
Abstract
Habitat area and fragmentation are confounded in many ecological studies investigating fragmentation effects. We thus devised an innovative experiment founded on fractal neutral landscape models to disentangle the relative effects of habitat area and fragmentation on arthropod community patterns in red clover (Trifolium pratense). The conventional approach in experimental fragmentation studies is to adjust patch size and isolation to create different landscape patterns. We instead use fractal distributions to adjust the overall amount and fragmentation of habitat independently at the scale of the entire landscape, producing different patch properties. Although habitat area ultimately had a greater effect on arthropod abundance and diversity in this system, we found that fragmentation had a significant effect in clover landscapes with ≤40 % habitat. Landscapes at these lower habitat levels were dominated by edge cells, which had fewer arthropods and lower richness than interior cells. Fragmentation per se did not have a direct effect on local-scale diversity, however, as demonstrated by the lack of a broader landscape effect (in terms of total habitat area and fragmentation) on arthropods within habitat cells. Fragmentation—through the creation of edge habitat—thus had a strong indirect effect on morphospecies richness and abundance at the local scale. Although it has been suggested that fragmentation should be important at low habitat levels (≤20–30 %), we show that fragmentation per se is significant only at intermediate (40 %) levels of habitat, where edge effects were neither too great (as at lower levels of habitat) nor too weak (as at higher levels of habitat).
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Habitat area has a pervasive influence on extinction risk and species diversity in most—if not all—ecological systems. Species extinction risk is typically treated as an inverse function of habitat patch size or the amount of habitat in the landscape, and the familiar species–area relationship, in which species richness increases with habitat area, has been virtually canonized as an ecological law (e.g., Lawton 1999). Because a reduction in habitat area (habitat loss) can lead to the fragmentation of habitat, the terms ‘habitat loss’ and ‘fragmentation’ are typically used interchangeably in the ecological literature. The expected consequences of habitat fragmentation are different than those of habitat loss, however (Fahrig 2003). Fragmentation more properly refers to the pattern of habitat loss resulting in an increased number of small habitat patches. Small patch size by itself is not diagnostic of fragmentation, however, as habitat loss alone can reduce patch size, such as through a targeted reduction in the size of existing fragments. Furthermore, patch isolation—a supposed consequence of fragmentation—can occur simply from a loss of habitat, such as by the complete removal of intervening habitat between patches. Thus, although fragmentation typically entails habitat loss, habitat loss can occur in ways that do not fragment the landscape. Therefore, patch-area and isolation effects may really be more a habitat-area effect than an effect of fragmentation per se (Fahrig 2003).
Habitat area and fragmentation effects are thus confounded in many studies (Ewers and Didham 2006). Although habitat area may have an overriding effect on species’ responses to landscape structure, we should not conclude that habitat fragmentation is never important, just as we should not assume that it always is. For example, fragmentation—the details of how habitat is arranged—is expected to be important below some threshold level of habitat (e.g., <20–30 %; Andrén 1994; Fahrig 1997). The exact threshold is likely to be species-specific, relative to how landscape structure influences the dispersal and reproductive potential of species (e.g., extinction thresholds; With and King 1999a). As a consequence, species are expected to exhibit different responses to habitat loss and fragmentation, based on their relative body size, trophic position, niche breadth (e.g., degree of habitat specialization), and other life-history traits (van Nouhuys 2005; Ewers and Didham 2006; Prugh et al. 2008; Öckinger et al. 2010). Nevertheless, we might reasonably expect that the cumulative effects of habitat loss and fragmentation on community patterns would be greatest at low levels of habitat, at which point most species’ extinction thresholds are likely to have been surpassed. The familiar form of the species–area relationship is perhaps a manifestation of this, in which case we would expect fragmentation (i.e., a pattern of habitat loss producing many small patches) to alter the shape or slope of the relationship, given that fragmentation has been shown to increase species’ extinction thresholds, causing species to go extinct at lower levels of habitat loss (With and King 1999a).
Along with an increase in the number of patches in the landscape, fragmentation also increases the amount of edge habitat. Habitat edges represent the interface between different biological communities, which may negatively impact habitat-interior species through an increased frequency or magnitude of species interactions resulting in higher rates of competition, predation, disease, or parasitism (e.g., Fagan et al. 1999; Olson and Andow 2008). Habitat loss alone can increase the amount of edge, however, which is maximized at intermediate levels of habitat loss (50 %). Thus, habitat area and edge effects may likewise be confounded or have synergistic effects on community patterns (Ewers et al. 2007). For studies that have controlled for both effects, however, edge effects are often stronger and more frequently observed than patch-area effects (Fletcher et al. 2007). Because fragmentation can greatly increase the amount of edge in the landscape, depending on the scale and pattern of habitat loss (With and King 1999b), species diversity is expected to be lower in fragmented landscapes as a result of negative edge effects.
Habitat fragmentation could thus have both direct and indirect effects on species diversity: direct effects at a landscape scale, operating on the shape or slope of the species–area relationship; and indirect effects on local-scale diversity through the creation of edge habitat. These effects need not be mutually exclusive, as local-scale edge effects on diversity may well translate into landscape-scale fragmentation effects. Indeed, edge effects may be the primary mechanism by which fragmentation effects manifest at the landscape scale. Nevertheless, teasing apart the direct and indirect effects of fragmentation, let alone the relative effects of habitat area and fragmentation, is a challenge in practice, given the inevitable confounding of the two in most investigations. Furthermore, since most fragmentation research is conducted at a patch scale, in which patch size or degree of isolation is measured or manipulated, it has been difficult to assess habitat area and fragmentation effects in an unambiguous fashion, or to disentangle the direct from indirect effects of fragmentation on community patterns. Fragmentation is really a landscape-scale phenomenon (McGarigal and Cushman 2002), and thus most patch-based fragmentation studies are only able to examine a small part of the patch-configuration state space, thereby limiting inferences about fragmentation effects at a landscape scale (Debinski and Holt 2000).
