
Abstract
Ongoing declines in biodiversity caused by global environmental changes call for adaptive conservation management, including the assessment of habitat suitability spatiotemporal dynamics potentially affecting species persistence. Remote sensing (RS) provides a wide-range of satellite-based environmental variables that can be fed into species distribution models (SDMs) to investigate species-environment relations and forecast responses to change. We address the spatiotemporal dynamics of species’ habitat suitability at the landscape level by combining multi-temporal RS data with SDMs for analysing inter-annual habitat suitability dynamics. We implemented this framework with a vulnerable plant species (Veronica micrantha), by combining SDMs with a time-series of RS-based metrics of vegetation functioning related to primary productivity, seasonality, phenology and actual evapotranspiration. Besides RS variables, predictors related to landscape structure, soils and wildfires were ranked and combined through multi-model inference (MMI). To assess recent dynamics, a habitat suitability time-series was generated through model hindcasting. MMI highlighted the strong predictive ability of RS variables related to primary productivity and water availability for explaining the test-species distribution, along with soil, wildfire regime and landscape composition. The habitat suitability time-series revealed the effects of short-term land cover changes and inter-annual variability in climatic conditions. Multi-temporal SDMs further improved predictions, benefiting from RS time-series. Overall, results emphasize the integration of landscape attributes related to function, composition and spatial configuration for improving the explanation of ecological patterns. Moreover, coupling SDMs with RS functional metrics may provide early-warnings of future environmental changes potentially impacting habitat suitability. Applications discussed include the improvement of biodiversity monitoring and conservation strategies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Despite the increasing number of conservation initiatives, the rate of biodiversity loss does not appear to be diminishing, nor do the pressures upon species and their habitats (Butchart et al. 2010). Recent quantitative scenarios of biodiversity change consistently indicate that this decline will continue throughout the twenty first century, with global environmental changes and land-use shifts driving alterations in terrestrial ecosystems (Pereira et al. 2010). As a response, the Aichi Biodiversity Targets for 2011–2020 were devised by parties of the Convention for Biological Diversity (CBD 2010) and supported the definition of new conservation targets for 2020 in Europe (European Union 2011). Measuring progress towards these goals is therefore crucial and requires robust and long-term monitoring data on status and trends to assess habitat changes potentially impacting biodiversity (Magurran et al. 2010).
Effective local-scale conservation requires an accurate identification of habitat areas supporting species at higher risk of decline, as well as an assessment of how land management constrains or contributes to maintain viable populations of wild species (Prendergast et al. 1999). As such, identifying the drivers determining species distributions is at the core of applied ecological research aimed to prevent further loss of biodiversity (Butchart et al. 2010). Species distribution models (SDMs) combine observations of species occurrence or abundance with the spatial representation of environmental factors to predict species distributions (Elith and Leathwick 2009; Guisan and Zimmerman 2000). They are widely used to describe patterns, to deliver spatiotemporal predictions at several scales (Elith and Leathwick 2009) and to address fundamental questions such as the ecological impacts of climate and land-use changes (Broennimann et al. 2006). SDMs have also shown their potential for management and conservation of vulnerable and rare species, for which distribution data are often scarce, biased and/or incomplete (Gogol-Prokurat 2011; Guisan et al. 2006; Lomba et al. 2010; Sousa-Silva et al. 2014).
Remote sensing (RS) has strongly contributed to improve ecosystems and biodiversity monitoring as well as the development of SDMs (Bradley and Fleishman 2008; He et al. 2015; Nagendra et al. 2013; Skidmore et al. 2015). By providing repeated and synoptic measures of the Earth surface, RS offers a cost-effective approach to measure biodiversity and predict changes in species composition (Rocchini et al. 2016). Technologies based on RS can also provide data on habitat quantity and quality, with positive feedbacks for conservation management (Mairota et al. 2015; Vaz et al. 2015). RS data products, available at multiple spatial and temporal resolutions, from multi- or hyperspectral sensors, as well as, light detection and ranging (LiDAR) and RADAR missions can further improve SDM performance and form the backbone for next-generation SDMs (He et al. 2015). Often, satellite data are used in the form of spectral vegetation indices, which are mathematical combinations of two or more spectral bands selected to describe the biophysical parameters of interest (Jones and Vaughan 2010). The normalised difference vegetation index (NDVI) is one of the most widely used vegetation indices in ecological applications (Pettorelli et al. 2014). NDVI is regarded as a proxy of chlorophyll content in plants, or vegetation vigour, and indirectly of aboveground net primary production, absorbed photosynthetically active radiation (Kerr and Ostrovsky 2003) and leaf area index (Wang et al. 2005).
Currently available time-series of satellite data with high-temporal resolutions (such as those available from the MODerate Resolution Imaging Spectroradiometer, MODIS) greatly enhance the ability to assess and monitor both inter-annual (e.g., trends, landscape change) and intra-annual (e.g., seasonal changes, phenology) aspects of vegetation and ecosystem dynamics (Cabello et al. 2012; Heumann et al. 2007; Pettorelli et al. 2014). Various metrics of vegetation functioning, either related with overall productivity/biomass (e.g., maximum annual value, relative range) or with phenology (e.g., dates of the start, end and peak of the growing-season), can be derived from NDVI time-series (Alcaraz et al. 2006; Jönsson and Eklundh 2004). These metrics were successfully used, for example, to model tree species distribution (Cord et al. 2014), to assess phenological changes (Jönsson et al. 2010), and to predict breeding bird species richness (Coops et al. 2009). Besides vegetation indices, other RS-based variables have been used for characterizing vegetation response, such as evapotranspiration (Pôças et al. 2013). MODIS products currently available also include an actual evapotranspiration dataset (Mu et al. 2011) with high-temporal resolution, which can also be used in SDM applications.
RS ability to inform on ecosystem and landscape dynamics is a valuable asset to anticipate changes in the status of threatened species and habitats (Cabello et al. 2012). For vulnerable species, integrative approaches combining RS and SDM can improve inference and prediction at the landscape level (Parviainen et al. 2013). Although much effort has been employed in developing methods to spatially predict habitat suitability for vulnerable and/or rare plant species at multiple spatial scales (Gogol-Prokurat 2011; Lomba et al. 2010), the effects of short-term environmental fluctuations on habitat suitability, distribution and abundance of those species are seldom addressed.
