscape positions (Chappell and Agee 1996; Collins and Roller 2013; Dodson and Root 2013; Chambers et al. 2016). Additionally, conifer seedlings must compete with other species that are well adapted for rapidly colonizing high-severity burned areas. A robust post-fire shrub response following high-severity fire in dry mixed-conifer forests is common (Powell 1994), and dense shrub cover can decrease conifer seed germination and seedling growth (Zavitkovski et al. 1969; Burns and Honkala 1990). However, shrub cover in stand-replacement patches may also ameliorate unfavorable environmental conditions, facilitating conifer establishment and survival in some cases (Shatford et al. 2007; Collins and Roller 2013).
Prior studies of post-fire regeneration in dry forests in the American west report widely divergent vegetation recovery trajectories. High-severity burned areas were regenerating rapidly in parts of the Pacific Northwest (Shatford et al. 2007; Donato et al. 2009; Crotteau et al. 2013) and northern Rockies (Kemp et al. 2016), while forest regeneration was considerably slower or absent in parts of the southern Rockies (Chambers et al. 2016), California (Collins and Roller 2013; Welch et al. 2016) and the Southwestern US (Haire and McGarigal 2010). Additionally, evidence is emerging that post-fire drought conditions are resulting in widespread regeneration failure (Stevens-Rumann et al. 2018), and some dry forests may be near a tipping point as they are unable to recover between fire events, resulting in extensive forest loss (Tepley et al. 2017). These studies, which provide evidence for both forest resilience and forest vulnerability to high-severity fire, highlight the importance of understanding post-fire regeneration patterns across different geographic regions that contribute to the gradient of environmental variability in western dry forest ecosystems.
In this study, we quantify conifer seedling regeneration in stand-replacement patches within four large fires that occurred in Oregon’s Blue Mountains to examine the influence of fire refugia pattern on forest reestablishment, while accounting for other biotic and abiotic factors known to influence post-fire tree regeneration. Although there has been concern about the vulnerability of Blue Mountain forests to high-severity fire for decades (Hall 1976), their current capacity to recover following stand-replacement fire is poorly understood.
Recent large fires in the Blue Mountains provide an opportunity to examine post-fire forest regeneration across broad temperature and precipitation gradients. In addition, new methods for characterizing fire refugia patterns as estimates of seed source using high-resolution aerial imagery may offer novel insights into the key drivers of post-fire forest recovery. Focusing on landscapes that burned between 2000 and 2005 we asked: (1) How does post-fire conifer regeneration following stand-replacement fire vary with patterns of fire refugia, quantified in terms of both point-wise distance to refugial seed source and a surface density of surrounding refugia? (2) How do fire severity, understory vegetation, and environmental gradients influence post-fire conifer regeneration in conjunction with patterns of fire refugia? (3) What are the temporal patterns of seedling establishment of ponderosa pine (Pinus ponderosa), the most widespread conifer species in our study area, following stand-replacement fire?
Methods
Study region
The Blue Mountains ecoregion extends from central Oregon east to the Snake River Plain near the Idaho border, and north to the Columbia River in southeastern Washington (Fig. 1). The Blue Mountains are composed of a series of small sub-ranges bisected by rugged river canyons. The majority of precipitation falls in the winter as snow, and thunderstorms that ignite wildfires are common during warm, dry summers (Burns 1983). The Cascade Mountains to the west effectively block marine air masses from reaching the drier, southern portion of the range (Johnson and Clausnitzer 1992), whereas the Columbia River provides an ingress for these systems, resulting in a comparatively cooler and wetter climate in the northern Blue Mountains (Heyerdahl et al. 2001). The forested extent of the Blue Mountains spans broad environmental gradients: from 1981 to 2010, the 30-year average maximum August temperature range was 16 to 34 °C, the average minimum January temperature range was -12 to 1 °C, and the annual precipitation range was 200 to 1800 mm across the study region (PRISM Climate Group 2018).

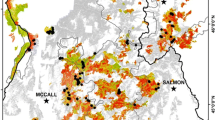
The study area and study fires in central and northeastern Oregon. a The study was performed in the Blue Mountains ecoregion. b All four study fires burned primarily on land managed by the US Forest Service (USFS). c–f Fire refugia (green) within our four study fires, mapped using 1-m resolution aerial imagery, illustrate the complex spatial patterning of surviving forest. Unforested areas outside of fire refugia (white) were not all forested prior to fire, particularly at the highest elevations in the Roberts Creek fire (meadows), and the lowest elevations in the 747 fire (grasslands and shrublands). Plot locations were located outside of fire refugia in areas forested prior to fire that experienced stand-replacement fire effects
Dry mixed-conifer forests in the Blue Mountains occupy the warmer, drier portions of these climatic gradients, and are composed primarily of ponderosa pine, Douglas-fir (Pseudotsuga menziesii), grand fir (Abies grandis), and lesser amounts of western larch (Larix occidentalis) and lodgepole pine (Pinus contorta). Grand fir and white fir (A. concolor) hybridize across their ranges in Oregon (Ott et al. 2015), and we refer to their variants here as grand fir. Dry mixed-conifer forests in the Blue Mountains contain components of western juniper (Juniperus occidentalis) at lower elevation sites, and Engelmann spruce (Picea engelmannii) and subalpine fir (A. lasiocarpa) at the highest elevation sites.
Fires burned frequently in the Blue Mountains prior to fire exclusion which began in the late 1800s. Pre-settlement fire return intervals in the southern Blue Mountains averaged 10–21 years (Johnston et al. 2016), and slightly longer in the northern part of the range where the climate is cooler and wetter (Heyerdahl et al. 2001). Historically, frequent fires produced a range of low- and mixed-severity effects that supported a heterogeneous landscape resilient to disturbance and variations in climate (Hessburg and Agee 2003; Stine et al. 2014). Contemporary fire return intervals for ponderosa pine and Douglas-fir/grand fir forests in the Pacific Northwest are now estimated to be an order of magnitude longer than prior to fire exclusion, and the amount and scale of high-severity fire effects is widely considered outside the historical range of variability (Reilly et al. 2017).
Study design
We collected data during the summer of 2017 in four large fires that burned in the years 2000, 2002, and 2005, 12–17 years prior to sampling (Fig. 1 c–f). We selected fires that burned primarily in designated roadless or wilderness areas to minimize the influence of active land management, and we avoided areas for which we had a record of timber harvest, fuels treatments, or tree replanting. All sampled fires were lightning-caused. We sampled exclusively in areas that were forested prior to fire, and that experienced 100% overstory mortality within our plots. The study was designed across a gradient of fire refugia density, which aimed to represent variability in two-dimensional landscape surface estimates of seed source.
The first step in quantifying the fire refugia surface density metrics was to identify fire refugia as patches of surviving overstory tree canopy. We mapped fire refugia using post-fire aerial imagery at 1-m resolution from the National Agriculture Imagery Program (NAIP). Our processing follows Chambers et al. (2016), with some modifications. Aerial imagery provides a finer resolution complement to widely used Landsat burn severity products, and the 1-m resolution allowed us to accurately map fire refugia within fire perimeters at the scale of single surviving trees. NAIP imagery was acquired for 2012 and 2014, 7–14 years after the fire events, and 3–5 years prior to field data collection in 2017. NAIP imagery dates were chosen to allow for the full realization of second-order fire effects (e.g., delayed mortality) and to limit possible misclassification of post-fire recolonization as surviving, pre-fire trees. Images were classified with a maximum likelihood algorithm using training samples to produce 1-m resolution raster grids with two classes, “refugia” and “non-refugia” (Fig. 1c–f). Classified rasters were subsequently converted into polygons based on a simple rule set: all adjacent refugia were considered members of the same polygon; polygons within 20 m of each other were aggregated; and gaps < 50 m2 were subsumed by the polygon within which they were embedded. The layers were edited manually to a limited degree to remove any un-forested areas and to include any surviving tree canopies missed during automated image classification. Maps were validated using the source 1-m NAIP imagery (total accuracy = 0.92–0.95, κ = 0.84–0.90) and ground-truthed using data collected in the field (total accuracy = 0.90–0.95, κ = 0.80–0.89).
