
Abstract
Context
An increase in the incidence of large wildfires worldwide has prompted concerns about the resilience of forest ecosystems, particularly in the western U.S., where recent changes are linked with climate warming and 20th-century land management practices.
Objectives
To study forest resilience to recent wildfires, we examined relationships among fire legacies, landscape features, ecological conditions, and patterns of post-fire conifer regeneration.
Methods
We quantified regeneration across 182 sites in 21 recent large fires in dry mixed-conifer forests of the U.S. northern Rockies. We used logistic and negative binomial regression to predict the probability of establishment and abundance of conifers 5–13 years post-fire.
Results
Seedling densities varied widely across all sites (0–127,500 seedlings ha−1) and were best explained by variability in distance to live seed sources (β = −0.014, p = 0.002) and pre-fire tree basal area (β = 0.072, p = 0.008). Beyond 95 m from the nearest live seed source, the probability of seedling establishment was low. Across all the fires we studied, 75 % of the burned area with high tree mortality was within this 95-m threshold, suggesting the presence of live seed trees to facilitate natural regeneration.
Conclusions
Combined with the mix of species present within the burn mosaic, dry mixed-conifer forests will be resilient to large fires across our study region, provided that seedlings survive, fire do not become more frequent, high-severity patches do not get significantly larger, and post-fire climate conditions remain suitable for seedling establishment and survival.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Large wildfires have been increasing worldwide over the past several decades (Kasischke and Turetsky 2006; Westerling et al. 2006; Pausas and Fernández-Muñoz 2012), a pattern predicted to continue in many forested regions with even moderate climate warming (Flannigan et al. 2009; Pechony and Shindell 2010; Littell 2011; Rogers et al. 2011). Large disturbances, including wildfires, shape ecosystem structure and function for decades to centuries, and shifts in their frequency, size, or intensity can have unknown implications for forest resilience (Turner 2010). We use the term “resilience” to describe the capacity of a system to absorb disturbance without transitioning into a qualitatively different state, emphasizing the maintenance of system structure and function (Holling 1973; Groffman et al. 2006; Moritz et al. 2011). Large, intense wildfires can reduce forest resilience by shifting post-fire species assemblages (Johnstone et al. 2010a, b) or initiating type conversions to non-forest vegetation (Savage and Mast 2005; Odion et al. 2010). Thus, understanding forest resilience to future wildfires, particularly in the context of climate change, depends upon identifying the mechanisms that influence tree regeneration, survival and composition.
Dry mixed-conifer forests of the western U.S. may be particularly vulnerable to ongoing and future shifts in wildfire activity (e.g., Williams et al. 2010), given the combined effects of 20th-century land use and land management practices on species composition, fuel loads, and fire regimes (e.g., Hessburg et al. 2000; Keeling et al. 2006; Naficy et al. 2010). Of particular concern in these ecosystems is the possibility that large, stand-replacing wildfires will remove viable seed sources over large areas, significantly delaying or preventing post-fire forest recovery (Stephens et al. 2013) and converting forested areas into a qualitatively different vegetation type. However, given that nearly all wildfires include patches of stand-replacing fire interspersed with low- to- moderate severity patches (Turner and Romme 1994; Baker et al. 2007; Odion et al. 2014), an alternative scenario is that spatial heterogeneity in fire effects will allow for forest recovery and resilience to fire (Halofsky et al. 2011).
Forest recovery, or conversely, type-conversion, after large mixed-severity wildfires depends upon a combination of factors, including spatial variability in fire effects, pre-fire species composition, species-specific persistence mechanisms, and post-fire abiotic (e.g., topography, climate) and biotic (e.g., canopy opening, competition) conditions (Stephens et al. 2013). In forests where fire-resilient and fire-resistant (Baker 2009; Keeley et al. 2011) species co-occur, like dry mixed-conifer forests in the U.S. northern Rockies, the diversity in species and stand structure may increase forest resilience to variable fire effects (Halofsky et al. 2011). Fire-resistant species, such as Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) and ponderosa pine (Pinus ponderosa Lawson & C. Lawson), have thick bark and high crown base heights that facilitate survival after low-intensity surface fires. Where mature trees survive, successful regeneration often occurs. Resprouting species, such as quaking aspen (Populus tremuloides Michx.), or serotinous species, such as lodgepole pine (Pinus contorta Douglas ex Loudon), however, are more likely to regenerate after stand-replacing fire (Turner et al. 1997; Franklin and Bergman 2011; McKenzie and Tinker 2012). Understanding how different landscape patterns of mixed-severity fires impact forest structure and regeneration can be particularly challenging, however, in part because of high variability in pre- and post-fire conditions and variability within and among fire events.
Studies of conifer regeneration from single mixed-severity fire events (e.g., Lentile et al. 2005; Donato et al. 2009; Crotteau et al. 2013) or small geographic regions (e.g., Shatford et al. 2007; Collins and Roller 2013), have helped highlight the singular importance of burn severity, patch size, or abiotic conditions to post-fire regeneration. It remains unclear, however, how these results scale up to multiple fire events across broad regions. Here, we quantified natural post-fire tree seedling regeneration in 21 large (>400 ha), individual mixed-severity fire events across a 21,000 km2 region, spanning most of the range of dry mixed-conifer forests in the U.S. northern Rockies. We combined field data with statistical modeling to quantify the relationships among post-fire seedling abundance and composition, wildfire patch metrics, and abiotic and biotic variables. The extent of our study gives us the unique opportunity to identify important mechanisms that drive patterns of forest recovery at broad scales and infer the resilience of dry mixed-conifer forests to future large fires and climate change.
Methods
Study region
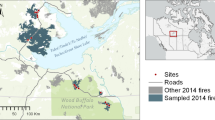
Our study region encompasses the range of dry mixed-conifer forests in the U.S. northern Rockies, spanning a four-degree south to north latitudinal gradient (Fig. 1). Dry mixed-conifer forests in this region are dominated by Douglas-fir and varying proportions of ponderosa pine, grand fir (Abies grandis (Douglas ex D. Don) Lindl.), and lodgepole pine. Across the study region, average temperatures range from 13.8 to 21.1 °C in July and −6.8 °C to −0.6 °C in January, and total precipitation ranges from 398 to 886 mm (PRISM 2014). The study region experiences warm, dry summers and wet winters. Soils are dominantly inceptisols and entisols derived from granitic Idaho batholith parent material (USDA 2014).

We sampled 182 sites in 21 individual fire events that burned in either 2000 or 2007 across central Idaho and western Montana. We stratified sites across gradients in elevation, aspect, and burn severity to represent the full range of climates in dry mixed-conifer forests (grey shading). Climate is defined here by the ratio of actual to potential evapotranspiration (Online Appendix A), shown below the map with boxplots for dry mixed-conifer forests (grey) and our sample sites (white). Boxplots delineate the 25th, 50th, and 75th percentiles and whiskers correspond to the 10th and 90th percentiles of the data.
