
Abstract
Understanding the factors determining the spatial distribution of species is a major challenge in ecology and conservation. This study tests the use of ecosystem functioning variables, derived from satellite imagery data, to explore their potential use in modeling the distribution of the European badger in Mediterranean arid environments. We found that the performance of distribution models was enhanced by the inclusion of variables derived from the Enhanced Vegetation Index (EVI), such as mean EVI (a proxy for primary production), the coefficient of variation of mean EVI (an indicator of seasonality), and the standard deviation of mean EVI (representing spatial heterogeneity of primary production). We also found that distributions predicted by remote sensing data were consistent with the ecological preferences of badger in those environments, which may be explained by the link between EVI-derived variables and the spatial and temporal variability of food resource availability. In conclusion, we suggest the incorporation of variables associated with ecosystem function into species modeling exercises as a useful tool for improving decision-making related to wildlife conservation and management.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Understanding the factors determining the spatial distribution of species is a major challenge in ecology and conservation biology (Brown et al. 1995). The European badger (Meles meles) is medium-sized carnivore widely distributed across Europe. In Mediterranean arid landscapes the species is not abundant or is absent due to extreme aridity (Virgós et al. 2005). Current spatial distribution models for the European badger use occurrence data in conjunction with environmental variables derived from GIS data sources, such as topographic, climatic, and land cover/use (Virgós and Casanovas 1999a; Jepsen et al. 2005; Newton-Cross et al. 2007). These models have improved our understanding of badger distribution and abundance (Newton-Cross et al. 2007) by reducing limitations associated with field sampling (e.g., high economic cost and limited geographic range). However, data derived by GIS cartography could include limitations of ecological representativeness such as not representing relevant landscape features for the target species or inadequate spatial resolution (Pearce et al. 2001).
The use of ecosystem functioning variables could improve spatial distribution modeling due to their capacity to reflect spatial variability of landscape features and faster response to environment changes (Pettorelli et al. 2011). Ecosystem functioning variables can be extracted from remote sensing imagery, available continuously, both spatially and temporally. This allows the employment of standardized spectral indexes for monitoring species on different spatiotemporal scales (Nilsen et al. 2005) reducing extrapolations. An example of potentially useful ecosystem functioning variables are the functional attributes derived by the Enhanced Vegetation Index (EVI). The EVI has been used in mammal ecology by Wang et al. (2010), Meynard et al. (2012), and Bardsen and Tveraa (2012). The EVI is linearly related to ecosystem carbon gains, and therefore, to net primary productivity (NPP) (Monteith 1981), which is used as a surrogate of ecosystem functioning (Alcaraz et al. 2006; Cabello et al. 2012b). Thus, measures derived from EVI can describe ecosystem functional attributes (Pettorelli et al. 2005). These attributes include the mean annual EVI (i.e., surrogate of primary production) (Huete et al. 1997; Sims et al. 2006) and the coefficient of variation of mean annual EVI (i.e., indicator of seasonality) (Alcaraz-Segura et al. 2012).
The resource dispersion hypothesis posits that the size of badger territories is mainly linked to the dispersion of food resources (Macdonald 1983; Kruuk 1989; Macdonald and Carr 1999). This hypothesis emphasizes the key role of patchiness of food quality in determining how large badger territories are. For example, habitat productivity tends to drive body condition, ultimately influencing fitness (Woodroffe 1995). As a consequence, reproductive success of females is largely dependent on food conditions, which in badgers are mainly linked to climate factors mediating food abundance (e.g., productivity of habitats) (Woodroffe and Macdonald 1995). Therefore, badger demography, abundance and social life is mainly shaped by food availability and predictability (seasonality), which can be assessed by ecosystem functional attributes derived of spectral vegetation indices (e.g., Nilsen et al. 2005; Pettorelli et al. 2005, 2006).
The purpose of this study is to test the use of ecosystem functional variables derived from EVI (e.g., mean annual EVI, coefficient of variation of mean annual EVI, and spatial deviation of mean annual EVI) to improve spatial distribution modeling of the European badger. With this aim, we first sampled badger occurrence in a representative arid landscape located in the southeastern Iberian Peninsula (Fig. 1). Secondly, we designed a variety of spatial distribution models based on environmental variables, with and without including EVI-derived variables. We also explored their performance based on a subset of previously sampled presence data and the habitat preferences of badger as described by other authors. Finally, we discuss the role of ecosystem functional dimension in species ecological modeling and conservation.

