
Abstract
Context
Forest insect outbreaks are influenced by ecological processes operating at multiple spatial scales, including host-insect interactions within stands and across landscapes that are modified by regional-scale variations in climate. These drivers of outbreak dynamics are not well understood for the western spruce budworm, a defoliator that is native to forests of western North America.
Objectives
Our aim was to assess how processes across multiple spatial scales drive western spruce budworm outbreak dynamics. Our objective was to assess the relative importance and influence of a set of factors covering the stand, landscape, and regional scales for explaining spatiotemporal outbreak patterns in British Columbia, Canada.
Methods
We used generalized linear mixed effect models within a multi-model interference framework to relate annual budworm infestation mapped from Landsat time series (1996–2012) to sets of stand-, landscape-, and regional-scale factors derived from forest inventory data, GIS analyses, and climate models.
Results
Outbreak patterns were explained well by our model (R 2 = 93%). The most important predictors of infestation probability were the proximity to infestations in the previous year, landscape-scale host abundance, and dry autumn conditions. While stand characteristics were overall less important predictors, we did find infestations were more likely amongst pure Douglas-fir stands with low site indices and high crown closure.
Conclusions
Our findings add to growing empirical evidence that insect outbreak dynamics are driven by multi-scaled processes. Forest management planning to mitigate the impacts of budworm outbreaks should thus consider landscape- and regional-scale factors in addition to stand-scale factors.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Defoliation by insects is a natural phenomenon that occurs in forest ecosystems around the globe. Species from the genus Choristoneura ssp. are the most important native forest defoliators in North America (Volney and Fleming 2007; Nealis 2008). While native defoliators are an integral part of forest ecosystems and important for maintaining ecosystem functioning and heterogeneity (Turner 2010), past outbreaks of the eastern spruce budworm (C. fumiferana Clemens), jack pine budworm (C. pinus pinus Freeman), western spruce budworm (C. freemani Razowski = C. occidentalis Freeman), and 2-year-cycle spruce budworm (C. biennis Free.) have caused widespread defoliation in coniferous forests of North America. Historic records indicate that outbreaks have increased in duration, intensity, and spatial extent during the last century (Swetnam and Lynch 1993; Flower et al. 2014), raising concerns about the future role of susceptible forests for provision of important ecosystem services such as carbon sequestration (Kurz et al. 2008; Dymond et al. 2010) and timber supply (Alfaro et al. 1982; Gray and MacKinnon 2006). To better predict and manage the impact of future defoliator outbreaks, an improved understanding of their underlying ecological processes is required.
Several studies have identified common stand factors influencing the dynamics of western spruce budworm outbreaks: stand species composition, stand structure, and site quality, among others (Hadley and Veblen 1993; Alfaro et al. 2001; Heppner and Turner 2006; Nealis and Régnière 2009; Nealis and Regniere 2014). Those studies, building on earlier recommendations (Carlson and Wulf 1989), led to the development of management strategies to reduce stand susceptibility to future outbreaks, such as changing the species composition or structure by thinning or similar silvicultural actions (MacLean et al. 2001). However, as research on eastern spruce budworm (Cappuccino et al. 1998; Campbell et al. 2008) and other forest insect species (Simard et al. 2012; Bouchard and Auger 2013; Foster et al. 2013) indicates, stand characteristics alone cannot fully explain observed spatiotemporal patterns of insect outbreaks. This suggests that the dynamics of insect outbreaks are also driven by processes that extend well beyond the stand scale (Raffa et al. 2008). Not accounting for larger-scale processes hampers predictive models of budworm dynamics and management strategies to mitigate the impacts of future outbreaks.
Landscape- to regional-scale variations in climate and forest composition/structure are important drivers of insect outbreak dynamics. Regional weather variability, in particular moisture deficits, governs budworm population dynamics through synchronizing budworm and host phenology and through determining the quantity and quality of food (Nealis 2012). Weather variability can thus trigger and synchronize the eruption of localized outbreaks (Flower et al. 2014). During outbreak build-up, higher budworm abundance leads to increased dispersal (Anderson and Sturtevant 2011) and thus to the expansion of distinct and randomly distributed infestation patches to more continuous, landscape-scale patterns (Sturtevant et al. 2004; Bouchard and Auger 2013; Turner and Gardner 2015). However, even within regional-scale homogenous climate, not all susceptible forests experience the same level of defoliation, suggesting that some factors at the landscape scale facilitate or dampen the expansion of budworm outbreaks. Several studies provide empirical evidence that outbreak severity is not only affected by stand-scale characteristics but also by the composition and configuration of host populations in the surrounding landscape (Radeloff et al. 2000; Campbell et al. 2008; Bouchard and Auger 2013). In particular, landscape-scale host abundance and configuration is thought to influence adult moth dispersal and predator–prey interactions. Thus, while stand-scale characteristics can explain variation in localized infestations patterns during periods of low budworm densities, it is the interaction of regional-scale weather variability and landscape structure that likely determines infestation patterns during outbreaks (Sturtevant et al. 2015). To improve our understanding—and capacity to predict—the spatiotemporal patterns of budworm outbreaks, we therefore need to consider processes and interactions across multiple scales, including the stand, landscape, and regional scale.
