
Abstract
Context
The relative influence of habitat loss versus configuration on avian biodiversity is poorly understood. However, this knowledge is essential for developing effective land use strategies, especially for grassland songbirds, which have experienced widespread declines due to land use changes. Habitat configuration may be particularly important to grassland songbirds as configuration of habitat affects the extent of edge effects on the landscape, which strongly influences habitat use by grassland birds.
Objectives
We examined the relative influence of grassland amount and a measure of grassland configuration per se (Landscape Shape Index; LSI) on the relative abundance and richness of grassland songbirds.
Methods
In 2013, 361 avian point counts were conducted across 47, 2.4 km radii landscapes in south-west Manitoba, Canada, selected to minimize the correlation between grassland amount and configuration. We used generalized linear mixed-effects models within a multi-model inference framework to determine the relative importance of grassland amount and configuration on songbird response variables.
Results
Effects of grassland amount and configuration were generally weak, but effects of configuration were greater than grassland amount for most species. Relative abundance and richness of obligate species, and Savannah sparrows, showed a strong negative response to LSI, while grasshopper sparrows responded positively to grassland amount.
Conclusion
Our results suggest that habitat configuration must be considered when managing landscapes for conservation of grassland songbirds. Maintaining large, intact tracts of grasslands and limiting development of roads that bisect grassland parcels may be an effective means of maintaining grassland songbird diversity and abundance in northern mixed-grass prairies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Grassland songbirds in North America have undergone widespread population declines (Sauer et al. 2011). While these declines have often been assumed to result from loss and fragmentation of native grasslands (Winter and Faaborg 1999), the relative influence of these landscape-scale processes on grassland songbirds remains unclear. Both habitat loss and fragmentation can influence biodiversity; however, understanding the relative influence of these variables on populations and communities is critical for developing effective management strategies (Fletcher et al. 2007), as their effects on wildlife may differ. Because habitat loss and fragmentation reflect changes in landscape composition and configuration respectively, land-use guidelines should differ if habitat loss or fragmentation is the main causal factor of population declines (Fletcher et al. 2007). For example, if habitat loss is the main driver of species declines, then management efforts should focus mainly on habitat conservation and restoration (Fahrig 1997). Conversely, if habitat fragmentation has a greater effect than habitat loss, it may be possible to mitigate the effects of habitat loss through effective land-use planning (Fahrig 1997), such as maintaining or restoring large contiguous grassland patches with low amounts of edge-affected habitat.
The effects of habitat amount and fragmentation on avian biodiversity have been well studied in some habitats (McGarigal and Cushman 2002), although not in grasslands (Shahan et al. 2017). Habitat loss results in a reduction in the amount of habitat in a landscape, which directly affects populations because individuals are inevitably lost from a landscape when habitat is removed (Bender et al. 1998; Schmiegelow and Mönkkönen 2002). Reductions in habitat amount can also have indirect negative impacts by altering species interactions (Kruess and Tscharntke 1994), reducing reproductive (Robinson et al. 1995; Kurki et al. 2000) and dispersal success (Belisle et al. 2001; Bender et al. 2003), and increasing local extinction rates (Fahrig 2001, 2002). Habitat fragmentation, defined as a change in habitat configuration in which contiguous habitat is broken apart (Fahrig 2003), can have important impacts that are independent of habitat loss. Habitat fragmentation results in an increase in the complexity of habitat configuration, resulting in an increase in the number of small habitat patches and ultimately increasing the amount of edge-affected habitat (Fahrig 2003). Two of the most important mechanisms that explain negative effects of habitat fragmentation on species (Fahrig 2003) are (1) resulting patches may be too small to support local populations or even a single home range, and (2) negative edge effects.
While habitat amount and configuration influence biodiversity through different ecological processes, estimating their relative importance is difficult because most configuration metrics typically covary with habitat amount across randomly selected landscapes (Fahrig 2003). For example, indices of habitat configuration such as patch size and isolation are often correlated with amount of habitat in the surrounding landscape; patch size generally decreases and patch isolation increases as amount of habitat declines. Although some studies have attempted to distinguish between effects of habitat amount and configuration per se, past efforts to remove the correlation between habitat amount and configuration have been met with difficulties. Some studies, for example, have attempted to distinguish between the effects of habitat amount and configuration by creating experimental landscapes and manipulating either the amount or configuration of habitat while holding all other factors constant (e.g., Caley et al. 2001; With and Pavuk 2011, 2012). Although this approach can allow researchers to isolate ecological effects of habitat configuration from habitat amount (or vice versa), it is logistically difficult to manipulate landscapes at a spatial scale that is relevant to conservation management (McGarigal and Cushman 2002). Alternatively, various methods have been used to separate effects of habitat amount and configuration in real landscapes by statistically controlling for their correlation, but this approach is often ineffective (reviewed in Smith et al. 2009). Smith et al. (2009) demonstrated that one of the most common approaches—using residuals of habitat configuration variables regressed against habitat amount as an uncorrelated index of configuration—can result in a bias in favour of either habitat amount or configuration, and the direction of the bias can vary depending on the relationship between the effects of the habitat amount and configuration predictor variables.
Given the problems associated with most of the statistical methods that have been used to distinguish between the effects of habitat amount and configuration, it is necessary to apply a different approach that explicitly separates the ecological impacts of habitat amount and configuration in real landscapes. One such approach is to control for the correlation between configuration and habitat amount in the experimental design (Pasher et al. 2013). Although several studies have used this approach to study the relative impacts of habitat amount and configuration on forest birds (e.g., McGarigal and McComb 1995; Trzcinski et al. 1999), this method has not yet been used to study the effects of habitat amount and configuration on grassland songbirds. Given that species’ sensitivity to configuration may differ among biomes (Ewers and Didham 2007), and that many grassland songbirds are sensitive to edge effects (e.g. Helzer and Jelinski 1999; Davis and Brittingham 2004; Sliwinski and Koper 2012), research in grasslands is needed to develop effective management strategies for conservation of declining grassland songbirds.
Numerous metrics have been developed to capture configuration changes associated with the division of habitat (Hargis et al. 1998). But to effectively capture the ecological consequences associated with habitat configuration independent of habitat amount, it is necessary to select a landscape metric that is (1) not highly correlated with habitat amount, and (2) is biologically relevant to the process or species of interest (Turner et al. 2001; Li et al. 2005). Shape metrics, rather than patch area and isolation metrics, generally have a weak correlation with habitat amount and may, therefore, be suitable for distinguishing between the effects of habitat amount and configuration (Wang et al. 2014). This group of metrics contains a number of different indices that can be used to describe the geometric complexity of patch shapes at the patch, class or landscape level. Within this group is the landscape shape index (LSI), which quantifies amount of edge for a given land cover class relative to that of a maximally compact and simple shape (i.e., a circle) of the same area (McGarigal and Marks 1995), capturing several configurational changes associated with the division of habitat (i.e., changes in edge amount, patch size and number of patches; Fig. 1; Saura and Carballal 2004).