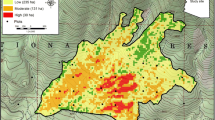
Maps of refugia were used to calculate landscape surfaces of distance-weighted refugia density. For each 1-m pixel, fire refugia density values were based on the amount and location of fire refugia cells within a 300 × 300-m moving window (with an additional row-column pair for the focal cell), using a two-dimensional, area-based, distance-weighted density approach. Within the moving window surface, the value of each cell (1 = refugia, 0 = non-refugia) was divided by its distance from the focal cell. These values were then summed to produce what we refer to here as refugia density for each focal cell (Fig. 2).

a Fire refugia density was calculated by dividing the value of each cell (1 = refugia, 0 = non-refugia) by its distance from the focal cell and summing these values for the 300 × 300-m window. b Cell weights calculated with the refugia density approach for a reduced window size (6 × 6, plus an additional row-column pair for the focal cell). Distributions on the top and left demonstrate cell weights exponentially decreasing with distance from the focal cell. The method was intended to account for the exponential decrease in seed dispersal and post-fire seedling regeneration with increased distance from seed source reported by previous studies. The resulting surface incorporates the potential contribution of nearby seed sources, as well as more distant seed sources at the scale of the moving window
The result of the refugia density calculation was a continuous metric ranging from zero, in areas where fire refugia were absent, to approximately 1000, in areas saturated with fire refugia. The refugia density surface metric extends the concept of a dispersal kernel, which describes the scatter of offspring around the parent plant in the form of a probability density function (Clark 1998), to a landscape dispersal kernel (Haire and McGarigal 2010). In their application of spatial seed dispersal kernels in dry forest ecosystems, Haire and McGarigal (2010) utilized Gaussian weights to model seed availability, with an input map of ponderosa pine canopy cover. Here, we developed the distance-weighted refugia density method with the goal of more closely approximating the widely documented exponential decrease in seed density (Greene and Johnson 2000) and post-fire seedling abundance with distance from seed source (e.g., Chambers et al. 2016; Kemp et al. 2016) and applied it to our maps of residual trees. The choice of window size was based on observed seed dispersal distances from prior studies. Kemp et al. (2016) reported the probability of post-fire seedling presence was low beyond 95 m in similar forests in the Northern Rockies. However, studies in dry mixed-conifer forests in the Klamath-Siskiyou region reported that longer dispersal events (> 95-m) were common (Shatford et al. 2007; Donato et al. 2009). The 300 × 300-m window size accounts for potential longer-distance seed dispersal (i.e., 150 m), while the distance-weighted approach reflects the exponentially decreasing probability of such dispersal events from more distant trees, compared to the likelihood of dispersal from nearby seed sources.
To identify field sampling locations we generated a population of random sample points stratified along the refugia density gradient for each study fire, focused on sites that experienced stand-replacement fire effects. We identified areas of stand-replacing/high-severity fire in the landscape using the differenced Normalized Burn Ratio (dNBR) derived from 30-m Landsat TM/ETM + satellite imagery from the Monitoring Trends in Burn Severity (MTBS) project. From the dNBR products we constrained sampling to areas that experienced moderate/high or high-severity (> 440 dNBR, Key and Benson 2006). We expected this constraint to exclude areas like grasslands, scree fields, and bare ground that were not forested prior to fire. The dNBR constraint was not imposed in the case of our smallest study fire (Fig. 1f: Burnt Cabin, 800 ha), and in this case we used field assessments to restrict sampling to areas forested prior to fire that experienced stand-replacement fire effects. We excluded all areas within 150 m of fire perimeters to limit the influence of seed sources outside the fire perimeter on both post-fire seedling regeneration and the methods used to quantify refugia density. Areas within 150-m of roads were also excluded to limit the influence of human-made, non-forested areas on the methods used to calculate refugia density, and to exclude the locations likely to have been most heavily impacted by fire suppression activities. To facilitate access, all sample plots were within either one km of a trail or three km of a road.
Final site selection was made in the field based on four criteria. We rejected plot locations if: (a) there was no evidence of forest prior to fire, (b) locations exhibited potentially confounding management histories, (c) surviving pre-fire trees (i.e., fire refugia) were present in the sample plot, or (d) the terrain was inaccessible or unsafe. Plot locations were separated by a minimum distance of 150 m to reduce possible spatial autocorrelation. While abiotic gradients were not explicitly included in our stratification, we attempted to representatively sample the range of aspects and elevations available within each fire while attending to our other constraints.
Field data collection
Field data were collected from 5.64-m radius circular plots (100 m2) in the summer of 2017. We tallied all post-fire tree seedlings within each plot and recorded species, height, and diameter at breast height (DBH; if height was > 1.37 m) for the three individuals of each tree species closest to plot center. Seedlings were recorded as overtopped if they were growing beneath the shrub canopy and no part of the seedling had emerged above the shrub canopy. Current-year seedlings, defined as seedlings that germinated in the spring of the year of sampling, were excluded due to uncertainty as to whether they would persist through the growing season. Where ponderosa pine seedlings were present, we estimated establishment dates for the three seedlings closest to plot center by counting branch whorls.
We quantified plot structure and composition to provide context for the tree regeneration data. We recorded DBH for all snags and downed trees that were present prior to fire and rooted in the plot, and converted these measurements into an estimate of total pre-fire basal area. Due to the amount of time that had passed since fire, we were unable to determine the mortality agent for all dead trees. However, our intent was not to quantify fire-induced mortality but rather to characterize pre-fire basal area as a proxy for site productivity. Elevation, aspect, slope position, and slope shape (flat, convex, or concave) were recorded for each plot. For all vascular plant species occupying greater than 0.25% of the plot area (0.25 m2) we recorded species, maximum height, and percent cover. Plant species abundance data were subsequently aggregated by lifeform: trees, shrubs (woody, non-tree species), herbs (grasses, forbs, and sedges). Total shrub cover was subsequently used in the analysis.
We collected point-wise estimates of distance to nearest refugial seed source in the field to complement our landscape surface metric of refugia density. We measured the distance from plot center to the three closest living, pre-fire seed sources for each species observed in the plot using a laser rangefinder (TruPulse 200). When seed sources for a particular species were beyond the range of the rangefinder (~500-m) or not visible from plot center, we assigned a value of 600 m to represent distances > 500 m in analyses. Spatial locations were recorded for each plot center using a handheld GPS (Garmin GPSMAP 64). Location data were used to assign a refugia density value to each field plot for subsequent analysis. Although some registration errors were likely as a result of our GPS units’ accuracy limitations, we believe it is unlikely that such errors introduced bias into the data.
To identify relationships between annual climate conditions and temporal trends in ponderosa pine seedling establishment, we collected a sample of seedlings and cross-referenced whorl-based age with age based on growth rings. We opportunistically sampled seedlings outside of wilderness areas within all four fires. We cut 21 ponderosa pine seedlings at the root-shoot boundary, and cross-sections from the samples were finely sanded until the cell structure was visible with a binocular microscope (AmScope, 45X). We counted the number of growth rings for each sample and compared these counts to our field-based, whorl-count age estimates. Field-based estimates were strongly correlated with age estimates derived from growth rings (R2 = 0.7), but the whorl-count method underestimated seedling age by an average of 2 years.
Climate environment
To characterize the climate space associated with each field plot, we analyzed both existing geospatial data and data collected in the field. We calculated heat load to estimate potential annual solar radiation based on slope, aspect, and elevation data measured in the field (McCune and Keon 2002, Eq. 3). To quantify climate at a coarser scale, we calculated mean annual temperature, mean annual precipitation, and mean annual climate moisture deficit (CMD) using data acquired from Climate WNA, an application that allows users to downscale climate data from moderate spatial resolution (800 × 800-m) grids to point locations (Wang et al. 2016). Hargreaves CMD is an annual metric that is the sum of the monthly difference between reference evaporation and precipitation (Zhao et al. 2012). Climate data were acquired for 2000 to 2015 and averaged across years for each plot location to characterize the general contemporary climatology of each plot. We included climate data for post-fire years only, beginning with year of burn, to quantify the post-fire conditions experienced by regenerating seedlings.
