
Abstract
The aim of this study was to evaluate the antioxidant effect of red seaweed Pyropia columbina on juvenile Pacú (Piaractus mesopotamicus). Two hundred and ten (n = 210) fish were fed for 90 days with a control extruded feed (CEF) or the same feed added with P. columbina (35 g kg−1) (PcEF). No significant difference in final body weight, specific growth rate, condition factor, and liver somatic index was detected between dietary treatments (p > 0.05). Fish fed with PcEF showed lower lipid peroxidation (28.2 ± 2.9 vs. 39.4 ± 3.7 mmol malondialdehyde mg−1 protein) and superoxide dismutase (23.3 ± 2.3 vs. 37.7 ± 3.0 U mg−1 protein) and reduced SOD/CAT ratio (5.5 ± 0.3 vs. 8.1 ± 0.7) in intestine than those fed with CEF (p < 0.05). These results also were observed in liver and white muscle, indicating a systemic effect. No difference in glutathione S-transferase and catalase (CAT) activity in intestine, liver, and white muscle was detected between dietary treatments (p > 0.05). PcEF showed higher hemoglobin (7.5 ± 0.4 vs. 6.6 ± 0.3 g 100 mL−1) and mean corpuscular hemoglobin concentration (18.4 ± 1.8 vs. 25.7 ± 2.7%) than those fed with CEF diet (p < 0.05). Beneficial effects on lipid metabolism were observed in fish fed with PcEF respect to control diet. Pyropia columbina could be used as a natural antioxidant ingredient in fish feeding contributing a better nutritional status of cultured fish.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Pacú (Piaractus mesopotamicus, Holmberg 1887) is a highly valued fish species for aquaculture due to its acceptance by consumers and high growth rate (Machado-Neto et al. 2018). In Argentina, Pacú is the most cultivated species, achieving a milestone by overtaking the production of the exotic rainbow trout in 2012 (Valladão et al. 2018).
Intensive fish farming involves a great challenge for the producer. Handling, biometry, vaccination, and stocking density in the intensive culture system induce changes in the physiological responses interfering with health, behavior status, adaptation, and welfare of fish, promoting situations of constant stress (Machado-Neto et al. 2018). In this regard, oxidative stress arises when the organism has elevated levels of reactive oxygen species (ROS). High levels of ROS can produce lipid peroxidation of cell membranes and may damage proteins, DNA, and other biological molecules. To neutralize the harmful effects of ROS, fish have developed a complex antioxidant system that includes enzymes, minerals, and vitamins. In addition, fish may obtain antioxidant compounds they cannot synthesize through their food (Biller-Takahashi et al. 2015), such as phenolic compounds, ascorbic acid, selenium, and β-carotene.
The recognition of seaweed as a natural source of functional ingredients has grown rapidly in last years (Kumar et al. 2008; Thirunavukkarasu et al. 2013; Magnoni et al. 2017). In fact, seaweeds have been associated frequently with health benefits due to the radical scavenging and singlet O2 quenching activity present in dry, raw and cooked preparations (Sachindra et al. 2010; Kumar and Brown 2013). Antioxidant compounds in seaweed have been suggested as an endogenous defense mechanism protecting against oxidative stress due to extreme environmental conditions (Aguilera et al. 2002). Red seaweeds such as Pyropia columbina are characterized by their pigments, including phycobiliproteins, halogenated compounds, mycosporine-like amino acids, sulphated polysaccharides, and polyphenols with antioxidant activity (Yuan et al. 2005; Duan et al. 2006; Yuan and Walsh 2006; Duan et al. 2006; Yuan and Walsh 2006; Cabrita et al. 2010; Cian et al. 2013), which make this group of seaweed promising supplements in aquatic feeds (Holdt and Kraan 2011). Studies in white shrimp Litopenaeus vannamei fed with a diet supplemented with Gracilaria vermiculophylla suggested a modulatory effect on antioxidant capacity when animals were subjected to biotic and abiotic stressors (Sirirustananun et al. 2011; Chen et al. 2012). Moreover, Magnoni et al. (2017) evaluated the effects of dietary supplementation with heat-treated seaweeds (5% G. vermiculophylla or 5% Ulva lactuca) on stress bioindicators in sea bream subjected to a hypoxic challenge. They found that the hepatic antioxidant enzyme activities were differently modulated by changes in environmental O2 level, particularly in sea bream fed with the red seaweed diet, suggesting that the antioxidant properties of heat-treated seaweed may have a protective role against oxidative stress.
Currently, no research is available on in vivo antioxidant effects of extruded fish feed with red seaweed P. columbina added. Even less is known about the in vivo effect of this red seaweed on antioxidant status of temperate water fish like Pacú. Therefore, the aim of this work was to evaluate the effects of red seaweed P. columbina incorporated in extruded fish feed as antioxidant supplement using a juvenile Pacú as model.
Materials and methods
Reagents
1-chloro-2,4-dinitrobenzene (138630), 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (10102946001), 4-hydroxybenzil alcohol (H20806), 4-hydroxybenzoic acid (240141), amino acid standard solution (AAS18), caffeic acid (C0625), diethyl ethoxymethylenemalonate (D94208), 2,2-diphenyl-1-picrylhydrazyl (D9132), α-aminobutyric acid (A1879), ferulic acid (128708), gallic acid (398225), malondialdehyde (63287), p-coumaric acid (C9008), sinapic acid (D7927), thiobarbituric acid (T5500), vanillic acid (W398802), and β-carotene (22040) were obtained from Sigma Chemical Co. (USA). Other reagents were of analytical grade and obtained from Chicarelli Laboratorios (San Lorenzo, Santa Fe, Argentina).