Comparison of the landscape shape index for three different patterns of habitat configuration. For a single square patch LSI = 1, but for habitat comprised of irregular shapes or multiple patches, LSI > 1. The LSI values for patterns a, b and c are 1, 1.5 and 3 respectively
Compared to the effects of patch size and isolation, effects of habitat shape have received considerably less attention (Davis and Brittingham 2004; Ewers and Didham 2007; Saura et al. 2008); however, shape may be an important predictor of grassland songbird abundance and occupancy because of the sensitivity of many species of grassland songbirds to habitat edge (Helzer and Jelinski 1999). Grassland songbirds may experience elevated rates of nest predation (Winter et al. 2000), brood parasitism (Davis and Sealy 2000), competition (Fletcher and Koford 2003) or displacement by invasive species near grassland edges (Gelbard and Harrison 2003). Microclimates and resource availability may also differ in proximity to edges (Koper et al. 2009). Indeed, edge effects have been reported for numerous species of grassland birds including Sprague’s pipit (Koper et al. 2009), chestnut-collared longspurs, and Baird’s sparrow (Sliwinski and Koper 2012). While a single measure of habitat configuration does not capture every ecological impact that results from the breaking apart of habitat, LSI is likely to be biologically meaningful for species that are sensitive to habitat edges, such as grassland songbirds.
Therefore, we designed this study to determine the relative influence of habitat amount and configuration on grassland songbirds by using a sampling design that incorporated landscapes with a range in the amount of habitat and LSI values while minimizing the correlation between each variable (sensu Trzcinski et al. 1999; Ethier and Fahrig 2011; Pasher et al. 2013).
Methods
Study area
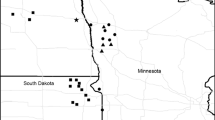
Our study was conducted in south-west Manitoba (MB), Canada, covering an area of approximately 17,000 square km from the Canada/USA border to Birtle, MB (50°25′21″N, 101°2′51″W), and the Manitoba/Saskatchewan border to Carberry, MB (49°52′8″N, 99°21′34″W; Fig. 2). The region is dominated by agricultural cropland, consisting primarily of cereal grains (e.g., wheat and oat) and oilseed (e.g., canola and sunflower), and interspersed with remnant mixed-grass prairie. Here, the mixed-grass prairie is dominated by native perennial grasses, sedges and forbs, with some low-lying shrubs such as wolf willow (Elaeagnus commutata) and western snowberry (Symphoricarpos occidentalis; Coupland 1950), and small stands of trembling aspen (Populus tremuloides) (Agriculture and Agri-Food Canada 2012). Common grass species include blue grama (Bouteloua gracilis), needle-and-thread (Hesperostipa comata), little bluestem (Schizachyrium scoparium) and western wheatgrass (Pascopyrum smithii); common forbs include pasture sage (Artemisia frigida), prairie crocus (Pulsatilla patens), and moss-phlox (Phlox hoodii). Following European settlement, many non-native grassland plant species were introduced to the mixed-grass prairie in southwestern Manitoba. Non-native plant species typical of the study region include leafy spurge (Euphorbia esula), Kentucky bluegrass (Poa pratensis), crested wheatgrass (Agropyron cristatum) and smooth brome (Bromus inermis; Wilson and Belcher 1989).

Example of the arrangement of point-count plots within 2.4 km-radii study landscapes (a) and locations of study landscapes in southwestern Manitoba, Canada, 2013 (b). Symbols represent different combinations of percent grassland cover (PG) and configuration: low grassland cover and high landscape shape index (LSI; configuration), low grassland cover and low LSI, high grassland cover and low LSI, and high grassland cover and high LSI
Selection of landscapes
We selected 47 non-overlapping landscapes from the study area that minimized the correlation between habitat amount and LSI. Landscape selection began by generating reference points (n = 255) within focal patches of grassland habitat using the editor tool in ArcMap 10. To ensure that candidate landscapes did not overlap, each reference point was separated by a minimum distance of 5 km. Points were located at least 500 metres from the nearest habitat edge, and at least 400 metres from oil wells and associated infrastructure to minimize confounding effects from energy infrastructure. We created a 2.4-km radius buffer around each point delineating the landscape boundary (i.e., an area of 18.1 km2). This spatial extent was based on a multi-scale analysis of grassland songbird response to landscape variables in southwestern Manitoba, which found that landscape structure was important most frequently within landscapes of 2.4-km radii or larger (Durán, 2009, unpublished data).
For each individual landscape (n = 255), we used Patch Analyst within ArcMap to derive data on percent grassland coverage, as an index of habitat amount, and landscape shape index (LSI) as an index of configuration. Landscape shape index was quantified as the total length of grassland edge divided by the minimum edge possible for the amount of habitat in the landscape (i.e., the circumference of a single circular patch). To quantify percent grassland cover and LSI, we used land cover data from a digitized land cover map of southwestern Manitoba from 2000 to 2002 Landsat Thematic Mapper imagery collected at a 30-m resolution with an overall thematic class accuracy of 82%, which are the best available data for this region (Agriculture and Agri-Food Canada 2012). The amount of non-cropland habitat across the Canadian prairies was relatively stable between the 1970s and 2011 (Prairie Habitat Joint Venture 2014), suggesting that these maps should be good representations of the amount of habitat available to our focal species during our surveys. Agricultural cropland, coniferous, deciduous and mixed-wood forest, open-deciduous shrub, road, wetland and grassland were among the 17 cover classes identified in the land cover dataset. Grasslands were defined as lands containing native or non-native prairie grasses and forbs with less than 10 percent shrub or tree cover (Agriculture and Agri-Food Canada 2012).