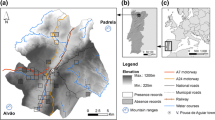
Study area location
Methods
Study area
We selected a representative area of arid landscapes in the southeastern Iberian Peninsula based on the Martonne aridity index (Martonne 1926) (Fig. 1), as it is easily calculated and mapped with GIS layers. Inside this area defined as Arid steppic by the Martonne index (range 5–15), we drew a 3.5 km-radius buffer zone on both sides of the two major rivers basins in the region, and then joined the two buffers (Fig. 1). In this form, we ensure inclusion of the potential home range estimated for European badgers in these environments (i.e., 9 km2) (Lara-Romero et al. 2012). The study area comprised 835 km2, with a temperature gradient (range of minimum mean temperatures: 7–12 °C, range of maximum mean temperatures: 23–28 °C) and an annual precipitation gradient (200–600 mm/m3) associated with a wide altitudinal gradient (0–1,400 m). Evapotranspiration ranges from 93 to 945 mm/year. Another important feature of the area is the diversity of land cover/use: xerophytic scrubs represent 48 % of the area, where Stipa tenacissima is the most abundant. Forested habitat is very scarce, corresponding mostly to scattered pine forests (Pinus halepensis). Crops occupy 27 % of the study area and include fruit orchards (especially abundant near the rivers), arable crops and greenhouses, in similar proportions.
Field survey data
A field survey was conducted from September 2010 to February 2011. The study area was divided into 5 × 5 km UTM (Universal Transverse Mercator) plots (out of total 66 plots) to organize the field surveys and not as the sampling unit. A survey to identify signs of badgers (i.e., footprints, latrines and setts) was carried out for 6 h in each plot. To maximize the detection of the species with the least effort, we selected places for survey such as paths and catchments for footprints, hills for latrines and easy to dig sloping areas for setts. These places are known to be usually used by badgers. The GPS (UTM) coordinates of each sign were noted with a measurement error of up to 10 m using a GPSmap® 60CS×-Garmin. To avoid spatial autocorrelation of environmental variables (see below), no signs within 100 m from each other were considered (see Appendix 5).
Environmental data
The study area was characterized based on twenty predictor variables (Table 1 and Appendix 4). Nine of these variables are commonly used in European badger ecology studies (e.g., Virgós and Casanovas 1999a; Revilla et al. 2000; Macdonald and Newman 2002; Jepsen et al. 2005; Rosalino et al. 2008; Lara-Romero et al. 2012), and were comprised by two climate variables, one topographic variable and six variables related to habitat structure represented by different land cover/use. The eleven remaining variables were derived from remote sensing data.
Final resolution of environmental data sets was adjusted to 100 × 100 m pixel size (i.e., sample unit), to agree with the predominant smallest spatial resolution of data (Ferrier and Watson 1997; Elith and Leathwick 2009). Some variables (i.e., land cover/use variables) were scaled to the relevant scale for badgers (i.e., their home range).
Topographic and climate variables
Topographic and climate variables were derived from spatial data layers of the Environmental Information Network of Andalusia (http://www.juntadeandalucia.es/medioambiente/site/web/rediam). ESRI® ArcMap™ 9.3 was used for their handling and processing. The topographic variable was mean slope, which has been described as a factor relevant for sett digging (Jepsen et al. 2005). It was estimated from the digital elevation model of Andalusia with a spatial and elevation pixel resolution of 20 × 20 m. This layer was resampled to 100 × 100 m.
Climate variables (Virgós and Casanovas 1999a; Johnson et al. 2002; Macdonald and Newman 2002) were mean annual rainfall and the mean maximum temperature, acquired with a resolution of 100 x 100 m, so no transformation was made.
Land cover and land use variables
The land cover and land use variables (Virgós and Casanovas 1999a; Revilla et al. 2000; Rosalino et al. 2008; Lara-Romero et al. 2012) were derived from Andalusian Land use/Land cover map (scale 1:25,000 from 2007), in vector format. This layer included the following classes: scattered scrub, dense scrub, woody crops, arable crops, mixed crops (woody and arable) and mosaic crops (crops and natural vegetation). The study area was first divided into 3 × 3 km plots. We estimated the area (km2) of each class and then we rasterized to 100 × 100 m pixel size. These variables were scaled because the percent cover is relevant for badgers (Lara-Romero et al. 2012), instead of using the class of land cover/use as categorical variable. We considered a 9 km2 area as the probable home range of the European badger in areas of low habitat suitability (Lara-Romero et al. 2012).
EVI variables
The eleven variables derived from remote sensing data were estimated based on the MOD13Q1 EVI product, generated from images captured by the MODIS sensor aboard the NASA’s TERRA satellite (www.modis.gsfc.nasa.gov) for a period of seven years. These images have the advantages of its high temporal resolution of 16 days (23 images/year) and spatial resolution appropriate to the scale of the study (231 × 231 m). The images were subjected to pixel quality filtering, in which those affected by heavy content of aerosols, clouds, shadows, snow or water were eliminated. The EVI is the index least affected by atmospheric conditions and presents fewer saturation problems for high levels of biomass (Huete et al. 2002).
