
Abstract
There is considerable concern over the occurrence of stand-replacing fire in forest types historically associated with low- to moderate-severity fire. The concern is largely over whether contemporary levels of stand-replacing fire are outside the historical range of variability, and what natural forest recovery is in these forest types following stand-replacing fire. In this study we quantified shrub characteristics and tree regeneration patterns in stand-replacing patches for five fires in the northern Sierra Nevada. These fires occurred between 1999 and 2008, and our field measurements were conducted in 2010. We analyzed tree regeneration patterns at two scales: patch level, in which field observations and spatial data were aggregated for a given stand-replacing patch, and plot level. Although tree regeneration densities varied considerably across sampled fires, over 50 % of the patches and approximately 80 % all plots had no tree regeneration. The percentage of patches, and to a greater extent plots, without pine regeneration was even higher, 72 and 87 %, respectively. Hardwood regeneration was present on a higher proportion of plots than either the pine or non-pine conifer groups. Shrub cover was generally high, with approximately 60 % of both patches and individual plots exceeding 60 % cover. Patch characteristics (size, perimeter-to-area ratio, distance-to-edge) appeared to have little effect on observed tree regeneration patterns. Conifer regeneration was higher in areas with post-fire management activities (salvage harvesting, planting). Our results indicate that the natural return of pine/mixed-conifer forests is uncertain in many areas affected by stand-replacing fire.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
There is considerable concern over the occurrence of stand-replacing fire in forest types historically associated with low- to moderate-severity fire. Characterizations of fire patterns in these forest types prior to Euro-American settlement vary in inferring the role of stand-replacing fire historically. Some studies demonstrate that stand-replacing fire was a relatively minor component of the fire regime, accounting for a small proportion of the total area burned (Brown et al. 2008; Scholl and Taylor 2010), and other studies demonstrating stand-replacing fire did occur at larger spatial extents, but occurred infrequently (Brown et al. 1999; Beaty and Taylor 2008). Irrespective of these inconsistencies, it almost ubiquitously acknowledged that contemporary patterns of stand-replacing fire in these forest types have deviated considerably from historical patterns (Graham 2003; Schoennagel et al. 2004; Keane et al. 2008; Miller et al. 2009a; but see Williams and Baker 2012). This is largely attributed to the changes in forest structure and composition associated with long-established fire suppression and past land management practices (timber harvesting, livestock grazing) (Parsons and Debenedetti 1979; Hessburg et al. 2005; Naficy et al. 2010; Collins et al. 2011). Another contributing factor could be that much of the area burned in contemporary wildfires coincides with fairly extreme fire weather conditions, as these are the conditions under which fires generally escape initial fire suppression efforts (Finney et al. 2011).
The impacts of this shift towards more extensive stand-replacing fire in forest types historically associated with low- to moderate-severity fire are largely unclear. Some studies suggest the early-successional plant communities that develop following stand-replacing fire contribute towards maintenance of landscape heterogeneity (Nagel and Taylor 2005; Skinner et al. 2006) and overall biodiversity (Odion et al. 2009; Swanson et al. 2011). However, given that many of the tree species associated with these forest types evolved largely with low- to moderate-intensity fire, and can withstand such fire, they generally have limited capacity to recover rapidly following extensive stand-replacing fire (Barton 2002; Goforth and Minnich 2008; Keeley 2012). Recent work from Pinus ponderosa forests in the southwestern U.S. demonstrated highly inconsistent pine regeneration in stand-replacing patches across 11 different fires, with half of the sampled patches having no pine regeneration 1–18 years post-fire (Roccaforte et al. 2012). Another similar study in the same region also reported very low pine regeneration densities for nearly half of the sampled sites 25 and 50 years post-fire (Savage and Mast 2005). Stand-replacing patches with poor conifer regeneration in these and other studies (Barton 2002; Strom and Fulé 2007; Goforth and Minnich 2008) tended to be dominated by shrubs and/or hardwoods, which suggests either very prolonged re-establishment of conifer forests (Nagel and Taylor 2005; Skinner et al. 2006) or conversion to new vegetation assemblages (Perry et al. 2011).
In contrast to the generally poor conifer regeneration following stand-replacing fire in forests historically associated with low- to moderate-severity fire, studies from mixed-conifer forests in the Klamath-Siskiyou Mountains, which have been historically associated with mixed-severity fire (Halofsky et al. 2011), demonstrated generally abundant conifer regeneration in stand-replacing patches 2–19 years post-fire (Shatford et al. 2007; Donato et al. 2009). Given the broad range of tree regeneration responses following stand-replacing fire across regions and the high variability even within a given region/forest type it is difficult to generalize what post-stand-replacing fire trajectories may be in conifer forest. This is particularly the case for Sierra Nevada mixed-conifer forests, which historically had a predominantly low- to moderate-severity fire regime (Swetnam and Baisan 2003; Stephens and Collins 2004; Scholl and Taylor 2010), but share several dominant tree species with Klamath-Siskiyou forests (Sugihara et al. 2006).
The re-establishment of trees following stand-replacing fire is a function of several factors, including species’ life history traits and interactions with other species (Connell and Slatyer 1977; Cattelino et al. 1979; Pickett et al. 1987), as well as stochastic processes (e.g. climate, seed production) (McCook 1994). In forests where dominant trees do not have serotinous cones nor vegetative reproduction, some of the key factors influencing tree establishment include seed dispersal (Cattelino et al. 1979), patch size (McDonald 1980; Bonnet et al. 2005; Haire and McGarigal 2010), and competition from other species (Connell and Slatyer 1977). Previous work in the Sierra Nevada has demonstrated considerable time (30–50 years) for conifers to establish following stand-replacing fire (Russell et al. 1998). The delayed tree establishment is largely due to the rapid and extensive establishment of shrubs via persistent soil seed banks (Russell et al. 1998; Nagel and Taylor 2005; Knapp et al. 2012).