Data analysis
We produced summary statistics from our seedling data to characterize the amount and composition of conifer seedlings regenerating in stand-replacement patches. We calculated the percentage of plots occupied by each species to identify the most common regenerating species, and totaled seedling counts for each species to determine the most abundant regenerating species. In addition, we calculated the mean and median seedling densities for each conifer species across all sample plots. To understand the relationship between non-tree, woody vegetation and regenerating seedlings, we calculated the average height for each conifer species, the percentage of each species overtopped by shrubs, and mean percent shrub cover. Lastly, we used boxplots to examine seedling densities across plots and among fires.
We developed statistical models to examine the variability in seedling abundance as a function of point-wise and surface density refugial seed source metrics (distance to seed source, refugia density) and ecological site characteristics described below. We fit generalized linear mixed models for the three most common conifer species in our study area: ponderosa pine, Douglas-fir, and grand fir individually, as well as a model for all conifer species combined. The all-conifer model included our three focal species, and western larch, subalpine fir, Engelmann spruce, and western juniper. We chose not to include lodgepole pine because we expected this species would not be as dependent on refugial seed source because of its capacity to produce serotinous cones. We considered both negative binomial and zero-inflated negative binomial models, and included fire identity (e.g., Hash Rock) as a random effects term to account for among-fire variability in seedling counts not otherwise explained by our fixed effects model parameters.
Our first step was to identify the best subset of refugial seed source metrics to parsimoniously represent seed sources potentially contributing to regeneration. We considered minimum, median, and mean values to represent point-wise distance to nearest seed source, and fire refugia density to represent the amount and pattern of surrounding fire refugia in two-dimensions. Mean and median distances to nearest seed source were calculated based on the distance to the three nearest surviving, pre-fire trees measured in the field. When fewer than three seed sources were visible, or one or more seed sources were beyond the range of the laser rangefinder, we averaged only measured distances and did not include the 600-m placeholder value. When no seed sources for a given species were visible from the plot center or within the range of the laser rangefinder, we used the value 600-m to allow for the inclusion of these plots in the analysis despite the absence of distance to seed source observations. Initial models were fit with a single seed source metric, as well as a combined model including both distance to seed source and refugia density. We compared all surviving seed source metrics based on ΔAIC (Akaike information criterion) and P-values, relative to a null model including only an intercept term.
Next, to understand the spatial variability of post-fire regeneration as a function of both abiotic and biotic factors, we developed a series of regression models that included the seed source metrics that explained the most variability in observed post-fire seedling abundance and a suite of site characteristics. Site characteristics were chosen based on results from similar studies and included elevation, understory vegetation, fire severity (dNBR), pre-fire basal area, and climate environment. We examined all site characteristics for collinearity, using a Spearman’s ρ > |0.7|. Temperature was excluded from the analysis because it was collinear with elevation, and because the GPS elevation data were likely more representative of variability among the sample points than the 800 × 800-m interpolated temperature data. Mean annual precipitation was collinear with CMD, and we chose to exclude precipitation in favor of CMD because CMD integrates both precipitation and temperature data as a measure of the moisture needed for vegetation growth that must be met from sources other than rain to avoid the impacts of drought (Wang et al. 2012). Exploratory plots indicated potential non-linear relationships between observed seedling densities and gradients of elevation and CMD (Supplemental Figs. 1 and 2). To account for this possibility, we included a quadratic term for elevation and CMD in our final models, in addition to linear terms. We also tested for an interaction between shrub cover and CMD as a way to test our prediction that seedling recruitment might diminish in hotter, drier locations due to physiological stress, but that shrub cover could ameliorate those unfavorable climatic conditions.
We evaluated the significance of model parameters based on P-values (α = 0.05) as well as ΔAIC comparisons relative to a full model. The effect sizes of surviving seed source parameters (distance to seed source, refugia density) were quantified in two ways. First, we compared model predictions for low (25th percentile of observed) and high (75th percentile of observed) values of distance to seed source and refugia density, while holding all other model parameters constant at their median values. Second, we generated a series of added variable plots to graphically examine the marginal effects of seed source parameters on predicted post-fire seedling abundance when all other model parameters are held constant at their median values. We tested the sensitivity of our models to the placeholder value of 600 m for distance to seed source by substituting larger values (1000 m, 2000 m) but observed no significant change in parameter estimates (Δ < 0.001). All analyses were implemented in R (ver. 3.4.0, R Core Team 2017). Model fitting and selection was performed using glmmTMB (Brooks et al. 2017), and model predictions were generated using lme4 (Bates et al. 2014).
Finally, to examine relationships between temporal trends in ponderosa pine seedling establishment and annual climate conditions, we averaged annual CMD for each fire. For each year and each fire, we plotted CMD values and the number of ponderosa pine seedlings established.
Results
Post-fire conifer regeneration was widespread across our study sites (n = 135). Regeneration was observed in 83% of sampled plots, and densities varied by five orders of magnitude, ranging from 0 to 67,800 stems ha−1. The median seedling density for all plots was 1100 stems ha−1 (Table 1). The most widely distributed species were ponderosa pine, grand fir, and Douglas-fir, which were present in 61, 54, and 47% of plots, respectively (Fig. 3a), and these species accounted for 54% of recorded seedlings. Western larch and lodgepole pine were less widespread, occurring in 30% and 28% of plots, respectively. Both larch and lodgepole pine tended to regenerate at high densities when present and accounted for 39% of seedlings tallied (Fig. 3b). The remaining 7% of seedlings consisted of western juniper, Engelmann spruce, subalpine fir, western white pine (Pinus monticola), and whitebark pine (Pinus albicaulis). The mean height for all seedlings was 100 cm (SE = 0.03), with ponderosa pine, Douglas-fir, and grand fir seedlings shorter on average than western larch and lodgepole pine (Table 1). Shrubs overtopped 40% of measured seedlings (Fig. 4), although the percentage overtopped varied considerably by species (Table 1).

a Percentage of plots occupied by the five most common regenerating conifer species. Ponderosa pine (PIPO), Douglas-fir (PSME), and grand fir (ABGR) were the most widespread species regenerating in our study area. b The total number of seedlings recorded across all field plots for the five most common conifer species. Grand fir was the most abundant species regenerating in our study area. Western larch and lodgepole pine were regenerating densely where present, which accounts for their high total seedling counts compared to ponderosa pine and Douglas-fir

Photos of regenerating seedlings above (highlighted in white) and below the shrub layer. a Many post-fire seedlings in our study area were overtopped by shrubs, b while others appear to have recently emerged from the shrub canopy
We sampled across broad environmental gradients; plot locations ranged in elevation from 900 to 2140 m (mean = 1600 m), and CMD ranged from 340 to 717 mm (mean = 505 mm) (Table 2). Average fire severity according to dNBR was 560 (152–966), and average pre-fire basal area was 17.4 (0–78) m2 ha−1. Average shrub cover was 54% (Fig. 5), and only one plot had 0% shrub cover. Average shrub height was 0.92 m (± 0.07 SE), slightly lower than the average seedling height. Ceanothus velutinus and C. sanguineus were the dominant shrub species and occurred in 88% of plots, averaging 33% cover where present. Other common shrub genera included Rosa spp., Arctostaphylos spp., Symphoricarpos spp., Salix spp., and Ribes spp.