We then selected landscapes to minimize the correlation between grassland amount and LSI while still maintaining a broad range in the possible values for each variable. We achieved this by systematically selecting landscapes that contained different combinations of percent grassland cover and LSI (i.e., low grassland cover and high LSI, low grassland cover and low LSI, high grassland cover and low LSI, and high grassland cover and high LSI (sensu Trzcinski et al. 1999; Ethier and Fahrig 2011; Pasher et al. 2013; Fig. 3). Landscapes representing each of these combinations of grassland cover and LSI were distributed throughout the study area to avoid confounding habitat amount or configuration with regional trends such as climate or geology (Fig. 2). In the final set of landscapes (n = 47), grassland amount ranged from 17 to 59% (\(\bar{x} = { 31}. 3\), STDV = 10.5), LSI ranged from 4.62 to 16.78 (\(\bar{x} = { 9}. 9\), STDV = 2.9), and the pairwise correlation between percent grassland amount and LSI was r = − 0.14.

Landscapes from the study area showing different combinations of grassland cover and landscape shape index (LSI; configuration): low grassland cover and high LSI, high grassland cover and high LSI, low grassland cover and low LSI and high grassland cover and low LSI. Lighter polygons represent grassland patches
Focal study species
Because we wanted to understand effects of grassland amount and configuration per se on songbirds, our focal species included obligate and facultative grassland songbird specialists. Obligate grassland specialists are defined as species that depend on grasslands for all or part of their lifecycle (Vickery et al. 1999) and included Baird’s sparrow (Ammodramus bairdii), bobolink (Dolichonyx oryzivorus), chestnut-collared longspur (Calcarius ornatus), grasshopper sparrow (Ammodramus savannarum), horned lark (Eremophila alpestris), Le Conte’s sparrow (A. leconteii), Savannah sparrow (Passerculus sandwichensis), sedge wren (Cistothorus platensis), Sprague’s pipit (Anthus spragueii), vesper sparrow (Pooecetes gramineus) and western meadowlark (Sturnella neglecta). Facultative grassland specialists are defined as species that use grasslands but can complete their lifecycle in their absence (Vickery et al. 1999) and included brown-headed cowbird (Molothrus ater), Brewer’s blackbird (Euphagus cyanocephalus), clay-colored sparrow (Spizella pallida), eastern kingbird (Tyrannus tyrannus), red-winged blackbird (Agelaius phoeniceus) and western kingbird (T. verticalis).
Songbird surveys
We used 5-min, 100-m radius point-counts to obtain songbird relative abundance and species richness within point-count plots (n = 4 per landscape). Point-count plots were arranged in a grid pattern within a focal patch of grassland habitat at the centre of each landscape (Fig. 2). Point-count plots were defined by the maximum radius of the point-count (i.e., an area of 3.14 ha); thus, to obtain independence among point-count plots, point-count plot centres were separated by a minimum distance of 250 m, and were located at least 100 m from the nearest habitat edge to avoid local edge effects. To standardize detection, point-counts were conducted between sunrise and 1000 h, on days with winds less than 20 km/h and no precipitation (Ralph et al. 1995). All birds seen or heard within 100-m of the point-count plot centre were recorded by observers trained in visual and auditory identification of grassland birds. Most point-count plots were surveyed twice between 25 May and 4 July 2013, with observers rotated between rounds; however, in a few cases where point-count plots became flooded between survey rounds, plots were only surveyed once, resulting in 361 point-counts. Relative abundance and species richness were not combined between rounds or across individual landscapes; instead, we included data from each point-count plot (n = 361) in our analysis and used two random effects (see Statistical analysis section) to model covariance among point-counts conducted at the same plot, and among plots located within the same landscape.
Songbird detection
We used unadjusted point-count plot data as an index of relative abundance for our focal species. However, we acknowledge that the use of unadjusted count data as an index of relative bird abundance can be problematic. If detection probabilities are not constant among sites, for example, differences in bird abundance will be confounded with variation in the proportion of individuals detected, making it difficult to draw inferences about actual changes in population densities. Further, population densities and richness may be underestimated (Nicols et al. 2000; Farnsworth et al. 2002; Alldredge et al. 2007; Schmidt et al. 2013; but see Hutto and Young 2003; Johnson 2008). However, in some cases, unadjusted counts are more appropriate and precise estimates of relative abundance than alternatives. One reason for this is that assumptions of distance sampling and other statistical methods used to correct for variation in detection probabilities are difficult to meet, especially in prairie systems (Henderson and Davis 2014; Richardson et al. 2014; Leston et al. 2015). Not meeting these assumptions results in detectability adjustments that increase rather than decrease bias (Efford and Dawson 2009). For example, a core assumption of distance sampling is that all individuals are detected at zero distance from the observer, but this assumption is rarely met in open habitats such as grasslands where birds may be more likely to move away from observers or vocalize less frequently when an observer is present (Hutto and Young 2003).
Indeed, past attempts to apply distance sampling to grassland passerines have yielded poor model fit due to few detections at 0 m (e.g., Rotella et al. 1999, Davis et al. 2013, Henderson and Davis 2014). Furthermore, the minimum number of observations required to obtain reliable estimates with distance sampling (e.g., 75–100 observations per species for point transects; Buckland et al. 1993: 302) is difficult to achieve in prairie systems, as most grassland birds occur at low densities (Rotella et al. 1999, Leston et al. 2015). Distances to birds are also difficult to estimate in grasslands (Leston et al. 2015). Other approaches to dealing with imperfect detection, such as removal sampling (Farnsworth et al. 2002) and N-mixture models (Royle 2004), assume closed populations, which is usually violated in prairie systems as individuals are rarely stationary during point-counts. Territory boundaries of individuals often shift during a single breeding season in response to nest failure or changes in habitat conditions (Leston et al. 2015). Because these assumptions of detectability adjustment could not be met in our grassland ecosystem, and perceptibility of most species of grassland songbirds is very high (Leston et al. 2015), we used unadjusted counts for all species. However, we note that detectability of 2 of our focal species, grasshopper sparrow and horned lark, is lower than that of our other study species (Leston et al. 2015). Results for these two species should be interpreted with caution.
Statistical analysis
We used generalized linear mixed-effects models (GLMM; PROC GLIMMIX procedure in SAS 9.3) and an information theoretic approach to model the responses of songbird relative abundance and species richness to changes in grassland amount and configuration. Our a priori models included (1) habitat amount (percent grassland), (2) landscape shape index, (3) habitat amount + landscape shape index, and (4) a null model. Two random effects, point-count plot and landscape, were included in each model to account for the hierarchical arrangement of data points; i.e., sampling round nested within point-count plot, and plot nested in landscape.