Raw materials and production of experimental diets
One kilogram of different specimens of Pyropia columbina was handpicked in Punta Maqueda (46° 00′ S, 67° 34′ W) in the spring of 2017. Punta Maqueda is located within the San Jorge Gulf, 30 km to the south of Comodoro Rivadavia, Argentina. The seaweed was processed according to Cian et al. (2014a). Briefly, P. columbina was transported to the laboratory and stored at 4 °C inside plastic bags. Seawater, sediments, organic debris, macro fauna, and epibionts were removed by scraping and subsequent washing with distilled water. Then the samples were dried at 100 ± 4 °C and ground with a particle size lower than 1 mm, using a laboratory hammer mill (Retsch, Haan, Germany). The powder obtained was passed through a 20-mesh sieve (0.85 mm) and stored at 4 °C in plastic bags until analysis or diet formulation. Chemical composition of P. columbina was determined using Association of Official Analytical Chemist (AOAC) (2000) methods. The protein, fat, ash, total dietary fiber, and moisture content of P. columbina in dry base were 24.61 g (100 g)−1, 0.25 g (100 g)−1, 6.46 g (100 g)−1, 48.09 g (100 g)−1, and 12.79 g (100 g)−1, respectively.
Control diet (control extruded feed, CEF) was formulated with commercial corn meal, soybean meal, bovine plasma protein concentrate, corn starch, vitamin-mineral mix, and canola oil according to Cian et al. (2018a). This formulation was made taking into account the nutrient requirement for Pacú (Bicudo et al. 2009). For P. columbina diet (P. columbina extruded feed, PcEF), the formulation was the same as CEF but 35 g kg−1 of commercial corn meal were replaced by P. columbina. The level of red algae was selected according to previous results obtained in our laboratory (Cian et al. 2018b).
Chemical composition, in vitro antioxidant capacity, amino acid profile, and phenolic acid profile of extruded feeds (CEF and PcEF)
For chemical analysis, extruded feeds were ground with a cyclone sample mill (UDY Corp, USA) using a 1-mm sieve.
Chemical composition and phosphorus of CEF and PcEF were determined using AOAC (2000) methods. Total starch was quantified according to Tovar et al. (1990). Iron, zinc and calcium contents of diets were measured by atomic absorption spectroscopy after dry mineralization using an atomic absorption spectrophotometer (Analyst 300, Perkin-Elmer, USA).
Chemical composition of diets is shown in Table 1. Chemical analysis confirmed diets were isocaloric and supplied a similar amount of macronutrients. Moreover, there were not significant differences in crude protein, crude lipid, total starch, ash, calcium, phosphorous, zinc, and iron between them (p > 0.05).
For amino acid analysis, samples of CEF and PcEF were hydrolyzed with 4 mL of 6 mol L−1 HCl. The solutions were sealed in tubes under nitrogen and incubated in an oven at 110 °C for 24 h. Amino acids were determined after derivatization with diethyl ethoxymethylenemalonate by high-performance liquid chromatography (HPLC), according to the method of Alaiz et al. (1992), using D,L-α-aminobutyric acid as internal standard. The HPLC system consisted in a Shimadzu Series LC-20AT pump, with Shimadzu SPD-M20A diode array detector, equipped with a 300 × 3.9 mm i.d. reversed-phase column (Novapack C18, 4 μm; Waters). Amino acid content was expressed as g kg−1 protein. Data were processed using Shimadzu LC solution software.
Extraction of soluble phenolic compounds from feed was performed according to Qiu et al. (2010), with some modifications. Briefly, 2 g of extruded feeds were extracted twice with 15 mL of 80% methanol for 1 h at room temperature, using a mechanical shaker. Then they were centrifuged at 3500 rpm for 5 min at room temperature (Cavour VT-3216, Argentina) and the supernatants obtained were combined. Subsequently, an acid hydrolysis for the cleavage of conjugated and condensed soluble phenolic compounds in samples was performed according to Cian et al. (2012). Phenolic acids were determined by high-performance liquid chromatography (HPLC) according to Leitao et al. (2011). The HPLC system consisted of a Shimadzu Series LC-20AT pump, with Shimadzu SPD-M20A diode array detector, equipped with a 250 mm × 4.6 mm i.d. reversed-phase column (Novapack C18, 5 μm; Gemini 110A C-18 Phenomenex column). Phenolic acid content was expressed as mg kg−1 dry feed. Data were processed using Shimadzu LC solution software.
In vitro antioxidant capacity of CEF and PcEF was evaluated by DPPH inhibition, β-carotene bleaching inhibition and ABTS inhibition methods according to Cian et al. (2014b). DPPH and ABTS inhibition were expressed as mg ascorbic acid equivalent g−1 dry feed and Trolox equivalent antioxidant capacity (TEAC) g−1 dry feed, respectively.
β-carotene bleaching inhibition was expressed as percent inhibition (%).
All determinations were performed in triplicate (n = 3).
Fish and feeding trial
Juvenile Pacú were obtained from a fish farm (Pez Campero, Paraná, Argentina). The experiment was performed in the Aquaculture Laboratory at the Instituto Nacional de Limnología (CONICET, Argentina) in a recirculating water system supplied with dechlorinated city (tap) water, and equipped with an external quartz-anthracite filter (Multiválvula Vulcano Filtro VC10). The recirculation aquatic system (RAS) was set at 15.1 L min−1 with artificial aeration and 12 h light/12 h dark photoperiod regime provided by artificial illumination. The replacement of 100% of the water in each tank was made every 45 min. Physico-chemical parameters of the water remained within the values recommended by Urbinati et al. (2010) for Pacú (temperature 24.0 ± 1 °C, dissolved oxygen 6.84 ± 0.62 mg L−1, pH 6.18 ± 0.21, electrical conductivity 162.17 ± 8.30 μS cm−1, and total ammonia nitrogen 0.26 ± 0.07 mg L−1).