a Observed seedling densities in four study fires. Seedling densities varied substantially between plots and among fires. Blue triangles represent mean seedling densities among plots for each fire. Three plots in the Roberts Creek fire with seedling densities > 40,000 seedlings ha−1 are not shown to improve figure interpretability. Horizontal lines represent the median, with first and third quantiles flanking either side. Median seedling densities calculated among plots for each fire were > 400 seedlings ha−1 in all four fires. b The abundance of ponderosa pine, Douglas-fir, and grand fir seedlings was not negatively associated with shrub cover. Shrub cover values could sum to greater than 100% when multiple species overlapped in space within a plot (e.g., 90% Ceanothus velutinus cover, 20% Arctostaphylos patula cover). (Color figure online)
The availability of surviving seed sources varied substantially across stand-replacement patches in our study fires. The median distance to nearest seed source recorded in the field was 27 m; distances ranged from 6 m (just outside the 5.64-m plot radius) to 600 m (estimated, beyond the range of our laser rangefinder). According to our field data, median distances to seed source for ponderosa pine, Douglas-fir, and grand fir were 51 m, 44 m, and 72 m, respectively. Mean refugia density across all plots was 275, with a range of 0 (no refugia cells within the moving window) to 750 (approximately 75% of cells classified as refugia). Because distance to seed source and refugia density both represent seed source availability we would expect them to be somewhat related, but we observed substantial variability in refugia density values along the distance to seed source gradient within the window used to calculate refugia density (Fig. 6a). Although our study was not explicitly designed to representatively sample all the area that experienced stand-replacement fire effects, a comparison of field-measured distance to seed source and distance to seed source inferred from our NAIP maps demonstrated that our field sampling reasonably approximated the distribution of distances to refugial seed source across our study area (Supplemental Fig. 5).

a Fire refugia distance-weighted density values (fire refugia density, x-axis) varied substantially along a gradient of distance to nearest seed source (y-axis) for plot locations within 150 m of a refugial seed source. Each point represents a field plot for one of the three species-specific models (ponderosa pine, Douglas-fir, grand fir). Fire refugia density varies between locations with relatively little refugial seed source within the 150 m window used to calculate refugia density (b), to locations with substantially more refugial seed source (c), even when distance to seed source is held constant. Distance to seed source measurements made in the field were species-specific, while the refugia density metric is a generic representation of potential seed source which does not differentiate between species. The spatial patterns of refugial seed source in our study area are complex (d), including sparse, isolated patches of surviving trees (foreground, left), variable density patches (middle ground, right), and large, dense patches that form the periphery of stand-replacement patches (background). Our refugia density metric allowed for us to account for some of this variability in fire refugia spatial pattern, providing an important complement to our species-specific measurements of distance to seed source
We identified mean distance to nearest seed source and refugia density for inclusion in our final statistical models of seedling recruitment based on our initial modeling of seed source metrics (Table 3). There was some evidence that mean distance to seed source explained more variability in the data than minimum and median distance in models of Douglas-fir regeneration, and we used this as the basis for choosing mean distance to seed source (hereafter, distance to seed source) as our point-wise metric of refugial seed source pattern. Combined models (distance to seed source + refugia density) represented substantial improvement over null models fit with only an intercept term for our three focal species and all species combined (Table 1). We considered both distance to seed source and refugia density important, complementary sources of information about surviving seed source pattern (Fig. 6), and we chose to include both in all of our final models.
Final models demonstrated spatial variability in post-fire regeneration as a function of both site characteristics and surviving seed source (Table 4). Distance to seed source was negatively associated with seedling regeneration in all models, but was not significant (P > 0.05) in the case of grand fir (Fig. 7). Refugia density was positively associated with post-fire conifer regeneration in all of our models, after accounting for distance to seed source, fire severity, elevation, CMD, heat load, pre-fire basal area, and shrub cover. Predicted seedling densities increased between 77% and 208% when refugia density increased from low (25th percentile observed) to high (75th percentile observed) values (Table 5). Conversely, predicted seedling densities decreased between 24% and 43% when distance to seed source increased from low (25th percentile observed) to high (75th percentile observed) values. Negative binomial models were ultimately chosen because they adequately accounted for the over-dispersion in the data and because they performed as well as or better than the more complex, zero-inflated negative binomial models, according to AIC. The standard deviation of the random effect term was 0, 0.57, 0.93, 0.92 for our ponderosa pine, Douglas-fir, grand fir, and all-species models, respectively.

Predicted seedling densities along distance to seed source and fire refugia density gradients. Predicted seedling densities decreased with distance to seed source and increased with refugia density. These added variable plots represent the effect size of refugial seed source parameters, with all other variables held constant at median values. Dotted lines represent 95% confidence intervals. Note y-axis scales vary between each pair of plots due to significant variation in predicted seedling densities for different species. Stars denote statistically significant relationships between seed source metrics and post-fire seedling abundance
While surviving seed source was an important driver of post-fire regeneration in all of our models, the influence of site characteristics varied among species. Douglas-fir regeneration was negatively associated with heat load and positively associated with burn severity (dNBR), despite our placement of all sample plots at locations with 100% canopy mortality. Shrub cover was positively related with ponderosa pine regeneration, but was not associated with Douglas-fir, grand fir, or all species combined. Polynomial terms included for elevation and CMD provided evidence of both convex (elevation) and concave (CMD) relationships with post-fire seedling abundance. With the exception of a positive relationship between ponderosa pine regeneration and CMD, elevation and CMD were not associated with seedling densities in any models fit without squared terms for these parameters (Supplemental Table 1). There was no evidence of an interaction between CMD and shrub cover for any species, and we did not include the interaction in final models.
Ponderosa pine regeneration was ongoing across all fires based on estimated establishment dates for 327 seedlings (Fig. 8). We underestimated seedling age by an average of two years, based on a comparison with our 21 seedling cross-sections, so we subtracted 2 years from the whorl-count establishment dates prior to analysis. The peak period of seedling establishment varied among fires, occurring as early as five years post-fire (Burnt Cabin and Roberts Creek) and as late as 11 years post-fire (747). Younger seedlings were more likely to be overtopped by shrubs than older seedlings. Overall, ponderosa pine seedling establishment dates showed no association with variability in annual climatic moisture deficit, although there was an increase in establishment in Burnt Cabin that coincided with a decline in moisture deficit in 2010.

Estimated year of establishment of ponderosa pine seedlings and annual climatic moisture deficit. There does not appear to be a strong association between seedling establishment and climate moisture deficit (CMD). Seedlings overtopped by shrubs are represented with black, and seedlings that have emerged above the shrub canopy or were not growing in close proximity to shrubs are represented with grey. Plots of seedlings established each year and mean annual CMD are paired for each fire. CMD was averaged across plots for each fire to characterize landscape-scale climatological trends, rather than the actual moisture conditions experienced by reestablishing seedlings at the plot scale. Two years were subtracted from seedling establishment dates to correct for the bias of field-based estimates based on whorl-counts (see Methods). Red dotted lines represent the 30-year average annual climate moisture deficit for each fire area. Triangles denote the year of fire
Discussion
Dry mixed-conifer forests in the Blue Mountains of Oregon show evidence of resilience to high-severity fire effects 12-17 years post-fire. In contrast to slow or absent post-fire forest regeneration reported in dry forests in other regions (Collins and Roller 2013; Chambers et al. 2016; Welch et al. 2016), conifer seedlings were widespread and relatively abundant in much of our study area, indicating that these forests have capacity to recover following stand-replacement fire, at least across the range of stand-replacement patch sizes we sampled. Consistent with previous studies (e.g., Donato et al. 2009; Chambers et al. 2016; Kemp et al. 2016), we found that proximity to surviving seed source is a key driver of post-fire conifer regeneration. Our high-resolution mapped surfaces of fire refugia allowed us to quantify the neighborhood density of fire refugia, providing novel insights into the influence of fire refugia pattern as well as the additive effect of multiple seed sources contributing to forest regeneration. High-severity fire in our study area resulted in substantial losses of mature forest structure that will take decades to be replaced. However, unlike other dry forest systems that appear to be transitioning to alternative, stable states following high-severity fire (Savage and Mast 2005; Coop et al. 2016; Tepley et al. 2017), we anticipate that in the absence of subsequent disturbance (e.g., reburn), dry mixed-conifer forest in the Blue Mountains within the range of conditions we sampled will recover following stand-replacement fire.