To circumvent these sorts of issues involving the design and confounding of fragmentation experiments, we devised an experimental landscape system inspired by neutral landscape models (Gardner et al. 1987; With 1997). Neutral landscape models are maps of statistical or mathematically generated patterns (e.g., habitat or resource distributions), which have a wide range of ecological applications, especially in the field of landscape ecology where they are commonly used to explore the interaction between landscape pattern and other ecological phenomena (With and King 1997). For example, fractal algorithms can be used to generate complex landscape patterns by tuning only two parameters that control the amount (habitat area, h) and clumping (habitat fragmentation, H) of the resulting habitat distribution (With 1997). Subsequently, we can create landscape patterns representing the extremes of fragmentation across a gradient of habitat loss. Patch-based properties, such as the number, size distribution, and distances among patches, thus emerge as a consequence of the whole-system properties of the landscape (defined by h and H). In effect, this provides a “top–down” approach to generating patch structure, as opposed to the more traditional “bottom–up” approach in manipulative experiments that vary patch-based properties to generate fragmented landscape patterns.
We have previously explored the relative effects of habitat area and fragmentation on arthropod richness over time with just such an experimental landscape system (With and Pavuk 2011). In that study, we demonstrated how habitat area—not fragmentation—was the predominant factor affecting arthropod richness within fractal landscapes of red clover (Trifolium pratense). Fragmentation had only a transient effect on arthropod richness, exhibiting a complex nonlinear relationship with habitat area in some surveys. Because that study was based on visual surveys and presence–absence data, however, we were only able to assay species richness (total number of different species types) for this particular component of the arthropod community. Thus, we have not previously evaluated fragmentation effects on higher-order community patterns (e.g., measures of diversity or species turnover), nor assessed the direct versus indirect effects of fragmentation on arthropod diversity in this system. Here, we focus on a different component of the arthropod community (microarthropods) from this same experimental landscape system, collected independently using a different sampling protocol. Because of their smaller size, it is possible that microarthropods might be more sensitive to the scale of our fragmentation experiment. We show that, while habitat area (h) generally has a greater effect than fragmentation on microarthropod diversity, the effect of fragmentation (H) was significant at intermediate habitat levels (40 % habitat) where edge effects were neither too great (as at lower habitat levels) nor too weak (as at higher habitat levels). Thus, fragmentation per se may have a weak direct effect on diversity at a landscape scale, which is restricted to intermediate habitat levels, but a strong indirect effect on local communities through edge effects.
Study area and methods
Experimental model landscape system
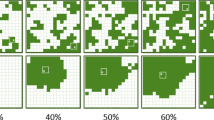
We established our experimental model landscape system (EMLS) on a 4-ha site at Bowling Green State University’s Ecology Research Station in northwest Ohio, USA. Plots (16 × 16 m2) were seeded to red clover (Trifolium pratense) as a fractal distribution of habitat at different levels (10, 20, 40, 50, 60, or 80 % clover) and degree of fragmentation (clumped, H = 1.0 vs. fragmented, H = 0.0), resulting in 12 landscape types (a combination of cover × fragmentation treatments; see With et al. 2002; With and Pavuk 2011). Plots were randomly assigned to a treatment and each landscape type (e.g., 20 % clumped) had three replicates, each with a different habitat distribution for that particular cover and fragmentation combination, for a total of 36 plots. Plots were maintained throughout the growing season (May–September) through a combination of hand-weeding (clover cells) and herbicide applications (non-clover cells) as required. The area between the plots (plots were separated by 16 m on all sides by bare ground) was also plowed periodically to keep it free of weeds, thereby enhancing the distinctiveness of the plots and reducing potential spillover effects from the matrix.
Microarthropod sampling
Microarthropod sampling was conducted on 8 days over a 2-week period at the end of the growing season (14–27 September 1997). Sampling was performed nearly 4 months after the establishment of the clover plots, which coincided with a period of maximum arthropod richness in this EMLS (With and Pavuk 2011). We randomly sampled 10 % of clover cells within each plot; clover cells to be sampled were selected using a random number generator within a spreadsheet. The number of cells sampled within each plot was thus proportional to the total habitat area within the plot: 2 cells plot−1 in 10 % landscapes, 5 cells plot−1 in 20 % landscapes, 10 cells plot−1 in 40 % landscapes, 12 cells plot−1 in 50 % landscapes, 15 cells plot−1 in 60 % landscapes, and 20 cells plot−1 in 80 % landscapes. Because of the large number of samples to be processed, we limited the number of plots sampled at higher habitat levels to three plots each (all three of the 50 % fragmented, 60 % clumped and 80 % fragmented plots), but sampled all plots at the other levels (six plots each at 10, 20, and 40 %; Fig. 1). Since most species’ thresholds may occur at <40 % habitat (Andrén 1994; Fahrig 1997), we anticipate that the effects of fragmentation (clumped vs. fragmented clover distribution) will be greatest below this level, making full sampling of all plots >40 % habitat unnecessary. Thus, we sampled a total of 243 clover cells from 27 of the 36 plots in our EMLS.