Understanding the multi-scaled processes underlying insect outbreaks requires an explicit view at the spatial and temporal outbreak patterns over large geographic extents, yet little data sources exist that track the dynamics of forest insects at a spatial and temporal resolution suitable for conducting landscape-scale analyses. Previous studies either used aerial overview survey maps (Bouchard and Auger 2013), which have known limitations in terms of spatial accuracy (Wulder et al. 2006); or maps created from aerial photo interpretation (Seidl et al. 2015), which often only cover small geographic extents (Wulder et al. 2004). Remote sensing, in particular the Landsat series of satellites, have the potential to fill this data gap (Kennedy et al. 2014). Recent advances in the Landsat time series based mapping of bark beetles and defoliator disturbances have allowed to map the spatiotemporal dynamics of insect infestation at an ecologically valuable spatial (30 m) and temporal (annual) resolution (Townsend et al. 2012; Meigs et al. 2015; Senf et al. 2015), thus allowing for the development of spatially and temporally explicit models of outbreak dynamics (Foster et al. 2013).
Our aim was to assess how ecological processes at multiple spatial scales drive budworm outbreak dynamics. We focused on the western spruce budworm, which is an important native defoliator of coniferous forests in western North American. Western spruce budworm primarily feeds on Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) and true fir (Abies spp.) and has recently been affecting large parts of the western United States and Canada (Hicke et al. 2012). Despite its economic and ecological importance for forests of western North America, the drivers of western spruce budworm outbreak dynamics are yet largely unknown. Using annual (1996–2012) Landsat-based maps covering the recent outbreak of western spruce budworm in British Columbia, Canada, our objective was to assess the relative importance and functional relationship of a set of factors hypothesized to influence western spruce budworm outbreak dynamics at stand, landscape, and regional scales. We hypothesized that—in addition to stand factors—the probability of western spruce budworm infestation is driven by spatiotemporal variability in the abundance and configuration of budworm populations and their hosts within the landscape, by regional variability in climate, and by the interactions among these factors.
Data and methods
Study area
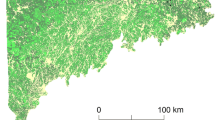
Our study area is located in British Columbia, Canada. It is delineated by the Interior Douglas-fir (IDF) biogeoclimatic zone (Hope et al. 1991) plus a 10 km buffer, which was added to mitigate possible edge effects in our analysis (Fig. 1). The study area, which is amongst the driest bioclimatic regions of Canada, is dominated by nearly homogeneous stands of Douglas-fir between 900 and 1200 m, mixed stands of Douglas-fir and ponderosa pine (Pinus ponderosa Douglas ex C. Lawson) between 600 and 900 m, and mixed stands of Douglas-fir and lodgepole pine (Pinus contorta Douglas) between 1200 and 1450 m above sea-level. Forests at higher elevations and further north in the study area are dominated by lodgepole pine, which is not a budworm host. The study area has experienced widespread defoliation by western spruce budworm in recent decades (Maclauchlan et al. 2006) and infestation by mountain pine beetle (Wulder et al. 2009). Forest health reports produced by the province of British Columbia indicate that western spruce budworm has defoliated more than one million hectares of forest in the province since 1999 (numbers obtained from: http://www2.gov.bc.ca/gov/content/environment/research-monitoring-reporting/monitoring/aerial-overview-surveys/summary-reports).

Map of western spruce budworm infestations (a) for the study area location in British Columbia, Canada (b). For better interpretability years of infestation were grouped in quintiles. Forest land indicates all land potentially covered by forest according to the Vegetation Resource Inventory, whereas forest represents actual forest cover in 1996 as mapped from Landsat. The inset (c) shows sampling location example with the three radial buffer sizes (1000, 2000, 5000 m) used for calculating landscape indices (host abundance and host edge density)
Response variable and sampling design
In a previous study, Senf et al. (2015) identified and attributed annual changes in the spectral-temporal signal of Landsat pixels (30 by 30 m) to either harvest/fire, bark beetle, or defoliator disturbances. In turn, this information supported a landscape-scale analysis of spatiotemporal western spruce budworm infestation patterns (Senf et al. 2016). Based on this information, the response variable used for subsequent analyses—annual spruce budworm infestation in a given Landsat pixel—was derived for the period 1996–2012. To minimize the number of data points (i.e., Landsat pixels) used, and thus the computational resources needed for data analyses, we applied a case-cohort sampling design. A case-cohort sampling design is efficient for rare events (as in our case where outbreaks pixels only represented 0.4% of all pixels) that allows for un-biased parameter estimation with minimum correction (King and Zeng 2001). We first randomly sampled 10,000 pixels from the whole study area and for each year and then, in a second step, randomly dropped absence samples in order to arrive at 1000 samples per year. The total sample size thus consisted of 17,000 Landsat pixels (1000 per year for 17 years, Table 1).
Potential predictors of western spruce budworm infestation
We assembled 13 predictor variables hypothesized to influence the dynamics of spruce budworm at the stand, landscape, and regional scale (Table 2).