Our study contributes to a growing body of literature demonstrating the variability in post-fire forest regeneration among different dry forest ecosystems and the difficulty associated with making broad generalizations about the resilience or vulnerability of dry forests to high-severity fire effects (Collins and Roller 2013). While differences in study design and time-since-fire make it challenging to directly compare our findings with those from previous regeneration studies, our results are consistent with those reported from dry forest ecosystems in the Klamath Siskiyou (Shatford et al. 2007; Donato et al. 2009), southern Cascades (Crotteau et al. 2013), and Northern Rockies ecoregions (Kemp et al. 2016) of the western United States. However, our results contrast with post-fire landscapes in parts of California, the Southern Rockies, and the Southwest. In the northern Sierra Nevada Mountains, seedlings were present in less than half of sample plots 2–11 years post-fire (Collins and Roller 2013), and in the Colorado Front Range, only 25% of plots contained seedlings 11–18 years post-fire (Chambers et al. 2016). Where seedlings were present following high-severity fire in the southern Rockies and Arizona, seedling densities were an order of magnitude lower on average than the seedling densities we observed in the Blue Mountains (Chambers et al. 2016; Owen et al. 2017). Our study, the first of its kind in the range of environmental conditions supported by the Blue Mountains of Oregon, highlights the importance of studying post-fire forest recovery across a wide range of dry forest ecosystems, as these ecosystems appear to vary substantially in their capacity to recover following high-severity fire.
We have provided evidence that the density of seed sources that survived fire in refugia is an important control on the process of forest regeneration following high-severity fire. The positive association we found between post-fire seedling abundance and distance-weighted refugia density demonstrates that there is an additive effect of multiple seed sources contributing to a site’s capacity to reestablish forest following fire: regeneration varies depending on the density of surviving seed source, even when distance to seed source is held constant. Our results may support the idea that the relatively high abundance of seeds produced by large, dense refugial patches increases the probability of longer-distance dispersal events (Shatford et al. 2007), while small, isolated refugia may influence regeneration in surrounding high-severity burned areas at comparatively smaller spatial scales because they generate fewer propagules.
While our refugia density metric does not explicitly examine patch characteristics, quantifying aspects of landscape context such as post-fire seed sources is an important way to evaluate the influence of spatial pattern on ecosystem processes like dispersal and regeneration (Turner 2005). Recognition of the importance of landscape context has been described as the essence of landscape ecology (Wiens 2011), particularly in relation to post-disturbance succession (Turner et al. 2015) Our two-dimensional, area-based refugia density metric enabled us to quantify a key landscape pattern attribute (refugial seed source) that strongly controls the process of post-fire forest regeneration. Landscape context can be defined in terms of both distance to, and area of, neighboring patches (Cushman et al. 2010), and we believe that distance to seed source and refugia density constitute complementary measures of seed source availability. However, unlike distance to seed source metrics that tacitly assume that all patches of surviving seed source are equal, refugia density is an area metric that accounts for a broad gradient of seed source patches ranging from isolated groups composed of a small number of surviving individual trees surrounded by stand-replacement fire effects, to large, contiguous patches of surviving forest that form the borders of high-severity patches (Fig. 6d).
Contemporary fires are capable of creating much larger stand-replacement patches than those within our study fires (Reilly et al. 2017), and the pace of forest recovery we identified here may not be representative of substantially larger stand-replacement patches in the Blue Mountains, should they occur. However, widespread regeneration in our study area indicates that adequate seed sources are present across much of the high-severity burned area in our study fires. Our field sampling effort was highest in the most recent fire (Table 2, Burnt Cabin, 2005). Because this area has had the least amount of time to regenerate forest, and because the percentage of plots that did not contain seedlings (29%) was higher than the dataset overall (17%), our results may be biased towards underestimating post-fire seedling regeneration. Overall, seedlings were absent in 23 of our study plots, but these plots were relatively close to a surviving seed source (median distance to nearest seed source = 27 m), indicating that seed availability was not the key constraint in most of these locations. Additionally, according to our maps of surviving forest across entire fire footprints, the area within stand-replacement patches far (> 150 m) from a refugial seed source was relatively small (9%) across our study fires (Supplemental Fig. 5).
Although our model results indicate that the amount and pattern of refugial seed sources are the dominant controls on post-fire regeneration in this ecosystem, other ecological site characteristics influenced post-fire seedling abundance in some cases. Fire severity was positively associated with Douglas-fir regeneration but did not appear to influence ponderosa pine, grand fir, or the group of all species combined, potentially because we restricted our sampling to stand-replacement patches. Stand-replacement fire in areas with dense pre-fire canopy will result in higher dNBR values compared to stand-replacement fire in areas with sparse pre-fire canopy (Miller and Thode 2007), and the positive association between Douglas-fir seedling abundance and fire severity may reflect site productivity (higher pre-fire biomass), rather than the direct influence of fire effects. Douglas-fir regeneration was also positively associated with pre-fire basal area, and we interpret these results as evidence that Douglas-fir regenerates at higher densities in more productive locations, as described by Kemp et al. (2016). Consistent with that study, we found that Douglas-fir was also the only species strongly influenced by heat load. Douglas-fir is more sensitive than ponderosa pine to heat and drought stress (Rother et al. 2015), and our results suggest that in the Blue Mountains its capacity to regenerate is limited more than other conifer species in landscape positions with high solar insolation. Additionally, our study design was intended to sample across broad environmental gradients, including less productive, low-elevation forests, but we did not specifically target the interface between forest and non-forest pre-fire vegetation. Variability in elevation and heat load may be more important drivers of post-fire forest recovery trajectories at lower elevations (Boag 2018) along this ecotone boundary than across the broad range of sites we sampled here.
We expected seedling abundance to increase with elevation and cooler, wetter climatic conditions (i.e., lower CMD), as observed by prior studies (Chappell and Agee 1996; Dodson and Root 2013; Chambers et al. 2016). Model results did not provide evidence of a linear association between post-fire seedling regeneration and either CMD or elevation, with the exception of a positive association between CMD and ponderosa pine. However, we did find evidence of non-linear relationships between seedling regeneration and both elevation and CMD. Species often respond to environmental gradients non-linearly, especially across broad environmental gradients like the ones we sampled here (Whittaker 1960), and in all four models, polynomial terms indicated seedling abundance may have a convex response along the elevation gradient. Seedling densities were highest at intermediate elevations, locations where regenerating seedlings may be buffered from both hot and cold temperature extremes, as found by Collins and Roller (2013) and others (Petrie et al. 2016). In contrast, results for CMD showed the opposite trend; seedling densities were lowest at intermediate CMD values in all four models (Supplemental Fig. 3). However, it may be the case that the resolution of available climate data was too coarse (800 × 800 m) to provide meaningful, interpretable results for a phenomenon like post-fire tree regeneration that is sensitive to factors operating at much finer spatial scales, and these data may be better suited to identifying general climatic trends through time, rather than variability across the range of data we sampled here.
Widespread shrub cover in stand-replacement patches in our study area does not appear to limit post-fire conifer regeneration. We did not find evidence of a negative relationship between post-fire seedling abundance and shrub cover, suggesting that competition between regenerating seedlings and other woody species does not substantially suppress reforestation in our study area. In addition, the positive association between ponderosa pine seedling abundance and shrub cover may indicate a facilitative relationship. Numerous seedlings in our study area appear to have recently emerged above the shrub canopy, suggesting that these sites have not been permanently “captured” by woody understory species (Savage and Mast 2005; Barton and Poulos 2018), and that a robust post-fire shrub response is compatible with the natural regeneration of stand-replacement patches in the Blue Mountains.