Here we describe a framework combining SDMs, landscape structural metrics and a time-series of remotely-sensed proxies of vegetation functioning to predict inter-annual variations in habitat suitability at the landscape level. The approach was implemented with Veronica micrantha, an Iberian endemic plant species with “vulnerable” conservation status and protected under European and National law. Our overarching goals were: (i) to assess and rank the predictive ability of RS variables related to vegetation functioning and actual evapotranspiration, as well as of variables related to landscape structural features, fire regime and soils to explain the distribution of the test-species, and to combined them through model-averaging; (ii) to evaluate temporal variations in habitat suitability by performing model hindcasting and thereby generating a time-series of habitat suitability; (iii) to assess the added-value of this time-series of habitat suitability by comparing the model performance of single-year versus multi-temporal averaged predictions; and (iv) to explore potential causes for inter-annual changes in habitat suitability related to alterations in precipitation, temperature, and land cover/use. Finally, we discuss the potential of our framework to improve the monitoring of vulnerable species and to support local conservation planning and management, while discussing potential caveats and perspectives for future developments and applications.
Methods
Study area and test-species distribution
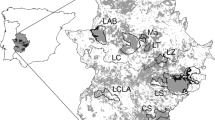
The study area is located in northern Portugal bounded by 7.785°W–7.471°W longitude and 41.391°N–41.640°N latitude. It comprises the full extent of Vila Pouca de Aguiar municipality with an area of roughly 437 km2 (Fig. 1). The elevation ranges from 225 m a.s.l. in the valley areas to 1200 m in the mountain tops (Supplementary material S1).

Main features of the study-area (a) and its location in mainland Portugal (b) and in Europe (c). (Color figure online)
V. micrantha Hoffmanns & Link (Scrophulariaceae; hereafter V. micrantha) is a perennial herbaceous plant with flowering typically occurring from May to July. Pollination is commonly entomophilous and the plant does not display a specific strategy for seed dispersal. It is an endemic species which distribution is restricted to the central- and north-western Iberian Peninsula (Sousa-Silva et al. 2014). In Portugal it occurs in the northern and centre regions, with approximately 500 reported individuals (CEC 2009). Our study-area encompasses some of the most central and large populations, hence its significance for the conservation of the species. The species has been reported to occur in open-spaces of deciduous woodland landscape matrices, heaths and herbaceous communities of forest fringes and riparian galleries, preferring shaded biotopes with humid soils (Peraza Zurita 2011; Sousa-Silva et al. 2014). Under the rainier conditions of the temperate Atlantic climate, V. micrantha also occurs in ruderal areas along rural tracks with forb vegetation.
Listed in the Habitats Directive (92/43/EEC of 21 May 1999, Annex II), V. micrantha is considered an important asset by both European and National laws. However, due to low number of individuals, fragmented subpopulations, and recorded decreasing trends in population size, V. micrantha is listed as Vulnerable in the 2011 IUCN Red List of Vascular Plants, with forest exploitation and conversion, and roads highlighted as main threats (Bilz et al. 2011; Peraza Zurita 2011).
Field data collection and sampling
SDMs were used to increase sampling efficiency and detectability as suggested by recent research (Guisan et al. 2006; Le Lay et al. 2010). Pre-existing V. micrantha distribution data for northern Portugal, collected mostly from herbaria sources, totalling 38 presence records represented in a 1 km2 regular grid, were used to develop a preliminary presence-only MaxEnt model (Phillips and Dudík 2008). A good predictive accuracy was obtained with the preliminary model (AUC = 0.81; Supplementary material S2). This model was spatially projected and used to select 32 sampling locations in the study-area, with selection probabilities proportional to habitat suitability which allowed maximizing detection probability.
Field campaigns to collect new data were performed during Spring and early Summer of 2011. An approximately linear transect along each 1 km2 quadrat diagonal was followed with controlled time for standardizing surveying effort proportionally to the landscape heterogeneity. Different land cover categories, occurring throughout the transect path, were surveyed to maximize the coverage of different vegetation and environmental conditions. When required, specific locations/habitats outside the diagonal where surveyed if the probability of occurrence was presumably high, following the assessment by experienced field botanists and supported by ancillary GIS data for the study-area. During fieldwork, five locations which were considered inaccessible for complete surveying were discarded. A total of 27 records were available after field surveys including 20 presence records and 7 absences.
Multi-model inference–hypothesis definition and variable selection
Multi-model inference (MMI), based on measures such as the Akaike information criterion (AIC), enables comparing and ranking multiple competing models (Burnham et al. 2011; Burnham and Anderson 2002; Symonds and Moussalli 2011). MMI requires an a priori definition of multiple competing hypotheses, ideally fewer than the sample size (Burnham et al. 2011). In this study, competing hypotheses were supported by previous research, literature review (Table 1 and, Supplementary material S3 for further details), expert-knowledge, as well as observations during in-field surveys regarding V. micrantha ecological requirements and distribution.
Predictor variables used in model development
The selection of predictor variables focused on landscape level ecological determinants of the test-species distribution related to vegetation functioning dynamics (VFD), landscape composition (percentage cover of certain land cover types) and configuration (spatial arrangement of landscape elements), water availability, soil types, and fire regime (Table 1). Modelling procedures were performed on a regular grid with 1 km2 units, following the available resolution for some of the predictor variables as well as for V. micrantha records. To examine multicollinearity effects, predictors were tested for pair-wise correlations using Spearman’s correlation coefficient and variance inflation factors (VIF; Supplementary material S4, S5). All pairwise correlations had an absolute value below 0.6 and thus predictors were all kept for subsequent analyses. In addition, no \(\sqrt {VIF} > 2\) was found in competing models signalling low to moderate multicollinearity.
Predictor variables were categorized into three groups: static, semi-dynamic, and dynamic. In the static group (Table 1), we considered those that either varied over geologic time (soil types, river density) or that were based on the analysis of a fixed time period (e.g., fire regime). In both cases, only a single block was used to cover the entire time-frame being analysed. For the distribution of soil types, the percentage cover of cambisols for each grid unit was calculated (PCAMB). To capture the indirect proximal effects of rivers on soil moisture and nutrient content levels we calculated the density of main rivers (RDENS, in m km−2). To analyse fire regime effects, data from the National Forest Fires Database (AFN 2011) was used to calculate the average area burned per year, between 2000 and 2009, for each grid unit (ABURN, in m2). Finally, to portray the temporal tendency of burned area, we used the trend slope (BTRND, in m2 year−1) for the same period using Sen-Theil’s method (Sen 1968).