The mean annual EVI is linearly related to total carbon gain (Running et al. 2000), and has been used as a surrogate of vegetation productivity (Alcaraz-Segura et al. 2012). The standard deviation of mean annual EVI is an indirect measure of spatial heterogeneity, so that a high standard deviation may indicate mixed patches, while a low standard deviation is common in homogeneous landscapes. This variable was estimated by calculating the standard deviation of mean annual EVI in the 3 × 3 km plots used to estimate the land cover/use variables. The coefficient of variation of mean annual EVI is a seasonal carbon gain descriptor (Alcaraz et al. 2006) that has been used as an indicator of ecosystem seasonality (Alcaraz-Segura et al. 2012). Furthermore, seasonality, although described by other variables, has proven decisive in modeling the habitat of several other species (Boyce 1978; Ferguson and McLoughlin 2000; Wiegand et al. 2008). In addition to these, the EVI autumn mean (September–November) and EVI spring mean (March–May) were also included as variables, because they represent the two growing seasons in Mediterranean arid landscapes (Cabello et al. 2012b).
EVI variables were resampled to 100 × 100 m by a bilinear resampling technique. It determines the new value of a cell based on a weighted distance average of the four nearest input cell centers. This is likely more realistic than using nearest-neighbor interpolation method (Phillips et al. 2006).
EVI of land cover and land uses variables
Five variables were created by calculating mean EVI for each class of land cover/use referred to above. These variables were also resampled to 100 × 100 m by a bilinear resampling technique.
Model building
MaxEnt
We used MaxEnt (Phillips et al. 2006) to model the spatial distribution of the European badger. The MaxEnt algorithm uses presence-only data. This is an advantage when working with a very low density of target species at large scales, as we expected in the study area based on Lara-Romero et al. (2012), due to the uncertainty in absences. Although MaxEnt has been criticized on several occasions (see recently Veloz 2009; Yackulic et al. 2012), it is widely used for modeling the spatial distribution of species for various purposes, e.g., testing model performance against other methods (Elith et al. 2006) and using several types of variables (Buermann et al. 2008), predicting species richness or diversity (Graham and Hijmans 2006), or forecasting distributions to estimate variations with climate change/land transformation (Yates et al. 2010). Finally, given that (1) the main goal of this study is to test the performance of models using ecosystem functional variables, and (2) prediction maps generated by MaxEnt are of interest as assessment tools, but are not the goal itself, we considered MaxEnt a valid tool for achieving our objectives.
Models
To test the utility of environmental functional variables in modeling the spatial distribution of the badger, we combined the twenty variables into four groups, with and without including ecosystem functional variables (Table 1). We defined these four groups because they were the most ecologically reasonable and of interest for comparison in keeping with the objectives of this study. These groups of variables, along with the badger presence data, were input to compute models. We used 10-fold cross-validation of the occurrence locations. Each partition was made by randomly selecting 75 % of the occurrence locations as training data, and the remaining 25 % as test data. Then, each one of the partitions, along with each of the four combinations of variables, was run in MaxEnt to compute the models. We made 10 random partitions rather than a single one in order to assess the average model behavior, and to allow for statistical testing of observed differences in performance (Phillips et al. 2006).
Model evaluation
Threshold-independent evaluation
We evaluated the performance of models created from different combinations of variables using all discriminating thresholds within the predicted area as suitable or unsuitable for badgers. We used (threshold-independent) receiver operating characteristic (ROC) analysis for this, as it uses a single measure, the area under the curve (AUC), to show model performance. With presence-only data, the AUCPO (i.e., AUC estimated with presence-only data) maximum was less than 1 (Wiley et al. 2003), so we do not know how close to optimal a given AUCPO was. Nevertheless, we were able to determine the statistical significance of the AUCPO and compare the performance of different models (Phillips et al. 2006). We employed a DeLong test (DeLong et al. 1998) to compare AUCPO values for each combination of variables. The DeLong test is designed to nonparametrically compare the difference between two AUCs from two correlated ROC curves. The Z score is defined as the difference of AUC divided by its standard error. Under the null hypothesis (the difference in AUC is zero) Z has a standard normal distribution (Chen et al. 2013). This test was computed in R (R Development Core Team 2008).
Information criteria
Following Warren and Seifert (2011), we implemented an Akaike information criterion corrected for small sample size (AICc) (Burnham and Anderson 2002) in the MaxEnt models. We standardized raw scores for each model, so that all scores within the study area added up to 1. Then we calculated the likelihood of the data in each model by taking the product of the suitability scores for each pixel showing presence. Both training and test data were used in calculating likelihood. The number of parameters was measured by counting all parameters with a nonzero weight in the .lambda file produced by MaxEnt. All AICcs were computed using ENMTools software (Warren et al. 2010).
Variable relative importance and response curves
We evaluated the relative importance of the variables using a jackknife test on the AUCPO found from test data. Thus AUCPO was estimated by (1) removing the corresponding variable, and then creating a model with the remaining variables, (2) creating a model using each variable alone, and (3) using all variables. Furthermore, we plotted the response curves for the variables which caused the widest variations in the AUCPO. Curves were estimated by generating a model using only the corresponding variable and disregarding those remaining (Phillips et al. 2006).