In this study we quantified shrub characteristics and tree regeneration patterns in stand-replacing patches for five fires in the northern Sierra Nevada, which occurred from 1999 to 2008. We analyzed tree regeneration patterns at two scales: the patch level, in which field observations and spatial data are aggregated for a given stand-replacing patch, and the plot level. The aim of performing analysis at the two scales was to capture both coarser-grained (e.g. patch size, patch edge) and finer-grained (e.g. local topography, shrub cover, distance-to-patch-edge) effects on tree regeneration (Bonnet et al. 2005; Haire and McGarigal 2010). Our initial intent was to sample patches that did not experience post-fire management activities (salvage-harvest, planting, etc.). However, field observations revealed evidence of post-fire activities in some sampled patches, which then led us to incorporate a post-fire management component in the study. Our specific objectives were to: (1) identify the influences on tree regeneration at the both the patch and plot levels (patch characteristics, dominant pre-fire vegetation, shrub cover, post-fire management, and topography); and (2) characterize tree regeneration patterns by tree species groups (Pinus, non-Pinus conifers, hardwood) at both scales.
Methods
Study area and field sampling
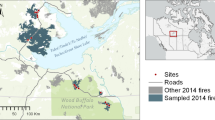
Our study area is located in the Plumas National Forest, and is situated in the northern Sierra Nevada at 39°56′N, 121°3′W (Fig. 1). Vegetation on this landscape is primarily mixed-conifer forest (Schoenherr 1992; Barbour and Major 1995), consisting of white fir (Abies concolor), Douglas-fir (Pseudotsuga menziesii), sugar pine (Pinus lambertiana), ponderosa pine (P. ponderosa), Jeffrey pine (P. jeffreyi), incense-cedar (Calocedrus decurrens), California black oak (Quercus kelloggii), and other less common hardwood species. Red fir (A. magnifica) is common at higher elevations, where it mixes with white fir. Montane chaparral and some meadows are interspersed throughout the forest. Tree density varies as a result of recent fire and timber management history, elevation, slope, aspect, and edaphic conditions. Historical fire occurrence, inferred from fire scars recorded in tree rings, suggests a fire regime with predominantly frequent, low- to moderate-severity fires occurring at intervals ranging from 7 to 12 years (Moody et al. 2006).

Fire perimeters, stand-replacing patches, and field plot locations for the five studied fires in the northern Sierra Nevada, California, USA
The study area was defined by five fairly recent fires that burned in relatively close proximity to one another within the Plumas National Forest: Lookout (1999), Pigeon (1999), Bucks Complex (1999), Storrie (2000), and Rich (2008) (Fig. 1; Table 1). We used satellite-derived estimates of fire severity to identify stand-replacing patches within these five fires. The fire severity estimates are based on the relative differenced Normalized Burn Ratio (RdNBR), which is computed from Landsat TM imagery (Miller and Thode 2007; Safford et al. 2008). This index has been used extensively to characterize recent fires and fire regimes (Holden et al. 2007; van Wagtendonk and Lutz 2007; Holden et al. 2009; Miller et al. 2009a; Miller et al. 2012). Miller et al. (2009b) report user’s and producer’s accuracies for RdNBR-based high severity classification to range between 70.7 and 85.3 %, indicating RdNBR robustly captures stand-replacing fire effects. Using classified RdNBR images (thresholds based on Miller and Thode (2007)) we employed the patch delineation algorithm PatchMorph (Girvetz and Greco 2007) to identify spatially contiguous patches of stand-replacing fire that were >2 ha. The PatchMorph tool is compatible with ArcGIS, and has been used previously to delineate stand-replacing patches (Collins and Stephens 2010). We chose a minimum patch size of 2 ha because we were interested in characterizing tree regeneration and shrub response within larger areas that were generally devoid of substantial tree seed sources.
Our field sampling efforts were constrained by access, crew safety concerns, and time. As such, we were only able to sample a subset of the patches identified by the PatchMorph tool (Fig. 1). Field plot locations were established using a systematic grid of points with 200 m spacing, which were based on a random starting location for each stand-replacing patch. We intensified sampling near patch edges to investigate potential distance-to-edge effects on tree and shrub dynamics. The rules for plot intensification were as follows, if grid points were: (1) <25 m from patch edge there were no additional plots, (2) 25–75 m from patch edge a plot was added that was 25 m towards the nearest patch edge, (3) 75–125 m from patch edge plots were added at 50 m both towards and away from the nearest patch edge, and (4) >125 m from patch edge there were no additional plots. We sampled 26 stand-replacing patches across the five fires, with a total of 277 field plots. Five of the 26 sampled patches were treated post-fire; three were salvage-logged only, and two were salvage-logged and planted. These treated patches accounted for 112 field plots.
At each plot we established three, ten-meter transects using a random starting azimuth for the first transect and then adding 120° for the next two (Fig. 2). Along each transect we measured shrub length, average shrub height, and ground cover. Ground cover was broken down into cover classes of woody shrubs, herbaceous plants, grass, litter, wood (downed trees), rock, and bare mineral soil. Tree regeneration was sampled on the same three transects. Trees with a minimum diameter-at-breast-height (dbh) of 0.1 cm were tallied by species on a 2 m wide belt, and established seedlings <1.4 m tall (no minimum size) tallied by species in circular plots at the end of the transect (Fig. 2). We measured live “overstory” and standing dead trees based on a variable radius plot using a 20 basal-area-factor prism.

