
Abstract
We explored the potential for using Pediastrum (Meyen), a genus of green alga commonly found in palaeoecological studies, as a proxy for lake-level change in tropical South America. The study site, Laguna La Gaiba (LLG) (17°45′S, 57°40′W), is a broad, shallow lake located along the course of the Paraguay River in the Pantanal, a 135,000-km2 tropical wetland located mostly in western Brazil, but extending into eastern Bolivia. Fourteen surface sediment samples were taken from LLG across a range of lake depths (2–5.2 m) and analyzed for Pediastrum. We found seven species, of which P. musteri (Tell et Mataloni), P. argentiniense (Bourr. et Tell), and P. cf. angulosum (Ehrenb.) ex Menegh. were identified as potential indicators of lake level. Results of the modern dataset were applied to 31 fossil Pediastrum assemblages spanning the early Holocene (12.0 kyr BP) to present to infer past lake level changes qualitatively. Early Holocene (12.0–9.8 kyr BP) assemblages do not show a clear signal, though abundance of P. simplex (Meyen) suggests relatively high lake levels. Absence of P. musteri, characteristic of deep, open water, and abundance of macrophyte-associated taxa indicate lake levels were lowest from 9.8 to 3.0 kyr BP. A shift to wetter conditions began at 4.4 kyr BP, indicated by the appearance of P. musteri, though inferred lake levels did not reach modern values until 1.4 kyr BP. The Pediastrum-inferred mid-Holocene lowstand is consistent with lower precipitation, previously inferred using pollen from this site, and is also in agreement with evidence for widespread drought in the South American tropics during the middle Holocene. An inference for steadily increasing lake level from 4.4 kyr BP to present is consistent with diatom-inferred water level rise at Lake Titicaca, and demonstrates coherence with the broad pattern of increasing monsoon strength from the late Holocene until present in tropical South America.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Pediastrum (Meyen) is a colonial green alga of the family Hydrodictyaceae. It is a potential bioindicator in palaeoenvironmental studies because: (1) it is abundant in freshwater lakes and wetlands and is often found in high concentrations in sediment cores; (2) it is globally distributed; (3) it survives the harsh chemical treatment of pollen preparations, and can therefore be analyzed along with pollen; and (4) it can be resolved to species, and often subspecies level, with the use of light microscopy (Komárek and Jankovská 2001; Van Geel 2001). Despite these strengths, the full potential of Pediastrum as a palaeoenvironmental proxy has remained unexplored.
Total Pediastrum concentrations are often recorded in palaeoenvironmental studies, but the majority of analysts do not distinguish among species, and Pediastrum concentrations are reported alongside other non-pollen palynomorphs (NPP) (Van Geel 2001). The interpretation of the “Pediastrum signal” varies geographically. In the northern mid-latitudes, fluctuations in total Pediastrum concentration are typically thought to reflect changes in trophic state and erosion in the catchment (Komárek and Jankovská 2001; Van Geel 2001). Although less commonly reported, varying Pediastrum concentrations in tropical and subtropical regions are interpreted as reflecting changes in temperature (Rull et al. 2008) and lake levels (Sylvestre 2002; Gosling et al. 2008), as well as being linked to both increased (Caballero et al. 1999; Brenner et al. 2006) and decreased (Lamb et al. 1999) macrophyte cover in lakes from the lower latitudes of the Americas and Africa, respectively.
However, given that species of Pediastrum have varying ecological tolerances (Komárek and Jankovská 2001; Van Geel 2001), qualitative and/or quantitative estimates of past environmental change might be inferred from fossil Pediastrum community composition. Studies that identified Pediastrum to higher taxonomic resolution have drawn links between community change and nutrient availability (Bradshaw et al. 2005), pH and DOC (Weckström et al. 2010), water quality (Jankovská and Pokorny 2002), and changes in physical lake variables, such as fluctuations in water levels (Kaufman et al. 2010) and littoral vegetation change (Danielsen et al. 2010).
Despite evidence that fossil Pediastrum communities reflect past limnological change, attempts to draw regional climate inferences from Pediastrum assemblages have often produced equivocal results (Nielsen and Sørensen 1992; Sarmaja-Korjonen et al. 2006; Komárek and Jankovská 2001). This is perhaps unsurprising, given that palaeoreconstructions which incorporate Pediastrum data are often focused on the large-magnitude climate events of the late glacial in the high to middle latitudes, where the landscape had been relatively recently deglaciated, and limnological change associated with landscape evolution (Engstrom et al. 2000) might mask effects of changing climate on Pediastrum assemblages. A palaeoclimate signal might, however, be inferred from Pediastrum in regions where the catchment has been relatively stable.
The aim of this study was to explore the potential of Pediastrum as a proxy for climate change in lowland, tropical South America. There are relatively few climate reconstructions across tropical South America. The majority are located in the central high Andes (Baker et al. 2001; Fritz et al. 2004) or along the Atlantic seaboard (Wang et al. 2004; Cruz et al. 2005, 2009), and palaeoclimate of the lowland interior is often extrapolated from these distant sites. Further palaeoclimate proxy development in the South American tropics is required because past climate in lowland regions is often inferred from fossil pollen, but the synergistic effects of changing precipitation, temperature and atmospheric CO2 concentrations on tropical vegetation are difficult to disentangle (Bush et al. 2007). Pediastrum has been reported in fossil pollen studies from lowland, tropical sites (Ledru et al. 2006) and these algae may provide an independent bioindicator to be used in conjunction with pollen analysis.
We present data on fossil Pediastrum community change from a site in the lowland southern hemisphere tropics of South America (SHTSA), a region for which there is widespread, coherent evidence of drought in the early to middle Holocene (Mayle and Power 2008; Whitney et al. 2011). Given that Pediastrum communities are found to vary among different lake habitats (Komárek and Jankovská 2001; Danielsen 2010; Kaufman et al. 2010), we hypothesize that lake-level change associated with a variable Holocene precipitation regime might be qualitatively inferred from varying relative abundances of open-water versus aquatic macrophyte-associated Pediastrum species.