Stand-scale predictors
Stand characteristics were obtained from British Columbia’s Vegetation Resource Inventory (VRI) database, which is a polygon-based inventory database populated and updated by photo-interpretation and field measurements (Leckie and Gillis 1995). We focused on six stand characteristics known to influence budworm habitat (Alfaro et al. 1985; Shepherd 1994; Nealis et al. 2009): age (years), crown closure (percent of forest floor covered by forest canopy), host abundance (density of hosts as a percentage of total tree density), site index (a metric of forest productivity measured as tree height at a given reference age) as measure of site productivity, and vertical complexity of the tree canopy (ordinal measure from 1 [very uniform] to 5 [very non-uniform]). The polygon-based inventory database was rasterized to a 30 by 30 m grid to match the spatial resolution of the Landsat-based map. Finally, we used a digital elevation model for estimating elevation and slope at each sampling location.
Landscape-scale predictors
We developed three predictors that describe the spatial distribution of host stands and budworm populations at landscape scale. First, we calculated the abundance and edge density of host tree patches within 1000, 2000, and 5000 m radial buffers centered on each sampled Landsat pixel to characterize the spatial composition and configuration of host forests within the surrounding landscape (Fig. 1c). The buffer sizes were chosen based on a study of another Choristoneura species (Radeloff et al. 2000), because little is known about western spruce budworm dispersal distances (Flower et al. 2014). Second, we calculated the Euclidian distance between each sampled pixel and the nearest western spruce budworm infestation in the previous year, assuming proximity to an infestation in the previous year increases the risk of infestation in the current year from dispersing adult moths (Anderson and Sturtevant 2011).
Regional-scale predictors
We acquired climate data from ClimateWNA (Wang et al. 2012), which generates climate time series for given locations using a down-scaled version of the regional-scale PRISM climate model (Daly et al. 2002). We generated seasonal values of average temperature and total precipitation for our study region for each year of our study period. From these data, we extracted four climate variables: summer precipitation and autumn precipitation in the previous year, as well as winter and spring temperature, which for all four we found evidence for importance for the development of regional scale western spruce budworm outbreaks in a previous study (Senf et al. 2016). Summer and autumn precipitation deficits may decrease Douglas-fir growth, vigor, and resistance to defoliation, allowing western spruce budworm populations to build (Flower et al. 2014). Extreme winter temperature can substantially reduce populations of over-wintering larvae. Spring temperatures, and to a lesser extent winter temperatures, influence the phenological synchrony of western spruce budworm egg hatch and the annual flush of Douglas-fir foliage on which newly emerged larvae feed (Swetnam and Lynch 1993; Nealis 2012).
Statistical analysis
We used hierarchical generalized linear mixed models (GLMM) with a logit link function to explain the annual probability of western spruce budworm infestation at the pixel level (Bolker et al. 2009). Using hierarchical GLMM allowed us to integrate fixed effects measured at varying spatial scales while simultaneously accounting for annual differences in western spruce budworm infestation probability not explained by our model. A model that does not consider the multi-level structure of the data might not detect minor effects and bias inferences made based on the model. The model used in this study is defined as:
with P(y i = 1) indicating the probability of infestation at pixel i, X i is a matrix containing the stand- and landscape-scale predictors at each pixel, the vector β contains the fixed effects at the stand-and landscape-scale, and α j is the intercept varying among years j. The varying intercept is modeled by a normal distribution:
with γ 0 representing the baseline probability of occurrence, U j is a matrix of regional-scale predictors available for each year j, the vector γ contains the regional-scale fixed effects, and σ 2 is the annual variability in infestation probability.
We first fitted a full model with all predictors shown in Table 2 as fixed effects. However, host abundance and host edge density were highly correlated among the three nested buffer sizes (Pearson r > 0.7 and variance inflation factor >5 for the 1000 and 2000 buffers), and host abundance within the 1000 and 2000 m buffer correlated moderately with host abundance at the stand scale (Pearson r > 0.55). To reduce multicollinearity in our model, we dropped host abundance and edge density measured within the 1000 and 2000 m buffers and only included both variables measured within the 5000 m buffer. That way, we reduced the variance inflation factor to <5, indicating low multicollinearity among predictors, while two variables characterizing host abundance and configuration in 5 km around each pixel and one variable characterizing host abundance at a smaller spatial scale (within a stand) were included in the model. We tested for intra-scale interactions between stand host abundance and other stand-scale predictors, for intra-scale interactions between landscape-scale host abundance and edge density and distance to infestation in the previous year, and for cross-scale interactions between the distance to infestation in the previous year and regional-scale weather variables. Intra-scale interactions for stand-scale variables were considered in order to control for species composition, on the one hand, and for effects of habitat structure on moth dispersal, on the other. Inter-scale interactions were chosen to represent the effect of weather on moth dispersal. We finally centered and scaled all predictors to zero mean and a standard deviation of one to improve model convergence and interpretability of the estimated parameters and interactions.