Results
Occurrence of European badger
The field survey yielded 94 presence locations, mainly associated with the two main rivers in the study area (see Appendix 1). Landscapes near the rivers had a larger supply of food resources for the European badger, because crops are abundant there (Fig. 1). These presence records are enough for this study since MaxEnt algorithm has been proved to works well at different sample size (Hernández et al. 2006). 51 of the records were footprints, 26 latrines, 15 setts and 2 road casualties.
Threshold-independent test
In 6 of the 10 partitions, combinations with all variables (ALL) yielded the models with the highest AUCPO (Table 2). In 8 of the 10 partitions, the AUCPO was higher for EVI and EVI LC than for the Land cover & Land uses models, which were the lowest in most of the partitions.
Information criteria
Table 3 shows Akaike weights found by models. It is accepted that models with AICc differences (∆AICc) <2 are plausible while models with ∆AICc values >10 are rejectable (Burnham and Anderson 2002). Thus, 6 of the 10 data partitions accepted EVI and LC & LU as the most parsimonious models, while one of the partitions accepted the EVI LC model. ALL models were not plausible in any of the partitions.
Relevant variables and their effects
We only analyzed the relative importance of variables from the ALL model, with the maximum AUCPO value. Area of mosaic crop (SMOCROP) caused a 2 % reduction in AUCPO (Fig. 2b). Therefore, this variable, along with others that caused a reduction of over 1 % (Area of scattered scrub (SSCRUB), EVI of mosaic crop (MOCEVI), mean maximum temperature (MMT) and coefficient of variation of mean EVI (EVICV)), provided the most useful information not present in the other variables. We considered reductions about 2 and 1 % as relevant, because these percentages were above the third quartile (0.84 %) of reduction values percentage. EVIMEAN alone had the highest AUCPO (87.4 % AUCPO with all variables) (Fig. 2a) and therefore, this variable provided the most useful information by itself. Apart from this, others like EVI spring, EVI autumn, EVI scattered scrub and standard deviation of mean EVI, were over 79 %.

Jackknife test of variable importance for European badger in the ALL model with maximum AUCPO. a Bars show the AUCPO with each variable modeled separately. Ratios above the bars show the AUCPO percentage of the reference value (0.831); b Bars show the AUCPO, when each variable is extracted from the model. The ratios above the bars show the ratio decreased by the AUCPO with respect to the reference value (0.831)
Variables such as scattered scrub area, mean maximum temperature, standard deviation of mean EVI and EVI of scattered scrub exerted a nonlinear effect on European badger habitat suitability, as predicted by MaxEnt (Appendix 2). On the contrary, mosaic crop area and mean annual EVI, exerted a positive linear effect, while EVI crop mosaic and coefficient of variation of mean EVI had a negative linear effect.
Discussion
Did the EVI-derived variables improve ecological niche modeling of the European badger in arid landscapes?
EVI variables provided useful information that improved the ecological niche modeling of European badger in arid Mediterranean landscapes. Based on the AUCPO and AICc criteria, models built with EVI variables, performed well in predicting the spatial distribution, while models without them were inferior (based on AUCPO). We suggest that the variables included in the EVI models underlie the spatiotemporal dynamic of badger food resources by describing vegetation productivity (EVIMEAN), seasonality (EVICV) and spatial heterogeneity (EVISTD). Thus, areas with high EVI mean and EVI spatial heterogeneity represented more suitable habitats for the European badger, while they rejected areas with high EVI seasonality.
Our study showed that despite the fact that rainfall (expressed here as mean annual rainfall, MRAIN) is considered the main driver of vegetation growth in Mediterranean environments (Nemani et al. 2003), it did not prove to be as good a predictor as the mean EVI (as proxy of primary productivity) for European badger distribution. The higher performance of EVI mean can be explained by the findings of Cabello et al. (2012b), in which productivity derived from EVI in drylands reflects the variation of the water use efficiency and its availability due to the features of vegetation and lithology. In addition, EVI mean also reflected the NPP for irrigated crops, which do not depend directly on rainfall (33 % of crops in the study area are irrigated).
Additionally, more seasonal environments in the study area (i.e., with high EVICV values) represented zones with low habitat quality for European badger. Johnson et al. (2002), suggested that badger densities across Europe are associated with seasonal constraints, or some other constraint(s) that covary with seasonality. EVI models predicted as suitable, landscapes with little annual variation in EVI values, corresponding with sites where the availability of food may be assured even in summer, the season experiencing the most extreme shortages in food. Similarly, Virgós and Casanovas (1999a) showed that a decrease in summer rainfall reduces badger occurrence in Mediterranean mountains.
We also found that badgers selected areas with high EVI spatial heterogeneity. Pita et al. (2009) described the Mediterranean rural landscape as a shifting mosaic that benefits diversity and presence of species as the European badger. The different types of traditional crops, along with patches of semi-natural vegetation, especially scrub and/or forest, yield a wide variety of food resources. We argue that the EVISTD variable might detect these mosaic landscapes. However, although this variable contributed positively to European badger habitat suitability, its effect was nonlinear, suggesting that badgers would not need such heterogeneity to survive in certain landscapes.