Site description
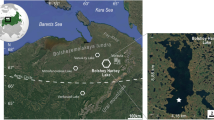
Our study site is located in the Pantanal, the world’s largest tropical wetland (~135,000 km2), located predominantly in western Brazil, but extending into easternmost Bolivia (Fig. 1). This wetland occupies a tectonic depression between the Brazilian shield and the Andean foreland (Clapperton 1993), and together with the western Brazilian highlands, forms the headwaters of the Paraguay River. Precipitation of the Pantanal is highly seasonal. Mean annual precipitation ranges from 1,000 mm in the north to 1,700 mm in the south of the Pantanal, the bulk of which falls in the austral summer, from December to February, during the South American summer monsoon. The monsoon causes flooding in low-lying areas of the Pantanal basin, and these floodwaters slowly drain into the Paraguay River system, eventually causing the Paraguay River and its associated tributaries and lakes to overflow (Alho 2005).

LLG is located along the Paraguay River, which flows through the adjacent Pantanal wetlands that experience annual flooding in the austral summer (December–January–February). Floodwaters slowly drain into the Paraguay River system, raising water levels in the wetlands, including its associated lakes (LLG) and tributaries. Inferred lake-level fluctuations at LLG are therefore representative of a broad hydrological catchment. The inset shows the position of the Pantanal wetlands, located mainly in western Brazil, but extending into easternmost Bolivia. Reproduced from Whitney et al. (2011), with permission from Elsevier
Laguna La Gaiba (LLG) (17°45′S, 57°40′W) is a large, shallow lake located along the course of the Paraguay River in the western Pantanal. Lake levels peak after the summer monsoon floodwaters have drained into the Paraguay River system, including LLG (Alho 2005). Changing hydrological conditions at this site thus reflect regional precipitation change (Hamilton 2002; Whitney et al. 2011). Maximum lake depth varies from about 4 to 6 m, depending on the season. LLG is comprised of two distinct portions: (1) the shallow northern part (~3 m deep) that merges with permanently flooded wetlands to the east and is characterized by dense mats of aquatic grasses and Eichhornia spp., and its northwestern margin is bounded by steep-sided hills covered with seasonally-dry tropical forest (SDTF) (Fig. 2). (2) The deeper, southern part of LLG is almost entirely surrounded by hills covered by SDTF. Few aquatic macrophytes line the southern shores of LLG, although Poaceae, Asteraceae, and floating mats of Eichhornia spp. have been identified. Basic analyses of lake water in July 2006 show the lake to be nutrient-limited and slightly alkaline (Table 1).

Schematic representation of the modern lake environment at LLG and the position of the 14 sites from which the topmost 1 cm of sediment was analyzed for modern Pediastrum assemblages. Site 0 is the core site. The 1-m contours were approximated from 273 points measured in July 2008 using a Lowrance Depth Sounder (X-65). Modified from Whitney et al. (2011)
Previous pollen analysis at LLG indicates that the catchment was continuously forested throughout the Holocene, thereby minimizing the influence of biome change or changes in soil erosion on the lake chemistry, and temperatures were near-to-modern from ~19.5 kyr BP until present (Whitney et al. 2011). Varying relative abundance of SDTF pollen types, however, indicates that the region experienced drought in the early to middle Holocene, coincident with the widespread drying trend in the southern hemisphere tropics of South America (SHTSA) (Mayle and Power 2008). Pediastrum results from this site can therefore be compared to independent evidence for lower-than-modern precipitation.
Materials and methods
Field
In 2001, two parallel, overlapping sediment cores were extracted from a floating platform in the deepest part of the basin using a Colinvaux-Vohnout drop-hammer piston corer (Colinvaux et al. 1999). The cores, measuring 5.6 and 4.8 m, were shipped in their aluminium core tubes and extruded in the laboratory. Extruded cores were wrapped in plastic film, aluminium foil and thick plastic sleeves, and stored in a dark room at 4°C. A surface core, comprising the uppermost 110 cm of unconsolidated sediments and overlapping the piston cores by 40 cm, was collected with a Perspex® tube and piston, and extruded in the field into air-tight plastic bags in consecutive 1-cm slices.
In July 2006, the uppermost 1 cm of sediment was collected at 19 sites across LLG (Fig. 2) using a Perspex® tube and piston. Samples were extruded in the field and stored in airtight plastic bottles. Eight 500-mL water samples were also collected from three different environments within LLG (shallow, macrophyte-dominated lake margins; deep, open water; river inflow/outflow), and basic water chemistry and limnological variables were measured at each site (Table 1). Because of the remoteness of the field site and the lack of appropriate storage for water samples in a hot climate, nitrite/nitrate concentrations were measured in the field using a Hach® field testing kit. The remaining water of each sample (~350 mL) was filtered through a 10-μm Nitex mesh and the residues were washed into air-tight plastic bottles to collect water-borne Pediastrum assemblages. Lugol’s iodine solution and 90% ethanol were added to the samples to preserve the algal communities.
Chronology and stratigraphy
The parallel piston cores were correlated by magnetic susceptibility and cross-checked through analysis of pollen at zonal boundaries. The surface core was correlated to the consolidated sediments of the piston cores by loss-on-ignition (550°C) analysis. The entire sediment core from LLG extends to ~45,000 years BP, the chronology of which is based on 18 AMS 14C dates analyzed by the NERC radiocarbon facility, East Kilbride, UK (Fig. 3). AMS 14C ages (±2σ) were calibrated using the “Fairbanks0107” calibration curve (Fairbanks et al. 2005), and the age-depth model was created using mixed-effect regression of all 18 dates (Heegaard et al. 2005). The Holocene section of the core is constrained by five dates (Table 2), including those obtained from bulk sediments sampled from strata in which carbonate content was negligible, estimated through loss-on-ignition at 1,000°C (Dean 1974). Holocene sediments are uniformly grey-brown clay with fine organic matter. Full details of chronology and stratigraphy are described in Whitney et al. (2011).