The effects of grassland amount and configuration on relative abundance and species richness were modeled separately for obligate and facultative grassland specialists. Species for which we had sufficient data to model relative abundance (observed in at least 10% of point-counts, and for which a negative binomial or Poisson distribution fit the data according to diagnostic criteria, below) were also analyzed individually, including Savannah sparrow, grasshopper sparrow, brown-headed cowbird, bobolink, red-winged blackbird, clay-colored sparrow and sedge wren. For obligate and facultative species richness, and obligate relative abundance, diagnostic graphs (e.g., quantile–quantile plots and residual histogram plots) showed that residuals were normally distributed and thus a Gaussian distribution was used for these models. For all other response variables, excluding Savannah sparrow, red-winged blackbird, clay-colored sparrow and facultative species relative abundance, we used a Poisson distribution. Deviance/df ratios indicated that Savannah sparrow, red-winged blackbird, clay-colored sparrow and facultative species counts were overdispered so we used a negative binomial probability distribution in these models (Quinn and Keough 2002, p. 372).
We used multi-model inference (MMI) based on Akaike’s information criterion corrected for small sample sizes (AICc) to determine the relative importance of habitat amount and configuration on grassland songbird relative abundance and species richness. The model-averaged parameter estimates were used to predict changes in relative abundance and species richness relative to habitat amount and configuration, and were used here as an index of biological importance. Although Cade (2015) points out that model-averaged estimates can be inaccurate when collinearity is present, we avoided that problem through our study design, which minimized correlation among independent variables. We used the summed AICc weights (wi) associated with each variable (j) (e.g., w + [j]; Burnham and Anderson 2002, p. 168) as an index of statistical confidence in the measured effect of each variable. Although summed Akaike weights are often interpreted as a measure of relative variable importance, where larger values indicate greater variable importance (Burnham and Anderson 2002), this inference is inappropriate (Cade 2015), particularly when comparing variables that are measured on different scales (Smith et al. 2009). Thus, in this study the summed Akaike weights represent the strength of statistical support for each variable, where larger values indicated a greater strength of support, but not necessarily a greater effect size, relative to other modeled variables. Furthermore, we note that the use of summed weights is appropriate here because all variables occurred in the model set an equal number of times (Smith et al. 2009). For consistency with the literature (e.g., Barbieri and Berger 2004; Schwenk and Donovan 2011; Davis et al. 2015), we considered the relative strength of support for the measured effect of each variable to be important where w + (j)> 0.5.
While standardized regression coefficients are sometimes used to infer relative variable importance when predictors are expressed in different units (Schielzeth 2010; Grueber et al. 2011), this approach can be problematic for several reasons. Because standardized parameter estimates are on a transformed scale they can be difficult to interpret biologically (Nakagawa and Cuthill 2007; Schielzeth 2010; Grueber et al. 2011). Also, because standardized estimates are sample dependent—meaning that parameters are standardized relative to their standard deviation and the range of measurements within the study system—it is difficult to translate standardized estimates to other study areas where the variance and range of values might be different (Schielzeth 2010). To avoid these challenges, we chose to evaluate the response of grassland birds to habitat amount and configuration in the original units.
However, the effect sizes of habitat amount and configuration cannot be directly compared because of the different units used. Instead, comparisons between habitat amount and configuration were interpreted relative to their units. For example, we used “percent change in relative abundance and species richness per point-count” relative to changes across the range of observed values in habitat amount and configuration. This allowed for useful comparisons in our study because the range of values for both habitat amount and configuration were similar (i.e., a 3.5-fold difference across the range of values for both habitat and shape). These percent changes were obtained from our predictive models, which are based on our model-averaged parameter estimates from the above generalized linear mixed-effects models (see Figs. 4 and 5). If, for example, our model predicted that the number of Savanah sparrow per point count is 6 when LSI is low (i.e., 4.62) and 3 when LSI is high (i.e., 16.78), a shift across the full range of possible LSI values from low to high would result in a 50% reduction in the number of Savannah sparrows per point count. We note that changes in relative abundance among focal species should be interpreted with awareness of the overall abundance of each species, as small changes in abundance result in a larger percent change for rare compared with common species.

Grassland songbird (a) relative abundance and (b) species richness relative to amount of grassland habitat in 2.4 km-radii landscapes in southwestern Manitoba, 2013. Solid line is based on model-averaged parameter estimates from generalized linear mixed-effects models examining the response of songbird relative abundance and species richness to changes in grassland amount. Points represent the mean number of individuals detected per point count within each landscape (i.e., number of individuals averaged across survey round and point count plot). Only those species for which the relative confidence, as assessed by multi-model selection, was > 0.5 are presented

Grassland songbird (a) relative abundance and (b) species richness relative to the landscape shape index (LSI) of grasslands in 2.4 km-radii landscapes in southwestern Manitoba, 2013. Solid line is based on model-averaged parameter estimates from generalized linear mixed-effects models examining the response of songbird relative abundance and species richness to changes in LSI. Points represent the mean number of individuals detected per point count within each landscape (number of individuals averaged across survey round and point count plot). Only those species for which the relative confidence, as assessed by multi-model selection, was > 0.5 are presented
Results
Habitat amount effects
The effects of habitat amount on grassland songbird relative abundance and richness were generally weak. Evidence for an effect of habitat amount was apparent for only 3 of the 7 species evaluated (w + [j] > 0.5, Tables 1 and 2). Although there was evidence for an effect of habitat amount on obligate and facultative species richness, and facultative species abundance (w + [j] > 0.5, Table 1), the observed effect sizes were small (Table 2). For example, a change across the full range of habitat amount, from low to high (i.e., 20–60%), resulted in a decrease in facultative species of 0.8 individuals (approximately 35%) per point-count plot and a change in facultative and obligate species richness of less than 0.5 species per point-count plot (approximately 15 and 20% respectively, Fig. 4).
Habitat amount did not have a large effect size, defined here as a change of less than 1 individual per point count, on any of the species for which there was evidence of an effect of habitat amount, except for grasshopper sparrow. However, as habitat amount increased from low to high (i.e., 20–60%), grasshopper sparrow relative abundance increased by 2 individuals per point count (greater than 100% increase; Fig. 4). Grasshopper sparrows have lower detectability than other species, but as there is no reason to expect their detectability to vary systematically with habitat amount, we cautiously note that this result provides evidence of the importance of landscape-scale habitat amount for this species.
Habitat configuration
Overall, the effects of LSI were stronger than the effects of habitat amount. Evidence for an effect of LSI was apparent for obligate species abundance and richness (w + [j] = 0.99, Table 1), and for 4 of the 7 species evaluated (w + [j] > 0.5, Tables 1 and 2). Both obligate species richness and abundance responded strongly and negatively to LSI. An increase in LSI from low to high (i.e., 5–17), for example, resulted in a decrease in obligate species abundance of 4 individuals (approximately 77% decline) per point-count plot and a decrease in obligate species richness of 1.6 species per point-count plot (approximately 73% decline, Fig. 5). However, the influence (effect size) of LSI on individual species, excluding Savannah sparrow, was generally weak (i.e., a change of less than 1 individual per point-count plot).