Dynamic variables tend to display moderate to strong temporal fluctuations, as in the case of variables related to VFD and evapotranspiration. For dynamic variables, we considered a number of blocks equal to the number of time-steps, totalling 10 years for the interval 2001–2010. VFD metrics, calculated from NDVI time-series, were used as proxy measures of vegetation properties related to productivity (e.g., maximum or mean annual value), seasonality (e.g., intra-annual range) and phenology (e.g., start or end of the growing-season) potentially related to V. micrantha habitat requirements at the landscape level. The computation of VFD metrics was based on Jönsson and Eklundh (2004), using free and open-access products derived from MODIS satellite data. Composites of 16-days of MODIS NDVI (MOD13Q1 product), with spatial resolution of 250 m, were used for each year of the 2001–2010 studied period. The double-logistic function-fitting method was applied to reduce noise effects on data, generating a smoothed NDVI curve for each year which was used to calculate VFD metrics (Eklundh and Jönsson 2010; Heumann et al. 2007). These two procedures were performed using the TIMESAT software (Jönsson and Eklundh 2004). Based on their intrinsic ecological properties and on exploratory statistical analysis (not shown), four VFD metrics were selected as predictor variables. All metrics were multiplied by 104 and used as integers in modelling procedures. The maximum value for the fitted function during the growing season (MAXVL) and seasonal amplitude (AMPLT) were selected to represent primary productivity. The time for the start of the growing-season (START, in days) and the time for the mid of the growing-season (zonal maximum; MIDSN, in days) were selected to represent phenology. All VFD variables were up-scaled to the same spatial resolution of species records (1000 m) using the mean (for AMPLT and START) or the maximum (for MAXVL and MIDSN). Finally, to characterize water availability we used the MODIS MOD16A3 actual evapotranspiration (EVPTR in mm.year−1) product, computed according to Mu et al. (2011) with a spatial resolution of 1000 m comprising the studied period between 2001 and 2010.
Semi-dynamic variables comprised those exhibiting low to moderate temporal variability or which are unavailable for all the time-steps used for modelling (e.g., land cover). In this case, two blocks covering the periods 2001–2005 and 2006–2010 were considered. To analyse the effects of landscape structure on the test-species distribution, several landscape pattern statistics were calculated for each grid unit. A modified version of Corine Land Cover (Supplementary material S6) available for mainland Portugal for years 2000 and 2006 was used. For landscape composition we calculated the percentages covered by agroforestry mosaics (PAGFO) and discontinuous low-density urban–rural areas (PURBD). For capturing landscape spatial configuration, median patch size (MEDPS, in m2) and the total length of patch edges (TEDGE, in m) were used.
For each previously specified hypotheses we devised a single model (Table 1; Supplementary material S3) as recommended in Burnham et al. (2011). In this step we also linked each predictor variable to its corresponding hypothesis/model thus providing a basis for model fitting procedures. For baseline comparison purposes we used an intercept-only model (M8).
Model development
Species occurrence (presence/absence) was related to predictor variables for the reference/calibration year of 2010 (Fig. 2) using Generalized Additive Models (GAMs). GAMs are an extension of Generalized Linear Models recognized as a powerful and versatile method (Guisan et al. 2002; Guisan and Zimmerman 2000) due to their ability to include non-linear and asymmetric responses in species-environment relations. For this purpose, we used the R package mgcv (Wood 2006), which includes a penalized regression spline approach with automatic smoothness selection (Wood 2004), to reduce overfitting problems. A maximum basis dimension equal to 2 was set for smooth terms, and bivariate models were combined through multi-model averaging, as described Lomba et al. (2010) to further prevent overfitting.

Flowchart representing model development to evaluate the spatiotemporal dynamics of habitat suitability and to address the added-value of multi-temporal data
For model comparison and ranking we used Akaike Information Criterion with a correction for finite sample size (AICc), following Symonds and Moussalli (2011). In order to rank competing models, ΔAICc was calculated to build four groups discriminated by their support to explain V. micrantha distribution patterns viz., substantial support: ΔAICc ≤ 2; some support: 2 < ΔAICc ≤ 7; little support: 7 < ΔAICc ≤ 10; no support: ΔAICc > 10 (Burnham and Anderson 2002). Multi-model averaging was performed using the “natural averaging” procedure (sensu Burnham and Anderson 2002) combining all models within the confidence set, i.e., for which ∆AICc < 2 through a weighted-average using Akaike weights.
In order to fully evaluate the competing models as well as the averaged-model, MMI procedures were integrated with holdout cross-validation (Fig. 2). To perform the holdout cross-validation, 500 evaluation rounds were implemented with a data split of 70 % for model training and 30 % for testing, and balancing the number of presences and absences. After these rounds we estimated the AICc using the mean (across all rounds) and from this other MMI parameters were calculated, namely: ΔAICc and the Akaike weights (wi).
Competing models goodness-of-fit was assessed through Nagelkerke’s generalized coefficient of determination (R2) and deviance (D2) averaged across all holdout cross-validation rounds.
To assess performance of competing and averaged models, the area under the receiver operating curve (AUC) and the true-skill statistic (TSS) were calculated for the test set.
In order to transform predicted probabilities into suitable/unsuitable areas we used a threshold minimizing the straight-line distance between the receiver operating curve plot and the upper-left corner of the unit square (Freeman and Moisen 2008).
Inter-annual analysis of habitat suitability
To assess the added-value of including multi-temporal RS variables with SDM, two types of model-averaged predictions (MAP) were tested: (i) single-year predictions using data solely for the calibration year of 2010; and, (ii) a multi-temporal mean, by model hindcasting and then averaging predictions across the entire 2001–2010 time-series. This was accomplished by replacing dynamic and semi-dynamic predictors at each time-step (Table 1; Fig. 2). These two types of MAPs were then evaluated and compared through holdout cross-validation for the same test sets used for assessing competing models.
Considering all areas predicted as suitable at least one time between 2001 and 2010, linear regression and Pearson’s pairwise correlation (ρ) were used to investigate the association between inter-annual anomalies in RS variables (MAXVL, EVPTR) and anomalies in average temperature of the coldest month and total annual precipitation (collected from E-OBS, www.ecad.eu/E-OBS/; and IPMA, https://www.ipma.pt; visited Dec-2014). We also analyzed the relation between anomalies in RS variables (MAXVL, AMPLT and EVPTR) on the amount of area predicted as suitable for each year. Anomalies were calculated by subtracting the median value (between 2001 and 2010) to annual values. The effect of disturbances, such as motorway construction and wildfires, on VFD metrics and actual evapotranspiration was also evaluated by comparing areas that were either affected or unaffected by those disturbances.