Similar initial conversions to shrublands after fire have been observed in the Klamath-Siskiyou and Sierra Nevada mountains, and both positive (Shatford et al. 2007; Collins and Roller 2013), and negative (Meigs et al. 2009; Welch et al. 2016; Tepley et al. 2017) associations between shrub cover and seedling abundance have been reported. Ceanothus, the dominant shrub genus in our study area, requires scarification for its seeds to germinate and responds vigorously after high-severity fire (Conrad et al. 1985). Ceanothus has been shown to both suppress conifer establishment and growth (Zavitkovski et al. 1969) as well as facilitate seedling survival by ameliorating unfavorable post-fire environmental conditions (Wahlenberg 1930). It is possible that both of these mechanisms are at work in our system, but that the net effect of shrub cover on seedling regeneration is not substantial for grand fir, Douglas-fir, or all species combined. Competition with herbaceous vegetation can also limit seedling regeneration in dry forest ecosystems (White 1985), but herbaceous cover in stand-replacement patches in our study area was low in comparison to that of woody understory species, and we chose not test for an effect of herbaceous cover on seedling abundance. However, it is possible that the continued expansion of invasive annual grasses like Ventenata dubia and Bromus tectorum into Blue Mountain dry forests could limit post-fire tree regeneration in the future (Downing 2018).
Ponderosa pine seedling establishment is ongoing in stand-replacement patches 12–17 years post-fire, even in areas dominated by shrubs. These results serve as an important reminder that reforestation of stand-replacement patches in dry forests can be a slow process that continues for decades (Haire and McGarigal 2010), and initial post-fire dominance by non-tree vegetation does not necessarily indicate an enduring vegetation type shift. The temporal patterns of regeneration we identified are consistent with previous studies conducted in ponderosa pine forests in the Southwest (Savage and Mast 2005; Haire and McGarigal 2010), and in dry mixed-conifer forests in the Klamath-Siskiyou region (Shatford et al. 2007), where a period of slow initial establishment was followed by a peak in regeneration. Other studies have reported a distinct regeneration pulse in the years immediately following fire (Donato et al. 2009; Rother and Veblen 2017; Tepley et al. 2017), but our data indicate that ponderosa pine seedlings either did not establish rapidly post-fire, or these early establishers did not survive to be recorded.
Our qualitative assessment of ponderosa pine seedling establishment dates and climatic moisture deficit did not identify a strong relationship, suggesting that ponderosa pine regeneration in our study area is constrained by factors other than landscape-scale climatic variability. In contrast, other studies have found that post-fire conifer establishment is concentrated in years with above-average growing season moisture (Rother and Veblen 2017), and decreased regeneration during dry periods suggests that climate warming may contribute to conversion to non-forest states following high-severity fire (Stevens-Rumann et al. 2018). The lack of an apparent relationship in our study may be a result of relatively low climatic variability during the post-fire time period; conditions may not have been hot and dry enough in the years following fire to significantly discourage ponderosa pine regeneration. In addition, ponderosa pine seed production is highly episodic, with good cone crops occurring only once every 4 to 6 years (Shepperd et al. 2006), and interannual variability in seed availability may obscure any climate signal present in our establishment data. Additionally, we recognize that the whorl counts we used to estimate establishment dates are biased towards underestimating true tree age (Hankin et al. 2018), and the lack of precision in these estimates limit our ability to detect a climate signal.
Although forest is reestablishing following high-severity fire in our study area, it is not clear the degree to which regenerating seedlings represent pre-fire forest composition, or, importantly, historical forest composition. As a result of fire exclusion, infilling by shade tolerants like grand fir is widespread across dry mixed-conifer forests in the Pacific Northwest (Merschel et al. 2014; Johnston 2017), and it is likely that pre-fire forest composition in our study area was departed from historical reference conditions due to the decades-long absence of frequent fire. Additionally, conditions may favor the establishment of more shade-tolerant tree species when shrubs like Ceanothus dominate high-severity burned sites (Zavitkovski and Newton 1968; Crotteau et al. 2013). We were unable to compare pre-fire forest composition to the composition of regenerating seedlings because bark loss and decay made it impossible to determine the species of some fire-killed trees, and future research could seek to determine if high-severity fire effects are reinforcing changes in dry mixed-conifer forest composition initiated by post-settlement fire exclusion.
This study provides evidence that the spatial pattern of forest that survived fire in refugia is a key driver of the process of post-fire forest regeneration. We acknowledge that this relationship between refugial seed source and seedling regeneration is just one facet of far more complex, iterative, pattern-process relationships (Swanson 1981). For example, the pattern of post-fire regeneration is itself the result of fire as an ecosystem process, which was influenced in turn by spatial patterns of topography, pre-fire fuels, and their feedbacks with the patterns of prior fires. As fire activity continues to increase and interest grows in the ecological implications of repeat disturbances like reburn (Prichard et al. 2017), additional research will be needed to better understand how pattern-process interactions reinforce one another over multiple disturbance events and the ways in which pre-fire patterns influence processes like post-fire forest regeneration.
Our ability to quantify refugial seed source pattern had a number of limitations. Seed sources may have been closer than measured when topography or individuals of other species obscured our view. Unlike distance to seed source, which was measured for each species, the methods we used to quantify refugia density did not identify species, and our capacity to model regeneration would almost certainly be stronger if refugia density reflected the tree composition of fire refugia. The techniques to achieve this level of discrimination among species do exist (Kuzmin et al. 2016), but their application was beyond the scope of this study and represents an opportunity for future refinement. In addition, our distance-weighted refugia density metric is useful because it incorporates both nearby and relatively distant seed sources, but the metric is limited by the fact that the user selects the window size. We chose what we believe to be an appropriate window size based on observed seed dispersal distances from numerous post-fire regeneration studies, including the work of Haire and McGarigal (2010), which reported that dispersal kernels scaled at 60–100 m were the strongest predictors of regeneration in ponderosa pine forests. However, their results may not translate to mixed-conifer forests, including tree species like Douglas-fir and grand fir that have lighter seeds than ponderosa pine (Burns and Honkala 1990). We observed many seedlings farther than 150 m from refugial seed sources, and future studies could investigate fire refugia spatial pattern at broader spatial scales (i.e. window sizes that account for seed sources beyond 150 m). Lastly, our models did not take into account the influence of prevailing winds on seed dispersal. In highly disturbed, topographically complex landscapes like those sampled here, integrating the strength and direction of prevailing winds constitutes a substantial analytical challenge. However, given the importance of wind dispersal for dry mixed-conifer tree species, future research could explore the influence of the interaction between prevailing winds and surviving seed sources on post-fire forest regeneration.
Management applications and conclusions
We sampled primarily in designated roadless and wilderness areas where many management interventions are either not feasible or not permitted. However, the strong influence of refugial seed source pattern on post-fire regeneration in the Blue Mountains may inform potential management strategies before, during, and after fire in more actively managed landscapes. Fuel treatments prior to fire that increase forests’ resistance to high-severity fire (Agee and Skinner 2005) could help conserve fire refugia and promote natural forest regeneration. Additionally, our findings suggest that actions that may reduce or eliminate fire refugia from the post-fire landscape both during fire events, like burnout operations, and after, like salvage logging, could interfere with post-fire forest regeneration. In areas where replanting after fire is a viable and desirable option, managers could target locations where severe fire effects eliminated refugial seed sources, thus inhibiting natural regeneration.
This study provides evidence that fire refugia confer resilience to dry mixed-conifer forests following high-severity fire. As the areas least changed by a fire event, fire refugia promote the surrounding forest matrix’s capacity to absorb and recover from significant fire-induced change. In dry mixed-conifer forests of the Blue Mountains, we found that fire refugia increased the ability of long fire-excluded landscapes to regenerate after high-severity fire. Aside from their role as propagule sources, fire refugia may serve additional ecosystem functions (Meddens et al. 2018), such as facilitating the survival of fire-sensitive plant species (Wood et al. 2011), providing faunal habitat otherwise absent in burned landscapes (Robinson et al. 2013), and buffering post-fire microclimates (Delong and Kessler 2000). Additional research will be needed to broaden our understanding of the role of fire refugia in dry mixed-conifer forest ecosystems, and to explore the generality of findings from these systems across a range of other fire-prone ecosystems globally.