To identify the models with the greatest likelihood from all possible predictor combinations, we followed a multi-model inference framework (Burnham et al. 2010). The performance of each model was assessed based on the marginal R 2 [R 2m , variance explained by the fixed effects; Nakagawa et al. (2013)], the conditional R 2 [R 2c , variance explained by both the fixed and random effects; Nakagawa et al. (2013)], the Akaike’s information criterion corrected for small sample sizes [AICc; Grueber et al. (2011)], and the corresponding Akaike weights (wAICc), which assigns a relative strength of evidence to each competing model (Link and Barker 2006). This information-theoretic framework is better suited to conducting statistical inference than classical methods, because it does not rely on arbitrary thresholds such as P < 0.05. Using the likeliest set of models (ΔAICc < 2), we used model averaging to calculate average estimates and confidence intervals for each predictor. To address a potential bias in our estimates due to spatial and temporal auto-correlation in the residuals, we used robust sandwich estimators of the variance–covariance matrix (Zeileis 2006). The relative influence of each predictor was assessed based on the summed Akaike weights (∑wAICc). All statistical analyses were performed in the R Language and Environment for Statistical Computing (R Core Team 2014), using the sandwich (Zeileis 2006), lme4 (Bates et al. 2015), and MuMIn (Bartoń 2009) packages.
Results
We identified nine models with ΔAICc <2 and very good explanatory power of 93–94% for the marginal R 2 and 95% for the conditional R 2 (Table 3). Thus, the random effect from annual variation in the intercept term explained only 1–2% of the observed variance in spatiotemporal outbreak patterns. The intercept of −7.81 indicated a very low baseline infestation probability of <0.01 at mean values of all predictors [after correction for the case–control sampling design; King and Zeng (2001)], with small annual variation (σ 2 ~0.67).
We identified several important predictors of budworm outbreak dynamics (Table 3). Distance to infestation in the previous year, host abundance at the stand- and landscape-scale, crown closure, site index, vertical complexity, autumn precipitation, elevation, and slope were included in all candidate models with ∆AICc < 2 (∑wAICc = 1). Spring temperature had lower but still moderate support from the candidate models (∑wAICc = 0.63). Age, edge density, summer precipitation, and winter temperature had lowest support from the candidate models (∑wAICc < 0.2). There was strong evidence for cross-scale interactions between autumn precipitation and distance to infestation in the previous year (∑wAICc = 1). Moreover, there was strong evidence for intra-scale interactions between landscape-scale host abundance and distance to infestation in the previous year (∑wAICc = 1), as well as for intra-scale interactions between stand-scale host abundance and site index and vertical complexity (both ∑wAICc = 1).
At the stand scale, host abundance was positively associated with infestation probability (Table 3); with a 50% increase in host abundance, the probability of infestation increased by a maximum of 14%. There was strong evidence that host abundance interacted with site index and vertical complexity (Table 3). For pure host stands, infestation probability decreased as site index increased (Fig. 2a), but no significant changes in infestation probability could be observed if host stands became vertically more complex (Fig. 2b). When Douglas-fir was not the dominant tree species in a stand, site index and vertical complexity had opposite effects. Specifically, high vertical complexity lead to low infestation probability, while high site index values were associated with high infestation probability (Fig. 2a, b). Crown closure was also important in our model (Table 3) with high infestation probabilities predicted for stands with high canopy closure (Table 3). Finally, infestation probability decreased with increasing site slope and with increasing elevation (Table 3).

Varying effects of stand characteristics on infestation probability depending on host species abundance at the stand scale. Note that in order to more easily visualize the effect on infestation probability in the graph, we fixed the distance to infestation in the previous year at 5 km. All other predictor variables were fixed at their mean value (i.e., zero for the scaled predictor variables)
At the landscape scale, we found strong evidence for the importance of proximity to budworm infestation in the previous year and for landscape-scale host abundance (Table 3). With increasing distance to an infestation in the previous year, the probability of infestation dropped substantially. Interestingly, the distribution of distances to infestations in the previous year changed remarkably throughout the outbreak cycle (Fig. 3). Before (<1999) and after (>2007) the outbreak, the distributions were nearly uniform. During the outbreak period (1999–2007), however, more than 90% of all newly initiated patches were within 5 km proximity to infestations in the previous year. Hence, while infestations seemed to occur independently from existing outbreak centers within the landscape before and after the outbreak, there was a clear pattern of spatial contagion observable during the outbreak (see also Fig. 1). According to our study, the average dispersal distance was 1.3 km during the outbreak (1999–2007).

Distributions of distance to infestation in the previous year for varying periods of the study area. The periods are grouped into pre-outbreak (1996–1998), outbreak build-up (1999–2001), outbreak (2002–2004), outbreak decline (2005–2007), and post-outbreak (2008–2011) conditions
A higher abundance of Douglas-fir patches in the surrounding landscape led to a higher probability of infestation, though the effect was moderated by the distance to infestations in the previous year (Fig. 4). For example, while landscape-scale host abundance had little effect on infestation probability when the closest infestation was more than 15 km away, the effect increased with proximity to infestations in the previous year. However, at very close proximity to an infestation in the previous year (<500 m), the effect of landscape-scale host abundance diminished and infestation probability was high, regardless of host abundance in the surrounding landscape.