Prior to the feeding trial, all fish were acclimated to the indoor rearing conditions for 2 weeks. At the start of the feeding experiment, 210 juvenile Pacú (initial body weight 12.4 ± 1.9 g) were randomly stocked in six 300-L tanks with 35 fish per tank. The two test diets, CEF (control) and PcEF (P. columbina), were randomly assigned to triplicate tanks. Fish were fed twice a day (5% biomass weight per day), during 90 days at 24 °C. The daily ration was divided into two, and fed to the fish at 09:00 and 14:00 h. Leftover diet was collected to prevent nutrient leaching. The fish were weighed every 3 weeks and their ration adjusted accordingly.
The experiment was conducted in accordance with national and institutional guidelines (CONICET 2005) for the protection of animal welfare and approved by the Committee of Ethics and Safety in Experimental Work (Scientific-Technological Center, CONICET Santa Fe, Argentina).
Sample collection and morphometric indexes
At the end of the feeding trial, fish from each dietary treatment were anesthetized in benzocaine 0.1 g L−1 as described by Parma de Croux (1990). Body weight (g) and total and standard length (cm) were recorded for each individual. Growth and morphometric parameters were calculated according to Cian et al. (2017) as follows: weight gain (WG, %) = 100 × [(final body weight (g) − initial body weight (g)) × initial body weight−1 (g)] and specific growth rate (%/day) = 100 × [(ln final weight (g) − ln initial weight (g)) / days of the trial].
Blood was collected immediately from the caudal vessel (Reichenbach-Klinke 1980). Plasma was separated from whole blood by centrifugation at 1409×g for 5 min, and stored at − 80 °C. Intestine, liver, white muscle, gills, and brain were dissected, quickly frozen in liquid nitrogen, and subsequently stored at − 80 °C until lipid peroxidation and enzymes involved in antioxidant system analysis. Before freezing, the wet weight of different tissues was determined. Condition factor (CF) and liver somatic index (LSI) were calculated according to Goede and Barton (1990).
Hematological parameters
Red blood cell (RBC) counts were performed with a Neubauer chamber. Hematocrit (Ht) was determined by micro-method. Hemoglobin concentration (Hb) was measured by cyanomethemoglobin method (Houston 1990). Mean cell volume (MCV), mean cell hemoglobin (MCH), and mean cell hemoglobin concentration (MCHC) were calculated from primary indices according to Cazenave et al. (2005).
All assays were performed in triplicate.
Analytical methods
Plasma glucose, triglyceride, cholesterol, and total protein levels were determined by spectrophotometric methods (Rossi et al. 2015). Glycogen, total protein, and lipid content were quantified in the homogenate of frozen tissues (liver and white muscle) according to Rossi et al. (2017).
Tissues homogenates for the determination of lipid peroxidation of intestine, liver, white muscle, gills, and brain were prepared from each individual fish. Briefly, tissues were homogenized using phosphate buffer (pH 7.4; 30 mmol L−1). The homogenate was centrifuged at 3200 rpm (4 °C) for 30 min, and the supernatant was collected and stored at − 80 °C. Lipid peroxidation of intestine, liver, white muscle, gills, and brain were determined by measuring thiobarbituric reactive substances (TBARS), according to Yagi (1976). Protein content of each extract was determined according to Bradford (1976).
Extracts for the determination of antioxidant enzyme activities from intestine, liver, white muscle, gills, and brain were prepared from each individual (not pooled), according to Bacchetta et al. (2014). The activity of glutathione-S-transferase (GST, EC 2.5.1.18) was determined using 1-chloro-2,4-dinitrobenzene (CDNB) as substrate, according to Habig et al. (1974). Catalase (CAT, EC 1.11.1.6) and superoxide dismutase (SOD, 1.15.1.1) activities were determined according to Beutler (1982) and Misra and Fridovich (1972), respectively. The enzyme activities were calculated in terms of sample protein content. All assays were performed in triplicate (n = 3).
Mineral body content
For determination of mineral body content, whole fish were ground using a Moulinex AD5661AR meat mincer in order to obtain a homogenate. Phosphorus content of homogenates was determined following AOAC (2000). Iron, zinc, and calcium content were measured by atomic absorption spectroscopy after dry mineralization. All determinations were performed in triplicate (n = 3).
Statistical analysis
Results were expressed as means with their standard deviation (± SD). One-way analysis of variance (ANOVA) was performed, and the statistical differences among samples were determined using LSD test (least significant difference). Significance was accepted at p < 0.05. All statistical analyses were performed with Statgraphics Centurion XV 15.2.06 (Statpoint Technologies, Inc., USA).
Results
Chemical composition, amino acid profile, phenolic acid profile, and in vitro antioxidant capacity of extruded feeds
Table 1 shows the chemical composition of control extruded feed (CEF) and P. columbina extruded feed (PcEF). Significant differences for crude protein, crude lipid, total starch, ash, and mineral content were not found (p > 0.05).
Table 2 shows the amino acid profile of diets. Aspartic (Asp), glutamic (Glu), and proline (Pro) contents of PcEF were higher than those found for CEF diet. However, leucine (Leu) and phenylalanine (Phe) contents were lower than those found for control diet. For all other amino acids, no significant difference between diets was observed.