Results
Performance of competing models
As measured by Deviance (D2) and R-squared (R2) statistics, which yielded very similar results, individual competing models explained variation in sample data reasonably well, with models M1 (D2 and R2 of 0.55 and 0.66, respectively), M6 (0.51, 0.65) and M7 (0.53, 0.66) obtaining the best scores (Table 2). These results were generally consistent with the AICc ranking, with exceptions for M5 (0.29, 0.55) and M3 (0.44, 0.62), although comparisons must be restricted due to the different degrees of freedom in competing models. The AUC evaluation (average value across all test sets) displayed relatively similar results, with competing models performance going from fair (M2, M3, M4, M5, M7, AUC = 0.74–0.79) to good (M1, AUC = 0.81 and M6, AUC = 0.85). The intercept-only model (M8) revealed no predictive ability regardless of the performance statistic.
Multi-model inference: model ranking and averaging
MMI results (Table 2) revealed high uncertainty from competing models, thus supporting the use of model-averaging procedures with Akaike weights [wi = (0, 1)] defining the relative contribution of each competing model to averaged predictions. In decreasing order of support, competing models M1 (VFD-productivity; ΔAICc = 0.00, wi = 0.25), M6 (Water availability; ΔAICc = 0.42, wi = 0.20), M5 (Soils; ΔAICc = 0.61, wi = 0.18), M7 (Fire disturbance; ΔAICc = 1.25, wi = 0.13) and M3 (Landscape composition; ΔAICc = 1.59, wi = 0.11) obtained substantial support and were thus included in the confidence set, later used for multi-model averaging. Competing models M2 (VFD-phenology; ΔAICc = 2.76, wi = 0.06) and M4 (Landscape configuration; ΔAICc = 3.50, wi = 0.04) obtained comparatively much less support. The intercept-only model (M8) obtained the lowest support (ΔAICc = 4.53, wi = 0.03).
Regarding the model-averaged predictions, although good results were attained for single-year predictions (AUC = 0.88, TSS = 0.82), the inclusion of RS data in multi-temporal predictions further increased model accuracy (AUC = 0.95, TSS = 0.92) (Table 3).
Determinants and inter-annual fluctuations of habitat suitability at the landscape level
A substantial portion of the study-area was considered suitable for the test species, at least at one time-step of the focal period (63 %). However, when focusing on areas most often predicted, this area decreased considerably (only 21 % for the entire focal period), as well as the spatial contiguity of suitable habitat (Fig. 3). Suitable areas are generally distributed along low-elevation areas and valleys, often close to rivers or areas with high water availability, and locations with high productivity and seasonality dynamics. Suitable areas are also related to agroforestry mosaics (with dispersed and intermixed agricultural and forest patches) and discontinuous urban–rural elements with edge and ruderal habitats (Table 2; Supplementary material S7). In addition, a negative association was detected between average burned area and habitat suitability (Figs. 3, 4; Supplementary material S7).

a Number of suitable habitat predictions between 2001 and 2010 for each grid unit. Burned area is displayed in proportional circles for enabling the comparison of suitable areas against fire-prone locations; b, c study-area broader geographical context. (Color figure online)

Plot representing the inter-annual variation in total number of suitable areas predicted (black line), along with auxiliary data related to the total amount of burned area in the study-site (in hectares; orange line) and, the variation in total annual precipitation (blue line) showing 3 years with severe drought conditions: 2004, 2005 and 2007. (Color figure online)
During the studied time-frame (2001–2010) a large decrease in habitat suitability or in the amount of suitable areas was recorded for years 2005 and 2006, related to the severe drought of 2004–2005 and with the high-incidence of wildfires in 2005 (Fig. 4). Following this period, a partial recovery of habitat suitability was observed in 2007, followed by a new decrease towards 2009, an apparent lag response to the 2007 drought conditions.
Effects of inter-annual changes in climate and land cover on habitat suitability dynamics
Overall, inter-annual anomalies (Fig. 5) in remotely-sensed variables of vegetation functioning (MAXVL) and evapotranspiration (EVPTR) were positively related to anomalies in annual precipitation (MAXVL: R2 = 0.63, ρ = 0.79; EVPTR: R2 = 0.23, ρ = 0.48) and average minimum temperature of the coldest month (MAXVL: R2 = 0.38, ρ = 0.62; EVPTR: R2 = 0.49, ρ = 0.70).

Biplots displaying the association between anomalies (annual value minus the median for the whole studied period 2001–2010) in maximum value of the growing season (left column) or Actual Evapotranspiration (right column) and anomalies in total precipitation from the previous year (identified as lag +1 year; first row) and average minimum temperature of the coldest month (second row) which show the effect of inter-annual variations in climatic conditions in remotely sensed indicators. Values for RS variables represent the anomaly in each year (from 2001 to 2010) in the median across the study-area, and considering only sites predicted as suitable at least one time in 2001–2010. Rsq coefficient of determination for the regression line, Pearson cor. Pearson correlation between variables
When comparing areas affected and unaffected by changes in land cover, derived from the construction of new motorways during 2002–2006 or from wildfires, inter-annual (i.e., non-seasonal) variation in remote-sensing variables (MAXVL, AMPLT and EVPTR) showed that these variables were sensitive to changes (Fig. 6). Both types of disturbances caused strong decreases in vegetation functioning variables (MAXVL, AMPLT; Fig. 6a–d), especially during the period 2002–2007 (i.e., throughout the motorways construction period) followed by some recovery towards 2008. For wildfires, declines were mostly observed in 2004–2006, during and after large fire events occurring in 2004 and, especially, 2005. Actual evapotranspiration (EVPTR; Fig. 6e, f) was overall less sensitive to changes induced by motorway construction and more sensitive to wildfires and drought conditions (especially in the period 2004–2005, followed by a slow recovery after these years). In turn, anomalies in number of predicted suitable areas by year recorded strong positive associations to RS variables (Fig. 7; MAXVL: R2 = 0.93, ρ = 0.97; AMPLT: R2 = 0.51, ρ = 0.72; EVPTR: R2 = 0.66, ρ = 0.81).