Terrain is characterized by steep topography, with dramatic ecotone boundaries and steep elevation gradients that encompass multiple biomes from river valleys to ridgetops. South-facing hillslopes at low elevations are dominated by ponderosa pine with some Douglas-fir, while north-facing slopes at the same elevation can maintain a mix of ponderosa pine, Douglas-fir, grand fir, and lodgepole pine. Low elevation dry mixed-conifer forests dominated by ponderosa pine historically experienced surface fires with return intervals of years to a few decades (Heyerdahl et al. 2008a, b), while higher elevations and more mesic sites experienced less frequent, mixed- or even high-severity fires (Arno et al. 2000; Baker et al. 2007; Odion et al. 2014).
Sampling design
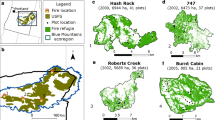
In the summer of 2012 and 2013, we sampled a total of 182 sites, stratified across the range of biotic and abiotic gradients characterizing dry mixed-conifer forests of the region. To identify potential sampling sites, we used a geographic information system (ArcGIS 10.0) to randomly select points within defined elevation, aspect, and burn severity classifications, in large (>400 ha) fires that burned in 2000 and 2007 (Gibson et al. 2014, Morgan et al. 2014). Thirty-meter resolution vegetation data from the LANDFIRE (2010) database and 10-m resolution digital elevations models (DEMs) were used to characterize forested regions within Idaho and western Montana. We used Relative differenced Normalized Burn Ratios (RdNBR; Miller and Thode 2007) derived from 30-m Landsat TM+ satellite imagery from the Monitoring Trends in Burn Severity (MTBS 2011) project to initially classify burn severity into four categories: (1) unburned-unchanged (2) low severity, (3) moderate severity, and (4) high severity. Sites were classified as “unburned” if they did not burn in either 2000, 2007, or in the 28- or 29- years covered by the MTBS data (i.e., since 1984). Within all four burn severity classes, we selected potential sites across three equally distributed elevation bands, with northeast- or southwest-facing aspects. Additional criteria for potential sites included a minimum polygon size of 0.81 ha (3 × 3 30-m pixels) to account for imprecision in satellite-derived data, separation of at least 120 m from other sites to minimize the potential for spatial autocorrelation, and proximity (within 2.5 km) to roads or wilderness research stations to maintain accessibility. Fifteen percent of sites were within federally designated wilderness areas. In all cases, sites that had been salvage logged or planted post fire were excluded from sampling based on communication with local USDA Forest Service personnel. Our stratification resulted in sample sites which spanned the full range of climates characterizing dry mixed-conifer forests in the region, as represented by the ratio of actual to potential evapotranspiration (Online Appendix 1; Fig. 1).
At each site, burn severity was field verified using environmental evidence including estimates of percent tree mortality, bole scorch, and shrub stem mortality. If a site did not fall within the desired burn severity classification, or was inaccessible, the location was offset by 30 m in cardinal directions until the desired stratification was attained. Live tree seedlings were counted within a 60-m long belt transect of variable width of 1–10 m, with transect width determined prior to sampling each plot based on visual estimates of seedling density. Vegetation cover, overstory tree basal area, and tree canopy cover were measured at 0, 30, and 60 m along each transect and averaged for a site. Vegetation cover was classified by lifeform (shrub, forb, graminoid, tree) within 1 m2 sub-quadrats. Overstory tree basal area was quantified using a 2 or 4 m2 ha−1 basal area factor prism on a variable radius plot, and canopy cover was recorded using a spherical crown densiometer (Forestry Suppliers, Inc., No. 43887). Overstory tree species, diameter at 1.37 m height, and percent mortality were also recorded and used to calculate the density of live and dead trees for each species. We quantified the distance to a live seed source by measuring the horizontal distance to the ten nearest live seed trees of all species from the transect center using a laser range finder (Truepulse® 360 B/Laser Technology) and averaging these distances for a site. Seed trees were confirmed to either have cones or be large enough to be reproductively mature. Distances greater than 500 m could not be measured. Additional site level data included slope, aspect, latitude, longitude, and elevation. Slope, aspect, and latitude were used to estimate potential heat load from direct solar radiation by calculating a heat load index (following McCune and Keon 2002).
Statistical analysis
We used a two-stage modeling approach to examine patterns of natural seedling regeneration as a function of three categories of response variables: legacies of the fire (e.g., burn severity, distance to live seed source), abiotic environmental variables (e.g., elevation, heat load), and biotic environmental variables (e.g., vegetation cover, tree basal area, and tree canopy cover; Table 1). We first used a logistic regression model to predict tree seedling presence or absence and then used a negative binomial or zero-inflated negative binomial model to predict tree seedling abundance (count). Logistic and count models were compared to evaluate whether the processes influencing seedling presence and abundance differed. All analyses were completed in R version 3.0.2 (R Core Team 2013) using the “MASS” package (Venables and Ripley 2002) for the negative binomial models and the “pscl” package (Zeileis et al. 2008; Jackman 2012) for the zero-inflated models. The logistic regression models are part of the standard statistical package in R.
Logistic regression models for seedling presence–absence
Five logistic regression models were developed using a binomial distribution to predict the presence or absence of tree seedlings. These models included a model for all species present on the site, and four species-specific models for species present on >15 % of the sites: Douglas-fir, ponderosa pine, lodgepole pine, and grand fir. All predictor variables were tested for collinearity using Spearman’s rank correlation. Tree canopy cover was dropped from each model because it was well correlated (ρ > 0.5) with overstory tree basal area and the average distance to a live seed source. Models were then constructed using the remaining predictor variables and a censor variable indicating whether seed sources were present and measured on a site (Table 1). We examined the sensitivity of the parameter estimates (β) using forward–backward stepwise selection with AIC selection criteria and determined there was little change in these estimates due to model reduction. Therefore, we kept the fully specified models with all variables for comparative purposes.
We assessed logistic model fit using the three summary measures: the deviance residual, a Hosmer–Lemeshow test, and the area under the curve (AUC) of a receiver operating characteristic (ROC) curve (Online Appendix 1). The AUC for each model can vary from 0.5 (random) to 1.0 (perfect prediction), where 0.7–0.8 is “acceptable”, 0.8–0.9 is “excellent”, and 0.9–1 is “outstanding” discrimination between the model predictions and the observed data (Fawcett 2006; Hosmer et al. 2013). Additionally, to discriminate between presences and absences based on the modeled probabilities, we used a classifier value calculated from the ROC curve that maximized the rate of true positives while minimizing false positives for each model (Fawcett 2006). Using this classifier value, we calculated the positive predictive rate (PPR; i.e., the ratio of true positives to the sum of true and false positives) and the negative predictive rate (NPR; i.e., the ratio of true negatives to total negatives) to evaluate how well models predicted the actual presence or absence of tree seedlings across our study sites. To avoid model overfitting, we subsequently used cross-validation techniques to evaluate the Hosmer–Lemeshow statistic and AUC for each model (Online Appendix 1; Hosmer et al. 2013).