Phenolic acid profile is shown in Table 3. Both diets presented a predominance of gallic acid, followed by 4-hydroxybenzoic acid, 4-hydroxybenzil alcohol, ferulic acid, p-coumaric acid, and caffeic acid. Vanillic and sinapic acid were not detected in both diets. PcEF showed significant higher content of gallic, 4-hydroxybenzoic, and p-coumaric acid than CEF.
As shown in Table 4, PcEF showed higher in vitro antioxidant capacity than CEF. In this regard, DPPH inhibition, β-carotene bleaching inhibition, and Trolox equivalent antioxidant capacity (TEAC) of PcEF were 46, 168, and 21% higher than those obtained for CEF, respectively.
Effects of diets on morphometric indexes, hematological parameters, mineral body content, and tissue energy reserves of juvenile Pacú
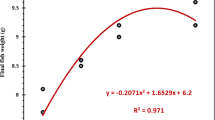
Fish promptly accepted both diets and no mortality occurred during the feeding trial. Moreover, significant differences in growth performance and condition factor were not detected between dietary treatments (p > 0.05) after 90 days of feeding trial (Table 5).
Except for hemoglobin and mean corpuscular hemoglobin concentration (MCHC), which were significantly higher for fish consuming PcEF (p < 0.05), there were no significant differences in hematological parameters between fish fed with both diets (p > 0.05). Hemoglobin and MCHC levels in fish fed with PcEF increased by 14 and 40% with respect to those fed with control diet, respectively. On the other hand, there were not significant differences in iron, zinc, calcium and phosphorus body content between diets (Table 5).
As shown in Table 6, fish fed with PcEF showed lower triglycerides and cholesterol in plasma than those consuming CEF diet. In spite of this, there was no significant difference in plasma glucose concentration. Moreover, no effect of PcEF diet on proteins, lipids and glycogen content from liver and white muscle was observed.
Effects of diets on enzymes involved in the antioxidant system and lipid peroxidation
Table 7 shows thiobarbituric reactive substances (TBARS) and the activity of glutathione S-transferase (GST), superoxide dismutase (SOD) and catalase (CAT) in intestine, liver, white muscle, gills and brain of fish fed with CEF and PcEF. Lipid peroxidation (TBARS) in intestine, liver and white muscle of fish fed with PcEF was lower than that found for tissues from fish fed with CEF. The reduction in TBARS content was more noticeable in the muscle (37%) then in the intestine (28%) and finally in the liver (17%). In spite of this, there were no significant differences in TBARS content of gills and brain between diets. The lower lipid peroxidation was accompanied by lower SOD activity. On the other hand, no effect of PcEF diet on GST and CAT activity of different tissues was observed. However, PcEF diet induced a change in SOD/CAT ratio in intestine, liver and white muscle (Fig. 1).

SOD/CAT ratio in intestine, liver, and white muscle of Pacú fed with CEF and PcEF diets. Data are expressed as mean ± SD. Different letters mean significant differences between samples analyzed by LSD test (p < 0.05)
Discussion
Composition and in vitro antioxidant capacity of extruded feeds
For most seaweed, Asp and Glu constitute a large part of the amino acid fraction, representing between 22 and 44% of the total amino acids (Fleurence 1999). Cian et al. (2014b) reported that P. columbina had 22.7 g Asp + Glu (100 g)−1 proteins, constituting the most abundant amino acid in this seaweed. Thus, the higher content of acidic amino acids of PcEF is due to the addition of P. columbina to the feed. It should be noted that PcEF diet has 63% more acidic amino acids than CEF.
Moreover, PcEF had higher content of phenolic acids than CEF (663.7 vs. 364.6 mg kg−1, respectively), gallic and 4-hydroxybenzoic acid being the most abundant. The increase of the phenolic acid content of the diet is due to the addition of P. columbina to feed. It is known, seaweeds are a very rich source of phenolic compounds such as epicatechin, gallic acid, and 4-hydroxybenzoic acid (Cian et al. 2012; Sabeena Farvin and Jacobsen 2013; Machu et al. 2015). Red seaweeds also contain particular phenolic compounds that differ in some respects from those of terrestrial plants, such as bromophenols (Liu et al. 2011). In this regard, it has been observed that 4-hydroxybenzoic acid is the most likely precursor of bromophenols (Whitfield et al. 1999), it being in high levels in P. columbina and therefore in the PcEF diet.
The higher in vitro antioxidant capacity of PcEF can be due to the antioxidant compounds provided by P. columbina. Regarding that, several studies have demonstrated the relationship between phenolics and antioxidant activity (Cian et al. 2013, 2014b). As mentioned above, PcEF had a higher content of phenolic acids which have been reported to be potent radical scavengers (Cian et al. 2013; Machu et al. 2015). In addition, red seaweeds are a rich source of iota, kappa, and lambda carrageenan, which have antioxidant activity demonstrated by in vitro and in vivo assays (Gómez-Ordóñez et al. 2012; Jiménez-Escrig et al. 2013; Cian et al. 2014a).
Finally, it has been reported that Asp and Glu are antioxidants (Cian et al. 2013) since they are hydrogen donors able to quench unpaired electrons or radicals (Quian et al. 2008). Note that PcEF had higher content of Asp + Glu than control diet.