The entire age-depth model for LLG is based on 18 AMS 14C dates and an assumed modern age for the uppermost 1 cm of sediment. Dates were calibrated using the Fairbanks calibration program (Fairbanks et al. 2005) and the relationship between age and stratigraphic position of dates was modelled using mixed-effect regression as described by Heegaard et al. (2005). The shaded area marks the section of core analyzed for this study, and black outer lines represent 2σ error. “x” denotes the rejected date. Reproduced from Whitney et al. (2011) with permission from Elsevier
Laboratory
Pediastrum assemblages were analyzed from Holocene-age sediments that were previously prepared and analyzed for pollen (Whitney et al. 2011) according to standard chemical digestion protocol (Fægri and Iversen 1989). Thirty-one samples were analyzed at approximately 500-yr resolution from 12.0 kyr BP until present. To further understand modern Pediastrum species distributions and taphonomy at LLG, 1 cm3 of sediment from the 19 surface sediment samples collected across the lake (Fig. 2) were also processed and analyzed for Pediastrum. Modern and fossil Pediastrum identifications were made with reference to Komárek and Jankovská (2001), Pasztaleniec and Poniewozik (2004), and Zamaloa and Tell (2005). Algae were identified to species and variety levels, although frequent poor preservation of fossil Pediastrum and the lack of voucher specimens prompted us to report only species level identification, thereby minimizing the possibility of misidentification. Photographs of commonly occurring Pediastrum species were taken using a Canon® 350D SLR digital camera attached to a triocular Olympus® BX40 microscope using a 2.5× Olympus® phototube.
Statistics and data presentation
Pediastrum species abundance was calculated as a proportion of the sum of all identified Pediastrum in each sample. A minimum of 30 identifiable coenobia (cell colonies) was counted in each sample, which equates to the approximate abundance of Pediastrum coenobia per 300 terrestrial pollen grains in the LLG sediment samples. Pediastrum concentrations are generally much lower here than at high-latitude sites, where as many as 500 coenobia per sample have been reported (Bradshaw et al. 2005; Sarmaja-Korjonen et al. 2006), most likely because of the limited nutrient availability in lowland tropical lakes, including LLG.
Redundancy analysis (RDA), a constrained ordination technique that combines Principal Component Analysis (PCA) and regression, was performed on square-root-transformed surface Pediastrum assemblages, with water depth as the constraining environmental variable. The significance of the constraining axis was determined using a permutation test (anova.cca) (Borcard et al. 2011). RDA was chosen over CCA because of the short environmental gradient exhibited in the modern dataset (ter Braak 1995). RDA was performed on a correlation matrix to emphasize the rarer taxa, particularly those that occur only in one habitat type. All ordination analyses were performed using Vegan package 1.17-12 (Orksanen et al. 2010) for R (version OS X GUI 1.40). Proportions of individual Pediastrum species in the modern surface samples are plotted against water depth in Fig. 7, and the relationship between water depth and species abundance was modelled using a generalized linear model with a binomial error structure to account for the boundedness of the response variable and the non-normal distribution of errors in proportion data (Crawley 2005).
Spatial autocorrelation of the surface Pediastrum assemblages was tested following a modified version of the simple procedure described in Telford and Birks (2009). Analysis began with removal of one of the modern surface samples, and the remaining dataset was ordered by geographic distance in the lake to the chosen sample. Modern surface samples were sequentially deleted by order of closest geographic neighbour until the dataset was reduced to three samples. After each deletion, constrained ordination analyses were performed (both CCA and RDA), using water depth as the constraining variable, and the eigenvalues of the constrained axis were used as a measure of ‘model performance’ after each deletion. The test was repeated for each of the remaining 13 samples. In addition to the ‘nearest neighbour’ deletions, we also determined model performance after a series of random deletions, in which constrained ordination (CCA and RDA) was performed after sequentially deleting random samples until only three modern samples remained. The test was repeated on 100 randomized datasets. Given that model performance did not deteriorate more rapidly for ‘nearest neighbour’ deletions compared with random deletions, we conclude that our modern dataset is not spatially autocorrelated (Telford and Birks 2009), and we therefore do not present those results here.
The fossil Pediastrum dataset was zoned using optimal splitting by information content, and three significant zonal boundaries were recognized using a broken-stick model (Bennett 1996). Principal component analysis (PCA) (correlation matrix) was also performed on fossil Pediastrum assemblages (Fig. 8) to aid the biostratigraphic interpretation in the fossil diagram (Kovach 1995) (Fig. 9). Plots of Pediastrum species abundance in surface samples, and stratigraphic fossil Pediastrum plots, were constructed using C2 (Juggins 2003).
Results
Modern Pediastrum
The attempt to collect Pediastrum from lake water samples was unsuccessful. No Pediastrum coenobia, or fragments identifiable as Pediastrum, were recovered. Algal concentrations were shown to be correlated inversely with river stage in the Paraguay River system, including associated lakes such as LLG (de Oliveira and Calheiros 2000), and lake and river levels were high at the time of sampling (July 2006). Pediastrum concentrations were probably too low to permit recovery of a sizeable number of coenobia in 350 mL of lake water, as a consequence of the season in which fieldwork was conducted.