Boxplots showing the effect of motorways (constructed during 2002–2006; left-side of the panel) and wildfires (right-side) on the temporal variation of predictor variables: seasonal amplitude (AMPLT), maximum value of the growing season (MAXVL) and actual evapotranspiration (EVPTR) for the 2001–2010 period. Plotted locations only include areas that were at least one time predicted as suitable by the averaged model. Grey boxes represent affected areas (by motorways or wildfires) while white boxes represent control/unaffected areas by both types of disturbances. Outliers were represented as circles, i.e., values lying outside the box 1.5 times the inter-quartile range

Biplots showing the association between the number of suitable areas predicted per year (one point by year between 2001 and 2010) and anomalies (calculated as the annual value minus the median for the whole studied period 2001–2010) in the maximum value of the growing season (left), seasonal amplitude (center) and actual evapotranspiration (right). Values for RS variables represent the anomaly in each year (from 2001 to 2010) in the median across the study-area, and considering only sites predicted as suitable at least one time in 2001–2010. Rsq coefficient of determination for the regression line, Pearson cor. Pearson correlation between variables
Discussion
Combining structural and functional determinants for better predictions of habitat suitability
MMI results emphasized the strong predictive ability of RS variables related to VFD (Table 2), highlighting their usefulness for developing robust SDMs and habitat suitability predictions (Bradley and Fleishman 2008; Cord et al. 2014; Zimmermann et al. 2007). Water availability, quantified from the MODIS actual evapotranspiration product, also contributed to explain landscape patterns of habitat suitability (Table 2), confirming its importance for SDMs and other ecological assessments (Fisher et al. 2011; Hawkins et al. 2003). Our results suggest that, despite the relatively coarse spatial resolution considered in the study, RS-based variables were capable of capturing vegetation functional attributes and the heterogeneity of landscape mosaics that are linked with V. micrantha suitable habitat. Typically, the species occurs in landscapes with high levels of productivity, seasonality and water availability (Supplementary material S7). High levels of productivity have been positively related to diversity (e.g., Bai et al. 2007), which matches our observations that the species is often found in species-rich and heterogeneous mosaics.
Additionally, we found a clear negative association between habitat suitability for V. micrantha and fire occurrence (average burned area per year, and its trend; see Supplementary material S7), another result with important consequences for conservation management (Driscoll et al. 2010; Gogol-Prokurat 2011).
Although with comparatively less relevance, features related to soils and to landscape composition and configuration yielded additional (and partially interrelated) contributions to explain habitat suitability (see Table 2). The distribution of agroforestry mosaics, low-density discontinuous urban–rural areas, and edge habitats were particularly relevant for V. micrantha, highlighting the importance of small-scale heterogeneous landscapes for conservation (Thornton et al. 2011). Phenological attributes of vegetation attained considerably less support for explaining the distribution of the test species (see Table 2), apparently contradicting results from other studies (e.g., Cord et al. 2014; Ivits et al. 2011) in which phenology attained higher predictive importance. To a certain extent, comparisons are hindered due to differences in the species tested. Still, this may reflect the fact that V. micrantha is associated to different vegetation types (e.g., deciduous woodlands fringes, heathlands and herbaceous communities) with a wide spectrum of vegetation phenologies’, which hampers the model ability to discriminate between suitable and unsuitable locations. Such limitations may be overcome through the inclusion of phenology metrics within modelling procedures to characterize the heterogeneity of flowering periods or different durations of the growing-season in a given landscape. Another approach would be to combine continuous vegetation functioning metrics into a discrete map of ecosystem functional types at the landscape level (Alcaraz et al. 2006). Such data could then be used to quantify landscape heterogeneity and analyse spatial patterns.
Overall, our results strongly emphasize the importance of simultaneously considering landscape attributes related to function, composition and spatial configuration to improve the explanation of ecological patterns (Vaz et al. 2015).
Habitat suitability dynamics and the conservation of vulnerable species
Based on multi-temporal remote-sensing data, our approach allowed exploring the effects of short-term fluctuations in environmental conditions on the spatiotemporal dynamics of landscape level of habitat suitability for our test-species (see Figs. 4, 5). Thus, it contributes to clarify the mechanisms underpinning species’ responses to fluctuations in habitat suitability derived from multiple and often overlapping sources of disturbance, such as land cover changes and wildfires, but also to strong inter-annual climatic variability (such as drought conditions; see Figs. 5, 6, 7). This is especially useful since these fluctuations may pose additional threats to vulnerable species with already small and highly fragmented populations and hence higher extinction risk (Bilz et al. 2011; Peraza Zurita 2011). Such added-value may be particularly useful when conservation plans are based on adaptive management strategies focused on maintaining key ecosystem processes and functions (Haney and Power 1996). In addition, by focusing on the landscape level, our approach presents several advantages for the focal species conservation namely because local and regional management frequently target this scale for planning and are typically multi-objective (e.g., Sayer et al. 2013), and also because, from a monitoring perspective, it allows to identify and assess landscape change processes and dynamics (Mairota et al. 2015; Nagendra et al. 2013) affecting habitat suitability and biodiversity.
Habitat suitability time-series can also provide useful inputs for systematic conservation planning (Pressey et al. 2007), allowing to assess and rank suitable habitat areas according to their relative stability in time. By identifying locations that have lost their suitability in a recent past due to environmental changes, our approach may help to spatially prioritize habitat protection and restoration actions with higher benefits for species persistence (Renton et al. 2012). Among other aspects, RS data could be used for monitoring the dynamics and the temporal stability of suitable locations and their habitat quality (Mairota et al. 2015; Vaz et al. 2015) and serve as early-warning system helping to identify critical changes in essential variables (Skidmore et al. 2015).
Contributions of the framework to improve the monitoring of vulnerable species
Incorporating high-temporal resolution data related to landscape-level vegetation functioning and evapotranspiration in SDMs conveyed better predictions and the ability to evaluate spatiotemporal habitat suitability dynamics. This has several obvious implications for improving the effectiveness of biodiversity monitoring (Kerr and Ostrovsky 2003; Pettorelli et al. 2014; Skidmore et al. 2015). First, this may provide some evidence that RS-based functional metrics could provide early-warnings of changes in ecosystem processes affecting habitat suitability, well before assessments based on structural metrics such as those derived from land cover maps (Bradley and Fleishman 2008; Cabello et al. 2012; Cord et al. 2014). Second, since the later typically have lower update frequencies and higher production costs (especially for moderate or high spatial resolution products such as those used here), a RS-based functional approach using freely accessible data (e.g., MODIS, Landsat, Sentinel-2) could also potentially improve the cost-efficiency of vulnerable species monitoring. Third, the spatiotemporal dynamics of habitat suitability and the RS variables used in SDMs can be employed by regional or local authorities to devise monitoring schemes to develop useful vegetation or ecosystem functioning indicators aimed to track (or even anticipate) shifts in biodiversity (Cabello et al. 2012; Nagendra et al. 2013; Pettorelli et al. 2014). Fourth, reporting on threatened species (such as V. micrantha), with periodical information requests on their conservation status (Sousa-Silva et al. 2014), could potentially benefit from the proposed framework to develop comprehensive indicators based on species’ spatiotemporal dynamics and to detail pressures affecting them. Finally, by taking advantage of an iterative and model-based design (as implemented in this study), existing data on species occurrence can be used to improve species detectability and cover data gaps especially for vulnerable and/or rare species (Guisan et al. 2006; Le Lay et al. 2010).