Effects of extruded feeds on juvenile Pacú
As mentioned before, significant differences in final body weight, specific growth rate, condition factor, and liver somatic index were not detected between dietary treatments after 90 days of feeding trial. Thus, the addition of P. columbina as antioxidant supplement (35 g kg−1 diet) did not affect the weight gain and growth performance of Pacú. This result is in agreement with that reported by Peixoto et al. (2016), who evaluated the role of dietary seaweed supplementation on growth performance, digestive capacity, and immune and stress responsiveness in European seabass (Dicentrarchus labrax). They found that dietary red seaweed supplementation (25 g Gracilaria kg−1 diet) had no effects on growth rate, voluntary feed intake, feed conversion ratio, and protein efficiency ratio. A similar effect was found for rainbow trout (Oncorhynchus mykiss) fed with diets containing 50 and 100 g of Pyropia dioica kg−1 diet (Soler-Vila et al. 2009). Moreover, these authors suggested that P. dioica can be included in diets for rainbow trout up to 10% without significant negative effects on weight gain and growth performance. However, at higher levels of red seaweed replacement, negative effects on the weight gain and growth performance of the fish have been reported. In this sense, Xu et al. (2011) found that dietary seaweed supplementation above 30% reduced the growth performance and feed utilization efficiency of the teleost fish Siganus canaliculatus.
Hemoglobin and MCHC were higher in fish fed with PcEF than CEF. This means a higher iron bioaccessibility in PcEF than CEF, since the level of iron was the same in both diets (Table 1). Cian et al. (2016) studied the effect of P. columbina on iron bioaccessibility. They found that chelating peptides obtained from P. columbina protein hydrolysates had the capacity of maintaining iron in a soluble and bioaccessible form. As mentioned above, Asp and Glu were the main amino acids in PcEF. At pH 7.0, carboxyl residues of the acidic amino acids (Asp, pKa = 3.86; Glu, pKa = 4.25) are fully dissociated, being able to form complexes with iron. These complexes allow keeping the iron soluble in physiological conditions, making it more bioaccessible. Taking into account that the intestinal pH of Pacú is between 6.4 and 8.0, the higher hemoglobin and MCHC in fish fed with PcEF could be due to the higher level of these acidic amino acids in the diet, which were provided by P. columbina. On the other hand, a higher iron bioavailability of a diet has important benefits. In first place, higher bioavailability improves iron stores, which allow maintaining normal physiological state in situations of environmental stress associated with less feed ingestion. In second place, a diet with higher iron bioavailability means lower levels of fortifying for reach the fish requirement, reducing feed cost.
A lipid-lowering effect on juvenile Pacú consuming P. columbina (PcEF) was observed. In this regard, diets supplemented with 20 and 40 g kg−1Chlorella decreased serum cholesterol levels in juvenile Japanese flounder (Kim et al. 2002). Other studies performed on various animals have recorded lowering of both plasma and liver cholesterol and lipid levels through seaweed supplementation to diets (Dvir et al. 2000; Matanjun et al. 2010; Gómez-Ordóñez et al. 2012; Villanueva et al. 2014; Cian et al. 2018b). On the other hand, the main unsaturated fatty acid of P. columbina is C20:5 (n-3) (Cian et al. 2014b), which would contribute to lowering cholesterol and triglycerides in Pacú. Ragaza et al. (2015) found that juvenile Japanese flounder (Paralichthys olivaceus) fed with a diet added with red seaweed Eucheuma denticulatum (30 g kg−1 diet) presented higher n-3 polyunsaturated fatty acid accumulation in dorsal muscle and reduced serum triglycerides and total cholesterol than those fed with a control diet. Since fatty acid profile of diets were not analyzed in this study, future studies that focus on this topic are recommended. Also, the fiber provided by P. columbina (carrageenans) could modify the intestinal flora, promoting the reduction of lipids in plasma. Thus, more research is needed in order to elucidate the lipid-lowering mechanism.
Fish fed with PcEF produced antioxidative environment in intestine of juvenile Pacú, reducing lipid peroxidation (TBARS) and superoxide dismutase (SOD) activity. A similar effect was found for liver and white muscle of fish fed with PcEF. These results suggest that P. columbina exerted an antioxidant effect at the systemic level. Note that SOD belongs to the antioxidant enzymatic pathways in response to oxidative stress (Ighodaro and Akinloye 2017). In this regard, it has been suggested that polyphenols from seaweeds can reduce the in vivo formation of reactive species (Iwai 2008), modifying the enzymatic antioxidant system (Kim et al. 2008, López-Oliva et al. 2013, Figueiredo et al. 2016, Llopart et al. 2017). Zdunczyk et al. (2002) found that flavones, catechins, anthocyanins and condensed tannins from different plant sources decreased the activity of erythrocyte SOD in rats. Estruch et al. (2011) reported that moderate consumption of red wine decreases erythrocyte superoxide dismutase activity in men. This effect was attributed to high polyphenol content of red wine. Moreover, SOD activity was reduced by hydroquinone, resorcinol, and pyrocatechol (OH-phenols) in sea bass (Dicentrarchus labrax) (Roche and Bogé 2000). Taking into account that gallic, 4-hydroxybenzoic and p-coumaric acid content of PcEF was higher than that of CEF diets, the diminution of SOD activity in intestine, liver and white muscle coupled with the unmodified catalase (CAT) activity could be due to phenolic compounds from P. columbina. In agreement, SOD/CAT ratio in intestine, liver and white muscle of Pacú fed with PcEF was lower than that found for CEF, indicating an improvement at systemic level. These results suggest that PcEF modulated the activity of SOD enzyme, yielding a system that can promote a reducing environment. This was confirmed by the lower TBARS levels in intestine, liver, and white muscle of Pacú fed with PcEF. In this regard, You et al. (2014) reported that the dietary inclusion of Ulva pertusa at levels up to 10% reduced TBARS content and modified SOD and CAT activities in liver of rabbitfish Siganus canaliculatus.