In contrast, Pediastrum coenobia were recovered from 14 surface sediment samples, including the core top (Fig. 4). The remaining five samples, taken near the river inflow and outflow, or from very shallow water, contained negligible concentrations of Pediastrum coenobia and were excluded from analysis. P. simplex (Meyen) (Fig. 5a) was the most abundant taxon in all samples (50–76%). P. musteri (Tell et Mataloni) (Fig. 5b) was also present in all samples (5–30%), and this species was most abundant in samples from the deeper, central and southern parts of the lake, with the exception of site 3 where it represented only 8%. P. musteri occurred at lowest abundance at the two shallowest sites (15, 16). P. boryanum (Turp.) Menegh. (Fig. 5c) was present in eight assemblages, and did not demonstrate a strong preference for any particular part of the lake, occurring in low abundance (3–10%) in samples 3, 5, 7, 8, 12, 13, and 15, with the exception of site 16, where it was the second most abundant taxon, reaching a value of 18%. P. duplex (Meyen) (Fig. 5d) was present in low abundance at eight sites across the lake and occurred in highest abundance at site 3 (15%). P. argentiniense (Bourr. et Tell) (Fig. 5e) occurred in seven modern assemblages, the majority of which were sampled from shallow to moderate water depths, typically located in the northern portion of the lake (sites 12, 15, 16) and at centrally-located sites near the river inflows and outflows (8, 10, 11). P. argentiniense also occurred at site 2 (7%) in the deeper, southern part of LLG. P. cf. angulosum (Ehrenb.) ex Menegh. (Fig. 5f) occurred in only four samples, three of which were sampled from the shallowest part of the lake (11, 15, 16). The remaining site containing P. cf. angulosum (13) was sampled from the highest measured water depth, despite its proximity to the macrophyte-dominated northeastern lake margins. P. subgranulatum (Racib.) was rare and did not demonstrate a particular habitat preference, occurring in low abundance in the core top and at sites 3, 6 and 16.

Relative percent abundance of Pediastrum taxa in surface sediments sampled from variable lake depths across LLG. Assemblages are ordered by increasing water depth

Common Pediastrum species found in the surface and fossil sediments at LLG. a P. simplex; b P. musteri; c P. boryanum; d P. duplex; e P. argentiniense; f P. cf. angulosum. Scale bars = 50 μm
Redundancy analysis (RDA) of the 14 analyzed modern samples was constrained against water depth using a correlation matrix (Fig. 6). The constrained axis (RDA1) accounts for 22% of the variance in the dataset, and the result of the permutation test indicates that Pediastrum assemblages are significantly correlated with water depth in the modern LLG environment (R 2adj = 0.16, p < 0.001). Assemblages sampled from the deepest part of the lake environment contain higher abundance of P. musteri, whereas P. argentiniense and P. angulosum exhibit positive axis 1 scores and characterize the assemblages sampled from the macrophyte-dominated shallows of the northern portion of the lake. P. boryanum is positively positioned along RDA1 (negatively correlated with water depth), but this is largely due to its high abundance at site 16, and the species proportion abundance plot of P. boryanum versus water depth (Fig. 7) does not show any clear trend.

Redundancy Analysis (RDA) of all modern Pediastrum assemblages with water depth as the constraining environmental variable (grey arrow). Refer to Fig. 2 for the position of the samples within the lake

Relative abundance of the six commonly reported Pediastrum species in the modern surface sediments of LLG, plotted against lake depth. The relationships between water depth and P. musteri, P. argentiniense, and P. cf. angulosum are described using a generalized linear model with a binomial error structure. Site 13 (open circle) was excluded from the P. cf. angulosum response model. See text for further details
PC1 accounts for 29% of the unexplained variance in the dataset, thus Pediastrum community structure is also influenced by an unmeasured environmental variable(s). The removal of an outlier sample (site 13) from the redundancy analysis increases the proportion of variance explained by RDA1 to 31%, higher than the unconstrained variance accounted for by PC1 (26%). Although site 13 is surrounded by macrophyte-dominated shallow water, the sample was extracted from within a deep river channel, where we recorded the highest water depth for this study. Thus, the assemblage comprises a mixture of both deep- and shallow-water taxa, illustrated by high abundances of P. musteri (>20%), similar to values in the deeper-water sites, and the presence of P. cf. angulosum, found to be most abundant in the shallowest sites (Fig. 7). Exclusion of site 13 from the RDA increases the R 2adj to 0.25.
Pediastrum proportion data were plotted against water depth (Fig. 7). P. argentiniense exhibits a significant negative correlation with water depth (p < 0.001), and P. musteri is positively correlated with water depth (p < 0.001) in the modern dataset. The relationship between water depth and P. cf. angulosum (p < 0.01) is significant only if site 13 is excluded from the model. Exclusion of site 13, however, does not alter the significance of the relationship between water depth and abundance of P. musteri and P. argentiniense.
Fossil Pediastrum
Zone-I: 12.0–9.8 kyr BP; 9 samples
The Pediastrum assemblages of this zone (Fig. 8), often poorly preserved, are dominated by P. simplex and P. argentiniense, reaching maximum values of 90 and 43%, respectively. P. musteri, common in the deep, southern part of the modern lake environment, is also present in low abundances. P. boryanum and P. cf. angulosum each comprise <10% of the assemblages in this zone. P. duplex has similarly low abundances, with the exception of the sample at 11.7 kyr BP, where it comprises 35% of the assemblage.

Principal Component Analysis (PCA) of fossil Pediastrum assemblages. Symbols refer to sample age
Zone-II: 9.8–3.0 kyr BP; 16 samples
Pediastrum duplex is the dominant taxon in Zone-II (17–93%) (Fig. 8). Also abundant are taxa characteristic of the shallow, macrophyte-dominated parts of the modern lake environment at LLG: P. argentiniense (5–43%) and P. cf. angulosum. The latter is present in over half the samples in this zone, reaching a maximum abundance of 10% at 7.4 and 6.7 kyr BP. P. musteri is rare or absent prior to 5.5 kyr BP, but rises to a zone maximum of 20% abundance at 4.7 kyr BP, and also occurs at the top of Zone II in samples dating from 3.5 and 3.3 kyr BP. P. simplex declines in abundance from the previous zone, and it is absent in all but one sample, occurring at 4.3 kyr BP (3%). The abundance of P. boryanum varies greatly in this zone; it is absent in five samples and reaches a maximum of 43% at 6.4 kyr BP.