Count models for regeneration abundance
Our tree seedling count data had a high proportion of zeros (26 % for all species combined; 35, 68, 74, and 86 % for Douglas-fir, ponderosa pine, lodgepole pine, and grand fir, respectively) and the distribution of seedling counts was strongly left-skewed. Therefore, we considered a number of alternative generalized linear models for discrete skewed data, following the procedures outlined by Zeileis et al. (2008). Because our data displayed significant overdispersion (e.g., the variance was larger than the mean), we modeled the abundance of all species and the abundance of Douglas-fir using a generalized linear model with a negative binomial distribution. We developed zero-inflated negative binomial models for the abundance of ponderosa pine, lodgepole pine, and grand fir (Online Appendix 1; Martin et al. 2005; Zuur et al. 2009; Hilbe 2011).
To compare the variables influencing presence and abundance of tree seedling regeneration, we kept all of the predictor variables used in the logistic regression model in the count models. Zero-inflated models were parameterized with an intercept-only predictor function for the zero portion of the model (Online Appendix 1; Zuur et al. 2012). In addition, an offset variable was included in each count model to correct for the variable sampling area. Five sites were removed from this analysis because we failed to record the transect area (n = 177).
We compared predicted and observed values from each of the count models using Spearman’s rank correlation. Model fit was assessed visually by plotting the Pearson’s residuals against the fitted values (Zuur et al. 2009). Furthermore, we performed a goodness-of-fit test using Monte Carlo simulations (n = 1000) to calculate the Pearson’s chi-squared statistic, where the expected probabilities from the parameterized model are used to generate new observations set as the “observed” counts. These observed counts are then compared to the “expected” counts drawn from a random negative binomial distribution. A significant lack of fit is indicated by a p-value ≤ 0.05. We also calculated the Pearson’s chi-squared test statistic and p-value using cross-validation for each model (n = 1000).
Patch size analysis
To determine the proportion of the landscape burned at high severity (i.e., stand-replacing wildfire), we used the classified MTBS data for the 21 sampled fire events and calculated the mean distance from each pixel classified as high severity to the nearest pixel of lower severity (i.e., classified as moderate, low, or unburned). Distance calculations were made from the raster images of the individual fires within R using the “gdistance” (Etten 2014) and “rgal” (Bivand et al. 2014) packages. Because pixel size in the MTBS dataset is 30 m, the minimum distance from a high severity pixel to an edge was 30 m if the two pixels were adjacent. We merged all fires for a single year (2000 or 2007) into a single raster and calculated the cumulative proportion of high burn severity pixels that were various distances from a pixel of lower burn severity. These distributions were compared to the average distance from live seed source trees measured on the ground at each of our high burn severity sites (n = 61).
Results
We counted over 10,000 seedlings of eight different species on 182 sites across our study region. Douglas-fir was the most abundant species, present on 120 sites (66 %; Table 2). Total seedling densities ranged from 0 to 1.3 × 106 seedlings ha−1 and ranged four orders of magnitude for the most abundant individual tree species, from 0 to 3763 seedlings ha−1 (Fig. 2). Seedling densities did not vary significantly between the two fire years (Mann–Whitney U test: W = 4378, p = 0.456). Nine sites had no seed sources within 500 m (the maximum detectable distance), and of these nine sites, only one had seedlings present, all of which were lodgepole pine (Fig. 2). Conifer species composition did not vary significantly between trees present before fire and seedlings present after the fire, with the exception of a slight increase in lodgepole pine present in sites that burned in 2007, and a decrease in subalpine fir on sites where it was present and burned in 2007 (Table 2).

Tree seedling density as a function of the distance to a live seed source. Each site is additionally colored by burn severity. The maximum distance that could be measured was 500 m from the transect center. Sites with no live seed sources within 500 m are presented in the right panel
Models for Conifer Seedling Presence
All species model
Distance to a live seed source was the most important variable predicting seedling presence (β = −0.014, p = 0.002; Table 3), with a lower probability of presence with increasing distance (Fig. 3a). We identified an optimal classifier probability of 0.87 with the ROC analysis (Table 3), above which seedlings were predicted to be present. This probability threshold corresponded to a maximum distance of 95 m from a live seed source (Fig. 3a). Sites with a higher pre-fire tree basal area also had a significantly higher probability of seedling presence (β = 0.072, p = 0.008; Table 3), though this pattern was most apparent at sites with low basal areas. All sites with a pre-fire tree basal area of 5 m2 ha−1 had a high probability of seedling establishment, and above a basal area of 20 m2 ha−1 this probability was >95 %.

Logistic regression model results. Relationship between the probability of seedling presence and distance to a live seed source for (a) all species and (b) the four most abundant species, when all other variables in the model are held at their median values. The shaded region between the dotted lines represents the 95 % confidence intervals on the predicted values for the all species model. In panel (b), the dashed line indicates that the relationship between distance and seedling presence was not significant for that species (p > 0.05). Confidence intervals are not shown in panel (b) because they overlap for all species
Our model correctly predicted the proportion of sites with seedling presence 93 % of the time (PPR) and seedling absences (NPR) 39 % of the time. The model distribution fit the data well (p > 0.08; Table 4). An AUC value of 0.826 indicated an excellent ability to discriminate between sites with and without seedlings across our extensive sample region (Table 4), which was robust to cross-validation (Online Appendix 2).
Species-specific models
Distance to a live seed source was the most significant variable influencing the presence of Douglas-fir and ponderosa pine seedlings (Table 3). Seed source distance was marginally significant for grand-fir, and insignificant for lodgepole pine (Table 3). The further a site was from a live seed source, the lower the probability of Douglas-fir, ponderosa pine, or grand fir presence (Fig. 3b). Ponderosa pine seedlings were most likely to be present within 60 m of a live seed tree (Table 4). Similarly, Douglas-fir was most likely to be present within roughly 75 m of a live seed source. Grand fir presence was probable as far as 165 m from a live seed tree (Fig. 3b). The censored variable, indicating whether or not a live seed source was measured on the site, was significant in these models only for ponderosa pine, lodgepole pine, and grand fir, with the odds of having seedlings present on sites without seed sources between three and seven percent less than sites with seed sources. Furthermore, distance to a live seed source was the only variable that was important in predicting the presence of ponderosa pine and grand fir tree seedlings across our study region (Table 3).