Discussion
Overall, we found that the effects of habitat amount and configuration on grassland songbird abundance and richness were variable and mostly weak, except for the strong negative effect of configuration on grassland obligates. These results are surprising, as most previous studies, including those conducted on forest-dwelling songbirds (e.g., McGarigal and McComb 1995; Trzcinski et al. 1999), report strong and consistently negative effects of habitat loss on populations and communities (reviewed in Fahrig 2003) but weak and mostly positive effects of habitat fragmentation independent of habitat amount (reviewed in Fahrig 2017). To our knowledge, ours is the first study to select study sites that minimize correlation between grassland amount and configuration, allowing us to evaluate the relative impacts of habitat amount and configuration per se on grassland birds at the landscape scale. Because our results indicate that habitat configuration was generally more influential than grassland amount, it seems that mechanisms that explain patterns of habitat use of songbirds in grasslands probably differ from those in less planar ecosystems.
The measure of configuration that we used increases with amount of habitat edge (Saura and Carballal 2004), and it is likely that amount of edge within landscapes explains many of the biological effects that we observed. Although other studies have documented avoidance of edges by grassland birds (e.g., Winter et al. 2000; Davis and Brittingham 2004; Renfrew et al. 2005; Koper et al. 2009), the mechanisms underlying edge effects in northern mixed-grass prairies are not well understood (Sliwinski and Koper 2012). Some evidence, however, suggests that increased predation may be an important factor contributing to negative edge effects for grassland songbirds in fragmented agricultural landscapes (Winter et al. 2000), as cropland may support higher densities of generalist predators by providing supplemental resources (Heske et al. 1999). While we did not investigate effects of grassland edges on predators, other studies conducted within the same region have reported increased predation of waterfowl nests by raccoons (Procyon lotor; Stoudt 1982) in response to land use changes such as agriculture (Sargeant et al. 1993; Larivière 2004). Moreover, on multiple occasions we observed frequent nest mesopredators such as striped skunk (Mephitis mephitis) and red fox (Vulpes vulpes) travelling along cropland and road edges adjacent to our survey pastures (J. Lockhart, personal observation). Taken together, we speculate that increased predation risk may be one mechanism explaining the negative edge effects observed in our study; however, further research is required to test this hypothesis. The explanation that nest predation may contribute to the landscape-scale edge effects observed in our study contradicts findings from Renfrew et al. (2005), who found that edge avoidance by grassland songbirds was not driven by predation risk in their study system. However, as noted by the authors, it is possible that they did not detect edge effects on nest predation because fieldwork was conducted in a highly fragmented region and, therefore, the entire study area may have been influenced by edge.
Although some studies have reported increased risk of nest parasitism by brown-headed cowbirds in proximity to grassland edges (e.g., Johnson and Temple 1990; Davis and Sealy 2000; Johnson and Igl 2001), we found only a weak response of brown-headed cowbird relative abundance to LSI. Much of the anthropogenic fragmentation in this region occurs through the creation of roads and croplands, which do not provide tall perch sites from which cowbirds can scan for host nests. Edges may therefore not be attractive to brown-headed cowbirds in this grassland system. However, we note that our sample size for brown-headed cowbirds was low (n = 123) compared to other studies, so results should be taken with caution. This low number of brown-headed cowbird detections during our surveys also suggests that cowbird densities are lower in our study area than in other regions, where their densities have negative impacts on other species (Brittingham and Temple 1983; Johnson and Temple 1990), which may also mean that cowbird abundances are relatively unlikely to cause extensive edge effects across this study region.
We observed some substantial differences in the responses of obligate and facultative grassland birds to grassland amount and configuration; for example, we detected negative correlations between LSI and relative abundance of grassland obligates but not facultative grassland species. This result may be due to habitat preferences. Previous studies (e.g., Winter et al. 2000) suggest that grassland specialists demonstrate “distributional-edge sensitivity”, which is a change in density relative to edges explained by innate preferences for intact grass-dominated habitats (Sliwinski and Koper 2012). Conversely, habitat generalists are more likely to display “demographic edge-sensitivity,” or reduced nesting success in response to edges (Winter et al. 2000). Further research to assess effects of grassland amount and configuration on nesting success are required to test this hypothesis.
Our observation that grassland amount had few impacts on songbirds is in contrast with prior theoretical work that predicts significant negative effects of declining habitat amount on wildlife (e.g., Venier and Fahrig 1996; Fahrig 1997, 2001). We speculate that one reason for this is that grasslands have been replaced with other relatively planar land uses, such as agriculture (Davis et al. 2006; Sliwinski and Koper 2012). Row crops and hay lands may not hinder movement of grassland birds among habitat patches (Winter et al. 2006; McDonald 2017), and may provide few perches for avian predators (Keyel et al. 2012). Moreover, some species of grassland birds may be able to subsist in agricultural landscapes because cropland provides some substitutable resources (i.e., landscape supplementation; Dunning et al. 1992). For example, some grassland birds forage and occasionally breed in adjacent croplands (Brotons et al. 2005; Winter et al. 2006), suggesting that the agricultural matrix around grassland patches may be relatively benign to some species. Our results are consistent with findings from some other empirical studies in grasslands that found only small to moderate impacts of habitat loss and fragmentation on grassland songbirds (e.g., Davis et al. 2006; Koper and Schmiegelow 2006, but see Renfrew and Ribic 2008). These results suggest that characteristics of the vegetation within the grasslands themselves might have a greater impact on habitat suitability than does landscape-level habitat structure. Occurrence of non-native grasses, such as smooth brome and crested-wheatgrass, may reduce the quality of breeding habitat for some species of grassland birds (e.g., Lloyd and Martin 2005; Fisher and Davis 2011), and grazing management alters habitat suitability for some species (Sliwinski and Koper 2015).
Alternatively, it is possible that we were unable to detect strong effects of grassland amount on most species because habitat amount in our study landscapes ranged from 17 to 59%, which may be below a critical threshold at which effects of habitat amount can be detected. Theoretical models predict that species responses to habitat loss are nonlinear, whereby populations undergo sudden declines and extinction risk increases below a critical amount of habitat (Lande 1987; With and King 1999; Fahrig 2001). Furthermore, it is hypothesized that below a critical amount of habitat, fragmentation effects become more pronounced (Andren 1994; Fahrig 1998). To determine whether these habitat amount thresholds exist for grassland songbirds, future empirical studies could be conducted in other regions that contain a wider range of possible values of grassland amount in the landscape (e.g., 10–90% grassland amount).