Both EVICV and EVISTD might depict variability of resources availability. EVICV represents temporal variability in the availability of resources because it is the dispersion of mean EVI throughout the year. In this sense, if EVI in summer and winter are significantly different, the annual temporal variability of EVI will be large. On the other hand, EVISTD represents spatial variability because it is the standard deviation of mean EVI into the potential territories of badgers. In consequence, high values indicate that a landscape will be more heterogeneous.
Was the predicted spatial distribution across arid lands consistent with the ecological preferences of the European badger?
The distribution predicted by the EVI models was coherent with the habitat preferences described for the European badger (see Appendix 3 for further details of predicted distributions by models). Our results reveal that badger′s presence in the study area was mainly associated with sites near rivers where there were several different types of crops and patches of natural vegetation. According to Lara-Romero et al. (2012), in Mediterranean drylands the European badger prefers mosaic landscapes consisting of fruit orchards and natural vegetation, which provide shelter and food resources. In these environments, the diet is diversified, with consumption of fruit increasing in some seasons (Barea-Azcón et al. 2010). Fruits, insects and vertebrates have also been described as relevant food resources for European badger in Mediterranean environments (Rodríguez and Delibes 1992; Revilla and Palomares 2002). Likewise, other authors have related the occurrence or abundance of these items with satellite-derived vegetation indices, such as EVI or Normalized Difference Vegetation Index (NDVI) (see Willems et al. 2009; Lafage et al. 2013; Tapia et al. 2013).
EVI and EVI for Land cover models discriminated better between suitable (i.e., mosaic landscapes with crops) and unsuitable areas (homogeneous patches of dense xerophytic scrubs) than the LC & LU models (see Table 2 and Appendix 3). EVI variables provided information for discriminating between two patches with the same type of land use and cover, but with different primary production, seasonality and spatial heterogeneity. The EVI for Land cover models exhibited an intermediate performance (Table 2). These models also used variables related to primary productivity. However, such variables were averaged based on the spatial classification derived by GIS cartography. These maps may not represent relevant landscape features for the target species or inadequate spatial resolution (Pearce et al. 2001).
Sites with high EVIMEAN and SMOCROP (area of mosaic crops) values represented the most suitable habitats for the European badger. However, the variable EVI mean of mosaic crops (MOCEVI) showed a negative effect on badger presence, which could be explained by the fact that 1—mosaic crop variable, in turn, encompasses different types of crops, and 2—badger presence records with high EVI values, are associated with non-irrigated almond crop, which would not favor badger presence in those areas. This suggests that in particular landscapes, the type of land use would be more decisive for badger than its associated productivity.
Removal of variables such as SWCROP (area of woody crop) and AEVI (EVI autumn), did increase performance, meaning that such variables reduced the generality of the model. This is, models made with these variables appear to be less transferable to other geographic areas or to projected future distributions by applying future conditions (Phillips 2006).
Regarding the potential bias of the selected study area on results, we consider that the study area contained enough variability to ensure that its effect was minimized. Probably, a larger buffer would provide similar results because the area between both rivers has not crops. In Mediterranean arid landscapes, the major landscape variability is generally associated with areas near rivers (Corbacho et al. 2003) and along altitudinal gradients, just what we defined with our study area.
Ecosystem functional dimension in species ecological modeling and conservation
The incorporation of remotely sensed characterization of the ecosystem functional dimension in management and monitoring of species and populations is gaining attention in conservation biology (Cabello et al. 2012a). Ecosystem functional dimension provides proxies showing biodiversity patterns and new tools and criteria that can assist in designing conservation planning and actions. Some examples are shown by Bardsen and Tveraa (2012), who used vegetation productivity estimated by EVI to advance knowledge of the reproductive biology of reindeer (Rangifer tarandus) in Norway; Oindo (2002) who predicted mammal species richness and abundance using multi-temporal NDVI data; or Wiegand et al. (2008), who studied the relationship between brown bear (Ursus arctos) habitat quality and the seasonal course of NDVI as a proxy for ecosystem functioning in the northern Iberian Peninsula.
Ecological modeling of the European badger in the Iberian Peninsula has to date been addressed mainly using landscape structural variables estimated from visual field observation (transect scale) (Virgós and Casanovas 1999b) and by GIS information (regional scale) (Rosalino et al. 2004). Even though these variables that reflect landscape structure are essential to modeling the species distribution (Rosalino et al. 2008), they do not reflect the role of ecosystem functioning indicators or their bidirectional relationship with the conservation of biodiversity and ecosystem processes (Cabello et al. 2012a). However, Pettorelli et al. (2005) and García-Rangel and Pettorelli (2013) point out some constrains of remote sensing data to wildlife studies such as select the most suitable processing to eliminate noise in the data, insufficient temporal resolution to precisely date phenological phenomena, and economic disadvantages due to many satellites still produce data that are not free.