Elevation and heat load index were also important in the species-specific models for Douglas-fir and lodgepole pine (Table 3). Douglas-fir and lodgepole pine tree seedlings were more likely to be present at higher elevations and on sites with a low heat load index (Table 3). Tree basal area was important in predicting the presence of Douglas-fir on a site but did not influence the presence of any of the other species (Table 3).
Each of the full and cross-validated species-specific regression models predicting tree seedling presence performed better than random, as indicated by median AUC values greater than 0.5 (Table 4; Online Appendix 2). Models correctly predicted the proportion of sites with seedling presence between 69 and 91 % of the time (PPR), and correctly predicted the proportion of sites with seedlings absent (NPR) between 47 and 91 % of the time (Table 4). Deviance chi-squared and Hosmer–Lemeshow statistics validated the fit of our data to a binomial distribution (p > 0.05), and were confirmed by cross-validation for most models (Online Appendix 2).
Models for conifer seedling abundance
All species model
Distance to the nearest live seed source (β = −0.007; p ≪ 0.001) and tree basal area (β = 0.032; p = 0.016) remained the most important predictors of seedling abundance in the negative binomial model (Table 3), with the same directionality whether predicting seedling presence or seedling abundance. There was significant fit between the data for our full all-species model and the negative binomial model distribution (Table 4), though this model cross validated poorly (Online Appendix 2). Some variability between the predicted estimates and the observed seedling counts was present in the model (Table 4). Though the negative binomial all-species model tended to overpredict the abundance of seedlings on sites, forty-five percent of the observed seedling counts fell within the range of predicted counts, +/− two standard errors (Online Appendix 2; Fig. A2.1).
Species-specific models
While the variables determining abundance of tree seedlings in the all species model and the ponderosa pine model did not change from the corresponding models predicting seedling presence, additional variables helped predict the abundance of Douglas-fir and grand fir across the study region (Table 3). For example, Douglas-fir abundance was significantly higher on sites with a longer time since fire (Table 3). Burn severity was also significant for Douglas-fir abundance; the expected counts of Douglas-fir seedlings were 8 and 13 times higher in sites burned at low and moderate severity, respectively, than in unburned sites. Although elevation and heat load influenced lodgepole pine presence, these variables were not important for determining its abundance (Table 3). Grand fir abundance was negatively related to both vegetation cover and tree basal area and distance to a live seed source was not important for its abundance on a site (Table 3).
The regeneration count models performed moderately well. Correlation between the predicted and observed counts ranged from 0.4 to 0.7 (Table 4). The negative binomial models correctly predicted 41–50 % and overpredicted between 31 and 60 % of observations for each species. A negative binomial distribution and a zero-inflated negative binomial distribution fit the observed data for all the full species-specific models except ponderosa pine, as indicated by the Pearson’s chi-squared goodness-of-fit tests (Table 4). In general, the species-specific count models did not cross-validate as well as the logistic regression models (Online Appendix 2).
Patch size and distance to seed source
Burn severity was not a significant factor influencing seedling presence or abundance for any of the species except Douglas-fir. Rather, distance to a live seed source overrode burn severity in the models. Specifically, sites within patches burned at high and moderate severity were further from live seed source trees than sites in either low or unburned patches (Fig. 4; Kruskal–Wallis Test: χ2 = 117.809, d.f. = 3, p << 0.001). Sites that burned at moderate severity were a median distance of 31 m from a live seed source, while sites that burned at high severity were a median distance of 122 m from a live seed source, compared to 12 and 13 m for unburned and low severity sites, respectively.

Relationship between burn severity and the average distance to a live seed source. Burn severity was initially categorized using satellite RdNBR data and subsequently field verified. Letters denote to statistical differences between the treatments. Patches burned at high severity have significantly further average distances to live seed trees than moderate, low, or unburned patches. Patches burned at low severity did not differ from unburned patches in the average distance to a live seed source
Patches burned at higher severity were characterized by high tree mortality, though they tended to be adjacent to patches with only partial tree mortality. The mean distance from an area within a patch burned at high severity to an edge of an unburned or low severity patch ranged from 33 to 118 m in fires that burned in 2000 and from 31 to 122 m in fires that burned in 2007. Over 85 % of the area burned in 2000 in high severity patches was less than 95 m from the nearest edge of a lower severity patch and 98 % of the area was within 200 m from an edge (Online Appendix 2; Fig. A2.2). Likewise, in the 2007 fire events, 75 % of the area burned by high severity fire was within 95 m of an edge and 94 % of the area was within 200 m of an edge burned with lower severity (Online Appendix 2; Fig. A2.2). The distribution of edge distances characterized from the satellite data is consistent with the distribution of our on-the-ground measurements of distance to seed source (Online Appendix 2; Fig. A2.2).
Discussion
Resilience of forests to large, severe wildfires is of ecological and management significance, particularly given ongoing climate change and the potential implications of increased forest density from prior forest and fire management (Keeling et al. 2006; Naficy et al. 2010). Our results highlight important interactions between the spatial distribution of patches burned at high severity and seed dispersal mechanism as the primary controls of post-fire regeneration. Abundant seedling regeneration and the presence of few large high severity patches across our study region suggest that successful reestablishment is widespread and these dry-mixed conifer forests will be resilient to recent large fires. Furthermore, our results have important implications for addressing the impacts of shifting fire regimes and climate change on current and future forest persistence.
Dispersal distance is a primary control on post-fire regeneration
In dry mixed-conifer forests of the U.S. northern Rockies, post-fire regeneration is strongly controlled by landscape structural characteristics resulting from burn patterns. Distance to a live seed source was the primary limitation on post-fire regeneration for all the species across our study region except lodgepole pine, where the presence of serotinous cones allowed for regeneration in the absence of live trees. Burned areas without nearby residual live seed trees had few or no conifer seedlings 5–13 years after fire, regardless of the severity with which the patch burned. Observed post-fire seedling regeneration of Douglas-fir, ponderosa pine, and grand fir mimicked the expected dispersal curves of wind-dispersed species, where the number of viable seeds deposited decreases exponentially with distance from the patch edge (Greene and Johnson 1996). This observed pattern suggests that dispersal limitation is more significant than density-dependent mortality in determining post-fire recruitment across our broad study region (Greene and Johnson 2000).
Our study adds to a growing body of literature emphasizing the importance of nearby live seed sources for post-fire regeneration (e.g., Keyser et al. 2008; Donato et al. 2009; Haire and McGarigal 2010). We identified a 95 m threshold from residual live seed sources for tree seedling establishment to occur, integrating differences in burn severity, abiotic and biotic environmental conditions among all 182 sites across our large study region. This threshold falls within the range of dispersal distances documented for many species found on our sites, which can range anywhere from 20 m up to 180 m for ponderosa pine and Douglas-fir seeds to between 40 and 120 m for the majority of grand fir seeds (McCaughey et al. 1986, Vander Wall 2003).