Examples of fractal habitat distributions used in the experimental landscape system. Red clover (Trifolium pratense) was planted at different coverages within plots (16 × 16 m) as either a clumped (H = 1.0, top) or fragmented (H = 0.0, bottom) distribution. Each landscape type (e.g., 10 % clumped) had three replicates, each with a different pattern, within the experimental array. Plots with 50 % clumped, 60 % fragmented, and 80 % clumped habitat distributions were not surveyed for this study (see text for explanation)
We used a hand-held D-vac unit attached to a mesh screen sieve bag (Model 122; D-vac, Ventura, CA, USA) to vacuum-sample arthropods within clover cells, by pushing the collection cone directly into the clover and sweeping it back and forth to maximize collections. D-vac sampling allows for more complete extraction of small-bodied arthropods that are missed by sweep nets or visual surveys (e.g., With and Pavuk 2011). The contents of the sieve bag collected from each cell were immediately emptied into individually labeled plastic bags (marked with the cell and plot number) and put on ice until they could be transported to the laboratory. In the laboratory, microarthropod samples were sorted and stored in individually labeled vials of 70 % ethanol until processing. During processing, microarthropods were examined under a dissecting scope and classified by one of us (D.M.P.) to the lowest taxonomic level possible. With so many individual arthropods to process (>24,000) and the high level of diversity, it was not possible to key these to species level, especially since identification of many of these microarthropods would require the expertise of a taxonomist specializing in that particular group. We thus use the term “morphospecies” in reference to the number of taxonomically distinct units within our samples, as has been done in many similar studies documenting arthropod responses to fragmentation (e.g., Bolger et al. 2000). This level of identification is sufficient for our objectives here, as we are more interested in community patterns than the response of individual species, although we do highlight a few morphospecies that contributed most to the differences among landscapes or cell types.
Microarthropod diversity
For each clover cell sample, we obtained the total number of microarthropods collected (abundance m−2) and several measures to characterize morphospecies diversity. Richness (S) was simply the total number of morphospecies in the sample. In addition, we calculated diversity indices (Shannon–Wiener, H′ and evenness, E) using standard methods (Magurran 1988). Evenness is a standardized index of diversity, based on the maximum diversity possible in the community [\( H^{\prime}_{\max } \) = ln(S)]; it is thus the proportion of maximum diversity observed in the community (E = H′ × \( H^{\prime}_{\max } \) −1). Evenness is maximized (E = 1) when all species are equally abundant (i.e., H′ = \( H^{\prime}_{\max } \)) and is low when the community is dominated by one or a few species (E → 0). Finally, at the landscape (plot) scale, we obtained morphospecies richness (S L) as the total number of morphospecies encountered across all cells sampled within that plot, and diversity measures (\( H^{\prime}_{\text{L}} \) and E L) derived from the total abundance of each species in a given plot (i.e., summed across all cells sampled).
Statistical analysis
Landscape effects on diversity at the plot scale
We used a mixed-modeling approach to examine the relative effect of habitat area and fragmentation on total abundance and diversity (S L, \( H^{\prime}_{\text{L}} \) and E L) of microarthropod communities within plots. We used PROC MIXED in SAS (SAS Institute, Cary, NC, USA) and treated cover as a fixed effect and plot as a random effect nested within the cover-by-fragmentation treatment (i.e., as a covariate). Significant effects were ultimately evaluated using Type III tests, which assess the independent effect of each predictor variable after controlling for the shared effects of any correlated predictors. This is a conservative approach, as it prevents the arbitrary attribution of shared variance to any single predictor, especially in unbalanced designs where variance among predictors is not orthogonal. Because fragmentation effects are expected to be greatest at <40 % habitat, however, we repeated this analysis for just those plots with 10–40 % habitat for a more robust assessment of treatment effects, given the balanced design at these habitat levels in which the variance between predictors is completely orthogonal. For both sets of analyses, sample sizes were the three replicate plots for each treatment combination (e.g., 20 % clumped).
Because sampling intensity varied among plots (10 % of clover cells were sampled per plot) and species richness is known to increase with sampling effort, we compared species accumulation curves using rarefaction procedures within the Species Diversity and Richness software program (SDR 4.0; Seaby and Henderson 2006). Rarefaction generates the expected number of species given a small number of individuals (or samples) drawn at random from the larger dataset, and thus effectively scales all samples down to the same number of individuals, thereby permitting equivalent comparisons of species richness among communities (landscape types, in this case). We performed both single-sample rarefactions (to provide curves for individual plots) and a pooled rarefaction across all plots, sampling without replacement to generate finite estimates of species richness.
Because habitat area ended up having a significant effect on morphospecies richness at the plot scale (see “Results”), we performed an analysis of similarity (ANOSIM) using the Community Analysis Package (CAP 4.0; Henderson and Seaby 2007). Analysis of similarity examines the relative similarity of samples within versus between groups, and is based on the Bray–Curtis percent dissimilarity index. The ANOSIM generates the R-statistic, which varies from 1 when all similar samples come from the same group to −1 when all similar samples come from different groups. A significant R-statistic in this context would thus indicate that community composition differed among plots with different habitat amounts. We then quantified the average percent dissimilarity (SIMPER) of arthropod communities within each group (e.g., among all six 10 % plots), as well as between groups (i.e., between all possible combinations of the six habitat levels = 15 pairwise comparisons). Finally, we used the Renkonen similarity index to assess the similarity of arthropod communities among all pairs of individual plots (n = 351 pairwise comparisons). Unlike qualitative indices of similarity that only assess presence–absence (e.g., Jaccards or Sorenson similarity), Renkonen similarity also accounts for relative species abundance within samples as \( \sum {\min (p_{i} ;q_{i} )} \), where p i and q i are the relative frequencies of species i in samples p and q, respectively. We selected this index because it is not as heavily influenced by sample size or number of species as some of the other similarity indices (Henderson and Seaby 2007).