Zone-III: 3.0 kyr BP to present; 6 samples
The Pediastrum assemblages of Zone-III (Fig. 8) are dominated by P. musteri and P. simplex. P. musteri abundances are highest at 1.9 kyr BP (60%) and decrease to 30% at the top of the zone. Conversely, P. simplex is rare or absent in the lower third of the zone, but comprises 50–70% of the assemblages from 1.4 kyr BP onward. P. argentiniense and P. duplex decline to <10% and <30% from the previous zone, respectively, and P. cf. angulosum all but disappears from the record. P. boryanum is present in one sample (3%) at the base of the zone, occurring at 2.8 kyr BP.
PCA of fossil assemblages
The principal component analysis (PCA) of the fossil assemblages (Fig. 8) shows negative scores for P. simplex and P. musteri along PCA1, whereas P. argentiniense and P. duplex exhibit high positive scores. PCA1 accounts for 44% of the variance in the dataset. Although the modern study does not demonstrate any relationship between water depth and P. simplex, it is correlated with P. musteri in the fossil dataset, a taxon found to be more abundant in deeper water in the surface dataset. Similarly, P. duplex and P. boryanum, commonly found in the modern assemblages, but not related to water depth in the modern lake environment, are correlated with the shallow-water indicator P. argentiniense along axis 1. P. cf. angulosum also shows positive PCA1 values, but, along with P. duplex, exhibits higher scores along PCA2, accounting for 26% of the variance in the dataset.
Discussion
Interpretation of the Pediastrum signal
Analysis of modern Pediastrum assemblages at LLG highlights P. musteri, P. argentiniense, and P. cf. angulosum as potential indicators of past lake-level fluctuations. P. musteri is shown to prefer the larger and deeper southern portion of LLG (Figs. 6 and 7) and P. argentiniense demonstrates a strong preference for the shallower, macrophyte-influenced, northern extent of the lake (Figs. 6 and 7). Although P. argentiniense has not been previously reported from fossil sediments, our results are consistent with modern studies from tropical regions of the Americas that indicate this taxon is often associated with aquatic vegetation (Komárek and Jankovská 2001). P. cf. angulosum is found in low abundance in the modern and fossil assemblages, but it may be an indicator of very low water levels because it is mostly restricted to the shallowest part of LLG, with the exception of site 13, which reflects its close proximity to the adjacent macrophyte-dominated shallows of the northeastern margin of LLG. Given our limited sampling of the shallowest portion of LLG, the interpretation of P. cf. angulosum as a shallow-water indicator must be viewed with caution. Previous studies, however, corroborate that P. angulosum is found in small water bodies and swamps, and mainly among aquatic plants (Komárek and Jankovská 2001).
Pediastrum simplex, P. duplex and P. boryanum are three abundant species in the fossil assemblages, but our limited modern study does not shed light on the habitat and/or environmental preference of these types. P. simplex is the most abundant species in the modern samples at LLG (Fig. 4), and it is reported to be found in relatively deep water (Komárek and Jankovská 2001), but the abundance of P. simplex does not vary with lake depth at LLG. It may be that the range of measured depths (2–5.2 m) is too narrow to demonstrate any response in P. simplex abundance, and this taxon is only absent from very shallow water. This hypothesis cannot be tested at LLG because sediments sampled from the shallowest portions of LLG contained very few Pediastrum coenobia, probably the result of periodic drying as a consequence of the high interannual variability of the Pantanal flooding regime (Alho 2005).
Pediastrum duplex dominates the assemblages in Zone-II, but again, the modern study at LLG did not demonstrate a relationship between water depth and abundance of this taxon. Although none of the Pediastrum taxa was reported at the taxonomic level of variety, as a consequence of the lack of reference material, the cell walls of many of the P. duplex coenobia in this zone are coarsely sculptured, consistent with descriptions of P. duplex var. rugulosum (Racib). Komárek and Jankovská (2001) report that this variety is often associated with aquatic macrophytes, the occurrence of which in Zone-II would be consistent with the high abundance of other macrophyte-associated taxa, P. argentiniense and P. cf. angulosum. Regardless of which variety of P. duplex dominates Zone-II, total P. duplex is of considerably higher abundance in the fossil record than the maximum 15% it reaches in the modern dataset, which means that there is no analogue for the fossil assemblages of Zone-II in the modern lake environment.
The PCA of the fossil dataset (Fig. 8) shows that P. simplex is correlated with the deeper-water indicator, P. musteri, and P. duplex has a positive Axis 1 score similar to P. argentiniense, shown to be a shallow-water indicator in our modern dataset. Therefore, these species might show a response to water depth in the modern dataset if a broader range of water depths were sampled. We cannot discount the possibility that they are primarily controlled by an unknown environmental variable that fluctuated throughout the Holocene, particularly because both species demonstrate high positive (P. duplex) and negative (P. simplex) scores along PCA2 in the fossil dataset. Further field sampling, and particularly, successful recovery of water-column assemblages alongside measured environmental variables, is needed to understand the ecology of these two taxa. Given that all but the late Holocene fossil assemblages do not have analogues in our modern dataset, interpretation of the P. simplex and P. duplex signals in the fossil dataset is tentative.
The P. boryanum signal in the fossil dataset is also equivocal. Although it has nine described subspecies (Komárek and Jankovská 2001), P. boryanum was not identified to subspecies level at LLG because its relatively small coenobia are highly susceptible to degradation, and the subspecies are often misidentified (Komárek and Jankovská 2001). The majority of the coenobia, however, match the descriptions for P. boryanum var. boryanum (Turp.) Menegh. and P. boryanum var. brevicorne (A.) Br. Although P. boryanum is reported to be cosmopolitan and a generalist, and therefore thought to be a poor indicator of past environmental conditions (Komárek and Jankovská 2001), its co-occurrence with macrophyte-associated taxa P. argentiniense and P. cf. angulosum in Zone-II might suggest it is associated with shallow water, but our limited modern dataset does not confirm this interpretation. Further analysis of the distribution of subspecies of this taxon, as well as those of P. duplex and P. simplex, might reveal a significant relationship with water depth, but such an analysis is beyond the scope of this study.