Our study is the first to show that incorporation of ecosystem functional variables (EVI-derived) improves the prediction of spatial distribution modeling of the European badger in arid landscapes, considered especially sensitive to Global Change (Lavorel et al. 1998). In this sense, Pettorelli et al. (2005) suggested that satellite-derived indexes, such NDVI or EVI, could be used to predict the ecological effects of environmental change on ecosystems functioning and animal population dynamics and distributions, due to their correlation with vegetation biomass and relationship with climate variables.
Finally, we found that EVI variables represented relevant ecological parameters for the description of the distribution of the European badger as they can indicate (1) a high NPP associated with orchards or fruit crops, very important for its survival in Mediterranean arid landscapes (Rodríguez and Delibes 1992; Lara-Romero et al. 2012), (2) seasonality in the primary production, which can be seen as a surrogate of habitat quality (Johnson et al. 2002), and (3) spatially heterogeneous landscapes which provide different food resources (Pita et al. 2009). However, these variables should be tested in other areas of its distribution range. Models including EVI variables perform better (based on AUCPO) than models not including these variables. Additionally, continuous availability, both spatially and temporally, of remote sensing data can improve the accuracy of monitoring and modeling wildlife for conservation purposes in arid ecosystems throughout the world.
References
Alcaraz D, Paruelo JM, Cabello J (2006) Current distribution of ecosystem functional types in the Iberian Peninsula. Glob Ecol Biogeogr 15:200–210
Alcaraz-Segura D, Paruelo JM, Epstein HE, Cabello J (2012) Environmental and human controls of ecosystem functional diversity in temperate South America. Remote Sens 5(1):127–154
Bardsen BJ, Tveraa T (2012) Density-dependence vs. density-independence—linking reproductive allocation to population abundance and vegetation greenness. J Anim Ecol 81:364–376
Barea-Azcón JM, Ballesteros-Duperón E, Gil-Sánchez JM, Virgós E (2010) Badger Meles meles feeding ecology in dry Mediterranean environments of the southwest edge of its distribution range. Acta Theriol 55(1):45–52
Boyce MS (1978) Climatic variability and body size variation in the muskrast (Ondatra zibethicus) of North America. Oecologia 36:1–19
Brown JH, Mehlman DW, Steven GC (1995) Spatial variance in abundance. Ecology 76:2028–2043
Buermann W, Saatchi S, Smith TB, Zutta BR, Chaves JA, Milá B, Graham CH (2008) Prediction species distributions across the Amazonian and Andean regions using remote sensing data. J Biogeogr 35(7):1160–1176
Burnham KP, Anderson DR (2002) Model selection and multimodel inference. A practical information-theoretic approach. Springer, New York
Cabello J, Fernández N, Alcaraz-Segura D, Oyonarte C, Piñeiro G, Altesor A, Delibes M, Paruelo JM (2012a) The ecosystem functioning dimension in conservation: insights from remote sensing. Biodivers Conserv 21:3287–3305
Cabello J, Alcaraz-Segura D, Ferrero R, Castro AJ, Liras E (2012b) The role of vegetation and lithology in the spatial and inter-annual response of EVI to climate in drylands of Southeastern Spain. J Arid Environ 79:76–83
Chen W, Samuelson FW, Gallas BD, Kang L, Sahiner B, Petrik N (2013) On the assessment of the added value of new predictive biomarkers. BMC Med Res Methodol 13:98
Corbacho C, Sánchez JM, Costillo E (2003) Patterns of structural complexity and human disturbance of riparian vegetation in agricultural landscapes of a Mediterranean area. Agric Ecosyst Environ 95:495–507
DeLong ER, DeLong DM, Clarke-Pearson DL (1998) Comparing the areas under two or more correlated receiver operating characteristic curves: a non-parametric approach. Biometrics 44:837–845
Elith J, Leathwick JR (2009) Species distribution models: ecological explanation and prediction across space and time. Annu Rev Ecol Evol Syst 40:677–697
Elith J, Graham CH, Anderson RP, Dudík M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton JMM, Peterson AT, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberón J, Williams S, Wisz MS, Zimmermann NE (2006) Novel methods improve prediction of species′ distributions from occurrence data. Ecography 29:129–151
Ferguson SH, McLoughlin PD (2000) Effect of energy availability, seasonality and geographic range on brown bear life history. Ecography 23:193–200
Ferrier S, Watson G (1997) An Evaluation of the effectiveness of environmental surrogates and modeling techniques in predicting the distribution of biological diversity. Environment Australia, Canberra Australia. http://www.environment.gov.au/archive/biodiversity/publications/technical/surrogates/
