
Abstract
Previous studies have used home range size to predict a species’ vulnerability to forest fragmentation. Northern bearded saki monkeys (Chiropotes satanas chiropotes) are medium-bodied frugivores with large home ranges, but sometimes they reside in forest fragments that are smaller than the species’ characteristic home range size. Here we examine how travel and spatial patterns differ among groups living in forest fragments of 3 size classes (1 ha, 10 ha, and 100 ha) versus continuous forest. We collected data in 6 research cycles from July–August 2003 and January 2005–June 2006 at the Biological Dynamics of Forest Fragments Project (BDFFP), north of Manaus, Brazil. For each cycle, we followed the monkeys at each study site from dawn until dusk for 3 consecutive days, and recorded their location. Although bearded saki monkeys living in 10-ha and 1-ha fragments had smaller day ranges and traveled shorter daily distances, they traveled greater distances than expected based on the size of the forest fragment. Monkeys in the small fragments revisited a greater percentage of feeding trees each day, traveled in more circular patterns, and used the fragments in a more uniform pattern than monkeys in the continuous forest. Our results suggest that monkeys in the small fragments maximize their use of the forest, and that the preservation of large tracts of forest is essential for species conservation. Species with large home ranges sometimes inhabit forest fragments, but doing so can alter behavior, demographics, and ecology, and the monkeys may be vulnerable to stochastic events.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
One means by which a species’ habitat is destroyed is through forest fragmentation. During this process, forested areas are cleared, leaving a mosaic of forest patches surrounded by a nonforested matrix (Gascon et al. 2001). As deforestation continues and the remaining forest becomes increasingly patchy, local climate (Achard et al. 2002; Bierregaard et al. 1992), species richness and distribution (Bierregaard et al. 1992; Laurance et al. 2000; Malcolm 1997), seed dispersal (Chapman and Onderdonk 1998; Estrada et al. 1999), and habitat suitability (Gascon et al. 2000) are affected. Often the remaining forest fragments are smaller than what is characteristically required for survival by some fauna. For example, large-bodied, frugivorous primates typically require large home ranges (Clutton-Brock and Harvey 1977; Johns and Skorupa 1987; Milton and May 1976; Onderdonk and Chapman 2000). Therefore, some researchers have stated that a highly frugivorous diet and a large home range limits, or excludes, the presence of such primate species in forest fragments (Gilbert and Setz 2001; Rylands and Keuroghlian 1988).
Forest fragmentation is one of the main threats to primate conservation (Strier 2007), yet the responses to fragmentation have varied and currently there is not a full understanding of the main factors that determine how a species will respond (Marsh 2003). In some studies, primates living in forest fragments have smaller home ranges (Cristóbal-Azkarate and Arroyo-Rodríguez 2007; Irwin 2008; Tutin 1999;Wong and Sicotte 2007), smaller group sizes (Irwin 2008), decreased biomass (Estrada et al. 1999), greater densities (Irwin 2007; Tutin 1999), different diets (Tutin 1999), and different behavioral budgets (Wong and Sicotte 2007) than primates living in continuous forest. In other studies, fragment size does not predict a primate’s activity budget and ecology (Bicca-Marques 2003; Cristóbal-Azkarate and Arroyo-Rodríguez 2007), group size does not predict home range size (Bicca-Marques 2003), group size is not associated with fragment size (Umapathy and Kumar 2003), and density is not greater in forest fragments (Wong and Sicotte 2007). Such variability in results illustrates the ecological complexities in forest fragmentation research, as variables such as fragment size (Harcourt and Doherty 2005; Wieczkowski 2004), matrix composition (Anderson et al. 2007), edge effects (Lehman et al. 2006; Mbora and Meikle 2004), predation (Tutin 1999), and seasonality of resources (Martins and Setz 2000) may affect species presence, distribution, and behavior. Further, the lack of clear patterns predicting species’ responses to forest fragmentation may be the result of these responses being site-specific (Onderdonk and Chapman 2000).
A home range is the area in which an individual engages in its regular activities (Burt 1943). There are many methods for estimating home range (Harris et al. 1990; Powell 2000; Worton 1987), as well as various applications of home range size and use measures. These applications include, but are not limited to, determining relationships between home range size and species’ characteristics (Harvey and Clutton-Brock 1981; Haskell et al. 2002; Mace and Harvey 1983), habitat preferences (Cederlund and Okarma 1988; Gese et al. 1988; Tufto et al. 1996), seasonal and annual fluctuations (Börger et al. 2006; Li et al. 2000; Wiktander et al. 2001), and sex differences (Attuquayefio et al. 1986; Fedigan et al. 1988), as well as applications to conservation and management (Bingham and Noon 1997; Bull and Holthausen 1993; Linnell et al. 2001). Further, home range size is often used to predict a species’ vulnerability to habitat loss, although the strength of such predictions have varied (Johns and Skorupa 1987; Onderdonk and Chapman 2000; Skorupa 1986).