Sampling design for all field plots
Data analysis
We performed analyses at two levels, individual plots and stand-replacing patches, with the intent of exploring both site- and patch-level influences on tree regeneration. For each plot and sampled stand-replacing patch, we assembled a suite of explanatory variables that characterize the pre- and post-fire vegetation, topography, and patch characteristics. Pre-fire vegetation was based on a 1997 Landsat-derived map produced by the U. S. Forest Service Region 5 (J. Miller, personal communication, 2011). The map provided broad vegetation type (mixed-conifer, mixed hardwood, mixed chaparral, and white fir) and canopy cover classes (25–39, 40–59, and 60–100 %). Post-fire vegetation variables were: live tree basal area, shrub cover, and average shrub height. Topographic variables included: slope steepness, aspect, and elevation. For the plot level analysis, the topographic variables were based on field observations, with elevation acquired via handheld GPS. For the patch level analysis these variables were averages for a given patch derived from a 10 m digital elevation model. Additional explanatory variables included post-fire management (salvage-logged only, salvage-logged and planted, or none), as well as distance-to-patch-edge at the individual plot level, and patch area and perimeter-to-area ratio at the patch level.
We used these explanatory variables to explore patterns in tree regeneration densities, which we binned in three groups: pines (P. ponderosa and P. lambertiana), non-pine conifers (A. concolor, A. magnifica, C. decurrens, and P. menziesii), and hardwoods (predominantly Quercus kelloggii). Hardwood regeneration was nearly all a result of basal sprouting from top-killed trees, which was determined by proximity to burned tree bases. At the plot level, tree tallies were multiplied by the appropriate scaling factor based the tree size/sampling area to obtain per hectare tree regeneration densities. Patch-level densities were the average of all plots within a given patch. Tree regeneration densities were log-transformed due the very large range in observed densities.
We explored possible relationships between these log-transformed tree densities and the explanatory variables using regression tree analysis. Regression tree analysis offers distinct advantages over traditional linear models because it can handle nonlinear or discontinuous relationships between variables, and high-order interactions (Breiman et al. 1984). In addition, the hierarchical structure and identification of potential threshold values for independent variables is well suited for explaining ecological phenomena (De’ath and Fabricius 2000; Collins et al. 2009; Thompson et al. 2011). The regression tree is constructed by repeatedly splitting the data into increasingly homogenous groups based on identified influential explanatory variables. We used the conditional inference tree technique in the PARTY library, within the statistical package R (Hothorn et al. 2009). This technique identifies influential explanatory variables using a partitioning algorithm that is based on the lowest statistically significant P value derived from Monte Carlo simulations. This minimizes bias and prevents over-fitting of the data, which is a common problem with regression trees (Hothorn et al. 2006; Thompson et al. 2011). Given the tremendous variability in seedling establishment observed in the field and from previous studies we report results based on an α-level of 0.1 rather than the typical 0.05.
There is a potential issue regarding independence among observations in our dataset that deserves attention. The fact that there are multiple plots within individual patches, and multiple patches within individual fires implies possible violation of the assumption that samples are independent, hence pseudoreplication (Hurlbert 1984). While this is an important consideration, particularly because it has the potential to confound interpretations and limit applicability of findings to other areas, it should be noted that fire is dynamic process that is capable generating highly heterogeneous patterns of effects based on fine-scale variations in vegetation/fuels, weather, and topography (e.g. Knapp and Keeley 2006; Collins et al. 2007). This heterogeneity contributes to an overall lack of spatial dependence among fire effects subsamples within a given fire (van Mantgem and Schwilk 2009). As van Mantgem et al. (2001) point out “it is not possible to replicate or randomize large-scale events such as wildfires” and as a result we may never overcome the potential for pseudoreplication in studies of actual wildfire. Perhaps by studying multiple fires across a fairly large and topographically diverse area we are able to at least partially address these concerns (van Mantgem et al. 2001).
Results
Although tree regeneration densities varied considerably across the five sampled fires, over 50 % of the sampled stand-replacing patches and approximately 80 % of all plots had no tree regeneration (Fig. 3). The percentage of patches and plots without pine regeneration was even higher, 72 and 87 %, respectively. However, nearly 20 % of the sampled patches had non-pine conifer regeneration in excess of 2,000 trees ha−1 (Fig. 3). Of the three species groups, hardwood was clearly the most common at intermediate densities, with almost 30 % of the sampled patches averaging between 400 and 1,400 trees ha−1. Shrub cover was generally high, with approximately 60 % of both the patches and individual plots exceeding 60 % cover (Fig. 4). Average shrub height across all plots with shrubs present was 86 cm (n = 263). Although there were some plots with no shrub cover (n = 14), all patches on average had shrub cover >0 (Fig. 4). The actual distribution of distance-to-patch-edge for established plots emphasizes intensification near patch edges, with 58 % of our plots occurring within 75 m of patch edges (Fig. 5).