Because dispersal distance acts as a primary filter on post-fire conifer regeneration, the size and spatial configuration of stand-replacing patches across the landscape become key drivers of post-fire successional trajectories (e.g., Haire and McGarigal 2010). Even within the large wildfires we sampled, more than 75 % of the area within patches burned by stand-replacing fire was less than 95 m from an edge, implying that the majority of burned area was close to live seed sources and thus had a high probability of successful natural regeneration. This finding is corroborated by several prior studies from diverse forest types; for example, 75 % of stand-replacement patches in subalpine forests in Yellowstone National Park were less than 200 m from a live forest edge (Turner et al. 1994), and 58 % of stand-replacement patches in a mixed-conifer forest in southern Oregon were within 200 m of a live forest edge (Donato et al. 2009). Thus, even within large fires, only a small proportion of the entire burned area, including patches burned with high severity, are far enough from seed sources to limit successful natural regeneration. This diversity in burned patch sizes creates landscapes that are largely resilient to mixed-severity fires, regardless of burning conditions or forest type.
Secondary controls on seedling regeneration
Where seed sources were available, seedling regeneration was highly variable, suggesting that seed source is a necessary but insufficient explanation of seedling abundance. Abundant regeneration depends upon successful germination, survival, and growth, all of which are influenced by stochastic processes and environmental conditions that vary at fine scales (e.g., Bonnet et al. 2005). Tree basal area was the primary biotic environmental variable influencing seedling abundance in our study. Basal area can be a proxy for site productivity as it measures both size and density of trees in a stand. Gradients in resource availability that make pre-fire stands productive, such as soil fertility and moisture availability, likely also influence post-fire germination and tree seedling survival (e.g., Clarke et al. 2005; Röder et al. 2008; Casady et al. 2010). Relative to distance to seed source, however, our results suggest only a minor influence of tree basal area on tree seedling regeneration, as all sites with a basal area <5 m2 ha−1 had a relatively high probability of tree seedling establishment, indicating that where trees occurred prior to fire, they are likely to regenerate post-fire. Marginal sites with few trees prior to the fire (those with a basal area <5 m2 ha−1) are less favorable for reestablishment. Stochastic processes that we did not quantify, such as variability in seed crops, microsite conditions, or favorable post-fire climate conditions (Brown and Wu 2005; League and Veblen 2006), could also account for some of the unexplained variability in seedling abundance across our study region. Seed masting events occur on average every 3–12 years in Douglas-fir and ponderosa pine stands in the U.S. northern Rockies (USFS 2012), for example, strongly limiting seed availability in intervening years. Likewise, seed predation can limit successful germination and subsequent tree seedling regeneration (Vander Wall 1994; Zwolak et al. 2010; Lobo 2014). Given that the post-fire seedling recruitment period can be an important stage within long-term forest succession, further research examining the influence of stochastic variables, especially weather and climate, on post-fire regeneration, will be important for understanding the potential implications of shifts in climate on longer-term forest dynamics.
The importance of environmental gradients was most pronounced for Douglas-fir regeneration. Moderate- and low-severity burns had significantly more Douglas-fir seedlings than unburned sites, suggesting that moderate increases in resource availability, such as increased light, nutrients, or mineral soil, favored seedling establishment and survival (e.g., York et al. 2003, Moghaddas et al. 2008). Recruitment was also more abundant on sites characterized by higher average elevation and lower heat load, representing cooler, wetter locations. High summer temperatures and water availability limit Douglas-fir growth across its range in the northern Rockies (Littell et al. 2008), and our data suggest that these factors also limit seedling establishment and survival. Douglas-fir abundance was also higher on sites with a longer time since fire. For a sporadic seed producing species like Douglas-fir, this is expected, as the probability of successful establishment accumulates over time. However, we caution the extrapolation of these results to sites outside of our study region given that this model did not cross-validate well, nor was this relationship significant for the other species examined. Further, this result suggests that regeneration for the other species we examined may have been pulsed in a single event post-fire, a result of seedling mortality over time, or due to our sample period not being long enough to pick up multiple pulses of successful post-fire establishment.
Post-fire tree seedling composition reflects pre-fire stand composition
Our species composition data show relatively little difference between the species composition of regenerating seedlings and the pre-fire mature tree composition. This suggests that the mix of burn severities, patch sizes, and environmental conditions across the landscape perpetuated the forest conditions that were present prior to fire. At the site scale, post-fire seedling composition may still vary as a function of pre-fire species composition of the live tree edge and species-specific tree regeneration mechanisms. For example, the abundance of seeds reaching the interior of patches depends upon a tree species’ dominance on the intact forest edge (Greene and Johnson 2004), where edge dominance increases the seed rain of that particular species (e.g., Greene and Johnson 1996). Infilling by shade tolerant species in the absence of fire could therefore alter the seed rain available to recolonize a patch post-disturbance (Perry et al. 2011), especially if those species are prolific seed producers. These factors, in combination with our model predictions of further dispersal distances for grand fir (i.e., 2–3 times the distance of Douglas-fir and ponderosa pine) and additional empirical evidence (McCaughey et al. 1986), suggest that grand fir has the potential to recolonize larger burned patches and increase in dominance post-fire (e.g., Crotteau et al. 2013). This effect may be especially pronounced in areas that have reduced tree cover of Douglas-fir and ponderosa pine from prior logging.
Species-specific regeneration mechanisms may also determine which dry mixed-conifer tree species recolonize different patches post-fire. Lodgepole pine, for example, can regenerate in the absence of a live seed source where it has serotinous cones stored in an aerial seed bank. Though serotiny can vary considerably across a landscape (Schoennagel et al. 2003), forests with high pre-fire serotiny can have prolific regeneration in stand-replacing patches (Turner et al. 1997; Schoennagel et al. 2003). Compared to the other dry mixed-conifer species we studied, lodgepole pine recruitment is likely to be favored in large high severity (i.e., stand-replacing) patches where live seed sources are limited.
Implications for forest resilience
Our results suggest that the spatial characteristics of mixed-severity fires and pre-fire species composition interact to promote resilience of dry mixed-conifer forests to large wildfires, even when those fires burn under a variety of weather conditions. Most of the area within patches burned by high-severity (i.e., stand-replacing) fire in the large regional fire events we studied were close to live trees which likely included seed sources for conifer regeneration. Over 80 % of the patches burned at low severity and approximately 40 % of patches burned at moderate severity that we sampled exceeded the tree seedling densities considered sufficient to regenerate a stand to its pre-fire density, which can range from 180 trees ha−1 to 370 trees ha−1 in dry mixed-conifer forests across the northern Rockies region (S. Fox, USFS; pers. comm.). Sparse natural tree regeneration was primarily observed in large, high severity patches (i.e., those patches with interiors >95 m from an edge and few surviving trees). Although burn severity was a poor predictor of ecological response in our study, it is directly related to dispersal distance, as by definition high severity patches have fewer residual live trees and are further from live seed sources. Therefore, the high heterogeneity of patch types and sizes within a fire (i.e., the burn mosaic) is key to maintaining current forest diversity and structure after future wildfires.