The northern bearded saki monkey (Chiropotes satanas chiropotes) is a medium-bodied (Ford and Davis 1992), highly frugivorous seed predator (Ayres 1981; Kinzey and Norconk 1990; van Roosmalen et al. 1981) that lives in large social groups (Kinzey and Norconk 1990; Norconk et al. 2003; van Roosmalen et al. 1981) and has large home ranges (Ayres 1981; van Roosmalen et al. 1981). The taxonomy of Chiropotes is a matter of debate (Bonvicino et al. 2003; Hershkovitz 1985; Silva and Figueiredo 2002; Veiga et al. 2008). We studied the subspecies Chiropotes satanas chiropotes. The conservation status of Chiropotes chiropotes, the current Red List designation for Chiropotes satanas chiropotes, is least concern (Veiga et al. 2008). Researchers studying bearded sakis (Chiropotes spp.) have found that the species’ home range varies from 200 to 559 ha (Ayres 1981; Boyle 2008a; van Roosmalen et al. 1981) in continuous forest; however, research in Pará, Brazil (Silva 2003; Veiga 2006) and Venezuela (Peetz 2001) revealed that bearded sakis occupy areas of 16–250 ha on islands formed by the flooding of hydroelectric lakes. Further, the monkeys have inhabited forest fragments ranging from 1 ha to 100 ha at the Biological Dynamics of Forest Fragments Project (BDFFP), located in the central Brazilian Amazon (Boyle 2008a; Gilbert 2003). Therefore, bearded saki home range size appears to be somewhat flexible, particularly in areas altered by humans.
Frugivorous primates with large home ranges, i.e., Ateles spp., are often absent from forest fragments (Estrada and Coates-Estrada 1996; Rylands and Keuroghlian 1988). The presence of bearded saki monkeys in BDFFP forest fragments provided a unique opportunity to examine bearded saki groups consistently living in forest fragments that were as small as 3% of the species’ home range in continuous forest. The bearded saki monkeys were present in forest fragments of various sizes, allowing us to examine the extent to which the monkeys modify their behavior and ecology according to fragment size. The objectives of our study were to 1) compare home ranges, day ranges, and spatial patterns of bearded saki groups in forest fragments of various sizes and in continuous forest; 2) compare travel patterns, i.e., distance and movement paths among these groups; and 3) analyze seasonal fluctuations in movement patterns.
Methods
Study Site and Subjects
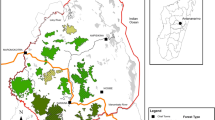
We collected focal individual movement data from July to August 2003, and then from January 2005 to April 2006, at the Biological Dynamics of Forest Fragments Project (BDFFP), located approximately 80 km north of Manaus, Brazil (2°30′S, 60°00′W). BDFFP is the site of a long-term project on fragmentation, facilitated by the Instituto Nacional de Pesquisas da Amazônia (INPA) in Manaus, Brazil, and the Smithsonian Tropical Research Institute (STRI). The forest fragments are categorized into size classes of 1, 10 and 100 ha (Fig. 1), and were initially isolated from the continuous forest by distances of 70–1000 m (Laurance et al. 2006). The forest is tropical moist terra firme, and the matrix surrounding the forest fragments ranges from pasture to tall secondary growth forest. Boyle (2008a) characterized the matrix for each of the forest fragments during the study, and Gascon and Bierregaard (2001) provided a detailed history of BDFFP and a review of the first 20 yr of research at the site.

Biological Dynamics of Forest Fragments Project (BDFFP) study site, located 80 km north of Manaus, Brazil. We surveyed 9 forest fragments throughout the Dimona, Porto Alegre, and Esteio ranches. We found bearded saki monkeys in both continuous forest sites (Cabo Frio and Km41), both 100-ha fragments (no. 2303 and no. 3304), 2 10-ha fragments (no. 2206 and no. 1202), and 1 1-ha fragment (no. 2107). Black polygons represent the forest fragments, gray areas represent the matrix, and white represents forest.
Six primate species reside in the BDFFP study area: red howlers (Alouatta macconnelli), black spider monkeys (Ateles paniscus), brown capuchins (Cebus apella), bearded saki monkeys (Chiropotes satanas chiropotes), golden-faced saki monkeys (Pithecia pithecia chrysocephala), and golden-handed tamarin monkeys (Saguinus midas). The presence of these 6 species in the BDFFP forest fragments has been variable, as some species, e.g., black spider monkey, have been virtually absent from most of the fragments (Boyle 2008b; Gilbert 2003; Gilbert and Setz 2001; Rylands and Keuroghlian 1988; Schwarzkopf and Rylands 1989).