Varying effects of landscape scale host abundance on infestation probability depending on the distance to infestation in the previous year
Considering the regional-scale predictors, infestation probability decreased if preceded by higher than average autumn precipitation (Table 3), but only if the distance to budworm infestations in the previous year was low (Fig. 5). For pixels in close proximity to budworm infestations in the previous year (<5 km), infestation probability increased significantly when average autumn precipitation was below 200 mm in the preceding year. Lower support was found for spring temperature, with slightly increased infestation probability following warmer springs (Table 3).

Varying effects of autumn precipitation on infestation probability depending on the distance to infestation in the previous year
Discussion
Using annual Landsat-based maps of the most recent outbreak in British Columbia, Canada, we tested the hypothesis that spatiotemporal patterns of western spruce budworm outbreaks are driven by multi-scale processes, that is, by ecological interactions among budworm, its host, and their physical environment at stand, landscape, and regional scales. Our results support this hypothesis, adding to the growing empirical evidence that insect outbreak dynamics are driven by ecological processes occurring across multiple scales (Raffa et al. 2008; Seidl et al. 2015; Turner and Gardner 2015). In particular, we showed that the probability of infestation was mainly driven by host abundance at the landscape scale, by stand structure and species composition, and by regional-scale precipitation patterns. In addition, we found that those processes interacted within and across scales to determine probability of infestation.
Species composition had the most important influence on infestation probability at the stand scale, with stands having a high percentage of Douglas-fir having a higher probability of infestation. This finding aligns well with previous studies of western spruce budworm (Swetnam and Lynch 1993; Maclauchlan et al. 2006) and similar budworm species (Alfaro et al. 2001). Stands with high proportions of Douglas-fir can sustain larger populations of western spruce budworm due to higher availability of food, and dispersal losses are reduced with decreasing proportion of non-host species (Brookes et al. 1985, 1987). The effects of other stand characteristics (i.e., crown closure, site index, and vertical complexity) were not as strong (i.e., smaller scaled estimates in the model), though higher infestation probabilities tended to be associated with a lower site index and higher crown closure (in Douglas-fir dominated stands). Denser stands tend to have low losses during larvae and moth dispersal and can thus sustain larger budworm populations (Swetnam and Lynch 1993; Nealis et al. 2009). Trees on high quality sites might accumulate high levels of secondary metabolites used for tree defense against budworm herbivore (Alfaro et al. 2001), resulting in lower probability of infestation. The reasons why stand-scale characteristics were generally weaker predictors of infestation probability compared to landscape- and regional-scale factors may be related to the geographic scale of the analysis (Sturtevant et al. 2015). For example, while stand characteristics might explain differences in infestation probability among stands with similar environmental conditions, those effects are likely overridden by larger-scale processes once considering the landscape or regional scale. Similar effects were described for the mountain pine beetle, where once stand-scale thresholds are passed, outbreak dynamics are mostly governed by processes at the landscape and regional scale (Raffa et al. 2008).
We found that the distance to infestations in the previous year was the strongest factor determining the probability of infestation at a given location. Our model suggests that spatiotemporal outbreak patterns are mainly driven by adult moth dispersal, which leads to the colonization and infestation of new habitat and to the eruption and synchronization of local population dynamics. This finding aligns well with several studies of eastern spruce budworm and gypsy moth across North America (Bouchard and Auger 2013; Foster et al. 2013). By integrating two factors describing the composition and configuration of host patches within a 5 km radial buffer centered on each pixel location, and their interactions with budworm dispersal (i.e., distance to infestation in the previous year), we tested the hypothesis that dispersal of adult moths is only possible if habitat is available in the surrounding landscape, that is within average dispersal distance. We found that a higher abundance of Douglas-fir within the landscape increased infestation probability and thus of successful dispersal of adult moths from distant populations (>500 m). Hence, it is the combined effect of adult moth dispersal and landscape-scale host availability that shaped the landscape-scale outbreak patterns observed in our study.
Regional weather variability, especially regional precipitation patterns, also had important effects on the probability of budworm infestation, but we found that the effect was strongly dependent on distance to infestation in the previous year. As shown by previous research, water stress in host trees is an important driver of regional-scale western spruce budworm outbreaks (Flower et al. 2014), supporting our findings of increased infestation probability following lower autumn precipitation. Autumn precipitation was also shown to be a good indicator of water stress in Douglas-fir in other regions (Mildrexler et al. 2016), highlighting the importance of regional scale moisture variability for the eruption and spread of localized budworm populations throughout its native range. The moderate support for spring temperature in our model might suggest a higher degree of synchrony between larvae development and Douglas-fir bud-break under warmer spring conditions, though we here acknowledge that the mechanisms controlling budworm and host phenology are much more complex (Nealis 2012). Interestingly, we did not find strong support for the importance of summer precipitation and winter temperature in our model, even though expected based on our prior knowledge. The missing effect of summer precipitation might be explained by the fact that late summer water deficits are more important for drought stress in Douglas-fir than early summer water deficits (Mildrexler et al. 2016). A missing effect for winter temperature indicates that overwintering larvae—once in their hibernacula—are insensitive to winter temperature variations. Finally, elevation and slope, which were both important stand-scale predictors in our model, might further explain local variation in precipitation and moisture availably. Precipitation generally increases with increasing elevation and highest precipitation values are found on the steep slopes of the coastal mountain range at the western fringe of the study area. Lower elevation sites with more moderate slopes might thus have higher risk of local water deficits supporting budworm population development.