Conclusions
We have demonstrated that extruded fish feed containing a low level of red seaweed P. columbina (35 g kg−1) exerted antioxidant effects on intestine, liver and white muscle in juvenile Pacú. Moreover, P. columbina promoted higher iron bioavailability observable through an increase in hemoglobin and mean corpuscular hemoglobin concentration, and lipid-lowering effect (lower plasma triglycerides and cholesterol). Pyropia columbina could be used as a natural antioxidant ingredient in fish feeding contributing a better nutritional status of cultured fish.
References
Aguilera J, Dummermuth A, Karsten U, Schriek R, Wiencke C (2002) Enzymatic defences against photo-oxidative stress induced by ultraviolet radiation in Arctic marine macroalgae. Polar Biol 25:432–441
Alaiz M, Navarro J, Giron J, Vioque E (1992) Amino acid analysis by high performance liquid chromatography after derivatization with diethylethoxymethylenemalonate. J Chromatogr 591:181–186
AOAC (2000) Official methods of analysis of AOAC International, 17th ed. Washigton DC, USA
Bacchetta C, Rossi A, Ale A, Campana M, Parma M, Cazenave J (2014) Combined toxicological effects of pesticides: a fish multi-biomarker approach. Ecol Indic 36:532–538
Beutler E (1982) Catalase. In: Beutler E (ed) Red cell metabolism, a manual of biochemical methods. Grune and Stratton Inc., New York, pp 105–106
Bicudo A, Sado R, Cyrino J (2009) Dietary lysine requirement of juvenile pacu Piaractus mesopotamicus (Holmberg, 1887). Aquaculture 297:151–156
Biller-Takahashi J, Takahashi L, Mingatto F, Urbinati E (2015) The immune system is limited by oxidative stress: dietary selenium promotes optimal antioxidative status and greatest immune defense in pacu Piaractus mesopotamicus. Fish Shellfish Immunol 47:360–367
Bradford M (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Cabrita M, Vale C, Rauter A (2010) Halogenated compounds from marine algae. Mar Drugs 8:2301–2317
Cazenave J, Wunderlin D, Hued A, Bistoni M (2005) Haematological parameters in a neotropical fish, Corydoras paleatus (Jenyns, 1842) (Pisces, Callichthyidae) captured from pristine and polluted water. Hydrobiologia 537:25–33
Chen Y, Sim S, Chiew S, Yeh S, Liou C, Chen J (2012) Dietary administration of a Gracilaria tenuistipitata extract produces protective immunity of white shrimp Litopenaeus vannamei in response to ammonia stress. Aquaculture 370-371:26–31
Cian R, Martínez-Augustin O, Drago S (2012) Bioactive properties of peptides obtained by enzymatic hydrolysis from protein byproducts of Porphyra columbina. Food Res Int 49:364–372
Cian R, Alaíz M, Vioque J, Drago S (2013) Enzyme proteolysis enhanced extraction of ACE inhibitory and antioxidant compounds (peptides and polyphenols) from Porphyra columbina residual cake. J Appl Phycol 25:1197–1206
Cian R, Salgado P, Drago S, González R, Mauri A (2014a) Development of naturally activated edible films with antioxidant properties prepared from red seaweed Porphyra columbina biopolymers. Food Chem 146:6–14
Cian R, Fajardo M, Alaiz M, Vioque J, González R, Drago S (2014b) Chemical composition, nutritional and antioxidant properties of the red edible seaweed Porphyra columbina. Int J Food Sci Nutr 65:299–305
Cian R, Garzón A, Betancur Ancona D, Chel Guerrero L, Drago S (2016) Chelating properties of peptides from red seaweed Pyropia columbina and its effect on iron bio-accessibility. Plant Foods Hum Nutr 71:96–101
Cian R E, Bacchetta C, Cazenave J, Drago SR (2017) Optimization of single screw extrusion process for producing fish feeds based on vegetable meals and evaluation of nutritional effects using a juvenile Piaractus mesopotamicus model. Anim Feed Sci Technol 234:54–64
Cian R, Bacchetta C, Cazenave J, Drago S (2018a) Extruded fish feed with high residual phytase activity and low mineral leaching increased P. mesopotamicus mineral retention. Anim Feed Sci Technol 240:78–87
Cian R, Llopart E, López-Oliva M, Weisstaub A, Zuleta A, Drago S (2018b) Low level of red seaweed Pyropia columbina added to extruded maize products promotes colonic and systemic antioxidant environment in growing Wistar rats. J Appl Phycol 30:637–648
CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas) (2005) Marco Ético de Referencia para las Investigaciones Biomédicas en Animales de laboratorio, de granja y obtenidos de la naturaleza, Buenos Aires, Argentina. https://www.web.conicet.gov.ar/documents/11716/0/OCR+RD+20050701-1047.pdf
Duan X, Zhang W, Li X, Wang B (2006) Evaluation of antioxidant property of extract and fractions obtained from a red alga, Polysiphonia urceolata. Food Chem 95:37–43
Dvir I, Chayoth R, Sod-Moriah U, Shany S, Nyska A, Stark A, Madar Z, Arad S (2000) Soluble polysaccharide and biomass of red microalga Porphyridium sp. alter intestinal morphology and reduce serum cholesterol in rats. Br J Nutr 84:469–476