García-Rangel S, Pettorelli N (2013) Thinking spatially: the importance of geospatial techniques of carnivore conservation. Ecol Inform 14:84–89
Graham CH, Hijmans RJ (2006) A comparison of methods for mapping species ranges and species richness. Glob Ecol Biogeogr 15:578
Hernández PA, Graham CH, Master LL, Albert DL (2006) The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 29:773–785
Huete AR, Liu HQ, Batchily K, van Leeuwen W (1997) A comparison of vegetation indices global set of TM images for EOS-MODIS. Remote Sens Environ 59:440–451
Huete AR, Didan K, Miura T, Rodriguez EP, Gao X, Ferreira LG (2002) Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens Environ 83:195–213
Jepsen JU, Madsen AB, Karlsson M, Groth D (2005) Predicting distribution and density of badger (Meles meles) setts in Denmark. Biodivers Conserv 14:3235–3253
Johnson DD, Jetz W, Macdonald DW (2002) Environmental correlates of badger social spacing across Europe. J Biogeogr 29:411–425
Kruuk H (1989) The social badger: ecology and behaviour of a group-living carnivore (Meles meles). Oxford University Press, Oxford
Lafage D, Secondi J, Georges A, Bouzillé J-B, Pétillon J (2013) Satellite-derived vegetation indices as surrogate of species richness and abundance of ground beetles in temperate floodplains. Insect Conserv Divers. doi:10.1111/icad.12056
Lara-Romero C, Virgós E, Escribano-Ávila G, Mangas JG, Barja I, Pardavila X (2012) Habitat selection by European badgers in Mediterranean semi-arid ecosystems. J Arid Environ 76:43–48
Lavorel S, Canadell J, Rambal S, Terradas J (1998) Mediterranean terrestrial ecosystems: research priorities on global change effects. Glob Ecol Biogeogr Lett 7:157–166
Macdonald DW (1983) The ecology of carnivore social behaviour. Nature 301:379–384
Macdonald DW, Carr GM (1999) Food security and the rewards of tolerance. In: Standen V, Foley R (eds) Comparative socioecology: the behavioural ecology of humans and animals, vol. 8. Blackwell Scientific, Oxford, pp 75–79
Macdonald DW, Newman C (2002) Population dynamics of badgers (Meles meles) in Oxford shires, U.K.: numbers, density and cohort life histories, and a possible role of climate change in population growth. J Zool Lond 256:121–138
Martonne E (1926) Areisme et indice d′aridité. Geogr Rev 17:397–414
Meynard CN, Pillay N, Perrigault M, Caminade P, Ganem G (2012) Evidence of environmental niche differentiation in the striped mouse (Rhabdomys sp.): inference from its current distribution in southern Africa. Ecol Evol 2(5):1008–1023
Monteith JL (1981) Evaporation and surface temperature. R Meteorol Soc 107:1–27
Nemani RR, Keeling CD, Hashimoto H, Jolly WM, Piper SC, Tucker CJ, Myneni RB, Running SW (2003) Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 300:1560–1563
Newton-Cross G, White PC, Harris S (2007) Modelling the distribution of badgers Meles meles: comparing predictions from field-based and remotely derived habitat data. Mamm Rev 37(1):54–70
Nilsen EB, Herfindal I, Linnell JD (2005) Can intra-specific variation in carnivore home-range size be explained using remote-sensing estimates of environmental productivity? Ecoscience 12(1):68–75
Oindo BO (2002) Predicting mammal species richness and abundance using multi-temporal NDVI. Photogram Eng Remote Sens 68(6):623–629
Pearce JL, Cherry K, Drielsma M, Ferrier S, Whish G (2001) Incorporating expert opinion and fine-scale vegetation mapping into statistical models of faunal distribution. J Appl Ecol 38:412–424
Pettorelli N, Vik JO, Mysterud A, Gaillard J-M, Tucker CJ, Stenseth NC (2005) Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol Evol 20(9):503–510
Pettorelli N, Gaillard JM, Mysterud A, Duncan P, Stenseth NC, Delorme D, Laere GV, Toïgo C, Klein F (2006) Using a proxy of plant productivity (NDVI) to track animal performance: the case of roe deer. Oikos 112:565–572
Pettorelli N, Ryan S, Mueller T, Bunnefeld N, Jedrzejewska B, Lima M, Kausrud K (2011) The Normalized Difference Vegetation Index (NDVI): unforeseen successes in animal ecology. Clim Res 46:15–27
Phillips SJ (2006) A brief tutorial on MaxEnt. AT & T Research. http://www.cs.princeton.edu/~schapire/maxent/tutorial/tutorial.doc
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259
Pita R, Mira A, Moreira F, Morgado R, Beja P (2009) Influence of landscape characteristics on carnivore diversity and abundance in Mediterranean farmland. Agric Ecosyst Environ 132:57–65
R Development Core Team (2008) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. http://www.R-project.org
Revilla E, Palomares F (2002) Spatial organization, group living and ecological correlates in low-density populations of Eurasian badgers, Meles meles. J Anim Ecol 71:497–512