Implications and conclusion
Our findings have several implications for understanding, management, and modelling of budworm disturbances. First, our study highlights that there are important drivers beyond stand-scale characteristics that need to be evaluated when developing management strategies to mitigate the impacts of future budworm outbreaks. In particular, our study suggests that reducing western spruce budworm habitat quantity and quality within the landscape might prevent the development of regional-scale outbreaks. Reducing habitat quantity and quality could be achieved by developing management strategies that mimic natural disturbances (Long 2009) and by increasing the landscape heterogeneity in terms of tree species composition, among other characteristics (Robert et al. 2012). Second, our study suggests that climate is an important driver of western spruce budworm outbreak development. Consequently, climate change will likely alter the duration, intensity, and spatial extent of future outbreaks (Murdock et al. 2013). While our study does not project how climate change might affect future budworm outbreaks, forest managers should be mindful of the uncertainties associated with climate change in long-term planning (Millar et al. 2007). Third, our study adds important information to a process-based understanding of budworm outbreaks, and could further enhance the development of mechanistic models (e.g., LANDIS, Sturtevant et al. 2004) simulating the landscape-scale progression of outbreaks. In particular, our study suggests that mechanistic models should incorporate processes across multiple scales to improve predictions of western spruce budworm outbreak dynamics under varying management and climate scenarios (Seidl et al. 2011; Sturtevant et al. 2015). Finally, our study demonstrates the usefulness of Landsat-based time series for assessing landscape- to regional-scale drivers of insect outbreaks. Many factors found important in our study would be difficult to tackle without the spatially and temporally explicit view offered by long-term landscape-scale observations based on globally available satellite data. Landsat allows for analyzing insect outbreaks over larger extents than would be possible with field data alone, and with higher ecological resolution than using aerial overview survey maps (Meigs et al. 2015).
References
Alfaro RI, Sickle GAV, Thomson AJ, Wegwitz E (1982) Tree mortality and radial growth losses caused by the western spruce budworm in a Douglas-fir stand in British Columbia. Can J For Res 12:780–787
Alfaro RI, Taylor S, Brown RG, Clowater JS (2001) Susceptibility of northern British Columbia forests to spruce budworm defoliation. For Ecol Manag 145:181–190
Alfaro RI, Thomson AJ, Sickle GAV (1985) Quantification of Douglas-fir growth losses caused by western spruce budworm defoliation using stem analysis. Can J For Res 15:5–9
Anderson DP, Sturtevant BR (2011) Pattern analysis of eastern spruce budworm Choristoneura fumiferana dispersal. Ecography 34:488–497
Bartoń K (2009) MuMIn: multi-model inference. R-Package. https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf. Accessed 11 Nov 2016
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, Stevens MH, White JS (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecology and Evolution 24:127–135
Bouchard M, Auger I (2013) Influence of environmental factors and spatio-temporal covariates during the initial development of a spruce budworm outbreak. Landscape Ecol 29:111–126
Brookes MH, Campbell RW, Colbert JJ, Mitchell RG, Stark RW (1987) Western Spruce Budworm. USDA Forest Service Technical Bulletin No. 1694. Washington, DC. http://digitalcommons.usu.edu/cgi/viewcontent.cgi?article=1318&context=barkbeetles. Accessed: 11 Nov 2016
Brookes MH, Colbert JJ, Mitchell RG, Stark RW (1985) Managing Trees and Stands Susceptible to Western Spruce Budworm. USDA Forest Service Technical Bulletin No. 1695. Washington, DC. https://naldc.nal.usda.gov/naldc/download.xhtml?id=CAT86871967&content=PDF/latest. Accessed: 11 Nov 2016
Burnham KP, Anderson DR, Huyvaert KP (2010) AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisons. Behav Ecol Sociobiol 65:23–35
Campbell EM, MacLean DA, Bergeron Y (2008) The Severity of Budworm-Caused Growth Reductions in Balsam Fir/Spruce Stands Varies with the Hardwood Content of Surrounding Forest Landscapes. For Sci 54:195–205
Cappuccino N, Lavertu D, Bergeron Y, Régnière J (1998) Spruce budworm impact, abundance and parasitism rate in a patchy landscape. Oecologia 114:236–242