Frequency distributions for sampled tree regeneration, by species group, for individual plots, and averaged for sampled stand-replacing patches. The density values on the horizontal axis represent the upper bound for each bin

Frequency distributions for sampled shrub cover for individual plots, and averaged for stand-replacing patches. The cover values on the horizontal axis represent the upper bound for each bin

Frequency distribution of distance-to-nearest-patch edge for field plots (n = 277). Distance values represent the upper bound for each bin
The regression tree analysis at the patch level identified shrub cover as the only significant predictor of both non-pine conifer and hardwood regeneration density (results not shown). For both species groups, average regeneration density tended to be higher in patches with average shrub cover over 25 %. The same analysis failed to identify any significant variables explaining the variability in pine regeneration density at the patch level. It is likely that this lack of significant explanatory variables is driven by the absence of pine regeneration on such a high number of patches. At the plot level, the most important factor influencing non-pine conifer regeneration was post-fire management (Fig. 6). The next most important explanatory factors were elevation and live tree basal area (Fig. 6). The highest non-pine conifer regeneration density was associated with plots that were salvaged-logged alone and occurred at elevations below 1,635 m (minimum elevation in this group was 1,515 m). In salvage + planting and no treatment plots, higher regeneration densities were in areas with live basal area over 4.6 m2/ha and on north-facing aspects. The lowest non-pine conifer regeneration density was associated with plots that were salvaged-logged alone and occurred at elevations above 1,635 m (Fig. 6). The significant factors explaining the variability in hardwood regeneration density were distance to nearest patch edge, slope, and shrub height. The highest hardwood densities were associated with plots further from patch edge, while the lowest densities were associated with plots closer to patch edges that were on flatter slopes (Fig. 6). Post-fire management activity was the only variable that significantly explained variability in plot level pine regeneration density. Not surprisingly, plots that were salvage-harvested and then planted had the highest pine densities. Plots that were salvage-harvested alone or were not treated had, for the most part, zero pine regeneration, with a few outliers (Fig. 6).

Regression tree output explaining the influence of the identified variables on plot level tree regeneration by species group. The length of the line from each split indicates the relative proportion of total sum of squares explained by that split. Box-and-whisker plots displaying regeneration density per hectare (log scale) and number of plots (n) are reported for the resulting groups at each terminal node. Diamonds in each box-and-whisker plot represent mean densities for each group
Discussion
The limitations inherent in tree-ring based fire reconstructions, such as incomplete spatial coverage and imperfect preservation of evidence, make it difficult to ascertain the historical role of stand-replacing fire in forests generally associated with low- to moderate-intensity fire (Bekker and Taylor 2001; Hessburg et al. 2007; Beaty and Taylor 2008; Brown et al. 2008). As a result, there is considerable uncertainty regarding what “natural” recovery is following extensive stand-replacing fire in these forest types. The absence of direct mechanisms for conifer regeneration following extensive stand-replacing fire (e.g. serotiny, sprouting, soil seed banks) in these forest types suggest that re-establishment of mature conifer forests would be slow. This has been demonstrated in previous studies within the Sierra Nevada (Conard and Radosevich 1982; Russell et al. 1998; Nagel and Taylor 2005). Our results demonstrating that over half of the sampled stand-replacing patches had no tree regeneration and almost three-quarters of the patches had no pine regeneration 2–11 years post-fire are consistent with the findings from these studies (Fig. 3). Excluding patches that had post-fire management activities, there was no pine regeneration in over 90 % of sampled patches (2 of 21 patches had pine regeneration). This lack of pine regeneration is particularly problematic given that one of the stated goals of the U. S. Forest Service both nationally and within the Sierra Nevada region is to restore forest structure and composition to that resembling historical forest conditions (USFS 2011).
The RdNBR threshold we used for identifying stand-replacing fire effects corresponds to approximately 95 % overstory mortality (Miller et al. 2009b). Our field observations confirmed this, with only 15 of our 277 plots recording live overstory trees. (According to the 1997 vegetation map all but 18 of our 277 plots were classified as mixed-conifer forest prior to burning in the five fires studied.) Based on the nearly complete overstory mortality and that these mixed-conifer forests are composed of non-serotinous conifers, seed for regenerating trees would need to come from surrounding areas (Goforth and Minnich 2008; Keeley 2012). As such, we expected some signature of patch characteristics (sizes, perimeter-to-area ratios) on conifer regeneration, e.g. smaller patches and patches with greater proportions of edge would have greater conifer regeneration. We found no effect of patch characteristics on conifer regeneration, despite the considerable range in patch sizes and shapes across our sampled patches (Fig. 1). This lack of patch effects, combined with failure to identify any effects of coarser-grained topography or pre-fire vegetation type/structure suggests that factors at scales other than those captured by our patch level variables govern tree regeneration following stand-replacing fire. Furthermore, it emphasizes the complex and perhaps highly stochastic nature of tree re-colonization in stand-replacing fire areas. Given this complexity, perhaps the site or plot level is more appropriate for investigating the influences on tree regeneration.
The lack of a significant distance-to-patch-edge effect on tree regeneration densities for both conifer groups is somewhat surprising. Given that for almost two-thirds of the patches we sampled, field observations took place 10–11 years post-fire, we expected there would have been sufficient time for tree establishment, particularly nearer to patch edges (Greene and Johnson 2000; Bonnet et al. 2005; Haire and McGarigal 2010). It does not appear that this lack of an effect is a product of our sampling intensity near patch edges (Fig. 5). Nearly 60 % of our plots were within 75 m of patch edges, which is within the two tree length distance that serves as a general “rule-of-thumb” for seed dispersal in Sierran mixed-conifer forests (McDonald 1980) (R. Tompkins, personal communication, 2010). Perhaps more time following stand-replacing fire will be needed for dispersal patterns in conifer establishment to emerge in these patches (Russell et al. 1998; Haire and McGarigal 2010).