Estruch R, Sacanella E, Mota F, Chiva-Blanch G, Antúnez E, Casals E, Deulofeu R, Rotilio D, Andres-Lacueva C, Lamuela-Raventos RM, de Gaetano G, Urbano-Marquez A (2011) Moderate consumption of red wine, but not gin, decreases erythrocyte superoxide dismutase activity: a randomised cross-over trial. Nutr Metab Cardiovasc Dis 21:46–53
Figueiredo F, Encarnação T, Campos M (2016) Algae as functional foods for the elderly. Food Nutr Sci 7:1122–1148
Fleurence J (1999) Seaweed proteins: biochemical, nutritional aspects and potential uses. Trends Food Sci Technol 10:25–28
Goede R, Barton B (1990) Organismic indices and autopsy-based assessment as indicators of health and condition of fish. Am Fish Soc Symp 8:93–108
Gómez-Ordóñez E, Jiménez-Escrig A, Rupérez P (2012) Effect of the red seaweed Mastocarpus stellatus intake on lipid metabolism and antioxidant status in healthy Wistar rats. Food Chem 135:806–811
Habig W, Pabst M, Jakoby W (1974) Glutathione S transferases. The first step in mercapturic acid formation. J Biol Chem 249:7130–7139
Holdt S, Kraan S (2011) Bioactive compounds in seaweed: functional food applications and legislation. J Appl Phycol 23:543–597
Houston A (1990) Blood and circulation. In: Schreck CB, Moyle PB (eds) Methods for fish biology. American Fisheries Society, Bethesda, pp 273–334
Ighodaro O, Akinloye O (2017) First line defense antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defense grid. Alex J Med. https://doi.org/10.1016/j.ajme.2017.09.001
Iwai K (2008) Antidiabetic and antioxidant effects of polyphenols in brown alga Ecklonia stolonifera in genetically diabetic KK-Ay mice. Plant Foods Hum Nutr 63:163–169
Jiménez-Escrig A, Gómez-Ordóñez E, Tenorio M, Rupérez P (2013) Antioxidant and prebiotic effects of dietary fiber co-travelers from sugar Kombu in healthy rats. J Appl Phycol 25:503–512
Kim K, Bai S, Koo J, Wang X (2002) Effects of dietary Chlorella ellipsoidea supplementation on growth, blood characteristics, and whole-body composition in juvenile Japanese flounder Paralichthys olivaceus. J World Aquacult Soc 33:425–431
Kim M, Kim J, Choi W, Lee S (2008) Effects of seaweed supplementation on blood glucose concentration, lipid profile, and antioxidant enzyme activities in patients with type 2 diabetes mellitus. Nutr Res Pract 2:62–67
Kumar S, Brown L (2013) Seaweeds as potential therapeutic interventions for the metabolic syndrome. Rev Endocr Metab Disord 14:299–308
Kumar C, Ganesan P, Suresh P, Bhaskar N (2008) Seaweeds as a source of nutritionally beneficial compounds-a review. J Food Sci Technol 45:1–13
Leitao C, Marchioni E, Bergaentzlé M, Zhao M, Didierjean L, Taidi B, Ennahar S (2011) Effects of processing steps on the phenolic content and antioxidant activity of beer. J Agric Food Chem 59:1249–1255
Liu M, Hansen P, Lin X (2011) Bromophenols in marine algae and their bioactivities. Mar Drugs 9:1273–1292
Llopart E, Cian R, López-Oliva M, Zuleta A, Weisstaub A, Drago S (2017) Colonic and systemic effects of extruded whole-grain sorghum consumption in growing Wistar rats. Br J Nutr 118:589–597
López-Oliva M, Pozuelo M, Rotger R, Muñoz-Martínez E, Goñi I (2013) Grape antioxidant dietary fibre prevents mitochondrial apoptotic pathways by enhancing Bcl-2 and Bcl-x L expression and minimising oxidative stress in rat distal colonic mucosa. Br J Nutr 109:4–16
Machado-Neto R, Wiolene M, Mariana C, Jéssica P, Débora B (2018) Growth and hematology of juvenile Piaractus mesopotamicus stocked at 10 up to 40kg/m3 for twenty-one days. Oceanogr Fish Open Access J 8:555733
Machu L, Misurcova L, Ambrozova J, Orsavova J, Mlcek J, Sochor J, Jurikova T (2015) Phenolic content and antioxidant capacity in algal food products. Molecules 20:1118–1133
Magnoni L, Martos-Sitcha J, Queiroz A, Calduch-Giner J, Gonçalves J, Rocha C, Abreu H, Schrama J, Ozorio R, Pérez-Sánchez J (2017) Dietary supplementation of heat-treated Gracilaria and Ulva seaweeds enhanced acute hypoxia tolerance in gilthead sea bream (Sparus aurata). Biol Open 6:897–908
Matanjun P, Mohamed S, Muhammad K, Mustapha N (2010) Comparison of cardiovascular protective effects of tropical seaweeds, Kappaphycus alvarezii, Caulerpa lentillifera, and Sargassum polycystum, on high-cholesterol/ high-fat diet in rats. J Med Food 13:792–800
Misra H, Fridovich (1972) The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J Biol Chem 247:3170–3175
Parma de Croux M (1990) Benzocaine (ethyl-p-aminobenzoate) as an anaesthetic for Prochilodus lineatus, Valenciennes (Pisces, Curimatidae). J Appl Ichthyol 6:189–192
Peixoto M, Salas-Leitón E, Pereira L, Queiroz A, Magalhães F, Pereira R, Abreu H, Reis P, Gonçalves J, Ozório R (2016) Role of dietary seaweed supplementation on growth performance, digestive capacity and immune and stress responsiveness in European seabass (Dicentrarchus labrax). Aquac Rep 3:189–197