Holocene lake-level changes at LLG
Drawing on the results from the modern study, abundance of P. argentiniense and P. cf. angulosum, and the low relative proportions of P. musteri, imply that LLG was most shallow and swampy in the early to middle Holocene. The high proportion of macrophyte-associated taxa from 9.8 to 3.0 kyr BP suggests that lower-than-modern lake levels allowed for aquatic vegetation to encroach toward the centre of the lake, where the core site lies. The Paraguay River and associated lakes, including LLG, are hydrologically connected to the Pantanal floodplains (Hamilton 2002; Alho 2005). Past fluctuations in river and lake levels are reflective of varying precipitation across the Pantanal basin and its river headwaters (Hamilton 2002; Whitney et al. 2011). Thus, the Pediastrum-inferred lowering of lake levels at LLG is indicative of regional precipitation decline.
Although interpretation of the Pediastrum dataset is based on a limited number of surface samples, the inferred lake-level fluctuations are coincident with marked changes in the pollen record (Fig. 8). The hills surrounding LLG were continuously covered by closed-canopy dry forest during the Holocene, but changes in the relative abundance of key pollen types are indicative of drier conditions in the early to middle Holocene, as previously demonstrated at this site by Whitney et al. (2011). Moraceae, a family most abundant in moisture-dependent vegetation communities (Mayle et al. 2000; Gosling et al. 2009), declines in this zone. Drier conditions are also inferred from peak abundance of Astronium and Anadenanthera, pollen types characteristic of drought-resistant, seasonally dry tropical forest (Mayle et al. 2000; Jardim et al. 2003; Gosling et al. 2009) (Fig. 9).

Relative percent abundance of fossil Pediastrum species in sediments, calculated as a percentage of all identified Pediastrum (black bars), and dominant pollen types, calculated as a percentage of the terrestrial sum (grey bars). Also shown are PCA Axis 1 sample scores (Fig. 8), broadly reflective of relative lake depth at LLG, and freshwater diatom data from L. Titicaca (Baker et al. 2001); both curves demonstrate lower-than-modern lake levels in the early to middle Holocene
The Pediastrum-inferred lowering of lake levels in the middle Holocene and the coincident decline in moisture-dependent vegetation are corroborated by widespread evidence of significant reduction in precipitation in the early to middle Holocene across tropical South America (Mayle and Power 2008). The timing of this event varies latitudinally. The onset of drought occurs earliest at more northerly sites and progresses southward, but generally this event is centred between 8.0 and 4.0 kyr BP. Evidence of drought includes peak dust concentrations and lowered snow accumulation in the Sajama ice cap (Thompson et al. 1998), diatom- and geochemically-inferred lake-level lowering in the high central Andes (Seltzer et al. 2000; Baker et al. 2001; Tapia et al. 2003), and high erosion along the Andean piedmont (May et al. 2008). At Lake Titicaca, from 6.0 to 5.0 kyr BP, lake levels were reduced by as much as 100 m compared to present (Baker et al. 2001), and reached the lowest levels of the past 25,000 years. Quantitative climate reconstructions from Laguna Bella Vista, located ~600 km northwest of LLG, demonstrate that mean annual precipitation may have declined by as much as 500 mm in the middle Holocene, relative to present (Burbridge et al. 2004; Punyasena et al. 2008).
Also reflected in the Pediastrum record at LLG is a gradual, rather than punctuated, rise in lake level from the middle Holocene until present. P. musteri abundance increases at 4.4 kyr BP, coincident with an initial decrease in the macrophyte-associated taxon P. argentiniense. This taxon declines further to reach lowest levels from 3.0 kyr BP until present. With the appearance of P. simplex at 1.4 kyr BP, the fossil assemblages resemble the modern Pediastrum communities at LLG, reflective of deeply flooded conditions.
This gradual change is consistent with a strengthening of the South American summer monsoon through the late Holocene to present, hypothesized to have been forced by increasing (austral) summer insolation in the Southern Hemisphere tropics as a consequence of precession orbital forcing (Baker et al. 2001; Cruz et al. 2009). There is evidence for rising precipitation at Lake Titicaca, where the shift from benthic and salinity-tolerant to planktonic diatoms began at 4.5 kyr BP, but the lake did not reach modern levels until 2.0–1.5 kyr BP (Baker et al. 2001; Tapia et al. 2003). Along the Andean piedmont to the west of the Pantanal, a gradual change toward landscape stability and rising moisture availability began at 4 kyr BP, with the onset of wetter conditions occurring at 2.9 kyr BP (May et al. 2008), coincident with our inferred lake-level rise at LLG.
Conclusions
Despite a limited modern Pediastrum dataset, P. musteri, P. argentiniense, and P. cf. angulosum are highlighted as possible indicator taxa for lake-level change in lowland tropical lakes. The Holocene lake-level changes inferred from the fossil Pediastrum assemblages at LLG correspond with the pattern of changing moisture availability previously demonstrated by pollen analysis at this site (Whitney et al. 2011), and the Holocene climate trend widely reported from the southern hemisphere tropics of South America (Mayle and Power 2008).
Though a pilot study, these findings demonstrate the potential of fossil Pediastrum as a proxy for lake-level change in tropical lakes. In tropical South America, the paucity of independent palaeoprecipitation records from the lowlands means that Andean palaeoclimate records, such as the record from Lake Titicaca (Baker et al. 2001), are sometimes relied upon to explain lowland palaeo-vegetation changes (Mayle and Power 2008). A key advantage of Pediastrum as a lake-level (precipitation) proxy is that it is often abundant in lowland tropical lakes and can be analyzed along with fossil pollen on the same slides, providing the opportunity to explore vegetation-climate relationships, not only at the same site, but in the same samples. This strategy may also enable us to differentiate between human land-use versus climate change, as drivers of Holocene vegetation dynamics in the lowland tropics.