Given the retrospective nature of our study we lack specific observations of the successional processes that took place immediately following stand-replacing fire (<1 year). Seed production and climate, which both exhibit high interannual variability, have considerable influence on tree establishment. We lack information on seed inputs from surrounding trees or seed banks in the soil, as well as specific measurements of available soil moisture. Together these shortcomings limit our ability to describe the mechanisms behind observed patterns in tree regeneration and shrub abundance. That said, our findings, and those of other studies (Russell et al. 1998; Goforth and Minnich 2008) suggest the lack of conifer regeneration in stand-replacing patches is largely due to interacting effects of unfavorable conditions for tree establishment immediately following the fires, i.e. low seed production and/or low soil moisture, and the presence of shrubs. The two most common genera of shrubs, Ceanothus and Arctostaphylos, have persistent soil seed banks, much of which remains viable after burning (Conard and Radosevich 1982; Knapp et al. 2012) and/or has fire-cued germination (Kauffman and Martin 1991). This allows for rapid colonization following stand-replacing fire. Once established, these shrubs drive down soil moisture which can make conifer establishment more difficult (Conard and Radosevich 1982; McDonald and Fiddler 1989; Gray et al. 2005; Nagel and Taylor 2005). Previous work in the Sierra Nevada has demonstrated that despite strong shrub occupancy, conifers can establish, but it may take several decades (Russell et al. 1998). The lengthy time for conifer establishment combined with the time it takes for trees to reach a size where they can resist fire encompasses a long period during which patches are vulnerable to repeat fires. Repeat fires in shrub-dominated patches are characteristically stand-replacing and would generally be lethal to conifer regeneration (Biswell 1974; Nagel and Taylor 2005). Given the documented increases in stand-replacing fire in these forests (Miller et al. 2009a; Miller and Safford 2012), along with the projections for further increases due to climate change (e.g. Safford et al. 2012), there is real potential for substantial ecosystem shifts towards shrub- and/or hardwood-dominated communities (Barton 2002; Goforth and Minnich 2008; Perry et al. 2011).
The identification of post-fire activity as the strongest predictor of pine regeneration warrants some attention. While it is no surprise that the highest pine densities occurred in plots that were salvage-harvested/planted, it is surprising how little pine regeneration there was in untreated plots (Fig. 6). We detected pine regeneration in only 5 of the 165 untreated plots. Other post-wildfire studies in similar forest types have also found much lower proportions of pine regeneration relative to other tree species (Russell et al. 1998; Barton 2002; Crotteau et al. 2013). Limitations in pine seed production, which is generally occurs on 4–7 year cycles (McDonald 1992), and dispersal relative other conifers, as well as seed predation are all factors that may be contributing to this (Zald et al. 2008). Of these factors seed predation by rodents may be particularly important (Fowells and Stark 1965), which can be greater both in burned areas relative to unburned areas, and for pine seeds relative to seeds of other conifers (Zwolak et al. 2010). The low pine regeneration in stand-replacing patches observed in ours and other studies only reinforces the compositional shift in mixed-conifer forests towards shade-tolerant tree species brought about by fire exclusion and past management practices (Moghaddas et al. 2008; Collins et al. 2011; Crotteau et al. 2013). While we did not sample outside of stand-replacing patches, it is possible that greater pine regeneration in areas burned less severely (Crotteau et al. 2013).
The findings indicating different factors contributing to greater non-pine conifer versus hardwood densities suggests different influences contributing to observed patterns for the two species groups (Fig. 6). Non-pine conifer densities on salvage-logged only plots, particularly at lower elevations, were well over an order of magnitude higher than all other plots (Fig. 6). It appears that the ground disturbance associated with salvage harvesting in these areas may have facilitated germination and seedling establishment, which was predominantly Abies sp. (Gordon 1979). This effect appears to be considerably pronounced at intermediate elevations (1,515–1,635 m), where exposure to temperature extremes may be moderated (Fig. 6). This was not the case for hardwood regeneration, which overall was present on a higher proportion of plots (22 %) than either the pine or non-pine conifer groups (13 and 17 %, respectively). Hardwood densities were highest on plots that were well interior to stand-replacing patches (Fig. 6), suggesting a possible insulating effect. One hypothesis is that browsing pressure, which can be fairly high on black oak (Bowyer and Bleich 1980), could be lesser on more interior portions of patches, perhaps due to the high shrub cover limiting access for browsing animals. The indication of greater hardwood densities in plots with taller shrubs (Fig. 6) supports this hypothesis. While there were several untreated plots that had non-pine conifer regeneration, with some sites having very high densities (~10,000 ha−1), the overwhelming majority of untreated plots had zero non-pine conifer regeneration (143 of 165 plots). The number of untreated plots without any hardwood regeneration was lower, only 121 of 165 plots.
Summary and management implications
A recent assessment of land cover change in California demonstrated that fire now accounts for a greater proportion of forest “loss” than any other activity (e.g. timber harvesting, development) (Sleeter et al. 2011). Given the observed and predicted future trends toward increasing temperatures and longer fire seasons (Westerling et al. 2006, 2011; Safford et al. 2012) it appears that fire-driven forest change will only continue to increase (Miller et al. 2009a). If this is the case, it begs the question of what type of forest, if any, will be coming back after fire, particularly after stand-replacing fire? Our results, and the results from other studies (Barton 2002; Goforth and Minnich 2008), indicate that the return of mixed-conifer forests is uncertain in many areas affected by stand-replacing fire. This is particularly the case for the pine component within mixed-conifer forests. The vigorous response of shrubs in the patches we sampled, coupled with the greater proportion of plots with hardwood regeneration suggest a compositional shift in the near term, if not a possible longer-term conversion of these mixed-conifer forests. If the predicted regional increases in future fire activity are realized (McKenzie et al. 2004; Westerling et al. 2011) then shorter intervals between successive fires are likely (Safford et al. 2012). Re-burns of these stand-replacing patches may result in a shift from mixed-conifer forests toward shrub- and hardwood-dominated systems, especially if the re-burns occur under more severe fire weather conditions (Perry et al. 2011). If the desired condition for mixed-conifer forests affected by stand-replacing fire is to have a mixed-conifer forests return within a reasonable timeframe then based on our results it appears that management intervention, mainly planting pine species, may be needed in some areas to insure the condition can be achieved.