In contrast to most of our other observations, habitat amount appeared to influence the abundance of grasshopper sparrows. Our results for grasshopper sparrow are consistent with other studies that have found positive effects of grassland amount on grasshopper sparrow abundance or occurrence (Johnson and Igl 2001; Davis and Brittingham 2004). The grasshopper sparrow is known to be area-sensitive (Herkert 1994), although the mechanisms for this are not known. Grasshopper sparrows generally inhabit large open grasslands dominated by relatively short bunch grasses (Smith 1963; Whitmore 1981), and they may be particularly sensitive to conspecific attraction (Fletcher 2006; Andrews et al. 2015). However, because grasshopper sparrows have lower detectability than other species, we note that these results should be taken with caution.
Conclusion
Few studies have analyzed the effects of habitat loss and fragmentation per se on grassland songbirds even though these landscape-level processes are often attributed to widespread declines of grassland birds. Although our study showed an absence of a strong impact of grassland amount and configuration, configuration was generally more important than grassland amount for prairie songbirds with the exception of grasshopper sparrows. These results have important management implications as they suggest that further habitat fragmentation could lead to more declines of grassland obligates. To minimize these negative impacts, large, intact tracts of grasslands must be conserved, and development of roads that bisect grassland parcels should be limited.
We caution, however, that management efforts focused on grassland fragmentation alone may have limited success in reversing declines of grassland songbird populations in this region given the fairly small responses of our focal species to landscape structure. Development of effective management strategies requires more information on effects of grassland structure on nesting success and survival rates, as density is not always correlated with habitat quality (Van Horne 1983; Vickery et al.1992; Winter and Faaborg 1999; Winter et al. 2006). Efforts to conserve grassland songbirds in southwestern Manitoba also may require more information on impacts of local ecological factors, such as vegetation characteristics and land management practices (e.g., grazing regimes and pesticide application). While the reasons for the continued declines of grassland songbird populations are likely multifaceted, evaluating the relative influence of local and landscape-level factors will help to inform guidelines for the conservation and management of habitat for grassland songbirds.
References
Agriculture and Agri-Food Canada (2012) Manitoba land cover 17 class 2000-2002 for agricultural regions—30 m resolution, 1st edn. Agriculture and Agri-Food Canada, Ottawa, ON. http://open.canada.ca/data/en/dataset/16d2f828-96bb-468d-9b7d-1307c81e17b8. Accessed September 2013
Alldredge MW, Pollock KH, Simons TR, Collazo JA, Shriner SA (2007) Time-of-detection methods for estimating abundance from point-count surveys. Auk 124:653–664
Andren H (1994) Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71:355–366
Andrews JE, Brawn JD, Ward MP (2015) When to use social cues: conspecific attraction at newly created grasslands. Condor 117:297–305
Barbieri MM, Berger JO (2004) Optimal predicative model selection. Ann Stat 32:870–897
Belisle M, Desrochers A, Fortin MJ (2001) Influence of forest cover on the movements of forest birds: a homing experiment. Ecology 82:1893–1904
Bender DJ, Contreras TA, Fahrig L (1998) Habitat loss and population decline: a meta-analysis of the patch size effect. Ecology 79:517–533
Bender DJ, Tischendorf L, Fahrig L (2003) Evaluation of patch isolation metrics for predicting animal movement in binary landscapes. Landscape Ecol 18:17–39
Brittingham MC, Temple SA (1983) Have cowbirds caused forest songbirds to decline? Bioscience 33:31–35
Brotons L, Wolff A, Guillaume P, Martin JL (2005) Effect of adjacent agricultural habitat on the distribution of passerines in natural grasslands. Biol Conserv 124:407–414
Buckland ST, Anderson DR, Burnham KP, Laake JL (1993) Distance sampling: estimating abundance of biological populations. Chapman and Hall, New York
Burnham KP, Anderson DR (2002) Model selection and multi-model inference: a practical information-theoretic approach. Springer, New York
Cade BS (2015) Model averaging and muddle multimodel inferences. Ecology 96:2370–2382
Caley MJ, Buckley KA, Jones GP (2001) Separating ecological effects of habitat fragmentation, degradation, and loss on coral commensals. Ecology 82:3435–3448
Coupland RT (1950) Ecology of mixed prairie in Canada. Ecol Monogr 20:271–315
Davis SK, Brigham RM, Shaffer TL, James PC (2006) Mixed-grass prairie passerines exhibit weak and variable responses to patch size. Auk 3:807–821
Davis SK, Brittingham M (2004) Area sensitivity in grassland passerines: effects of patch size, patch shape, and vegetation structure on bird abundance and occurrence in southern Saskatchewan. Auk 121:1130–1145
Davis SK, Fisher RJ, Skinner SL, Shaffer TL, Brigham RM (2013) Songbird abundance in native and planted grassland varies with type and amount of grassland in the surrounding landscape. J Wildl Manag 5:908–919
Davis AJ, Phillips ML, Doherty PF (2015) Nest success of Gunnison sage-grouse in Colorado, USA. PLOS ONE. https://doi.org/10.1371/journal.pone.0136310
Davis SK, Sealy SG (2000) Cowbird parasitism and nest predation in fragmented grasslands of Southwestern Manitoba. In: Smith JNM, Cook TL, Rothstein SI, Robinson SK, Sealy SG (eds) Ecology and management of cowbirds and their hosts. University of Texas Press, Austin, pp 220–228
Dunning JB, Danielson BJ, Pulliam HR (1992) Ecological processes that affect populations in complex landscapes. Oikos 65:169–175
Durán SM (2009) An assessment of the relative importance of wildlife management areas to the conservation of grassland songbirds in south-western Manitoba. Dissertation, University of Manitoba
Efford MG, Dawson DK (2009) Effect of distance-related heterogeneity on population size estimates from point counts. Auk 126:100–111
Ethier K, Fahrig L (2011) Positive effects of forest fragmentation, independent of forest amount, on bat abundance in eastern Ontario, Canada. Landscape Ecol 26:865–876