Using this approach, improved monitoring can also be achieved in cases where small-scale landscapes are prone to frequent and various disturbances, especially those related to fire occurrence, or then submitted to drastic changes in land cover/use (e.g., large infrastructures). In this sense, landscape-level vegetation functioning attributes derived from spectral vegetation indices time-series (e.g., NDVI) have proved useful for their capacity to provide a synthetic overview of ecosystem changes caused by multiple drivers (Brook et al. 2008). Moreover, our results show that, as expected, RS variables were sensitive to inter-annual variations in temperature and precipitation (Figs. 5, 6). The tested multi-temporal mean approach (Fig. 2; Table 3) can be used to generate improved predictions by averaging habitat suitability time-series and thereby capturing long-term trends. Moreover, analysing or aggregating multi-temporal predictions (using the mean as in this case, or other statistical measures), instead of relying on a preliminary multi-temporal aggregation of RS variables (e.g., Coops et al. 2009; Cord et al. 2014), can provide a more robust framework to evaluate shifts and trends in suitable habitat dynamics and to assess pressures affecting it (Figs. 5, 6, 7).
Limitations, sources of uncertainty, and future prospects
As in any practical application of models in conservation and management, uncertainties underlying models and their predictions should be carefully handled and efficiently communicated to stakeholders (Guisan et al. 2013). These uncertainties mainly derive from caveats that vulnerable and rare species distributions pose to model development, due to the frequently small occurrence datasets (Lomba et al. 2010; Wiser et al. 1998). Overfitting may also be an issue, however model-averaging parsimonious bivariate models in a MMI framework, similarly to Lomba et al. (2010), may prove beneficial.
Temporal (Tuanmu et al. 2011) as well as spatial transferability (Randin et al. 2006) also pose some limitations that require careful inspection. Here, given the short time-span of this study, niche conservatism was assumed. In a recent review, Peterson (2011) found that ecological niche characteristics are strongly preserved over short to moderate time-frames. However, to fully evaluate temporal transferability, historical time-series on species occurrence would be required (not available in our case) coupled with appropriate methods (e.g., Rapacciuolo et al. 2014). Given the relatively small extent of the study-area [compared to the full species range in Iberia (Sousa-Silva et al. 2014)], spatial transferability may be more problematic, reducing model forecast ability for other regions (Randin et al. 2006). As such, our results for V. micrantha are mostly applicable to the test-area, but the framework can be easily generalized to its whole range as more data becomes available to fully encompass the species environmental niche.
RS based variables also pose some challenges in a modelling context due to various ‘noise’ sources (Hird and McDermid 2009; Jönsson and Eklundh 2004). However, appropriate pre-processing methods—such as geometric, atmospheric, as well as smoothing/filtering routines (Hird and McDermid 2009; Jönsson and Eklundh 2004)—can be used to minimize those problems. Therefore, the selection of an appropriate algorithm for time-series smoothing and its parametrization are crucial aspects to adequately fill data gaps and filter noise from undetected clouds or poor atmospheric conditions (Kandasamy et al. 2013).
Additional tests implementing the approach presented here are required, including other species with different life-history traits, ranges and habitat attributes, involving sensitivity analyses as well as methods to cope with model uncertainties. Besides species presence/absence, other parameters could also be modelled with RS multi-temporal indices, such as abundance, habitat quality or population fitness. This will further improve the integration of multi-temporal RS data in SDMs as a way to inform and advance biodiversity monitoring and conservation.
References
Alcaraz D, Paruelo J, Cabello J (2006) Identification of current ecosystem functional types in the Iberian Peninsula. Glob Ecol Biogeogr 15:200–212. doi:10.1111/j.1466-822X.2006.00215.x
Autoridade Florestal Nacional (National Forestry Authority). http://www.afn.min-agricultura.pt/portal/dudf/cartografia/cartograf-areas-ardidas-1990-2009. Accessed 10 Jan 2011. Cartografia nacional de áreas ardidas (national mapping of burnt areas)
Bai Y et al (2007) Positive linear relationship between productivity and diversity: evidence from the Eurasian Steppe. J Appl Ecol 44:1023–1034. doi:10.1111/j.1365-2664.2007.01351.x
Bilz M, Kell SP, Maxted N, Lansdown RV (2011) European red list of vascular plants. European Union, Luxembourg
Bowman DMJS, Murphy BP (2010) Fire and biodiversity. In: Sodhi NS, Ehrlich PR (eds) Conservation Biology for All. Oxford University Press, New York, pp 163–180
Bradley BA, Fleishman E (2008) Can remote sensing of land cover improve species distribution modelling? J Biogeogr 35:1158–1159. doi:10.1111/j.1365-2699.2008.01928.x
Broennimann O, Thuiller W, Hughes G, Midgley GF, Alkemade JMR, Guisan A (2006) Do geographic distribution, niche property and life form explain plants’ vulnerability to global change? Glob Change Biol 12:1079–1093. doi:10.1111/j.1365-2486.2006.01157.x
Brook BW, Sodhi NS, Bradshaw CJA (2008) Synergies among extinction drivers under global change. Trends Ecol Evol 23:453–460. doi:10.1016/j.tree.2008.03.011
Burnham KP, Anderson DR (2002) Model selection and multi-model inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Burnham K, Anderson D, Huyvaert K (2011) AIC model selection and multimodel inference in behavioral ecology: some background, observations, and comparisons. Behav Ecol Sociobiol 65:23–35. doi:10.1007/s00265-010-1029-6
Butchart SHM et al (2010) Global biodiversity: indicators of recent declines. Science 328:1164–1168. doi:10.1126/science.1187512