References
Barbour MG, Major J (eds) (1995) Terrestrial vegetation of California: new expanded. California Native Plant Society, Davis
Barton AM (2002) Intense wildfire in southeastern Arizona: transformation of a Madrean oak-pine forest to oak woodland. For Ecol Manage 165:205–212
Beaty RM, Taylor AH (2008) Fire history and the structure and dynamics of a mixed conifer forest landscape in the northern Sierra Nevada, Lake Tahoe Basin, California, USA. Forest Ecol Manage 255:707–719
Bekker MF, Taylor AH (2001) Gradient analysis of fire regimes in montane forests of the southern Cascade range, Thousand Lakes Wilderness, California, USA. Plant Ecol 155:15–28
Biswell HH (1974) Effects of fire on chaparral. In: Kozlowski TT, Ahlgren CE (eds) Fire and ecosystems. Academic Press, New York, pp 321–364
Bonnet VH, Schoettle AW, Shepperd WD (2005) Postfire environmental conditions influence the spatial pattern of regeneration for Pinus ponderosa. Can J For Res 35:37–47
Bowyer RT and Bleich VC 1980. Ecological relationships between southern mule deer and California black oak. In Plumb TR (ed) Proceedings of the Symposium on Ecology, Management, and Utilization of California Oaks, pp. 292–296. U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station, Berkeley
Breiman L, Friedman JH, Olshen RA, Stone CG (1984) Classification and regression trees. Wadsworth, Belmont
Brown PM, Kaufmann MR, Shepperd WD (1999) Long-term, landscape patterns of past fire events in a montane ponderosa pine forest of central Colorado. Landscape Ecol 14:513–532
Brown PM, Wienk CL, Symstad AJ (2008) Fire and forest history at Mount Rushmore. Ecol Appl 18:1984–1999
Cattelino PJ, Noble IR, Slatyer RO, Kessell SR (1979) Predicting the multiple pathways of plant succession. Environ Manage 3:41–50
Collins BM, Stephens SL (2010) Stand-replacing patches within a ‘mixed severity’ fire regime: quantitative characterization using recent fires in a long-established natural fire area. Landscape Ecol 25:927–939
Collins BM, Kelly M, van Wagtendonk JW, Stephens SL (2007) Spatial patterns of large natural fires in Sierra Nevada wilderness area. Landscape Ecol 22:545–557
Collins BM, Miller JD, Thode AE, Kelly M, van Wagtendonk JW, Stephens SL (2009) Interactions among wildland fires in a long-established Sierra Nevada natural fire area. Ecosystems 12:114–128
Collins BM, Everett RG, Stephens SL (2011) Impacts of fire exclusion and managed fire on forest structure in an old growth Sierra Nevada mixed-conifer forest. Ecosphere 2:51
Conard SG, Radosevich SR (1982) Post-fire succession in white fir (Abies concolor) vegetation of the northern Sierra Nevada. Madrono 29:42–56
Connell JH, Slatyer RO (1977) Mechanisms of succession in natural communities and their role in community stability and organization. Am Nat 111:1119–1144
Crotteau JS, Varner JM, Ritchie MW (2013) Post-fire regeneration across a fire severity gradient in the southern Cascades. For Ecol Manage 287:103–112
De’ath G, Fabricius KE (2000) Classification and regression trees: a powerful yet simple technique for ecological data analysis. Ecology 81:3178–3192
Donato DC, Fontaine JB, Campbell JL, Robinson WD, Kauffman JB, Law BE (2009) Conifer regeneration in stand-replacement portions of a large mixed-severity wildfire in the Klamath-Siskiyou Mountains. Can J For Res 39:823–838
Finney MA, McHugh CW, Grenfell IC, Riley KL, Short KC (2011) A simulation of probabilistic wildfire risk components for the continental United States. Stoch Environ Res Risk Assess 25:973–1000
Fowells HA and Stark NB (1965). Natural regeneration in relation to environment in the mixed conifer forest type of California. General Technical Report PSW-24. U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station, Berkeley, p 14
Girvetz EH, Greco SE (2007) How to define a patch: a spatial model for hierarchically delineating organism-specific habitat patches. Landscape Ecol 22:1131–1142
Goforth BR, Minnich RA (2008) Densification, stand-replacement wildfire, and extirpation of mixed conifer forest in Cuyamaca Rancho State Park, southern California. For Ecol Manage 256:36–45
Gordon DT (1979) Successful natural regeneration cuttings in California true firs. General Technical Report PSW-140. U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station, Berkeley, p 14
Graham RT, Technical Editor (2003). Hayman fire case study. General Technical Report RMRS-GTR-114. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ogden, p 396
Gray AN, Zald HSJ, Kern RA, North M (2005) Stand conditions associated with tree regeneration in Sierran mixed-conifer forests. For Sci 51:198–210
Greene DF, Johnson EA (2000) Tree recruitment from burn edges. Can J For Res 30:1264–1274
Haire SL, McGarigal K (2010) Effects of landscape patterns of fire severity on regenerating ponderosa pine forests (Pinus ponderosa) in New Mexico and Arizona, USA. Landscape Ecol 25:1055–1069
Halofsky JE, Donato DC, Hibbs DE, Campbell JL, Cannon MD, Fontaine JB, Thompson JR, Anthony RG, Bormann BT, Kayes LJ, Law BE, Peterson DL and Spies TA (2011) Mixed-severity fire regimes: lessons and hypotheses from the Klamath-Siskiyou Ecoregion 2: art40