Landscape effects on local-scale diversity
To assess landscape effects on local (clover cell) diversity, we partitioned the analysis by fragmentation treatment (clumped vs. fragmented), owing to the unbalanced sampling design, and essentially took a regression approach to assess whether local abundance (individuals m−2) and diversity (S, H′, and E) varied across the range of habitat levels surveyed within each treatment (clumped: 10, 20, 40, and 60 %; fragmented: 10, 20, 40, 50, and 80 %). We used PROC MIXED in SAS and treated cover as a fixed effect (with four or five levels, depending upon level of fragmentation) and plot as a random effect nested within the cover class (as a covariate). For each analysis, we explored the fit of several models that included either a linear, quadratic, or cubic function, but in each case the linear model was best (based on AICc). Significant effects of habitat cover were ultimately evaluated using Type III tests within each fragmentation type. Significant differences between fragmentation types were determined by evaluating whether the 95 % confidence intervals of the mean parameter estimates for the fixed effects overlapped. For this analysis, sample sizes varied among habitat levels, depending on the number of cells and plots sampled (e.g., 10 % of clover cells were sampled within each plot).
We again performed various analyses of community patterns using the CAP 4.0 software (Henderson and Seaby 2007). Because habitat area and fragmentation ultimately had no effect on local-scale diversity (see “Results”), we used the Renkonen similarity index to assess similarity of arthropod communities among all clover cells. We also sought to describe community structure with a principal components analysis of the between-sample variance–covariance matrix, after first performing a square-root transformation of the data because species varied greatly in abundance and the dataset included many zeroes. Our objective in using PCA was to reduce morphospecies complexity within these communities (270 individual axes or morphospecies; see “Results”) to a few orthogonal axes that best explained the variation among samples. In this case, we retained principal components that explained >5 % of the variation in the dataset.
Edge effects on arthropod diversity
Clover cells could also be characterized in terms of whether they were located within the interior of a habitat patch (the focal cell was surrounded on all sides by clover cells) or at the edge of the patch (the focal cell had at least one edge adjacent to the bareground matrix). Although roughly equal numbers of edge and interior cells were sampled across all plots (130 vs. 113, respectively), 68 % of all clover cells surveyed from fragmented landscapes (100/147) were edge cells, whereas only 31 % (30/96) of samples in clumped landscapes were from edge cells. Since very few interior cells were surveyed in fragmented landscapes with <50 % cover (only 12/87 = 14 %), and since habitat area and fragmentation were generally not important predictors of microarthropod diversity within clover cells (except total abundance within cells; see “Results”), we performed a two-tailed t test (using Satterthwaite degrees of freedom because the test for unequal variance was significant) to evaluate differences in diversity measures and total abundance between interior and edge cells.
Because morphospecies richness was significantly different between edge and interior cells (see “Results”), we performed an ANOSIM using the CAP 4.0 software (Henderson and Seaby 2007). A significant R-statistic in this context indicates that community composition differed between edge and interior cells. We also quantified the percent dissimilarity (SIMPER) among samples within each group (within edge or interior cells) as well as between groups (edge vs. interior cells).
Results
In total, our samples contained 24,106 individuals representing 270 morphospecies (n = 243, 1-m2 clover cells). Clover cells averaged 28 morphospecies (27.8 ± 0.48 SE, range 6–50) and about 100 individuals m−2 (99.2 ± 3.34, range 9–256 m−2) across all plots (n = 27). Most of these morphospecies were rare, with 77 % occurring in <10 % of clover cells we sampled. Only 7 % were found in ≥50 % cells sampled and were thus considered common to this EMLS; about half (47 %) of these common morphospecies were Dipterans (Online Resource 1). Arthropod diversity within clover cells was fairly even (H′ = 2.65 ± 0.021, range 1.58–3.53; E = 0.81 ± 0.006, range 0.52–0.99). At the plot scale, our experimental landscapes averaged 76 morphospecies per plot (75.9 ± 6.07, range 28–142), which is almost three times (2.7×) the average number of morphospecies found within individual clover cells. Microarthropod communities at the plot scale thus had higher diversity, but lower evenness, than at the local cell scale (H′ = 3.02 ± 0.07, range 2.21–3.74; E = 0.71 ± 0.014, range 0.61–0.88). Across the entire system (pooling across all individuals from all morphospecies from all plots), arthropod diversity was fairly high with only moderate evenness (3.41 ± 0.09; E = 0.61 ± 0.014; n = 270 species).
Landscape effects on diversity at the plot scale
Habitat area had a significant effect on morphospecies richness at the plot scale (F 5,20 = 55.89, P < 0.0001). Plots with 80 % habitat had 3.4× more morphospecies than plots with 10 % habitat, a difference of about 91 morphospecies (Fig. 2a). The greatest rate of increase in species occurred between 10 and 40 % habitat, in which morphospecies richness more than doubled (a 2.2× increase). Beyond 50 % habitat, morphospecies richness increased at a more modest rate (a 1.5× increase from 50 to 80 % habitat). Overall, morphospecies richness scaled as z = 0.58 on a log–log plot of species versus habitat area (S = cA z, where z is the slope of the relationship between richness, S, and area, A; R 2 = 0.98), meaning that a species was gained for roughly every 1.7 % increase in habitat area.

a Overall effect of habitat area (% clover) on morphospecies richness for microarthropods within plots of red clover (Trifolium pratense; cf. Fig. 1). b Effect of habitat fragmentation, as a function of habitat area (%), on plot-level richness (S L) for microarthropods in red clover. Note that data point for 10 % clumped and fragmented plots overlap
Fragmentation had a far weaker effect on plot-scale richness, being only marginally significant in the full analysis (F 1,20 = 3.72, P = 0.07) and just reaching the criterion for significance in an analysis restricted to the 10–40 % cover range (F 1,14 = 4.54, P = 0.05). The effect of fragmentation was evident only at 40 % habitat: clumped landscapes averaged 88.7 morphospecies (4.91 SE, n = 3 plots) compared to 73.0 morphospecies (5.57 SE, n = 3 plots) in fragmented landscapes, a difference of about 16 morphospecies (Fig. 2b). Overall, the difference between clumped and fragmented landscapes was about nine morphospecies across the 10–40 % habitat range (clumped = 63.4 ± 7.70, n = 9 plots; fragmented = 55.0 ± 5.86, n = 9), but was <1 when considered across the full range of habitat (clumped = 76.5 ± 8.96, n = 12 plots; fragmented = 75.3 ± 8.52, n = 15).