Carlson CE, Wulf NW (1989) Silvicultural strategies to reduce stand and forest susceptibility to the western spruce budworm. In: USDA Forest Service Agricultural handbook No. 676. Washington, DC. https://naldc.nal.usda.gov/download/CAT89916903/PDF. Accessed 11 Nov 2016
Daly C, Gibson WP, Taylor GH, Johnson GL, Pasteris P (2002) A knowledge-based approach to the statistical mapping of climate. Clim Res 22:99–113
Dymond CC, Neilson ET, Stinson G, Porter K, MacLean DA, Gray DR, Campagna M, Kurz WA (2010) Future Spruce Budworm Outbreak May Create a Carbon Source in Eastern Canadian Forests. Ecosystems 13:917–931
Flower A, Gavin DG, Heyerdahl EK, Parsons RA, Cohn GM (2014) Drought-triggered western spruce budworm outbreaks in the interior Pacific Northwest: a multi-century dendrochronological record. For Ecol Manag 324:16–27
Foster JR, Townsend PA, Mladenoff DJ (2013) Spatial dynamics of a gypsy moth defoliation outbreak and dependence on habitat characteristics. Landscape Ecol 28:1307–1320
Gray DR, MacKinnon WE (2006) Outbreak patterns of the spruce budworm and their impacts in Canada. For Chronicle 82:550–561
Grueber CE, Nakagawa S, Laws RJ, Jamieson IG (2011) Multimodel inference in ecology and evolution: challenges and solutions. J Evol Biol 24:699–711
Hadley KS, Veblen TT (1993) Stand response to western spruce budworm and Douglas-fir bark beetle outbreaks, Colorado Front Range. Can J For Res 23:479–491
Heppner D, Turner J (2006) Spruce weevil and western spruce budworm forest health stand establishment decision aids. J Ecosyst Manag 7:45–49
Hicke JA, Allen CD, Desai AR, Dietze MC, Hall RJ, Ted Hogg EH, Kashian DM, Moore D, Raffa KF, Sturrock RN, Vogelmann J (2012) Effects of biotic disturbances on forest carbon cycling in the United States and Canada. Glob Change Biol 18:7–34
Hope GD, Mitchell WR, Lloyd DA, Erickson WR, Harper WL, Wikeen BM (1991) Interior Douglas-fir Zone. In: Meidinger D, Pojar J (eds) Ecosystems of British Columbia. British Columbia Ministry of Forests. Victoria, British Columbia, Canada, pp 153–166
Kennedy RE, Andréfouët S, Cohen WB, Gómez C, Griffiths P, Hais M, Healey SP, Helmer EH, Hostert P, Lyons MB, Meigs GW, Pflugmacher D, Phinn SR, Powell SL, Scarth P, Sen S, Schroeder TA, Schneider A, Sonnenschein R, Vogelmann JE, Wulder MA, Zhu Z (2014) Bringing an ecological view of change to Landsat-based remote sensing. Front Ecol Environ 12:339–346
King G, Zeng L (2001) Logistic regression in rare events data. Polit Anal 9:137–163
Kurz WA, Stinson G, Rampley GJ, Dymond CC, Neilson ET (2008) Risk of natural disturbances makes future contribution of Canada’s forests to the global carbon cycle highly uncertain. Proc Natl Acad Sci USA 105:1551–1555
Leckie DG, Gillis MD (1995) Forest inventory in Canada with emphasis on map production. For Chronicle 71:74–88
Link WA, Barker RJ (2006) Model weights and the foundations of multimodel inference. Ecology 87:2626–2635
Long JN (2009) Emulating natural disturbance regimes as a basis for forest management: a North American view. For Ecol Manag 257:1868–1873
Maclauchlan LE, Brooks JE, Hodge JC (2006) Analysis of historic western spruce budworm defoliation in south central British Columbia. For Ecol Manag 226:351–356
MacLean DA (1980) Vulnerability of fir/spruce stands during uncontrolled spruce budworm outbreaks: a review and discussion. For Chronicle 56(213):221
MacLean DA, Erdle TA, MacKinnon WE, Porter KB, Beaton KP, Cormier G, Morehouse S, Budd M (2001) The Spruce Budworm Decision Support System: forest protection planning to sustain long-term wood supply. Can J For Res 31:1742–1757
Meigs GW, Kennedy RE, Gray AN, Gregory MJ (2015) Spatiotemporal dynamics of recent mountain pine beetle and western spruce budworm outbreaks across the Pacific Northwest Region, USA. For Ecol Manag 339:71–86
Mildrexler D, Yang Z, Cohen WB, Bell DM (2016) A forest vulnerability index based on drought and high temperatures. Remote Sens Environ 173:314–325
Millar CI, Stephenson NL, Stephens SL (2007) Climate change and forests of the future: managing in the face of uncertainty. Ecol Appl 17:2145–2151
Murdock TQ, Taylor SW, Flower A, Mehlenbacher A, Montenegro A, Zwiers FW, Alfaro R, Spittlehouse DL (2013) Pest outbreak distribution and forest management impacts in a changing climate in British Columbia. Environ Sci Policy 26:75–89
Nakagawa S, Schielzeth H, O’Hara RB (2013) A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol Evol 4:133–142
Nealis V (2008) Spruce budworms, Choristoneura Lederer (Lepidoptera: Tortricidae). In: Capinera J (ed) Encyclopedia of entomology. Springer, New York, pp 3524–3531
Nealis VG (2012) The phenological window for western spruce budworm: seasonal decline in resource quality. Agric For Entomol 14:340–347