Qiu Y, Liu Q, Beta T (2010) Antioxidant properties of commercial wild rice and analysis of soluble and insoluble phenolic acids. Food Chem 121:140–147
Quian Z, Jung W, Kim S (2008) Free radical scavenging activity of a novel antioxidative peptide purified from hydrolysate of bullfrog skin, Rana catesbeiana Shaw. Bioresour Technol 99:1690–1698
Ragaza J, Koshio S, Mamauag R, Ishikawa M, Yokoyama S, Villamo S (2015) Dietary supplemental effects of red seaweed Eucheuma denticulatum on growth performance, carcass composition and blood chemistry of juvenile Japanese flounder, Paralichthys olivaceus. Aquac Res 46:647–657
Reichenbach-Klinke H (1980) Enfermedades de los peces. Acribia, Zaragoza
Roche H, Bogé G (2000) In vivo effects of phenolic compounds on blood parameters of a marine fish (Dicentrarchus labrax). Comp Biochem Physiol C 125:345–353
Rossi A, Cazenave J, Bacchetta C, Campana M, Parma M (2015) Physiological and metabolic adjustments of Hoplosternum littorale (Teleostei, Callichthyidae) during starvation. Ecol Indic 56:161–170
Rossi A, Bacchetta C, Cazenave J (2017) Effect of thermal stress on metabolic and oxidative stress biomarkers of Hoplosternum littorale (Teleostei, Callichthyidae). Ecol Indic 79:361–370
Sabeena Farvin K, Jacobsen C (2013) Phenolic compounds and antioxidant activities of selected species of seaweeds from Danish coast. Food Chem 138:1670–1681
Sachindra N, Airanthi M, Hosokawa M, Miyashita K (2010) Radical scavenging and singlet oxygen quenching activity of extracts from Indian seaweeds. J Food Sci Technol 47:94–99
Sirirustananun N, Chen J, Lin Y, Yeh S, Liou C, Chen L, Sim S, Chiew S (2011) Dietary administration of a Gracilaria tenuistipitata extract enhances the immune response and resistance against Vibrio alginolyticus and white spot syndrome virus in the white shrimp Litopenaeus vannamei. Fish Shellfish Immunol 31:848–855
Soler-Vila A, Coughlan S, Guiry M (2009) The red alga Porphyra dioica as a fish-feed ingredient for rainbow trout (Oncorhynchus mykiss): effects on growth, feed efficiency, and carcass composition. J Appl Phycol 21:617–624
Thirunavukkarasu R, Pandiyan P, Balaraman D, Subaramaniyan K, Edward Gnana Jothi G, Manikkam S, Sadaiyappan B (2013) Isolation of bioactive compound from marine seaweeds against fish pathogenic bacteria Vibrio alginolyticus (VA09) and characterisation by FTIR. J Coast Life Med 1:26–33
Tovar J, Bjoerck I, Asp N (1990) Starch content and amylolysis rate in precooked legume flours. J Agric Food Chem 38:1818–1823
Urbinati E, Goncalves F, Takahashi L (2010) Pacu (Piaractus mesopotamicus). In: Baldisseroto B, Gomes LC (eds) Espécies nativas parapiscicultura no Brasil, second edn. UFSM, Santa Maria, pp 225–246
Valladão G, Gallani S, Pilarski F (2018) South American fish for continental aquaculture. Rev Aquac 10:351–369
Villanueva M, Morcillo M, Tenorio M, Mateos-Aparicio I, Andrés V, Redondo-Cuenca A (2014) Health-promoting effects in the gut and influence on lipid metabolism of Himanthalia elongata and Gigartina pistillata in hypercholesterolaemic Wistar rats. Eur Food Res Technol 238:409–416
Whitfield F, Helidoniotis F, Shaw K, Svoronos D (1999) Distribution of bromophenols in species of marine algae from eastern Australia. J Agric Food Chem 47:2367–2373
Xu S, Zhang L, Wu Q, Liu X, Wang S, You C, Li Y (2011) Evaluation of dried seaweed Gracilaria lemaneiformis as an ingredient in diets for teleost fish Siganus canaliculatus. Aquac Int 19:1007–1018
Yagi K (1976) A simple fluorometric assay for lipoperoxide in blood plasma. Biochem Med 15:212–216
You C, Zhang W, Wang S, Cheng C, Li Y (2014) Evaluation of green alga Ulva pertusa as a dietary ingredient for rabbitfish Siganus canaliculatus juveniles. J Aquac Res 1(1):005
Yuan Y, Walsh N (2006) Antioxidant and antiproliferative activities of extracts from a variety of edible seaweeds. Food Chem Toxicol 44:1144–1150
Yuan H, Zhang W, Li X, Lü X, Li N, Gao X, Song J (2005) Preparation and in vitro antioxidant activity of κ-carrageenan oligosaccharides and their oversulfated, acetylated, and phosphorylated derivates. Carbohydr Res 340:685–690
Zdunczyk Z, Frejnagel S, Wróblewska J, Juśkiewicz J, Oszmiański J, Estrella I (2002) Biological activity of polyphenol extracts from different plant sources. Food Res Int 35:183–186
Acknowledgements
All authors read and approved the final manuscript.
Funding
This work was funded by the projects PICT-2013-1804 and CAI + D 2011 PI 0292 LI from ANPCyT and Universidad Nacional del Litoral, respectively.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Rights and permissions
About this article

Cite this article
Cian, R.E., Bacchetta, C., Rossi, A. et al. Red seaweed Pyropia columbina as antioxidant supplement in feed for cultured juvenile Pacú (Piaractus mesopotamicus). J Appl Phycol 31, 1455–1465 (2019). https://doi.org/10.1007/s10811-018-1648-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-018-1648-2