Although our seedling regeneration data represent a limited snapshot in time, mortality of seedlings has been shown to decline markedly and remain constant after the year of germination (Pausas et al. 2002, Calvo et al. 2013). Seedlings that establish and survive the first year of growth are likely to remain an important feature of long-term forest structure. In our study, 94 % (n = 2226) and 72 % (n = 2518) of the seedlings that we sampled that regenerated after the 2000 and 2007 fires, respectively, were greater than 1 year old. Long-term studies of Douglas-fir forests in the western Cascades, USA, indicate that density dependent mortality does occur, especially with canopy closure, after about 25–32 years post disturbance. However, decreases in stem density are offset by changes in biomass, where biomass loss is maintained at a relatively low and constant level (Lutz and Halpern 2006). Although continued seedling recruitment and mortality is likely to occur in the stands we sampled, the combination of high seedling densities and large proportions of well established seedlings suggests that if current conditions remain stable, these seedlings are likely to persist.
Although there is considerable concern surrounding large fire events, patch scale heterogeneity present across the burned landscape in the U.S. northern Rockies suggests that these forests will recover to pre-disturbance species composition and diversity. Our conclusion is predicated upon several factors: significant seedling mortality does not occur, forests remain unburned long enough for live trees to reach reproductive maturity, the proportion of high-severity patches far from seed sources does not increase significantly, and climate change does not shift post-fire environmental conditions so as to limit successful tree seedling establishment and growth. The range of suitable climate conditions for growth of mature tree species in the U.S. northern Rockies may shift considerably in the coming decades (Rehfeldt et al. 2006; Rehfeldt et al. 2008), and it is likely that the regeneration niche of seedlings is even narrower (Grubb 1977; Jackson et al. 2009). Additionally, extrapolations of statistical fire-climate relationships suggest a potential two to five-fold increase in the median area burned in the U.S. northern Rockies by mid-century (Littell 2011), implying an increase not only in frequency, but also fire size. Short fire-return intervals may limit the potential for tree regeneration success and remove future seed sources (Keeley et al. 1999; Johnstone and Chapin 2006; Brown and Johnstone 2012). More intense and severe fire may also favor tree species that are well adapted to regenerate in the absence of live seed sources nearby (e.g., lodgepole pine and grand fir).
Understanding how large, mixed-severity fires impact the regeneration and resilience of forests will become increasingly important for making sound forest and fire management decisions in a warmer, more fire prone future. Conifer regeneration and habitat restoration are important management priorities following fire, and abundant natural post-fire tree regeneration may limit the area managers need to treat to meet these objectives. In large, high-severity patches, sparse natural regeneration may result in delayed successional trajectories or altered vegetation states. Managers aiming to insure post-fire recovery should therefore focus regeneration efforts on areas within high-severity patches that are far (>100 m) from live seed sources. As area burned continues to increase, the amount of area burned severely will also increase (Dillon et al. 2011). If the size of high-severity patches and the relative proportions they occupy on the burned landscape increases with future climate change, or if post-fire environmental conditions shift significantly relative to the past several decades (Rehfeldt et al. 2006), the resilience of dry mixed-conifer forests to large wildfires that we documented will be increasingly compromised.
References
Arno SF, Parsons DJ, Keane RE (2000) Mixed-severity fire regimes in the northern Rocky Mountains: consequences of fire exclusion and options for the future. In: Cole D, McCool S, Borrie W, O’Laughlin J (eds) Wilderness science in a time of change, Missoula, MT. 1999. vol 5: Wilderness ecosystems, threats, and management. USDA Forest Service, Rocky Mountain Research Station, pp 225-232
Baker WL (2009) Fire Ecology in Rocky Mountain Landscapes. Island Press, United States
Baker WL, Veblen TT, Sherriff RL (2007) Fire, fuels and restoration of ponderosa pine-Douglas fir forests in the Rocky Mountains, USA. J Biogeogr 34:251–269
Bivand R, Keitt T, Rowlingson B (2014) rgdal: Bindings for the Geospatial Data Abstraction Library. R package version 0.8-16. Available from http://CRAN.R-project.org/package=rgdal
Bonnet VH, Schoettle AW, Shepperd WD (2005) Postfire environmental conditions influence the spatial pattern of regeneration for Pinus ponderosa. Can J For Res 35(1):37–47
Brown CD, Johnstone JF (2012) Once burned, twice shy: Repeat fires reduce seed availability and alter substrate constraints on Picea mariana regeneration. For Ecol Manage 266:34–41
Brown PM, Wu R (2005) Climate and disturbance forcing of episodic tree recruitment in a southwestern ponderosa pine landscape. Ecology 86(11):3030–3038
Calvo L, Torres O, Valbuena L, Luis-Calabuig E (2013) Short Communication. Recruitment and early growth of Pinus pinaster seedlings over five years after a wildfire in NW Spain. Forest Systems 22(3):582–586
Casady GM, van Leeuwen WJ, Marsh SE (2010) Evaluating post-wildfire vegetation regeneration as a response to multiple environmental determinants. Environ Model Assess 15(5):295–307
Clarke PJ, Knox KJE, Wills KE, Campbell M (2005) Landscape patterns of woody plant response to crown fire: disturbance and productivity influence sprouting ability. J Ecol 93(3):544–555
Collins BM, Roller GB (2013) Early forest dynamics in stand-replacing fire patches in the northern Sierra Nevada, California. USA. Landscape Ecol 28(9):1801–1813
Crotteau JS, Varner M III, Ritchie MW (2013) Post-fire regeneration across a fire severity gradient in the southern Cascades. For Ecol Manage 287:103–112
Dillon GK, Holden ZA, Morgan P, Crimmins MA, Heyerdahl EK, Luce CH (2011) Both topography and climate affected forest and woodland burn severity in two regions of the western US, 1984 to 2006. Ecosphere 2(12):art130