References
Alho CJR (2005) The Pantanal. In: Fraser LH, Keddy PA (eds) The World’s largest wetlands: ecology and conservation. Cambridge University Press, Cambridge
Baker PA, Seltzer GO, Fritz SC, Dunbar RB, Grove MJ, Tapia PM, Cross SL, Rowe HD, Broda JP (2001) The history of South American tropical precipitation for the past 25,000 years. Science 291:640–643
Bennett KD (1996) Determination of the number of zones in a biostratigraphical sequence. New Phytol 132:155–170
Borcard D, Gillet F, Legendre P (2011) Numerical ecology with R. Springer, New York
Bradshaw EG, Rasmussen P, Nielsen H, Anderson NJ (2005) Mid- to late-Holocene land-use change and lake development at Dallund So, Denmark: trends in lake primary production as reflected by algal and macrophyte remains. The Holocene 15:1130–1142
Brenner M, Hodell DA, Leyden BW, Curtis JH, Kenney WF, Gu BH, Newman JM (2006) Mechanisms for organic matter and phosphorus burial in sediments of a shallow, subtropical, macrophyte-dominated lake. J Paleolimnol 35:129–148
Burbridge RE, Mayle FE, Killeen TJ (2004) Fifty-thousand-year vegetation and climate history of Noel Kempff Mercado National Park, Bolivian Amazon. Quat Res 61:215–230
Bush MB, Gosling WD, Colinvaux PA (2007) Climate change in the lowlands of the Amazon basin. In: Bush MB, Flenley JR (eds) Tropical rainforest responses to climate change. Springer, Berlin, pp 53–76
Caballero M, Lozano S, Ortega B, Urrutia J, Macias JL (1999) Environmental characteristics of Lake Tecocomulco, northern basin of Mexico, for the last 50,000 years. J Paleolimnol 22:399–411
Clapperton C (1993) Quaternary geology and geomorphology of South America. Elsevier Academic Publishers, Netherlands
Colinvaux PA, De Oliveira PE, Moreno PJE (1999) Amazon pollen manual and atlas. Harwood Academic Publishers, Amsterdam
Crawley MJ (2005) Statistics: a introduction using R. Wiley, Chichester
Cruz FW, Burns SJ, Karmann I, Sharp WD, Vuille M, Cardoso AO, Ferrari JA, Dias PLS, Viana O (2005) Insolation-driven changes in atmospheric circulation over the past 116,000 years in subtropical Brazil. Nature 434:63–66
Cruz FW, Vuille M, Burns SJ, Wang XF, Cheng H, Werner M, Edwards RL, Karmann I, Auler AS, Nguyen H (2009) Orbitally driven east-west antiphasing of South American precipitation. Nat Geosci 2:210–214
Danielsen R (2010) Dissimilarities in the recent histories of two lakes in Portugal explained by local scale environmental processes. J Paleolimnol 43:513–534
de Oliveira MD, Calheiros DF (2000) Flood pulse influence on phytoplankton communities of the south Pantanal floodplain, Brazil. Hydrobiologia 427:101–112
Dean WE (1974) Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignition: comparison with other methods. J Sediment Petrol 44:242–248
Engstrom DR, Fritz SC, Almendinger JE, Juggins S (2000) Chemical and biological trends during lake evolution in recently deglaciated terrain. Nature 408:161–166
Fægri K, Iversen J (1989) Textbook of pollen analysis. Wiley, Chichester
Fairbanks RG, Mortlock RA, Chiu T-C, Cao L, Kaplan A, Guilderson TP, Fairbanks TW, Bloom AL (2005) Marine radiocarbon calibration curve spanning 0 to 50,000 years B.P. based on paired 230Th/234U/238U and 14C dates on pristine corals. Quat Sci Rev 24:1781–1796
Fritz SC, Baker PA, Lowenstein TK, Seltzer GO, Rigsby CA, Dwyer GS, Tapia PM, Arnold KK, Ku T-L, Luo S (2004) Hydrologic variation during the last 170,000 years in the southern hemisphere tropics of South America. Quat Res 61:95–104
Gosling WD, Bush MB, Hanselman JA, Chepstow-Lusty A (2008) Glacial-interglacial changes in moisture balance and the impact on vegetation in the southern hemisphere tropical Andes (Bolivia/Peru). Palaeogeogr Palaeoclimatol Palaeoecol 259:35–50
Gosling WD, Mayle FE, Tate NJ, Killeen TJ (2009) Differentiation between Neotropical rainforest, dry forest, and savannah ecosystems by their modern pollen spectra and implications for the fossil pollen record. Rev Palaeobot Palynol 153:70–85
Hamilton SK (2002) Hydrological controls of ecological structure and function in the Pantanal wetland (Brazil). In: McClain ME (ed) The Ecohydrology of South American rivers and wetlands. IAHS Press, Centre for Ecology and Hydrology, Oxfordshire, pp 133–158
Heegaard E, Birks HJB, Telford RJ (2005) Relationships between calibrated ages and depth in stratigraphical sequences: an estimation procedure by mixed-effect regression. The Holocene 15:612–618
Jankovská V, Pokorny J (2002) Palaeoecology of a medieval fishpond system (Vajgar, Czech Republic). Folia Geobot 37:253–273
Jardim A, Killeen TJ, Fuentes A (2003) Guía de los Árboles y Arbustos del Bosque Seco Chiquitano, Bolivia. Fundacíon Amigos de la Naturaleza Noel Kempff (FAN), Santa Cruz
Juggins S (2003) User guide C2. Software for ecological and palaeoecological data analysis and visualization, User guide version 1.3. University of Newcastle, Newcastle upon Tyne
Kaufman DS, Anderson RS, Hu FS, Berg E, Werner A (2010) Evidence for a variable and wet younger dryas in southern Alaska. Quat Sci Rev 29:1445–1452
Komárek J, Jankovská V (2001) Review of the green algal genus Pediastrum: Implication for pollen analytical research. Bibl Phycol 108. J Cramer Berlin 1–127
Kovach W (1995) Multivariate data analysis. In: Maddy D, Brew JS (eds) Statistical modelling of quaternary science data. Technical guide no. 5, Quaternary research association, Cambridge