In terms of plot-scale diversity (\( H^{\prime}_{\text{L}} \) and E L), only habitat area had a significant effect on \( H^{\prime}_{\text{L}} \) (F 5,20 = 3.07, P = 0.03), but only fragmentation had a significant effect on E L (F 1,20 = 4.55, P = 0.05). Communities were more diverse in 80 % plots than 10 % plots (\( H^{\prime}_{\text{L}} \) 10 %: 2.72 ± 0.157, n = 6 plots; 80 %: 3.49 ± 0.148, n = 3 plots), but communities in fragmented landscapes were more even than those in clumped landscapes (E fragmented: 0.74 ± 0.017, n = 15 plots; clumped: 0.69 ± 0.020, n = 12 plots). None of these differences held up when only the plots in the 10–40 % range were analyzed, although the effect of fragmentation on E L was marginally significant (F 1,14 = 3.88, P = 0.07), with communities in fragmented landscapes more even than those in clumped landscapes (fragmented: 0.76 ± 0.026, n = 9 plots; clumped = 0.69 ± 0.023, n = 9 plots).
Rarefaction demonstrated how plots with little clover habitat (10 %) were less effective in sampling the larger regional species pool than plots with more habitat (e.g., 80 %; Fig. 3a). Although there was variation among plots in the rate at which morphospecies accumulate, curves from 10 % plots generally lie on the same slope as plots with more habitat. Fewer samples from 10 % plots resulted in fewer individuals and thus fewer morphospecies than plots with more habitat. Overall, morphospecies richness in this system appeared to asymptote around 12,500 individuals (Fig. 3b). Given that clover cells averaged about 100 individuals, this translates into a minimum sample of 125 clover cells (about half the number we actually sampled) to effectively characterize morphospecies richness in this system.

Rarefaction curves for a individual plots that vary in the amount of habitat (fragmented plots shown here) and b for all plots combined (dotted line 95 % confidence envelope)
Microarthropod communities within a given landscape type (as defined by habitat area) were more similar than expected by chance, and thus community composition differed significantly among plots with different levels of habitat (R = 0.41, P = 0.001, ANOSIM). The greatest differences among communities occurred between the 10 or 20 % plots and the 60 or 80 % plots (R ~ 1.0 for these pair-wise comparisons), although 10–20 % plots were significantly different from all other plots, including each other (Table 1A). The average percent dissimilarity was ultimately greatest between communities in 10 % plots and those from either 60 or 80 % plots (Table 1B). Overall, the average percent dissimilarity among plots was 56.9 % (3.90 SE, n = 15 pairwise comparisons).
In terms of Renonken similarity, microarthropod communities within plots had 57 % of morphospecies in common (0.57 ± 0.007, n = 351 pairwise comparisons), indicating a moderate degree of similarity and thus species turnover among plots (i.e., β-diversity). Although 54 % of community comparisons had at least this degree of similarity, only 16 % shared more than 70 % species and <2 % (1.7 %) shared more than 80 % species. The two most-similar communities (83 % similarity) came from two of the 40 % clumped plots, which shared 62 species and another 30–33 species unique to one plot or the other. The two least-similar communities (25.1 % similarity) were between a 10 % fragmented and 40 % fragmented plot, which had only 14 morphospecies in common and 62 other morphospecies that were found in one community or the other (14 in the 10 % fragmented plot and 48 in the 40 % fragmented plot).
Landscape effects on local-scale diversity
Habitat area did not have a significant effect on local (clover cell) microarthropod abundance, richness, or diversity at either level of fragmentation. Cells within clumped landscapes averaged 30.0 morphospecies (0.74 SE, range 14–50) and 119.6 individuals/clover cell (5.60 SE, range 32–256, n = 96 cells), whereas cells within fragmented landscapes averaged 26.4 morphospecies (2.89 SE, range 6–45) and 85.9 individuals/clover cell (9.49 SE, range 9–228, n = 147 cells). Although clover cells from fragmented landscapes averaged about four fewer species than clumped landscapes, the difference was not significant [clumped: \( \overline{Y} \) = 26.5 + 0.08(cover), 95 % CI: 20.8, 31.7; fragmented: \( \overline{Y} \) = 22.4 + 0.07(cover), 95 % CI: 17.8, 27.1]. However, clover cells within fragmented landscapes averaged significantly fewer individuals than cells in clumped landscapes [clumped: \( \overline{Y} \) = 116.2 + 0.08(cover), 95 % CI: 75.6, 156.7; fragmented: \( \overline{Y} \) = 79.8 + 0.11(cover), 95 % CI: 50.9, 108.7]. Despite this difference in abundance, local diversity (H′) and evenness (E) were unaffected by habitat area or fragmentation. Microarthropod communities within clover cells from clumped landscapes had a similar degree of diversity and evenness (H′ = 2.61 ± 0.035, range 1.67–3.53; E = 0.77 ± 0.01, range 0.52–0.97, n = 96 cells) to those in fragmented landscapes (H′ = 2.67 ± 0.026, range 1.58–3.32; E = 0.83 ± 0.007, range 0.57–0.99, n = 147 cells).