Donato DC, Fontaine JB, Campbell JL, Robinson WD, Kauffman JB, Law BE (2009) Conifer regeneration in stand-replacement portions of a large mixed-severity wildfire in the Klamath-Siskiyou Mountains. Can J For Res 39(4):823–838
Fawcett T (2006) An introduction to ROC analysis. Pattern Recog Lett 27(8):861–874
Flannigan MD, Krawchuk MA, de Groot WJ, Wotton BM, Gowman LM (2009) Implications of changing climate for global wildland fire. Int J Wildland Fire 18(5):483–507
Franklin J, Bergman E (2011) Patterns of pine regeneration following a large, severe wildfire in the mountains of southern California. Can J For Res 41(4):810–821
Gibson CE, Morgan P, Wilson AM (2014) Atlas of digital polygon fire extents for Idaho and western Montana. 2nd edn. Forest Service Research Data Archive, Fort Collins, CO. DOI: 10.2737/RDS-2009-0006-2
Greene DF, Johnson EA (1996) Wind dispersal of seeds from a forest into a clearing. Ecology 77(2):595–609
Greene DF, Johnson EA (2000) Tree recruitment from burn edges. Can J For Res 30(8):1264–1274
Groffman PM, Baron JS, Blett T et al (2006) Ecological Thresholds: The key to sucessful environmental management or an important concept with no practical application? Ecosystems 9:1–13
Grubb PJ (1977) The maintenance of species-richness in plant communities: the importance of the regeneration niche. Biol Reviews 52(1):107–145
Haire SL, McGarigal K (2010) Effect of landscape patterns of fire severity on regenerating ponderosa pine forests (Pinus ponderosa) in New Mexico and Arizona, USA. Landscape Ecol 25:1055–1069
Halofsky JE, Donato DC, Hibbs DE et al (2011) Mixed-severity fire regimes: lessons and hypotheses from the Klamath-Siskiyou Ecoregion. Ecosphere 2(4):art40
Hessburg PF, Smith BG, Salter RB, Ottmar RD, Alvarado E (2000) Recent changes (1930s–1990s) in spatial patterns of interior northwest forests. USA. For Ecol Manage 136(1–3):53–83
Heyerdahl EK, McKenzie D, Daniels LD, Hessl AE, Littell JS, Mantua NJ (2008a) Climate drivers of regionally synchronous fires in the inland Northwest (1651–1900). Int J Wildland Fire 17(1):40–49
Heyerdahl EK, Morgan P, Riser JP (2008b) Multi-season climate synchronized historical fires in dry forests (1650-1900), Northern Rockies. USA. Ecology 89(3):705–716
Hilbe JM (2011) Negative Binomial Regression. Cambridge University Press, Cambridge, U.K.
Holling CS (1973) Resilience and stability of ecological systems. Annu Rev Ecol Syst 4:1–23
Hosmer DW, Lemeshow S, Sturdivant RX (2013) Applied Logistic Regression. John Wiley & Sons, Hoboken, New Jersey
Jackman S (2012) pscl: Classes and Methods for R Developed in the Political Science Computational Labratory. Department of Political Science, Stanford University, Stanford, California. Available from http://cran.r-project.org/web/packages/pscl/pscl.pdf
Johnstone JF, Chapin FS III (2006) Fire interval effects on successional trajectory in boreal forests of Northwest Canada. Ecosystems 9(2):268–277
Johnstone JF, Hollingsworth TN, Chapin FS, Mack MC (2010a) Changes in fire regime break the legacy lock on successional trajectories in Alaskan boreal forest. Global Change Biol 16(4):1281–1295
Johnstone JF, McIntire EJB, Pedersen EJ, King G, Pisaric MJF (2010b) A sensitive slope: estimating landscape patterns of forest resilience in a changing climate. Ecosphere 1(6):art14
Kasischke ES, Turetsky MR (2006) Recent changes in the fire regime across the North American boreal region—Spatial and temporal patterns of burning across Canada and Alaska. Geophys Res Lett 33(9):L09703
Keeley JE, Ne’eman G, Fotheringham C (1999) Immaturity risk in a fire-dependent pine. J of Mediterranean Ecol 1:41–48
Keeley JE, Pausas JG, Rundel PW, Bond WJ, Bradstock RA (2011) Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci 16(8):406–411
Keeling EG, Sala A, DeLuca TH (2006) Effects of fire exclusion on forest structure and composition in unlogged ponderosa pine/Douglas-fir forests. For Ecol Manage 237(1–3):418–428
Keyser TL, Lentile LB, Smith FW, Shepperd WD (2008) Changes in forest structure after a large, mixed-severity wildfire in ponderosa pine forests of the Black Hills, South Dakota. USA. For Sci 54(3):328–338
LANDFIRE (2010) Existing vegetation type layer. U.S. Geological Survey, Department of Interior. Available from http://landfire.cr.usgs.gov/viewer/ (accessed 30 March 2012)
League K, Veblen T (2006) Climatic variability and episodic Pinus ponderosa establishment along the forest-grassland ecotones of Colorado. For Ecol Manage 228(1–3):98–107
Lentile LB, Smith FW, Sheppard WD (2005) Patch structure, fire-scar formation, and tree regeneration in a large mixed-severity fire in the South Dakota Black Hills, USA. Can J For Res 35:2875–2885
Littell JS (2011) Impacts in the next few decades and the next century: fire and climate. In: Council NR (ed) Climate Stabilization Targets: Emissions, Concentrations, and Impacts over Decades to Millennia. The National Academies Press, Washington, D.C., pp 178–180
Littell JS, Peterson DL, Tjoelker M (2008) Douglas-fir growth in mountain ecosystems: water limits tree growth from stand to region. Ecol Monogr 78(3):349–368
Lobo N (2014) Conifer seed predation by terrestrial small mammals: A review of the patterns, implications, and limitations of top-down and bottom-up interactions. For Ecol Manage 328:45–54
Lutz JA, Halpern CB (2006) Tree mortality during early forest development: a long-term study of rates, causes, and consequences. Ecol Monogr 76(2):257–275
McCaughey WW, Schmidt WC, Shearer RC (1986) Seed dispersal characteristics of conifers in the inland mountain West. In: Shearer RC (ed) Conifer tree seed in the inland mountain West, Missoula, MT. USFS Gen Tech Rep INT-023. Intermountain Research Station, pp 50–62
McCune B, Keon D (2002) Equations for potential annual direct incident radiation and heat load. J of Veg Sci 13:603–606
McKenzie D, Tinker D (2012) Fire-induced shifts in overstory tree species composition and associated understory plant composition in Glacier National Park, Montana. Plant Ecol:1-18
Miller JD, Thode AE (2007) Quantifying burn severity in a heterogeneous landscape with a relative version of the delta Normalized Burn Ratio (dNBR). Remote Sens Environ 109(1):66–80
Moghaddas JJ, York RA, Stephens SL (2008) Initial response of conifer and California black oak seedlings following fuel reduction activities in a Sierra Nevada mixed conifer forest. For Ecol Manage 255(8–9):3141–3150
Morgan P, Heyerdahl EK, Miller C, Wilson AM, Gibson CE (2014) Northern Rockies pyrogeography: An example of fire atlas utility. Fire Ecol 10(1):14–30