Hessburg PF, Agee JK, Franklin JF (2005) Dry forests and wildland fires of the inland Northwest USA: contrasting the landscape ecology of the pre-settlement and modern eras. For Ecol Manage 211:117–139
Hessburg PF, Salter RB, James KM (2007) Re-examining fire severity relations in pre-management era mixed conifer forests: inferences from landscape patterns of forest structure. Landscape Ecol 22:5–24
Holden ZA, Morgan P, Crimmins MA, Steinhorst RK, Smith AM (2007) Fire season precipitation variability influences fire extent and severity in a large southwestern wilderness area United States. Geophys Res Let 34:L16708
Holden ZA, Morgan P, Evans JS (2009) A predictive model of burn severity based on 20-year satellite-inferred burn severity data in a large southwestern US wilderness area. For Ecol Manage 258:2399–2406
Hothorn T, Hornik K, Zeileis A (2006) Unbiased recursive partitioning: a conditional inference framework. J Comput Graph Stat 15:651–674
Hothorn T, Hornik K, Strobl C and Zeileis A (2009) PARTY: a laboratory for recursive partitioning. (http://CRAN.R-project.org/package=party)
Hurlbert SH (1984) Psuedoreplication and the design of ecological field experiments. Ecol Monogr 54:187–211
Kauffman JB, Martin RE (1991) Factors influencing the scarification and germination of three montane Sierra Nevada shrubs. Northwest Sci 65:180–187
Keane RE, Agee JK, Fulé PZ, Keeley JE, Key CH, Kitchen SG, Miller R, Schulte LA (2008) Ecological effects of large fires on US landscapes: benefit or catastrophe. Int J Wildland Fire 17:696–712
Keeley JE (2012) Ecology and evolution of pine life histories. Ann For Sci 69:445–453
Knapp EE, Keeley JE (2006) Heterogeneity in fire severity within early season and late season prescribed burns in a mixed-conifer forest. Int J Wildland Fire 15:37–45
Knapp EE, Weatherspoon CP, Skinner CN (2012) Shrub seed banks in mixed conifer forests of northern California and the role of fire in regulating abundance. Fire Ecol 8:32–48
McCook LJ (1994) Understanding ecological community succession. causal models and theories, a review. Vegetatio 110:115–147
McDonald PM (1980) Seed dissemination in small clearcuttings in north-central California. General Technical Report PSW-150. U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station, Berkeley, p 5
McDonald PM and Fiddler GO (1989) Competing vegetation in ponderosa pine plantations: ecology and control. General Technical Report PSW-113. U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station, Berkeley, p 26
McDonald PM (1992) Estimating seed crops of conifer and hardwood species. Can J For Res 22:832–838
McKenzie D, Gedalof Z, Peterson DL, Mote P (2004) Climate change, wildfire, and conservation. Conserv Biol 18:890–902
Miller JD, Thode AE (2007) Quantifying burn severity in a heterogeneous landscape with a relative version of the delta normalized burn ratio (dNBR). Remote Sens Environ 109:66–80
Miller JD, Safford HD, Crimmins M, Thode AE (2009a) Quantitative evidence for increasing forest fire severity in the Sierra Nevada and southern Cascade Mountains, California and Nevada, USA. Ecosystems 12:16–32
Miller JD, Knapp EE, Key CH, Skinner CN, Isbell CJ, Creasy RM, Sherlock JW (2009b) Calibration and validation of the relative differenced Normalized Burn Ratio (RdNBR) to three measures of fire severity in the Sierra Nevada and Klamath Mountains, California, USA. Remote Sens Environ 113:645–646
Miller JD, Safford HD (2012) Trends in wildfire severity 1984–2010 in the Sierra Nevada, Modoc Plateau and southern Cascades, California, USA. Fire Ecol 8:41–57
Miller JD, Skinner CN, Safford HD, Knapp EE, Ramirez CM (2012) Trends and causes of severity, size, and number of fires in northwestern California, USA. Ecol Appl 22:184–203
Moghaddas JJ, York RA, Stephens SL (2008) Initial response of conifer and California black oak seedlings following fuel reduction activities in a Sierra Nevada mixed conifer forest. For Ecol Manage 255:3141–3150
Moody TJ, Fites-Kaufman J, Stephens SL (2006) Fire history and climate influences from forests in the northern Sierra Nevada, USA. Fire Ecol 2:115–141
Naficy C, Sala A, Keeling EG, Graham J, DeLuca TH (2010) Interactive effects of historical logging and fire exclusion on ponderosa pine forest structure in the northern Rockies. Ecol Appl 20:1851–1864
Nagel TA, Taylor AH (2005) Fire and persistence of montane chaparral in mixed conifer forest landscapes in the northern Sierra Nevada, Lake Tahoe Basin, California, USA. J Torrey Bot Soc 132:442–457
Odion DC, Moritz MA, DellaSala DA (2009) Alternative community states maintained by fire in the Klamath Mountains USA. J Ecol. doi:10.1111/j.1365-2745.2009.01597.x
Parsons DJ, Debenedetti SH (1979) Impact of fire suppression on a mixed-conifer forest. For Ecol Manage 2:21–33
Perry DA, Hessburg PF, Skinner CN, Spies TA, Stephens SL, Taylor AH, Franklin JF, McComb B, Riegel G (2011) The ecology of mixed severity fire regimes in Washington, Oregon, and Northern California. Forest Ecol Manage 262:703–717
Pickett STA, Collins SL, Armesto JJ (1987) Models, mechanisms and pathways of succession. Bot Rev 53:335–371