Cabello J et al (2012) The ecosystem functioning dimension in conservation: insights from remote sensing. Biodivers Conserv 21:3287–3305. doi:10.1007/s10531-012-0370-7
CBD (2010) Strategic plan for biodiversity 2011–2020 and the Aichi targets. CBD, Montreal
CEC (2009) Composite report on the conservation status of habitat types and species as required under article 17 of the habitats directive, COM(2009) 358 final. Commission of European Communities, Brussels
Coops NC, Wulder MA, Iwanicka D (2009) Exploring the relative importance of satellite-derived descriptors of production, topography and land cover for predicting breeding bird species richness over Ontario, Canada. Remote Sens Environ 113:668–679. doi:10.1016/j.rse.2008.11.012
Cord AF, Klein D, Mora F, Dech S (2014) Comparing the suitability of classified land cover data and remote sensing variables for modeling distribution patterns of plants. Ecol Model 272:129–140. doi:10.1016/j.ecolmodel.2013.09.011
Driscoll DA et al (2010) Fire management for biodiversity conservation: key research questions and our capacity to answer them. Biol Conserv 143:1928–1939. doi:10.1016/j.biocon.2010.05.026
Eklundh L, Jönsson P (2010) TIMESAT 3.0. Software Manual. Lund University, Lund
Elith J, Leathwick J (2009) Species distribution models: ecological explanation and prediction across space and time. Annu Rev Ecol Evol Syst 40:677–697
European Union (2011) The EU biodiversity strategy to 2020. European Union, Luxembourg. doi:10.2779/39229
Fisher JB, Whittaker RJ, Malhi Y (2011) ET come home: potential evapotranspiration in geographical ecology. Glob Ecol Biogeogr 20:1–18. doi:10.1111/j.1466-8238.2010.00578.x
Freeman EA, Moisen GG (2008) A comparison of the performance of threshold criteria for binary classification in terms of predicted prevalence and kappa. Ecol Model 217:48–58. doi:10.1016/j.ecolmodel.2008.05.015
Gogol-Prokurat M (2011) Predicting habitat suitability for rare plants at local spatial scales using a species distribution model. Ecol Appl 21:33–47. doi:10.1890/09-1190.1
Guisan A, Zimmerman NE (2000) Predictive habitat distribution models in ecology. Ecol Model 135:147–186
Guisan A, Edwards TC Jr, Hastie T (2002) Generalized linear and generalized additive models in studies of species distributions: setting the scene. Ecol Model 157:89–100. doi:10.1016/s0304-3800(02)00204-1
Guisan A, Broennimann O, Engler R, Vust M, Yoccoz NG, Lehmann A, Zimmermann NE (2006) Using niche-based models to improve the sampling of rare species. Conserv Biol 20:501–511. doi:10.1111/j.1523-1739.2006.00354.x
Guisan A et al (2013) Predicting species distributions for conservation decisions. Ecol Lett 16:1424–1435. doi:10.1111/ele.12189
Haney A, Power R (1996) Adaptive management for sound ecosystem management. Environ Manag 20:879–886. doi:10.1007/BF01205968
Hawkins BA et al (2003) Energy, water, and broadscale geographic patterns of species richness. Ecology 84:3105–3117. doi:10.1890/03-8006
He KS et al (2015) Will remote sensing shape the next generation of species distribution models? Remote Sens Ecol Conserv 1:4–18. doi:10.1002/rse2.7
Heumann BW, Seaquist JW, Eklundh L, Jönsson P (2007) AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982–2005. Remote Sens Environ 108:385–392. doi:10.1016/j.rse.2006.11.025
Hird JN, McDermid GJ (2009) Noise reduction of NDVI time series: an empirical comparison of selected techniques. Remote Sens Environ 113:248–258. doi:10.1016/j.rse.2008.09.003
Ivits E, Buchanan G, Olsvig-Whittaker L, Cherlet M (2011) European farmland bird distribution explained by remotely sensed phenological indices. Environ Model Assess 16:385–399. doi:10.1007/s10666-011-9251-9
Jones HG, Vaughan RA (2010) Remote sensing of vegetation. Principles, techniques, and applications, 1st edn. Oxford University Press, Oxford
Jönsson P, Eklundh L (2004) TIMESAT—a program for analyzing time-series of satellite sensor data. Comput Geosci 30:833–845. doi:10.1016/j.cageo.2004.05.006
Jönsson AM, Eklundh L, Hellström M, Bärring L, Jönsson P (2010) Annual changes in MODIS vegetation indices of Swedish coniferous forests in relation to snow dynamics and tree phenology. Remote Sens Environ 114:2719–2730. doi:10.1016/j.rse.2010.06.005
Kandasamy S, Baret F, Verger A, Neveux P, Weiss M (2013) A comparison of methods for smoothing and gap filling time series of remote sensing observations—application to MODIS LAI products. Biogeosciences 10:4055–4071. doi:10.5194/bg-10-4055-2013
Kerr JT, Ostrovsky M (2003) From space to species: ecological applications for remote sensing. Trends Ecol Evol 18:299–305
Le Lay G, Engler R, Franc E, Guisan A (2010) Prospective sampling based on model ensembles improves the detection of rare species. Ecography 33:1015–1027. doi:10.1111/j.1600-0587.2010.06338.x
Lomba A, Pellissier L, Randin C, Vicente J, Moreira F, Honrado J, Guisan A (2010) Overcoming the rare species modelling paradox: a novel hierarchical framework applied to an Iberian endemic plant. Biol Conserv 143:2647–2657. doi:10.1016/j.biocon.2010.07.007
Magurran AE et al (2010) Long-term datasets in biodiversity research and monitoring: assessing change in ecological communities through time. Trends Ecol Evol 25:574–582. doi:10.1016/j.tree.2010.06.016
Mairota P et al (2015) Challenges and opportunities in harnessing satellite remote-sensing for biodiversity monitoring. Ecol Inform 30:207–214. doi:10.1016/j.ecoinf.2015.08.006
Mu Q, Zhao M, Running SW (2011) Improvements to a MODIS global terrestrial evapotranspiration algorithm. Remote Sens Environ 115:1781–1800. doi:10.1016/j.rse.2011.02.019
Nagendra H, Lucas R, Honrado JP, Jongman RHG, Tarantino C, Adamo M, Mairota P (2013) Remote sensing for conservation monitoring: assessing protected areas, habitat extent, habitat condition, species diversity, and threats. Ecol Indic 33:45–59. doi:10.1016/j.ecolind.2012.09.014