Lamb H, Roberts N, Leng M, Barker P, Benkaddour A, van der Kaars S (1999) Lake evolution in a semi-arid montane environment: response to catchment change and hydroclimatic variation. J Paleolimnol 21:325–343
Ledru M-P, Ceccantini G, Gouveia SEM, Lopez-Saez JA, Pessenda LCR, Ribeiro AS (2006) Millennial-scale climatic and vegetation changes in a northern Cerrado (Northeast, Brazil) since the Last Glacial Maximum. Quat Sci Rev 25:1110–1126
May JH, Zech R, Veit H (2008) Late quaternary paleosol-sediment-sequences and landscape evolution along the Andean piedmont, Bolivian Chaco. Geomorphology 98:34–54
Mayle FE, Power MJ (2008) Impact of a drier early-mid-holocene climate upon Amazonian forests. Philos Trans R Soc Lond B Biol Sci 363:1829–1838
Mayle FE, Burbridge R, Killeen TJ (2000) Millennial-scale dynamics of southern Amazonian rain forests. Science 290:2291–2294
Nielsen H, Sorensen I (1992) Taxonomy and stratigraphy of late-glacial Pediastrum taxa from Lysmosen, Denmark—a preliminary study. Rev Palaeobot Palynol 74:55–75
Oksanen J, Blanchet FG, Kindt R, Legendre P, O’Hara B, Simpson GL, Solymos P, Stevens MHH, Wagner H (2010) vegan: community ecology package. R package version 1.17-12. Available at: www.cran.r-project.org
Pasztaleniec A, Poniewozik M (2004) Pediastrum species (Hydrodictyaceae, Sphaeropleales) in phytoplankton of Sumin Lake (Leczna-Wlodawa Lakeland). Acta Soc Bot Pol 73:39–46
Punyasena SW, Mayle FE, McElwain JC (2008) Quantitative estimates of glacial and holocene temperature and precipitation change in lowland Amazonian Bolivia. Geology 36:667–670
Rull V, Lopez-Saez JA, Vegas-Vilarrubia T (2008) Contribution of non-pollen palynomorphs to the paleolimnological study of a high-altitude Andean lake (Laguna Verde Alta, Venezuela). J Paleolimnol 40:399–411
Sarmaja-Korjonen K, Seppänen A, Bennike O (2006) Pediastrum algae from the classic late glacial Bølling Sø site, Denmark: Response of aquatic biota to climate change. Rev Palaeobot Palynol 138:95–107
Seltzer G, Rodbell D, Burns S (2000) Isotopic evidence for lateq climatic change in tropical South America. Geology 28:35–38
Sylvestre F (2002) A high-resolution diatom reconstruction between 21,000 and 17,400 14C yr BP from the southern Bolivian Altiplano (18–23°S). J Paleolimnol 27:45–57
Tapia PM, Fritz SC, Baker PA, Seltzer GO, Dunbar RB (2003) A Late quaternary diatom record of tropical climatic history from Lake Titicaca (Peru and Bolivia). Palaeogeogr Palaeoclimatol Palaeoecol 194:139–164
Telford RJ, Birks HJB (2009) Evaluation of transfer functions in spatially structured environments. Quat Sci Rev 28:1309–1316
ter Braak CJF (1995) Ordination. In: Jongman RHG, ter Braak CJF, van Tongeren OFR (eds) Data analysis in community and landscape ecology. University of Cambridge Press, Cambridge
Thompson LG, Davis ME, Mosley-Thompson E, Sowers TA, Henderson KA, Zagorodnov VS, Lin PN, Mikhalenko VN, Campen RK, Bolzan JF, Cole-Dai J, Francou B (1998) A 25,000-year tropical climate history from Bolivian ice cores. Science 282:1858–1864
Van Geel B (2001) Non-pollen palynomorphs. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments: terrestrial, algal, and siliceous indicators, vol 3. Kluwer, Dordrecht, pp 99–119
Wang XF, Auler AS, Edwards RL, Cheng H, Cristalli PS, Smart PL, Richards DA, Shen CC (2004) Wet periods in northeastern Brazil over the past 210 kyr linked to distant climate anomalies. Nature 432:740–743
Weckström K, Weckström J, Yliniemi L-M, Korhola A (2010) The ecology of Pediastrum (Chlorophyceae) in subarctic lakes and their potential as paleobioindicators. J Paleolimnol 43:61–73
Whitney BS, Mayle FE, Punyasena SW, Fitzpatrick KA, Burn MJ, Guillen R, Chavez E, Mann D, Pennington RT, Metcalfe SE (2011) A 45 kyr palaeoclimate record from the lowland interior of tropical South America. Palaeogeogr Palaeoclim Palaeoecol 307:177–192
Zamaloa MD, Tell G (2005) The fossil record of freshwater micro-algae Pediastrum Meyen (Chlorophyceae) in southern South America. J Paleolimnol 34:433–444
Acknowledgments
Funding for this research was provided by the Natural Science and Engineering Research Council (NSERC) (BSW) and the School of Geosciences, The University of Edinburgh (BSW). Radiocarbon analyses were granted by the Natural Environment Research Council (NERC), UK, (FEM) and we thank Dr. Charlotte Bryant for help and guidance on this topic. Fieldwork grants were obtained from the National Geographic Society (FEM) and the Royal Society (FEM). Logistical fieldwork support was provided by the ‘Noel Kempff Mercado’ Natural History Museum and the San Matías National Park authorities (especially Sr. Morales). We thank Don Chango and family for their hospitality on the shores of Laguna La Gaiba. William Gosling, Phillip Metcalfe, René Guillén, Ezequiel Chavez, Michael Burn and Huw Jones assisted with the coring and collection of samples. Thanks also to Dr. Hans Sluiman, Royal Botanic Gardens Edinburgh, for his advice on the identification of Pediastrum. We also thank two anonymous reviewers whose comments improved this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Whitney, B.S., Mayle, F.E. Pediastrum species as potential indicators of lake-level change in tropical South America. J Paleolimnol 47, 601–615 (2012). https://doi.org/10.1007/s10933-012-9583-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-012-9583-8