Nealis VG, Noseworthy MK, Turnquist R, Waring VR (2009) Balancing risks of disturbance from mountain pine beetle and western spruce budworm. Can J For Res 39:839–848
Nealis VG, Régnière J (2009) Risk of dispersal in western spruce budworm. Agric For Entomol 11:213–223
Nealis VG, Regniere J (2014) An individual-based phenology model for western spruce budworm (Lepidoptera: Tortricidae). Can Entomol 146:306–320
R Core Team (2014) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. https://www.r-project.org/. Accessed 11 Nov 2016
Radeloff VC, Mladenoff DJ, Boyce MS (2000) The changing relation of landscape patterns and jack pine budworm populations during an outbreak. Oikos 90:417–430
Raffa KF, Aukema BH, Bentz BJ, Carroll AL, Hicke JA, Turner MG, Romme WH (2008) Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. BioScience 58:501
Robert L-E, Kneeshaw D, Sturtevant BR (2012) Effects of forest management legacies on spruce budworm (Choristoneura fumiferana) outbreaks. Can J For Res 42:463–475
Seidl R, Fernandes PM, Fonseca TF, Gillet F, Jönsson AM, Merganičová K, Netherer S, Arpaci A, Bontemps J-D, Bugmann H, González-Olabarria JR, Lasch P, Meredieu C, Moreira F, Schelhaas M-J, Mohren F (2011) Modelling natural disturbances in forest ecosystems: a review. Ecol Model 222:903–924
Seidl R, Muller J, Hothorn T, Bassler C, Heurich M, Kautz M (2015) Small beetle, large-scale drivers: how regional and landscape factors affect outbreaks of the European spruce bark beetle. J Appl Ecol 53:530–540
Senf C, Pflugmacher D, Wulder MA, Hostert P (2015) Characterizing spectral–temporal patterns of defoliator and bark beetle disturbances using Landsat time series. Remote Sens Environ 170:166–177
Senf C, Wulder MA, Campbell EM, Hostert P (2016) Using Landsat to assess the relationship between spatiotemporal patterns of western. Can J Remote Sens. doi: 10.1080/07038992.2016.1220828. Accessed 11 Nov 2016
Shepherd RF (1994) Management strategies for forest insect defoliators in British Columbia. For Ecol Manag 68:303–324
Simard M, Powell EN, Raffa KF, Turner MG (2012) What explains landscape patterns of tree mortality caused by bark beetle outbreaks in Greater Yellowstone? Glob Ecol Biogeogr 21:556–567
Sturtevant BR, Gustafson EJ, Li W, He HS (2004) Modeling biological disturbances in LANDIS: a module description and demonstration using spruce budworm. Ecol Model 180:153–174
Sturtevant B, Cooke B, Kneeshaw D, MacLean D (2015) Modeling insect disturbance across forested landscapes: insights from the spruce budworm. In: Perera AH, Sturtevant BR, Buse LJ (eds) Simulation modeling of forest landscape disturbances. Springer, New York, pp 93–134
Swetnam TW, Lynch AM (1993) Multicentury, regional-scale patterns of western spruce budworm outbreaks. Ecol Monogr 63:399–424
Townsend PA, Singh A, Foster JR, Rehberg NJ, Kingdon CC, Eshleman KN, Seagle SW (2012) A general Landsat model to predict canopy defoliation in broadleaf deciduous forests. Remote Sens Environ 119:255–265
Turner MG (2010) Disturbances and landscape dynamics in a changing world. Ecology 91:2833–2849
Turner MG, Gardner RH (2015) Landscape ecology in theory and practice. Springer, New York
Volney WJA, Fleming RA (2007) Spruce budworm (Choristoneura spp.) biotype reactions to forest and climate characteristics. Glob Chang Biol 13:1630–1643
Wang T, Hamann A, Spittlehouse DL, Murdock TQ (2012) ClimateWNA—high-resolution spatial climate data for western North America. J Appl Meteorol Climatol 51:16–29
Wulder MA, Hall RJ, Coops NC, Franklin SE (2004) High spatial resolution remotely sensed data for ecosystem characterization. BioScience 54:511–521
Wulder MA, Dymond CC, White JC, Leckie DG, Carroll AL (2006) Surveying mountain pine beetle damage of forests: a review of remote sensing opportunities. For Ecol Manag 221:27–41
Wulder MA, White JC, Grills D, Nelson T, Coops NC, Ebata T (2009) Aerial overview survey of the mountain pine beetle epidemic in British Columbia: communication of impacts. J Ecosyst Manag 10:45–58
Zeileis A (2006) Object-oriented computation of sandwich estimators. J Stat Softw 16:1–16
Acknowledgements
Cornelius Senf gratefully acknowledges financial support from the Elsa Neumann Scholarship of the Federal State of Berlin. The research presented here contributes to the Landsat Science Team (http://landsat.usgs.gov/Landsat_Science_Team_2012-2017.php).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Senf, C., Campbell, E.M., Pflugmacher, D. et al. A multi-scale analysis of western spruce budworm outbreak dynamics. Landscape Ecol 32, 501–514 (2017). https://doi.org/10.1007/s10980-016-0460-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-016-0460-0