Roccaforte JP, Fule PZ, Chancellor WW, Laughlin DC (2012) Woody debris and tree regeneration dynamics following severe wildfires in Arizona ponderosa pine forests. Can J For Res 42:593–604
Russell WH, McBride JR, Rowntree R (1998) Revegetation after four stand-replacing fires in the Lake Tahoe Basin. Madrono 45:40–46
Safford HD, Miller JD, Schmidt D, Roath B, Parsons A (2008) BAER soil burn severity maps do not measure fire effects to vegetation: a comment on Odion and Hanson (2006). Ecosystems 11:1–11
Safford HD, North M, Meyer MD (2012) Chapter 3: climate change and the relevance of historical forest conditions. In: North M (ed) Managing Sierra Nevada forests. U. S. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany, pp 23–46
Savage M, Mast JN (2005) How resilient are southwestern ponderosa pine forests after crown fires? Can J For Res 35:967–977
Schoenherr AA (1992) A natural history of California. University of California Press, Berkeley
Schoennagel T, Veblen TT, Romme WH (2004) The interaction of fire, fuels, and climate across Rocky Mountain forests. Bioscience 54:661–676
Scholl AE, Taylor AH (2010) Fire regimes, forest change, and self-organization in an old-growth mixed-conifer forest, Yosemite National Park, USA. Ecol Appl 20:362–380
Shatford JPA, Hibbs DE, Puettmann KJ (2007) Conifer regeneration after forest fire in the Klamath-Siskiyous: How much, how soon? J Forest 105:139–146
Skinner CN, Taylor AH, Agee JK (2006) Klamath Mountains bioregion. In: Sugihara NG, van Wagtendonk JW, Fites-Kaufman J, Shaffer KE, Thode AE (eds) Fire in California’s ecosystems. University of California Press, Berkeley, pp 170–194
Sleeter BM, Wilson TS, Soulard CE, Liu J (2011) Estimation of late twentieth century land-cover change in California. Environ Monit Assess 173:251–266
Stephens SL, Collins BM (2004) Fire regimes of mixed conifer forests in the north-central Sierra Nevada at multiple spatial scales. Northwest Sci 78:12–23
Strom BA, Fulé PZ (2007) Pre-wildfire fuel treatments affect long-term ponderosa pine forest dynamics. Int J Wildland Fire 16:128–138
Sugihara NG, van Wagtendonk JW, Shaffer KE, Fites-Kaufman J, Thode AE (2006) Fire in California’s ecosystems. University of California Press, Berkeley
Swanson ME, Franklin JF, Beschta RL, Crisafulli CM, Dellasala DA, Hutto RL, Lindenmayer DB, Swanson FJ (2011) The forgotten stage of forest succession: early-successional ecosystems on forest sites. Front Ecol Environ 9:117–125
Swetnam TW, Baisan CH (2003) Tree-ring reconstructions of fire and climate history in the Sierra Nevada and southwestern United States. In: Veblen TT, Baker WL, Montenegro G, Swetnam TW (eds) Fire and climatic change in temperate ecosystems of the western Americas Ecological Studies, vol 160. Springer, New York, pp 158–195
Thompson JR, Foster DR, Scheller R, Kittredge D (2011) The influence of land use and climate change on forest biomass and composition in Massachusetts, USA. Ecol Appl 21:2425–2444
USFS (2011) Region five ecological restoration: leadership intent. March 2011. U.S. Forest Service, Pacific Southwest Region, p 4
van Mantgem PJ, Schwartz M, Keifer M (2001) Monitoring fire effects for managed burns and wildfires: coming to terms with pseudoreplication. Nat Areas J 21:266–273
van Mantgem PJ, Schwilk DW (2009) Negligible influence of spatial autocorrelation in the assessment of fire effects in a mixed conifer forest. Fire Ecol 5:116–125
van Wagtendonk JW, Lutz JA (2007) Fire regime attributes of wildland fires in Yosemite National Park, USA. Fire Ecol 3:34–52
Westerling A, Bryant B, Preisler H, Holmes T, Hidalgo H, Das T, Shrestha S (2011) Climate change and growth scenarios for California wildfire. Climatic Change 109:445–463
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006) Warming and earlier spring increase western US forest wildfire activity. Science 313:940–943
Williams MA, Baker WL (2012) Spatially extensive reconstructions show variable-severity fire and heterogeneous structure in historical western United States dry forests. Glob Ecol Biogeogr. doi:10.1111/j.1466-8238.2011.00737.x
Zald HSJ, Gray AN, North M, Kern RA (2008) Initial tree regeneration responses to fire and thinning treatments in a Sierra Nevada mixed-conifer forest, USA. For Ecol Manage 256:168–179
Zwolak R, Pearson DE, Ortega YK, Crone EE (2010) Fire and mice: seed predation moderates fire’s influence on conifer recruitment. Ecology 91:1124–1131
Acknowledgments
We would like to thank Christopher Dow and Anu Kramer for their assistance in the field under some of the toughest field sampling conditions we have been involved in. Anu Kramer also helped with GIS support. Jay Miller kindly provided the pre-fire vegetation maps. We thank the Plumas National Forest staff, in particular Ryan Tompkins, for providing information and input on this work. This work was funded by the Storrie Fire Restoration Project and the Plumas-Lassen Administration Study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Collins, B.M., Roller, G.B. Early forest dynamics in stand-replacing fire patches in the northern Sierra Nevada, California, USA. Landscape Ecol 28, 1801–1813 (2013). https://doi.org/10.1007/s10980-013-9923-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-013-9923-8