Ewers RM, Didham RK (2007) The effect of fragment shape and species’ sensitivity to habitat edges on animal population size. Conserv Biol 4:926–936
Fahrig L (1997) Relative effects of habitat loss and fragmentation on population extinction. J Wildl Manag 61:603–610
Fahrig L (1998) When does fragmentation of breeding habitat affect population survival? Ecol Modell 105:273–292
Fahrig L (2001) How much habitat is enough? Biol Conserv 100:65–74
Fahrig L (2002) Effect of habitat fragmentation on the extinction threshold: a synthesis. Ecol Appl 12:346–353
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol Syst 34:487–515
Fahrig L (2017) Ecological responses to habitat fragmentation per se. Annu Rev Ecol Evol Syst 48:1–23
Farnsworth GL, Pollock KH, Nichols JD, Simons TR, Hines JE, Sauer JR (2002) A removal model for estimating detection probabilities from point-count surveys. Auk 119:414–425
Fisher RJ, Davis SK (2011) Post-fledging dispersal, habitat use, and survival of Sprague’s pipits: are planted grasslands a good substitute for native? Biol Conserv 144:263–271
Fletcher RJ (2006) Emergent properties of conspecific attraction in fragmented landscapes. Am Nat 168:207–219
Fletcher RJ, Koford RR (2003) Spatial responses of bobolinks (Dolichonyx oryzivorus) near different types of edges in northern Iowa. Auk 120:799–810
Fletcher RJ, Ries L, Battin J, Chalfoun AD (2007) The role of habitat area and edge in fragmented landscapes: definitively distinct or inevitably intertwined? Can J Zool 85:1017–1030
Gelbard JL, Harrison S (2003) Roadless habitats as refuges for native grasslands: interactions with soil, aspect, and grazing. Ecol Appl 13:404–415
Grueber CE, Nakagawa S, Laws RJ, Jamieson IG (2011) Multimodel inference in ecology and evolution: challenges and solutions. J Evol Biol 4:699–711
Hargis CD, Bissonette JA, David JL (1998) The behavior of landscapes metrics commonly used in the study of habitat fragmentation. Landscape Ecol 13:167–186
Helzer CJ, Jelinski DE (1999) The relative importance of patch area and perimeter-area ratio to grassland breeding birds. Ecol Appl 9:1448–1458
Henderson AE, Davis SK (2014) Rangeland health assessment: a useful tool for linking range management and grassland bird conservation? Rangeland Ecol Manag 67:88–98
Herkert JR (1994) Status and habitat selection of the Henslow’s sparrow in Illinois. Wilson Bull 106:35–45
Heske EJ, Robinson SK, Brawn JD (1999) Predator activity and predation on songbird nests on forest-field edges in east-central Illinois. Landscape Ecol 14:345–354
Hutto RL, Young JS (2003) On the design of monitoring programs and the use of population indices: a reply to Ellingson and Lukacs. Wildl Soc Bull 31:903–910
Johnson DH (2008) In defense of indices: the case of bird surveys. J Wildl Manag 72:857–868
Johnson DH, Igl LD (2001) Area requirements of grassland birds: a regional perspective. Auk 118:24–34
Johnson RG, Temple SA (1990) Nest predation and brood parasitism of tallgrass prairie birds. J Wildl Manag 54:106–111
Keyel AC, Bauer CM, Lattin CR, Romero LM, Reed JM (2012) Testing the role of patch openness as a causal mechanism for apparent area sensitivity in a grassland specialist. Oecologia 169:407–418
Koper N, Schmiegelow FKA (2006) A multi-scaled analysis of avian response to habitat amount and fragmentation in the Canadian dry mixed-grass prairie. Landscape Ecol 21:1045–1059
Koper N, Walker D, Champagne J (2009) Nonlinear effects of distance to habitat edge on Sprague’s pipits in southern Alberta, Canada. Landscape Ecol 24:1287–1297
Kruess A, Tscharntke T (1994) Habitat fragmentation, species loss, and biological control. Science 264:1581–1584
Kurki S, Nikula A, Helle P, Linden H (2000) Landscape fragmentation and forest composition effects on grouse breeding success in boreal forests. Ecology 81:1985–1997
Lande R (1987) Extinction thresholds in demographic models of territorial populations. Am Nat 130:624–635
Larivière S (2004) Range expansion of raccoons in the Canadian prairies: review of hypotheses. Wildl Soc Bull 32:955–963
Leston L, Koper N, Rosa P (2015) Perceptibility of prairie songbirds using double-observer point counts. Gt Plains Res 1:53–61
Li X, He HS, Bu R, Wen Q, Chang Y, Hu Y, Li Y (2005) The adequacy of different landscape metrics for various landscape patterns. Pattern Recognit 38:2626–2638
Lloyd JD, Martin TE (2005) Reproductive success of chestnut-collared longspurs in native and exotic grassland. Condor 107:363–374
McDonald L (2017) The influence of patch size, landscape composition, and edge proximity on songbird densities and species richness in the northern tall-grass prairie. Dissertation, University of Manitoba
McGarigal K, Cushman SA (2002) Comparative evaluation of experimental approaches to the study of habitat fragmentation effects. Ecol Appl 12:335–345
McGarigal K, Marks BJ (1995) FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. General technical report PNW-GTR-351 USDA Forest Service, Pacific Northwest Research Station, Portland, OR
McGarigal K, McComb WC (1995) Relationships between landscape structure and breeding birds in the Oregon Coast range. Ecol Monogr 65:235–260
Nakagawa S, Cuthill IC (2007) Effect size, confidence interval and statistical significance: a practical guide for biologists. Biol Rev 82:591–605
Nicols JD, Hines JE, Sauer JR, Fallon FW, Fallon JE, Heglund PJ (2000) A double-observer approach for estimating detection probability and abundance from point counts. Auk 117:393–408
Pasher J, Mitchell SW, King DJ, Fahrig L, Smith AC, Lindsay KE (2013) Optimizing landscape selection for estimating relative effects of variables on ecological responses. Landscape Ecol 28:371–383
Prairie Habitat Joint Venture (2014) Prairie Habitat Joint Venture implementation plan 2013-2020: the Prairie Parklands. Report of the Prairie Habitat Joint Venture. Environment Canada, Edmonton, AB
Quinn GP, Keough MJ (2002) Experimental design and data analysis for biologists. Cambridge University Press, Cambridge, UK
Ralph JC, Droege S, Sauer JR (1995) Managing and monitoring birds using point counts: standards and applications. Forest Service general technical report PSW-GTR-149, U.S. Department of Agriculture, Albany, CA
Renfrew RB, Ribic CA (2008) Multi-scale models of grassland passerine abundance in a fragmented system in Wisconsin. Landscape Ecol 23:181–193