Similarity among diversity indices does not mean that communities are necessarily similar in composition, however. Microarthropod communities within clover cells had only 40 % of morphospecies in common (0.4 ± 0.001 SE, n = 29,403 pairwise comparisons; Renonken similarity), indicating a fairly low level of similarity and thus a high degree of species turnover among cells (i.e., high β-diversity). Although 53 % of community comparisons had at least this degree of similarity, only 25 % shared at least half of their species and only 6 % shared more than 60 % species. Two communities from clover cells within different 80 % fragmented landscapes had all 23 of their morphospecies in common (100 % similarity). The two least-similar communities came from clover cells within a 40 % clumped landscape and an 80 % fragmented landscape, which had only two morphospecies in common and 39 morphospecies that were found in one community or the other (0.8 % similarity).
A principal components analysis of local microarthropod communities revealed three principal components that explained 35.4 % of the variation among samples. The first principal component explained 21.5 % of the variance and loaded most heavily on Drosophila sp.1 (factor loading = 0.70); the second principal component accounted for 8.7 % of the variation and was negatively correlated with the abundance of Drosophila sp. 2 (−0.59) and positively correlated with the abundance of the collembolan Entomobyra sp. (0.56); and the third component explained 5.2 % of the variation and was inversely related to the abundance of Drosophila sp. 2 (−0.54). These three morphospecies were among the most abundant and commonly encountered in our EMLS, with Drosophila sp. 1 occurring in 96 % of clover cells sampled (27.4 ± 1.52 individuals m−2, range 1–105 in cells where it occurred), Drosophila sp. 2 was found in 82 % of samples (10.9 ± 0.82, range 1–74), and Entomobyra sp. sampled in 76 % of clover cells (9.7 ± 0.93, range 1–82). Less than 3 % of morphospecies (2.6 % of 270) occurred with >70 % frequency in our samples.
Edge effects on arthropod diversity
Interior cells had significantly more morphospecies (30.2 ± 0.64, range 6–49) than did edge cells (25.7 ± 0.65, range 11–50; t 241 = −4.93, P < 0.000). Total arthropod abundance was also about 40 % greater in interior cells (116.7 ± 5.16 individuals m−2, range 9–256) than in edge cells (84.0 ± 3.90, range 13–236; t s216 = −5.06, P < 0.000). Although diversity (H′) was similar between edge and interior cells (edge 2.63 ± 0.027, range 1.58–3.53; interior 2.67 ± 0.033, range 1.67–3.32), arthropod communities within edge cells were significantly more even (0.82 ± 0.007, range 0.57–0.99) than those in interior cells (0.79 ± 0.009, range 0.52–0.97; t 241 = 2.60, P = 0.011).
Based on an ANOSIM, microarthropod communities within edge cells were significantly different from those in interior cells (R = 0.079, P = 0.001, ANOSIM). On average, the degree of dissimilarity between communities in edge versus interior cells was 65.1 % (SIMPER between-group comparison). Much of the difference between edge and interior cells was driven by the relative abundances of the most common morphospecies: about 20 % of the difference between cell types was due to Drosophila sp. 1, which was about 60 % more abundant in interior cells than in edge cells (interior 32.7 ± 2.49, range 0–105; edge 20.8 ± 1.64, range 0–104; Fig. 4). Similarly, Drosophila sp. 2 contributed to about 8 % of the difference, although this species was more abundant in edge than interior cells (interior 8.6 ± 1.23, range 0–74; edge 9.4 ± 0.85, range 0–41; Fig. 4), and the collembolan Entomobyra sp. accounted for about 7 % of the difference (interior 9.3 ± 1.13, range 0–64; edge 5.6 ± 0.98, range 0–82). No other species contributed more than 5 % to the average dissimilarity between edge and interior cells.

Relative difference in abundance between interior and edge clover cells for the most-prevalent microarthropods that contributed to 70 % of the cumulative morphospecies richness in this system
Fifteen morphospecies were common to both cell types, although their relative abundance and degree of contribution to the similarity among local communities varied between cell types. Within interior cells, communities shared an average of 38 % of their morphospecies, with Drosophila sp. 1, Agromyzidae sp. 1, Entomobyra sp., and Drosophila sp. 2 contributing to 58 % of the similarity between samples. Edge cells also had a low degree of similarity (36 %), and communities were dominated by the three most abundant Dipterans (Drosophila sp. 1, Drosophila sp. 2, and Agromyzidae sp. 1), whose cumulative contribution to community similarity was 57 %. Beyond these “core” morphospecies, however, interior cells had an additional 17 morphospecies (total = 21 morphospecies) that contributed to most (90 %) of the similarity among cells (Fig. 5); six of these morphospecies were not found in the top 90 % of morphospecies within edge cells (Atheta sp., Myzus persicae, Syrphidae sp. 1, Telephanus velox, Syrphidae sp. 2, and Sminthuridae sp. 1). Edge cells had a total of 19 morphospecies that contributed to most of the similarity among cells (Fig. 5); four of these morphospecies were not found in the top 90 % of morphospecies encountered in interior cells (Empoasca fabae, Acyrthosiphon pisum, Notoxus monodon, Agromyzidae sp. 2).

Species-accumulation curves within edge and interior cells, depicting the morphospecies that contributed to the top 90 % of microarthropods within each cell type
Thus, out of nearly 300 morphospecies encountered in this EMLS, fewer than two dozen morphospecies accounted for much of the similarity among samples, with three or four morphospecies being nearly ubiquitous.