Parviainen M, Zimmermann NE, Heikkinen RK, Luoto M (2013) Using unclassified continuous remote sensing data to improve distribution models of red-listed plant species. Biodivers Conserv 22:1731–1754. doi:10.1007/s10531-013-0509-1
Peraza Zurita MD (2011) Veronica micrantha. IUCN red list of threatened species. Version 2011.2. http://www.iucnredlist.org/apps/redlist/details/162008/0]. IUCN, http://www.iucnredlist.org/apps/redlist/details/162008/0. Accessed March 2012
Pereira HM et al (2010) Scenarios for global biodiversity in the 21st century. Science 330:1496–1501. doi:10.1126/science.1196624
Peterson AT (2011) Ecological niche conservatism: a time-structured review of evidence. J Biogeogr 38:817–827. doi:10.1111/j.1365-2699.2010.02456.x
Pettorelli N, Laurance WF, O’Brien TG, Wegmann M, Nagendra H, Turner W (2014) Satellite remote sensing for applied ecologists: opportunities and challenges. J Appl Ecol 51:839–848. doi:10.1111/1365-2664.12261
Phillips SJ, Dudík M (2008) Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31:161–175
Pôças I, Cunha M, Pereira LS, Allen RG (2013) Using remote sensing energy balance and evapotranspiration to characterize montane landscape vegetation with focus on grass and pasture lands. Int J Appl Earth Obs Geoinf 21:159–172. doi:10.1016/j.jag.2012.08.017
Prendergast JR, Quinn RM, Lawton JH (1999) The gaps between theory and practice in selecting nature reserves. Conserv Biol 13:484–492. doi:10.1046/j.1523-1739.1999.97428.x
Pressey RL, Cabeza M, Watts ME, Cowling RM, Wilson KA (2007) Conservation planning in a changing world. Trends Ecol Evol 22:583–592. doi:10.1016/j.tree.2007.10.001
Randin CF, Dirnböck T, Dullinger S, Zimmermann NE, Zappa M, Guisan A (2006) Are niche-based species distribution models transferable in space? J Biogeogr 33:1689–1703. doi:10.1111/j.1365-2699.2006.01466.x
Rapacciuolo G, Roy DB, Gillings S, Purvis A (2014) Temporal validation plots: quantifying how well correlative species distribution models predict species’ range changes over time. Methods Ecol Evol 5:407–420. doi:10.1111/2041-210X.12181
Renton M, Shackelford N, Standish RJ (2012) Habitat restoration will help some functional plant types persist under climate change in fragmented landscapes. Glob Change Biol 18:2057–2070. doi:10.1111/j.1365-2486.2012.02677.x
Reside AE, VanDerWal J, Kutt A, Watson I, Williams S (2012) Fire regime shifts affect bird species distributions. Divers Distrib 18:213–225. doi:10.1111/j.1472-4642.2011.00818.x
Rocchini D et al (2016) Satellite remote sensing to monitor species diversity: potential and pitfalls. Remote Sens Ecol Conserv 2:25–36. doi:10.1002/rse2.9
Sayer J et al (2013) Ten principles for a landscape approach to reconciling agriculture, conservation, and other competing land uses. Proc Natl Acad Sci 110:8349–8356. doi:10.1073/pnas.1210595110
Sen PK (1968) Estimates of the regression coefficient based on Kendall’s tau. J Am Stat Assoc 63:1379–1389
Skidmore AK et al (2015) Agree on biodiversity metrics to track from space. Nature 523:403–405
Sousa-Silva R, Alves P, Honrado J, Lomba A (2014) Improving the assessment and reporting on rare and endangered species through species distribution models. Glob Ecol Conserv 2:226–237. doi:10.1016/j.gecco.2014.09.011
Symonds M, Moussalli A (2011) A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav Ecol Sociobiol 65:13–21. doi:10.1007/s00265-010-1037-6
Thornton D, Branch L, Sunquist M (2011) The influence of landscape, patch, and within-patch factors on species presence and abundance: a review of focal patch studies. Landsc Ecol 26:7–18. doi:10.1007/s10980-010-9549-z
Tuanmu M-N, Viña A, Roloff GJ, Liu W, Ouyang Z, Zhang H, Liu J (2011) Temporal transferability of wildlife habitat models: implications for habitat monitoring. J Biogeogr 38:1510–1523. doi:10.1111/j.1365-2699.2011.02479.x
Vaz AS et al (2015) Can we predict habitat quality from space? A multi-indicator assessment based on an automated knowledge-driven system. Int J Appl Earth Obs Geoinf 37:106–113. doi:10.1016/j.jag.2014.10.014
Wang Q, Adiku S, Tenhunen J, Granier A (2005) On the relationship of NDVI with leaf area index in a deciduous forest site. Remote Sens Environ 94:244–255. doi:10.1016/j.rse.2004.10.006
Wever LA, Flanagan LB, Carlson PJ (2002) Seasonal and interannual variation in evapotranspiration, energy balance and surface conductance in a northern temperate grassland agricultural and forest. Meteorology 112:31–49. doi:10.1016/s0168-1923(02)00041-2
Wiser SK, Peet RK, White PS (1998) Prediction of rare-plant occurrence: a southern Appalachian example. Ecol Appl 8:909–920
Wood SN (2004) Stable and efficient multiple smoothing parameter estimation for generalized additive models. J Am Stat Assoc 99:673–686. doi:10.1198/016214504000000980
Wood S (2006) Generalized additive models: an introduction with R, 1st edn. Chapman and Hall/CRC, Boca Raton
Zimmermann NE, Edwards TC, Moisen GG, Frescino TS, Blackard JA (2007) Remote sensing-based predictors improve distribution models of rare, early successional and broadleaf tree species in Utah. J Appl Ecol 44:1057–1067. doi:10.1111/j.1365-2664.2007.01348.x
Acknowledgments
J. Gonçalves was financially supported by FCT (Portuguese Science Foundation) through PhD grant SFRH/BD/90112/2012. I. Pôças was supported by FCT through postdoctoral grant SFRH/BPD/79767/2011. B. Marcos was supported by FCT and FEDER/COMPETE (project IND_CHANGE; PTDC/AAG-MAA/4539/2012; FCOMP-01-0124-FEDER-027863). R. Sousa-Silva was supported by a PhD grant in the framework of the FORBIO Climate project, financed by BRAIN.be. A. Lomba was supported by FCT through postdoctoral grant SFRH/BPD/80747/2011.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Daniel Sanchez Mata.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Gonçalves, J., Alves, P., Pôças, I. et al. Exploring the spatiotemporal dynamics of habitat suitability to improve conservation management of a vulnerable plant species. Biodivers Conserv 25, 2867–2888 (2016). https://doi.org/10.1007/s10531-016-1206-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-016-1206-7