References
Adie H, Kotze DJ, Lawes MJ (2017) Small fire refugia in the grassy matrix and the persistence of Afrotemperate forest in the Drakensberg mountains. Sci Rep 7:1–10
Agee JK, Skinner CN (2005) Basic principles of forest fuel reduction treatments. For Ecol Manage 211:83–96
Allen CD, Savage M, Falk DA, Suckling KF, Thomas W, Schulke T, Stacey PB, Morgan P, Hoffman M, Jon T (2002) Ecological restoration of southwestern ponderosa pine ecosystems: a broad perspective. Ecol Appl 12:1418–1433
Barton AM, Poulos HM (2018) Pine vs. oaks revisited: conversion of Madrean pine-oak forest to oak shrubland after high-severity wildfire in the Sky Islands of Arizona. For Ecol Manage 414:28–40
Bates D, Mächler M, Bolker B, Walker S (2014) Fitting linear mixed-effects models using lme4. J Stat Softw 67:48
Boag AE (2018) Climate change and wildfire: implications for forest management in the Blue Mountains of eastern Oregon. University of Colorado, Boulder
Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Mächler M, Bolker BM (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9:378–400
Burns RM (1983) Silvicultural systems for the major forest types of the United States. Agricultural Handbook 445. USDA Forest Service, Washington, D.C., USA
Burns RM, Honkala BH (1990) Silvics of North America. Agricultural Handbook 654. USDA Forest Service, Washington, D.C., USA
Camp A, Oliver C, Hessburg P, Everett R (1997) Predicting late-successional fire refugia pre-dating European settlement in the Wenatchee mountains. For Ecol Manage 95:63–77
Chambers ME, Fornwalt PJ, Malone SL, Battaglia MA (2016) Patterns of conifer regeneration following high severity wildfire in ponderosa pine – dominated forests of the Colorado Front Range. For Ecol Manage 378:57–67
Chappell CB, Agee JK (1996) Fire severity and tree seedling establishment in Abies magnifica forests, Southern Cascades, Oregon. Ecol Appl 6:628–640
Clark JS (1998) Why trees migrate so fast: confronting theory with dispersal biology and the paleorecord. Am Nat 152:204–224
Collins BM, Roller GB (2013) Early forest dynamics in stand-replacing fire patches in the northern Sierra Nevada, California, USA. Landscape Ecol 28:1801–1813
Conrad SG, Jaramillo AE, Kermit Cromack J, Rose S (1985) The role of the genus Ceanothus in western forest ecosystems. USDA Forest Service General Technical Report, PNW-GTR-182. Pacific Northwest Research Station, Portland, OR, USA
Coop JD, Parks SA, Mcclernan SR, Holsinger LM (2016) Influences of prior wildfires on vegetation response to subsequent fire in a reburned southwestern landscape. Ecol Appl 26:346–354
Crotteau JS, Varner JM, Ritchie MW (2013) Post-fire regeneration across a fire severity gradient in the southern Cascades. For Ecol Manage 287:103–112
Cushman SA, Evans JS, McGarigal K (2010) Spatial complexity, informatics, and wildlife conservation. In: Cushman SA, Huettmann F (eds) Spatial complexity, informatics, and wildlife conservation. Springer, New York
Delong SC, Kessler WB (2000) Ecological characteristics of mature forest remnants left by wildfire. For Ecol Manage 131:93–106
Dodson EK, Root HT (2013) Conifer regeneration following stand-replacing wildfire varies along an elevation gradient in a ponderosa pine forest, Oregon, USA. For Ecol Manage 302:163–170
Donato DC, Fontaine JB, Campbell JL, Robinson WD, Kauffman JB, Law BE (2009) Conifer regeneration in stand-replacement portions of a large mixed-severity wildfire in the Klamath-Siskiyou Mountains. Can J For Res 39:823–838
Donato DC, Harvey BJ, Turner MG (2016) Regeneration of montane forests 24 years afer the 1988 Yellowstone fires: a fire-catalyzed shif in lower treelines? Ecosphere 7:1–16
Downing WM (2018) Fire refugia function and composition in dry mixed-conifer forests of Oregon’s Blue Mountains. Oregon State University
Greene DF, Johnson EA (2000) Tree recruitment from burn edges. Can J For Res 30:1264–1274
Hagmann RK, Franklin JF, Johnson KN (2013) Historical structure and composition of ponderosa pine and mixed-conifer forests in south-central Oregon. For Ecol Manage 304:492–504
Haire SL, McGarigal K (2010) Effects of landscape patterns of fire severity on regenerating ponderosa pine forests (Pinus ponderosa) in New Mexico and Arizona, USA. Landscape Ecol 25:1055–1069
Hall FC (1976) Fire and vegetaion in the Blue Mountains–Implications for land managers. In: Page Proceedings of the annual Tall Timbers fire ecology conference. Tall Timbers Research Station, Tallahassee, Florida, pp. 155–170
Hankin LE, Higuera PE, Davis KT, Dobrowski SZ (2018) Accuracy of node and bud-scar counts for aging two dominant conifers in western North America. For Ecol Manag 427:365–371
Harvey BJ, Donato DC, Turner MG (2016) High and dry: post-fire tree seedling establishment in subalpine forests decreases with post-fire drought and large stand-replacing burn patches. Glob Ecol Biogeogr 25:655–669
Hessburg PF, Agee JK (2003) An environmental narrative of Inland Northwest United States forests, 1800-2000. For Ecol Manage 178:23–59
Hessburg PF, Agee JK, Franklin JF (2005) Dry forests and wildland fires of the inland Northwest USA: contrasting the landscape ecology of the pre-settlement and modern eras. For Ecol Manage 211:117–139
Heyerdahl EK, Brubaker LB, Agee JK (2001) Spatial controls of historical fire regimes: A multiscale example from the interior west, USA. Ecology 82:660–678
Johnson CG, RR Clausnitzer (1992) Plant associations of the Blue and Ochoco Mountains. R6-ERW-T-036092. Pacific Northwest Research Station, Portland, OR, USA
Johnston JD (2017) Forest succession along a productivity gradient following fire exclusion. For Ecol Manage 392:45–57
Johnston JD, Bailey JD, Dunn CJ (2016) Influence of fire disturbance and biophysical heterogeneity on pre-settlement ponderosa pine and mixed conifer forests. Ecosphere 7:1–19
Kemp KB, Higuera PE, Morgan P (2016) Fire legacies impact conifer regeneration across environmental gradients in the U.S. northern Rockies. Landscape Ecol 31:619–636
Key CH, Benson NC (2006) Landscape assessment: ground measure of severity, the composite burn index. In: Lutes DC (ed) FIREMON: fire effects monitoring and inventory system. USDA Forest Service General Technical Report 164-CD. Rocky Mountain Research Station, Fort Collins, Colorado, USA, pp LA8–LA15.