Revilla E, Palomares F, Delibes M (2000) Defining key habitats for low density populations of Eurasian badgers in Mediterranean environments. Biol Conserv 95:269–277
Rodríguez A, Delibes M (1992) Food habits of badgers (Meles meles) in an arid habitat. J Zool 227:347–350
Rosalino LM, Macdonald DW, Santos-Reis M (2004) Spatial structure and land cover use in a low density Mediterranean population of Eurasian badgers. Can J Zool 82:1493–1502
Rosalino LM, Santos MJ, Beiber P, Santos-Reis M (2008) Eurasian badger habitat selection in Mediterranean environments: does scale really matter? Mamm Biol 73:189–198
Running SW, Thornton PE, Nemani R, Glassy JM (2000) Global terrestrial gross and net primary productivity from the earth observing system. In: Sala O, Jackson R, Mooney H (eds) Methods in ecosystem science. Springer, New York, pp 44–57
Sims DA, Rahman AF, Cordova VD, El-Masri BZ, Baldocchi DD, Flanagan LB, Goldstein AH, Hollinger DY, Misson L, Monson RK, Oechel WC, Schmid HP, Wofsy SC, Xu L (2006) On the use of MODIS EVI to asses gross primary productivity of North American ecosystem. J Geophys Res 111:G04015
Tapia L, Domínguez J, Regos A, Vidal M (2013) Using remote sensing data to model European wild rabbit (Oryctolagus cuniculus) occurrence in a highly fragmented landscape in northwestern Spain. Acta Theriol. doi:10.1007/s13364-013-0169-2
Veloz SD (2009) Spatially autocorrelated sampling falsely inflates measures of accuracy for presence-only niche models. J Biogeogr 36:2290–2299
Virgós E, Casanovas JG (1999a) Environmental constraints at the edge of a species distribution, the Eurasian badger (Meles meles L.): a biogeographic approach. J Biogeogr 6:559–564
Virgós E, Casanovas JG (1999b) Badger Meles meles sett site selection in low density Mediterranean areas of Central Spain. Acta Theriol 44(2):173–182
Virgós E, Revilla E, Domingo-Roura X, Mangas JG (2005) Conservación del tejón en España: síntesis de resultados y principales conclusiones. In: Virgós E, Revilla E, Mangas JG, Domingo-Roura X (eds) Ecología y conservación del tejón en ecosistemas mediterráneos. Sociedad Española para la Conservación y Estudio de los Mamíferos (SECEM), Málaga, pp 283–294
Wang T, Ye X, Skidmore AK, Toxopeus AG (2010) Characterizing the spatial distribution of giant pandas (Ailuropoda melanoleuca) in fragmented forest landscape. J Biogeogr 37:865–878
Warren DL, Seifert SN (2011) Ecological niche modeling with Maxent: the importance of model complexity and the performance of model selection criteria. Ecol Appl 21(2):335–342
Warren DL, Glor RE, Turelli M (2010) ENMTools: a toolbox for comparative studies of environmental niche models. Ecography 33:607–611
Wiegand T, Naves J, Garbulsky MF, Fernández N (2008) Animal habitat quality and ecosystem functioning: exploring seasonal patterns using NDVI. Ecol Monogr 78(1):87–103
Wiley EO, McNyset KM, Peterson AT, Robins CR, Stewart AM (2003) Niche modeling and geographic range predictions in the marine environment using a machine-learning algorithm. Oceanography 16(3):120–127
Willems EP, Barton RA, Hill RA (2009) Remotely sensed productivity, regional home range selection, and local range use by an omnivorous primate. Behav Ecol 20:985–992
Woodroffe R (1995) Body condition affects implantation date in the European badger, Meles meles. J Zool 236:183–188
Woodroffe R, Macdonald DW (1995) Female/female competition in European badgers Meles meles: effects on breeding success. J Anim Ecol 64:12–20
Yackulic CB, Chandler R, Zipkin EF, Royle JA, Nichols JD, Campbell Grant EH, Veran S (2012) Presence-only modelling using MAXENT: when can we trust the inferences? Methods Ecol Evol 4(3):236–243
Yates CJ, McNeill A, Elith J, Midgley GF (2010) Assessing the impacts of climate change and land transformation on Banksia in the South West Australian Floristic Region. Divers Distrib 16:187–201
Acknowledgments
J.R-M received funding from the Centro Andaluz para la Evaluación y Seguimiento del Cambio Global (CAESCG). The Oklahoma Biological Survey provided support for AJC. Funding was also received from the Andalusian Government (Projects GLOCHARID and SEGALERT P09–RNM-5048), the ERDF, andthe Ministry of Science and Innovation (Project CGL2010-22314, subprogram BOS, National Plant I + D + I 2010).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Requena-Mullor, J.M., López, E., Castro, A.J. et al. Modeling spatial distribution of European badger in arid landscapes: an ecosystem functioning approach. Landscape Ecol 29, 843–855 (2014). https://doi.org/10.1007/s10980-014-0020-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-014-0020-4