Discussion
By independently controlling habitat area and fragmentation in the design of our experimental landscape system, we were able to reduce the inevitable confounding that plagues most fragmentation research and tease apart their relative effects on arthropod community patterns. In so doing, we demonstrated that habitat area generally had a greater effect than fragmentation on the diversity of microarthropods in these clover landscapes, which is consistent with previous research involving a multi-year survey of macroarthropods in this same system (With and Pavuk 2011). Given that fragmentation and habitat area effects are expected to be correlated, however, might the conservative nature of our tests (Type III) simply have overlooked the potential for significant fragmentation effects once this shared variance was removed? For example, we note that habitat fragmentation had a significant effect on microarthropod richness in the analysis of just those plots with ≤40 % habitat, in which the sampling design was balanced and the effects were completely orthogonal (i.e., no shared variance between habitat area and fragmentation). However, the effect of fragmentation was really only significant at 40 % habitat (Fig. 2b). Landscape pattern (fragmentation) may be less important at <40 % owing to the overwhelming contribution of edge effects in that domain.
Fragmentation creates more edge than does habitat loss alone. Although the amount of edge is maximized at intermediate habitat levels, fragmented fractal landscapes (H = 0.0) have significantly more edge than clumped fractal landscape patterns (H = 1.0) across all levels of habitat (see fig. 4 in With and King 1999b). This effect of fragmentation on the relative amount of edge was also reflected in the samples from our experimental landscape system: a random sampling of cells from fragmented landscapes comprised mostly edge (68 %) and those from clumped landscapes were mostly interior (69 %) cells. In particular, 67 % of samples in landscapes with ≤40 % habitat were from edge cells. At 10 % habitat, almost all samples (83 %) came from edge cells, compared to 70 % of samples from 20 % plots and 60 % of samples from 40 % plots. Thus, plots with 10–20 % habitat were likely dominated by edge effects regardless of landscape pattern (so habitat distribution, H, was unimportant), whereas habitat distribution (the degree of fragmentation, H) was important in 40 % landscapes because there was now sufficient habitat area to mitigate edge effects via a clumped habitat distribution. Thus, while edge effects are expected to dominate at low levels of habitat (e.g., ≤20 % habitat), fragmentation effects (differences in habitat configuration, H) may matter most at intermediate habitat levels (e.g., 40 %), before becoming swamped out or negligible at higher habitat levels. Although it has been suggested that fragmentation is only important at low levels of habitat (e.g., <20–30 %; Andrén 1994; Fahrig 1997), the results of our study show that fragmentation per se (a landscape-wide property) is really only important at intermediate levels of habitat, whereas edge effects (a patch-scale property) dominate at lower habitat levels (regardless of habitat distribution). In essence, then, we have identified that the domain of fragmentation effects in this system occurs at an intermediate habitat level (40 %), where edge effects are neither too great nor too weak. Our study design has thus enabled us to parse out the effects due to edge from those due to fragmentation per se.
In terms of edge effects, we found that microarthropod communities within edge cells had lower abundance and richness than those within interior cells. Species richness may have been lower in edge cells partly because the matrix was devoid of vegetation, which would preclude the spillover of generalist species from neighboring habitats. Nevertheless, population densities within interior cells were higher than in edge cells, which may reflect (1) a higher perceived quality by foragers, (2) a higher reproductive output, especially if habitat quality is higher in interior cells, (3) a lower mortality as a result of lower predation or parasitism rates, which tend to be higher along habitat edges, (4) a more benign microclimate, since edges tend to be hotter and drier than the interior, or (5) some combination of these factors (Tscharntke et al. 2002a, b). In turn, higher population densities might then translate into higher morphospecies richness within interior cells, which averaged nearly five more morphospecies per clover cell than edge cells, if clover cells are simply sampling the larger landscape or regional species pool at random.
An increase in the number of random samples (either in terms of more individuals or habitat cells) should result not only in a greater number of morphospecies but also a higher degree of species turnover among cells or plots, resulting in lower similarity among communities (whether among cells or landscape types). Indeed, we found evidence in support of a random-assembly process at both the plot and local scales within this system. At the plot scale, rarefaction analysis demonstrated how plots with 10 % habitat basically followed the same species-accumulation curve as 80 % plots, which indicates that plots with less habitat area (i.e., fewer clover cells) are sampling the regional species pool less effectively than plots with more habitat. Further, out of nearly 300 morphospecies, most (77 %) occurred in fewer than 10 % of samples. Subsequently, the biggest difference in community composition occurred between 10 % and the 60 or 80 % plots, which also had the highest degree of dissimilarity (82 %, or only 18 % similarity).
At the local scale, local communities had an average similarity of 40 %, suggesting a high degree of species turnover among clover cells. In particular, there was a high degree of dissimilarity (65.1 %) in the community compositions of edge versus interior cells. In spite of this, there were some consistent differences in community composition between edge and interior cells. Although the relative abundance of the three most common morphospecies in this system (two Drosophila species and one collembolan) was responsible for much of the difference among cell types, there were ten morphospecies that were predominantly found in one cell type or the other (six in interior cells, four in edge cells). In particular, potato leafhoppers (Empoasca fabae) and pea aphids (Acyrthosiphon pisum) were more commonly encountered in edge cells than in interior cells, which could either reflect patterns of colonization (edges are more likely to be colonized through passive dispersal than interior cells) or differences in predation or parasitism rates. Parasitism rates in particular are expected to be higher along habitat edges (e.g., Thies and Tscharntke 1999), which should suppress densities of herbivores like aphids within edge cells. We found that at least one aphid parasitoid, the braconid wasp Aphidius sp., had a higher relative abundance in edge cells, which may indicate that this parasitoid was attracted to the higher aphid densities present there, producing a positive correlation rather than the negative one expected if this morphospecies were actively suppressing aphid populations. The effect of fragmentation and habitat edge on rates of parasitism in this system deserves further study and will be explored elsewhere.