Kuzmin A, Korhonen L, Manninen T, Maltamo M (2016) Automatic segment-level tree species recognition using high resolution aerial winter imagery. Eur J Remote Sens 49:239–259
Landesmann JB, Morales JM (2018) The importance of fire refugia in the recolonization of a fire-sensitive conifer in northern Patagonia. Plant Ecol 219:455–466
McCune B, Keon D (2002) Equations for potential annual direct incident radiation and heat load. J Veg Sci 13:603–606
Meddens AJ, Kolden CA, Lutz JA, Smith AM, Cansler CA, Abatzoglou JT, Meigs GW, Downing WM, Krawchuk MA (2018) Fire refugia: what are they, and why do they matter for global change? BioScience 68(12):944–954
Meigs GW, Donato DC, Campbell JL, Martin JG, Law BE (2009) Forest fire impacts on carbon uptake, storage, and emission: the role of burn severity in the Eastern Cascades, Oregon. Ecosystems 12:1246–1267
Merschel AG, Spies TA, Heyerdahl EK (2014) Mixed-conifer forests of central Oregon: Effects of logging and fire exclusion vary with environment. Ecol Appl 24:1670–1688
Miller JD, Safford H (2012) Trends in wildfire severity: 1984 to 2010 in the Sierra Nevada, Modoc Plateau, and southern Cascades, California, USA. Fire Ecol 8:41–57
Miller JD, Thode AE (2007) Quantifying burn severity in a heterogeneous landscape with a relative version of the delta Normalized Burn Ratio (dNBR). Remote Sens Environ 109:66–80
Ott TM, Strand EK, Anderson CL (2015) Niche divergence of Abies grandis-Abies concolor hybrids. Plant Ecol 216:479–490
Ouarmim S, Paradis L, Asselin H, Bergeron Y, Ali A, Hély C (2016) Burning potential of fire refuges in the boreal mixedwood forest. Forests 7:246
Owen SM, Sieg CH, Sánchez Meador AJ, Fulé PZ, Iniguez JM, Baggett LS, Fornwalt PJ, Battaglia MA (2017) Spatial patterns of ponderosa pine regeneration in high-severity burn patches. For Ecol Manage 405:134–149
Petrie MD, Wildeman AM, Bradford JB, Hubbard RM, Lauenroth WK (2016) A review of precipitation and temperature control on seedling emergence and establishment for ponderosa and lodgepole pine forest regeneration. For Ecol Manage 361:328–338
Powell D (1994) Effects of the 1980 s western spruce budworm outbreak on the Malheur National Forest in northeastern Oregon. USDA Forest Service Technical Report R6-FI&D-TP-12-94. Pacific Northwest Region Forest Insects and Disease Group, Portland, OR, USA
Prichard SJ, Stevens-Rumann CS, Hessburg PF (2017) Tamm review: shifting global fire regimes: lessons from reburns and research needs. For Ecol Manage 396:217–233
Reilly MJ, Dunn CJ, Meigs GW, Spies TA, Kennedy RE, Bailey JD, Briggs K (2017) Contemporary patterns of fire extent and severity in forests of the Pacific Northwest, USA (1985–2010). Ecosphere 8:e01695
Robinson NM, Leonard SWJ, Ritchie EG, Bassett M, Chia EK, Buckingham S, Gibb H, Bennett AF, Clarke MF (2013) Refuges for fauna in fire-prone landscapes: their ecological function and importance. J Appl Ecol 50:1321
Rother MT, Veblen TT (2017) Climate drives episodic conifer establishment after fire in dry ponderosa pine forests of the Colorado Front Range, USA. Forests 8:1–14
Rother MT, Veblen TT, Furman LG (2015) A field experiment informs expected patterns of conifer regeneration after disturbance under changing climate conditions. Can J For Res 45:1607–1616
Safford HD (2013) Natural Range of Variation (NRV) for yellow pine and mixed conifer forests in the bioregional assessment area, including the Sierra Nevada, southern Cascades, and Modoc and Inyo National Forests. Unpublished report. USDA Forest Service, Pacific Southwest Region, Vallejo, CA
Savage M, Mast JN (2005) How resilient are southwestern ponderosa pine forests after crown fires? Can J For Res 35:967–977
Shatford JPA, Hibbs DE, Puettmann KJ (2007) Conifer regeneration after forest fire in the Klamath-Siskiyous: how much, how soon? J For 105:139–146
Shepperd WD, Edminster CB, Mata SA (2006) Long-term seedfall, establishment, survival, and growth of natural and planted ponderosa pine in the Colorado Front Range. West J Appl For 21:19–26
Stephens SL, Agee JK, Fule PZ, North MP, Romme WH, Swetnam TW, Turner MG (2013) Managing forests and fire in a changing climate. Science 342:41–42
Stevens-Rumann CS, Kemp KB, Higuera PE, Harvey BJ, Rother MT, Donato DC, Morgan P, Veblen TT (2018) Evidence for declining forest resilience to wildfires under climate change. Ecol Lett 21:243–252
Stine P, Hessburg P, Spies T, Kramer M, Fettig CJ, Hansen A, Lehmkuhl J, O’Hara K, Polivka K, Singleton P, Charnley S, Merschel A, White R (2014) The ecology and management of moist mixed-conifer forests in eastern Oregon and Washington: a synthesis of the relevant biophysical science and implications for future land management. USDA Forest Service, General Technical Report, PNW-GTR-897. Pacific Northwest Research Station, Portland, Oregon, USA
Swanson FJ (1981) Fire and geomorphic processes. In: Bonnicksen TM, Christensen NL, Lotan JE (eds) Mooney HA. Fire regimes and ecosystem properties, USDA General Technical Report WO-26. USDA Forest Service, pp 401–444
Tepley AJ, Thompson JR, Epstein HE, Anderson-Teixeira KJ (2017) Vulnerability to forest loss through altered postfire recovery dynamics in a warming climate in the Klamath Mountains. Glob Change Biol 23:4117–4132
Turner MG (1989) Landscape ecology: the effect of pattern on process. Annu Rev Ecol Syst 20:171–197
Turner MG (2005) Landscape ecology in North America: past, present, and future. Ecology 86:1967–1974
Turner MG, Gardner RH, O’neill RV, O’Neill RV (2015) Landscape ecology in theory and practice. Springer, New York
Turner MG, Romme WH, Gardner RH, Hargrove WW (1997) Effects of fire size and pattern on early succession in Yellowstone National Park. Ecol Monogr 67:411–433
Vander Wall SB (2003) Effects of seed size of wind-dispersed pines (Pinus) on secondary seed dispersal and the caching behavior of rodents. Oikos 100:25–34
Wahlenberg WG (1930) Effect of ceanothus brush on western yellow pine plantations in the northern Rocky Mountains. J Agric Res 41:601–612
Wang T, Hamann A, Spittlehouse D, Carroll C (2016) Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS ONE 11:1–17
Wang T, Hamann A, Spittlehouse DL, Murdock TQ (2012) ClimateWNA-high-resolution spatial climate data for western North America. J Appl Meteorol Climatol 51:16–29
Welch KR, Safford HD, Young TP (2016) Predicting conifer establishment post wildfire in mixed conifer forests of the North American Mediterranean-climate zone. Ecosphere 7:e01609
White AS (1985) Presettlement regeneration patterns in a Southwestern Ponderosa Pine Stand. Ecology 66:589–594
Whittaker RH (1960) Vegetation of the Siskiyou Mountains, Oregon and California. Ecol Monogr 30:279–338
Wiens JA (2011) Central concepts and issues of landscape ecology. In: Gutzwiller KJ (ed) Applying landscape ecology in biological conservation. Springer, Berlin, pp 3–21
Wood SW, Murphy BP, Bowman DMJS (2011) Firescape ecology: how topography determines the contrasting distribution of fire and rain forest in the south-west of the Tasmanian Wilderness World Heritage Area. J Biogeogr 38:1807–1820
Zavitkovski J, Newton M (1968) Ecological importance of snowbrush ceanothus velutinus in the Oregon Cascades. Ecology 49:1134–1145
Zavitkovski J, Newton M, El-Hassan B (1969) Effects of snowbrush on growth of some conifers. J For 67:242–246
Zhao J, Maguire DA, Mainwaring DB, Kanaskie A (2012) Climatic influences on needle cohort survival mediated by Swiss needle cast in coastal Douglas-fir. Trees Struct Funct 26:1361–1371
Acknowledgements
This research was funded by the National Fire Plan through agreements between the Aldo Leopold Wilderness Research Institute and Oregon State University (16-JV-11221639-101) and Western Colorado University (16-JV-11221639-107). Access to field sites was facilitated by the Ochoco, Umatilla, and Malheur National Forests. We thank Jean McCalmont, Julie VanSandt, and Claire Tortorelli for assistance with data collection, and Marc-André Parisien for comments on earlier drafts. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Downing, W.M., Krawchuk, M.A., Meigs, G.W. et al. Influence of fire refugia spatial pattern on post-fire forest recovery in Oregon’s Blue Mountains. Landscape Ecol 34, 771–792 (2019). https://doi.org/10.1007/s10980-019-00802-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-019-00802-1