Renfrew RB, Ribic CA, Nack JL (2005) Edge avoidance by nesting grasslands: a futile strategy in a fragmented landscape. Auk 122:618–636
Richardson AN, Koper N, White KA (2014) Interactions between ecological disturbances: burning and grazing and their effects on songbird communities in northern mixed-grass prairies. Avian Conserv Ecol 2:5
Robinson SK, Thompson FR, Donovan TM, Whitehead DR, Faaborg J (1995) Regional forest fragmentation and the nesting success of migratory birds. Science 267:1987–1990
Rotella JT, Madden EM, Hansen AJ (1999) Sampling considerations for estimating density of passerines in grasslands. Stud Avian Biol 19:237–243
Royle JA (2004) N-mixture models for estimating population size from spatially replicated counts. Biometrics 60:108–115
Sargeant AB, Greenwood RJ, Sovada MA, Shaffer TL (1993) Distribution and abundance of predators that affect duck production—Prairie Pothole region. Resource Publication 194. United States Department of the Interior Fish and Wildlife Service, Washington, DC
Sauer JR, Hines JE, Fallon JE, Pardieck KL, Ziolkowski DJ, Link WA (2011) The North American breeding bird survey, results and analysis 1966-2009. Version 3.23.2011 USGS Patuxent Wildlife Research Center, Laurel, MD
Saura S, Carballal P (2004) Discrimination of native and exotic forest patterns through shape irregularity indices: an analysis in the landscape of Galicia, Spain. Landscape Ecol 19:647–662
Saura S, Torras O, Gil-Tena A, Pascual-Hortal L (2008) Shape irregularity as an indicator of forest biodiversity and guidelines for metric selection. In: Lafortezza R, Sanesi G, Chen J, Crow TR (eds) Patterns and processes in forest landscapes. Springer, Netherlands, pp 167–189
Schielzeth H (2010) Simple means to improve the interpretability of regression coefficients. Methods Ecol Evol 1:103–113
Schmidt JH, McIntyre CL, MacCluskie MC (2013) Accounting for incomplete detection: what are we estimating and how might it affect long-term passerine monitoring programs? Biol Conserv 160:130–139
Schmiegelow FKA, Mönkkönen M (2002) Habitat loss and fragmentation in dynamic landscapes: avian perspectives from the boreal forest. Ecol Appl 12:375–389
Schwenk WC, Donovan TM (2011) A multispecies framework for landscape conservation planning. Conserv Biol 25:1010–1021
Shahan JL, Goodwin BJ, Rundquist BC (2017) Grassland songbird occurrence on remnant prairie patches is primarily determined by landscape characteristics. Landscape Ecol 32:971–988
Sliwinski M, Koper N (2012) Grassland bird responses to three edge types in a fragmented mixed-grass prairie. Avian Conserv Ecol 7:6
Sliwinski M, Koper N (2015) Managing mixed-grass prairies for songbirds using variable cattle stocking rates. Rangeland Ecol Manag 68:470–475
Smith AC, Koper N, Francis CM, Fahrig L (2009) Confronting collinearity: comparing methods for disentangling the effects of habitat loss and fragmentation. Landscape Ecol 24:1271–1285
Smith RL (1963) Some ecological notes on the grasshopper sparrow. Wilson Bull 75:159–165
Stoudt JH (1982) Habitat use and productivity of canvasbacks in southwestern Manitoba, 1961-72. U.S. Fish and Wildlife Service Special Science report 248
Trzcinski MK, Fahrig L, Merriam G (1999) Independent effects of forest cover and fragmentation on the distribution of forest breeding birds. Ecol Appl 9:586–593
Turner MG, Gardner RH, O’Neil RV (2001) Landscape ecology in theory and practice: patterns and process. Springer, New York
Van Horne B (1983) Density as a misleading indicator of habitat quality. J Wildl Manag 47:893–901
Venier L, Fahrig L (1996) Habitat availability causes the species abundance-distribution relationship. Oikos 76:564–570
Vickery PD, Hunter ML, Wells JV (1992) Is density an indicator of breeding success? Auk 109:706–710
Vickery PD, Tubaro PL, Da Silva JMC, Peterjohn BG, Herkert R, Cavalcanti RB (1999) Conservation of grassland birds in the western hemisphere. Stud Avian Biol 19:2–26
Wang X, Blanchet G, Koper N (2014) Measuring habitat fragmentation: an evaluation of landscape pattern metrics. Methods Ecol Evol 5:634–646
Whitmore RC (1981) Structural characteristics of grasshopper sparrow habitat. J Wildl Manag 45:811–814
Wilson SD, Belcher JW (1989) Plant and bird communities of native prairie and introduced Eurasian vegetation in Manitoba, Canada. Conserv Biol 3:39–44
Winter M, Faaborg J (1999) Patterns of Area sensitivity in grassland-nesting birds. Conserv Biol 13:1424–1436
Winter M, Johnson DH, Faaborg J (2000) Evidence for edge effects on multiple levels: artificial nests, natural nests, and distribution of nest predators in Missouri tallgrass prairie fragments. Condor 102:256–266
Winter M, Johnson DH, Shaffer JA, Donovan TM, Svedarsky WD (2006) Patch size and landscape effects on density and nesting success of grassland birds. J Wildl Manag 1:158–172
With KA, King AW (1999) Extinction thresholds for species in fractal landscapes. Conserv Biol 13:314–326
With KA, Pavuk DM (2011) Habitat area trumps fragmentation effects on arthropods in an experimental landscape system. Landscape Ecol 26:1035–1048
With KA, Pavuk DM (2012) Direct versus indirect effects of habitat fragmentation on community patterns in experimental landscapes. Oecologia 170:517–528
Acknowledgements
We thank the many landowners for their support, E. Prokopanko and M. Garcia for their assistance in the field, and L. Fahrig, M. Manseau and two anonymous reviewers for their comments and constructive feedback on earlier drafts of the manuscript. Funding for this work was provided by the Manitoba Graduate Scholarship, Environment and Biodiversity Stewardship Fund (Manitoba Conservation), Ghostpine Environmental Service LTD Prairie Research Award, Manitoba Hydro Graduate Scholarship, Richard C. Goulden Memorial Award, Natural Sciences and Engineering Research Council, Canadian Foundation for Innovation, the Manitoba Research and Innovations Fund, and Clayton H. Riddell Endowment Fund, University of Manitoba.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Lockhart, J., Koper, N. Northern prairie songbirds are more strongly influenced by grassland configuration than grassland amount. Landscape Ecol 33, 1543–1558 (2018). https://doi.org/10.1007/s10980-018-0681-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-018-0681-5