Taken together, these sorts of differences in the abundance, richness, and composition of microarthropods between edge and interior cells are consistent with the expectation that community dynamics within fragmented habitats ought to be “hyperdynamic” (sensu Laurance 2002) as a result of greater variability in the occurrence and relative abundance of species, increased species turnover, and altered trophic structure, all of which may be driven by edge effects. Given that most (66 %) of the samples from landscapes with ≤40 % habitat were edge cells, and edge cells have fewer morphospecies than interior cells, it stands to reason that edge effects are really driving the significant community response to fragmentation at lower levels of habitat in this system. However, fragmentation per se is not having a direct effect on local-scale diversity, as demonstrated by the lack of a broader landscape effect (in terms of either habitat area or fragmentation) on local communities within habitat cells. Fragmentation—through the creation of edge habitat—thus had a strong indirect effect on morphospecies richness and abundance at the local scale. Fragmentation effects appear to be edge-mediated in this system, and we suspect in many other systems as well. Although edge effects may well drive community responses to fragmentation, it appears that fragmentation per se really only has a significant effect at intermediate habitat levels (e.g., 40 %), where edge effects are neither too great (as at low habitat levels) nor too weak (as at very high habitat levels). Just as we should take care to avoid equating habitat loss with fragmentation, we should similarly avoid confounding fragmentation effects with edge effects.
References
Andrén H (1994) Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71:355–366
Bolger DT, Suarez AV, Crooks KR, Morrison SA, Case TJ (2000) Arthropods in urban habitat fragments in southern California: area, age and edge effects. Ecol Appl 10:1230–1248
Debinski DM, Holt RD (2000) A survey and overview of habitat fragmentation experiments. Conserv Biol 14:342–355
Ewers RM, Didham RK (2006) Confounding factors in the detection of species responses to habitat fragmentation. Biol Rev 81:117–142
Ewers RM, Thorpe S, Didham RK (2007) Synergistic interactions between edge and area effects in a heavily fragmented landscape. Ecology 88:96–106
Fagan WF, Cantrell RS, Cosner C (1999) How habitat edges change species interactions. Am Nat 153:165–182
Fahrig L (1997) Relative effects of habitat loss and fragmentation on population extinction. J Wildl Manag 61:603–610
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Syst 34:487–515
Fletcher RJ Jr, Ries L, Battin J, Chalfoun AD (2007) The role of habitat area and edge in fragmented landscapes: definitively distinct or inevitably intertwined? Can J Zool 85:1017–1030
Gardner RH, Milne BT, Turner MG, O’Neill RV (1987) Neutral models for the analysis of broad-scale landscape pattern. Landsc Ecol 1:19–28
Henderson PA, Seaby RMH (2007) Community Analysis Package 4.0. Pisces Conservation, Lymington
Laurance WF (2002) Hyperdynamism in fragmented habitats. J Veg Sci 13:595–602
Lawton JH (1999) Are there general laws in ecology? Oikos 84:177–192
Magurran AE (1988) Ecological diversity and its measurement. Princeton University Press, New Jersey
McGarigal K, Cushman SA (2002) Comparative evaluation of experimental approaches to the study of habitat fragmentation effects. Ecol Appl 12:335–345
Öckinger E, Schweiger O, Crist TO, Debinski DM, Krauss J, Kuussaari M, Petersen JD, Pöyry J, Settele J, Summerville KS, Bommarco R (2010) Life-history traits predict species responses to habitat area and isolation: a cross-continental synthesis. Ecol Lett 13:969–979
Olson D, Andow D (2008) Patch edges and insect populations. Oecologia 155:549–558
Prugh LR, Hodges KE, Sinclair ARE, Breshares JS (2008) Effect of habitat area and isolation on fragmented animal populations. Proc Natl Acad Sci USA 105:20770–20775
Seaby RM, Henderson PA (2006) Species diversity and richness, Version 4. Pisces Conservation, Lymington
Thies C, Tscharntke T (1999) Landscape structure and biological control in agroecosystems. Science 285:893–895
Tscharntke T, Steffan-Dewenter I, Kruess A, Thies C (2002a) Characteristics of insect populations on habitat fragments: a mini-review. Ecol Res 17:229–239
Tscharntke T, Steffan-Dewenter I, Kruess A, Thies C (2002b) Contribution of small habitat fragments to conservation of insect communities of grassland-cropland landscapes. Ecol Appl 12:354–363
van Nouhuys S (2005) Effects of habitat fragmentation at different trophic levels in insect communities. Ann Zool Fenn 42:433–447
With KA (1997) The application of neutral landscape models in conservation biology. Conserv Biol 11:1069–1080
With KA, King AW (1997) The use and misuse of neutral landscape models in ecology. Oikos 79:219–229
With KA, King AW (1999a) Extinction thresholds for species in fractal landscapes. Conserv Biol 13:314–326
With KA, King AW (1999b) Dispersal success in fractal landscapes: a consequence of lacunarity thresholds. Landsc Ecol 14:73–82
With KA, Pavuk DM (2011) Habitat area trumps fragmentation effects on arthropods in an experimental landscape system. Landsc Ecol 26:1035–1048
With KA, Pavuk DM, Worchuck JL, Oates RK, Fisher JL (2002) Threshold effects of landscape structure on biological control in agroecosystems. Ecol Appl 12:52–65
Acknowledgments
This research was supported by a grant from the National Science Foundation (DEB-9610159). We gratefully acknowledge the assistance provided by H. Beecheler, S. Cadaret, J. Brubaker, and S. Grimes in helping to establish and maintain the clover plots, and R. Kappel for help with microarthropod sampling. We also thank Dr. Leigh Murray (KSU Department of Statistics) for advice on statistical design and analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Diethart Matthies.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
With, K.A., Pavuk, D.M. Direct versus indirect effects of habitat fragmentation on community patterns in experimental landscapes. Oecologia 170, 517–528 (2012). https://doi.org/10.1007/s00442-012-2325-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-012-2325-9