Moritz MA, Hessburg PF, Povak NA (2011) Native Fire Regimes and Landscape Resilience. In: McKenzie D, Miller C, Falk DA (eds) The Landscape Ecology of Fire. Springer Science + Business Media B.V, New York, NY, pp 51–86
MTBS (2011) Monitoring Trends in Burn Severity Project Data Access. U.S. Geologic Survey, Department of the Interior. Available from www.mtbs.gov/dataaccess.html (accessed 30 March 2012)
Naficy C, Sala A, Keeling EG, Graham J, DeLuca TH (2010) Interactive effects of historical logging and fire exclusion on ponderosa pine forest structure in the northern Rockies. Ecol Appl 20(7):1851–1864
Odion DC, Hanson CT, Arsenault A et al (2014) Examining historical and current mixed-severity fire regimes in ponderosa pine and mixed-conifer forests of western North America. PLoS ONE 9(2):e87852
Odion DC, Moritz MA, DellaSala DA (2010) Alternative community states maintained by fire in the Klamath Mountains. USA. J Ecol 98(1):96–105
Pausas JG, Ouadah N, Ferran A, Gimeno T, Vallejo R (2002) Fire severity and seedling establishment in Pinus halepensis woodlands, eastern Iberian Peninsula. Plant Ecol 169(2):205–213
Pausas J, Fernández-Muñoz S (2012) Fire regime changes in the western Mediterranean Basin: from fuel-limited to drought-driven fire regime. Clim Change 110(1–2):215–226
Pechony O, Shindell DT (2010) Driving forces of global wildfires over the past millennium and the forthcoming century. Proc Natl Acad Sci USA 107(45):19167–19170
Perry DA, Hessburg PF, Skinner CN et al (2011) The ecology of mixed severity fire regimes in Washington, Oregon, and Northern California. For Ecol Manage 262:703–717
PRISM (2014) PRISM Climate Group. Oregon State University, Corvallis, OR, USA. Available from http://prism.oregonstate.edu (accessed June 3 2014)
R Core Team (2013) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available from http://www.R-project.org/
Rehfeldt GE, Crookston NL, Warwell MV, Evans JS (2006) Empirical analyses of plant-climate relationships for the western United States. Int J Plant Sci 167(6):1123–1150
Rehfeldt GE, Ferguson DE, Crookston NL (2008) Quantifying the abundance of co-occuring conifers along Inland Northwest (USA) climate gradients. Ecology 89(8):2127–2139
Röder A, Hill J, Duguy B, Alloza JA, Vallejo R (2008) Using long time series of Landsat data to monitor fire events and post-fire dynamics and identify driving factors. A case study in the Ayora region (eastern Spain). Remote Sens Environ 112(1):259–273
Rogers BM, Neilson RP, Drapek R et al (2011) Impacts of climate change on fire regimes and carbon stocks of the U.S. Pacific Northwest. J Geophys Res: Biogeosci 116(G3):G03037
Savage M, Mast JN (2005) How resilient are southwestern ponderosa pine forests after crown fires? Can J For Res 35(4):967–977
Schoennagel T, Turner MG, Romme WH (2003) The influence of fire interval and serotiny on postfire londgepole pine density in Yellowstone National Park. Ecology 84(11):2967–2978
Shatford J, Hibbs D, Puettmann K (2007) Conifer regeneration after forest fire in the Klamath-Siskiyous: how much, how soon? J For 105(3):139–146
Stephens S, Agee J, Fulé P et al (2013) Managing forests and fire in changing climates. Science 342(6154):41–42
Turner MG (2010) Landscape ecology in North America: past, present, and future. Ecology 86(8):1967–1974
Turner MG, Hargrove WW, Gardner RH, Romme WH (1994) Effects of fire on landscape heterogenity in Yellowstone National Park. Wyoming. J Veg Sci 5(5):731–742
Turner MG, Romme WH (1994) Landscape dynamics in crown fire ecosystems. Landscape Ecol 9(1):59–77
Turner MG, Romme WH, Gardner RH, Hargrove WW (1997) Effects of fire size and pattern on early succession in Yellowstone National Park. Ecol Monogr 67(4):411–433
USDA (2014) Web Soil Survey. US Department of Agriculture. Available from http://websoilsurvey.sc.egov.usda.gov/App/WebSoilSurvey.aspx (accessed Dec. 3 2014)
USFS (2012) Ch. 500: Planning for Cone and Seed Production. Forest Service Handbook Northern Region (R1) Seed Handbook. USDA Forest Service, Missoula, MT, USA. pp 31
van Etten J (2014) gdistance: Distances and routes on geographical grids. R package version 1.1-5. Available from http://CRAN.R-project.org/package=gdistance
Vander Wall SB (1994) Removal of wind-dispersed pine seeds by ground-foraging vertebrates. Oikos 69(1):125–132
Venables WN, Ripley BD (2002) Modern Applied Statistics with S. Springer, New York, NY
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006) Warming and earlier spring increase western U.S. forest wildfire activity. Science 313(5789):940–943
Williams AP, Allen CD, Millar CI et al (2010) Forest responses to increasing aridity and warmth in the southwestern United States. Proc Natl Acad Sci USA 107(50):21289–21294
York RA, Battles JJ, Heald RC (2003) Edge effects in mixed conifer group selection openings: tree height response to resource gradients. For Ecol Manage 179(1–3):107–121
Zuur A, Ieno EN, Walker N, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer Science & Business Media, LLC. New York, NY, U.S.A
Zuur AF, Savaliev AA, Ieno EN (2012) Zero inflated models and generalized linear mixed models with R. Highland Statistics Ltd., Newburg, U.K.
Zwolak R, Pearson DE, Ortega YK, Crone EE (2010) Fire and mice: seed predation moderates fire’s influence on conifer recruitment. Ecology 91(4):1124–1131
Acknowledgments
We thank K. Baker, M. Chaney, and A. Wells for assistance with data collection, S. Busby, R. Ramsey, and O. Guthrie for assistance with data collection and entry, Tim Johnson for helpful insights and assistance with statistical analysis, Zack Holden for providing the R script for calculating the distances to patch edges, and John Abatzoglou for providing downscaled climate data. This work was supported by grants from the National Aeronautics and Space Administration under award NNX11AO24G (PM), the National Science Foundation under awards DGE-0903479 (PM, KBK) and IIA-0966472 (PEH), the Joint Fire Science Program Graduate Research Innovation program under award 12-3-1-13 (KBK, PEH), and the University of Idaho Stillinger Trust Forest Science fellowship (KBK).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Rights and permissions
About this article

Cite this article
Kemp, K.B., Higuera, P.E. & Morgan, P. Fire legacies impact conifer regeneration across environmental gradients in the U.S. northern Rockies. Landscape Ecol 31, 619–636 (2016). https://doi.org/10.1007/s10980-015-0268-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-015-0268-3