Bearded saki monkeys were present in the BDFFP study area before the creation of the study site; however, they were absent from all of the study’s forest fragments immediately after the initial isolation process (Rylands and Keuroghlian 1988). Although the species recolonized several of the forest fragments 7–19 yr later, some of the fragments have never hosted bearded sakis (Boyle 2008a; Gilbert 2003; Gilbert and Setz 2001). Bearded saki monkeys had smaller group size and greater density (individual/ha) in the forest fragments, and the species was unlikely to cross the matrix unless there was tall secondary growth forest present (Boyle 2008a). Therefore, the monkeys were permanent residents of the isolated forest fragments during the study period, but they traveled in and out of 2 100–ha fragments that were linked to continuous forest by forested corridors.
Data Collection
We surveyed nine forest fragments—4 1-ha (nos. 1104, 2107, 2108, and 3114), 3 10-ha (nos. 1202, 2206, and 3209), and 2 100-ha fragments (nos. 2303 and #3304)—and 2 areas of continuous forest (Cabo Frio and Km41) for bearded saki monkeys. Each cycle consisted of a rotation through the 9 forest fragments and 2 continuous forest sites. It took ca. 2.5 mo to complete each cycle. We conducted 1 census cycle in July–August 2003, 4 cycles in 2005, and 1 cycle in 2006. The durations of the cycles were as follows: July 18– August 12, 2003 (cycle 2003), January 17–April 4, 2005 (cycle 1), April 13–June 22, 2005 (cycle 2), August 5–October 10, 2005 (cycle 3), October 19–December 12, 2005 (cycle 4), January 18–March 27, 2006 (cycle 5). We compared the cycles to determine seasonal differences in the monkeys’ spatial use of the forest fragments. Each study group was habituated to human presence.
On the first day in each study area for each cycle, we conducted a primate census of all six species by walking line transects along already established trails, following the methods of Rylands and Keuroghlian (1988) and Gilbert (2003). If bearded sakis were present in the study site during the primate census, we designated them the focal study group for that study site’s cycle, and subsequently relocated and followed them for 3 consecutive days. There was never >1 group of bearded sakis in a forest fragment. We identified the groups throughout the study period using group size, group composition, and several key recognizable individuals.
On locating a group of bearded sakis, we tracked the same group from the time they awoke in the morning until the time they settled down for the night (ca. 0530–1730 h). Using instantaneous and group scan sampling techniques (Altmann 1974), every 5 min we recorded the group’s geographic location via a handheld GPS receiver. If individuals were eating fruit, flowers, or leaves, we marked the feeding tree with plastic flagging and assigned a unique number. The system allowed for subsequent identification of the plant species, as well as the monitoring of revisits by monkeys to feeding trees during a cycle and throughout the year. The spatial data represent 7224 instantaneous scans (602 contact hours) of the bearded saki monkeys.
Spatial Analysis: Home and Day Range
We mapped daily travel routes and the subjects’ feeding trees via ArcView 3.3. We determined home range (ha), day range (ha), and daily distance traveled (km) via the Home Range Extension (Rodgers and Carr 2002) for ArcView. We calculated home range and day range via a minimum convex polygon (Odum and Kuenzler 1955). We did not use kernel density estimators (Worton 1987), another method for estimating home range, to calculate day range because they were inconsistent and overestimated the size of the area when sample size was low (Boyle et al. 2009). We calculated the percentage of each forest fragment used by bearded saki monkeys via a minimum convex polygon for all bearded saki data points. Because minimum convex polygon does not provide data on the proportion of time spent in various areas of the home range, we used kernel density estimators to determine the core areas used by the bearded saki monkeys. Because the calculations were based on total points for each study site, kernel density estimators performed well, due to large sample sizes. The average percentage difference between the 2 methods of home range estimation (±standard error) was 4.5% (±1.1).
We calculated home range size for 4 bearded saki groups (continuous forest sites Km41 and Cabo Frio, and 10-ha fragments nos. 1202 and 2206). We did not calculate home range for the 2 groups in the 100-ha fragments (nos. 2303 and 3304) because they frequently left and reentered the fragments using forested corridors. It was not possible to monitor their overall use of the BDFFP landscape because we did not follow them outside of the forest fragments. We also did not calculate home range for the sole individual in the 1-ha fragment because the monkey was present for only 1 cycle in 2003. We do not know whether the individual left the fragment or died.
We calculated day range (ha) and area used per hour (ha/h) for each day in each study site to evaluate the monkeys’ use of the forest. The primary purpose for calculating day range was to determine whether the monkeys in the fragments were maximizing the entire forest fragment daily. We determined an hourly average in addition to the daily area total because contact hours with the monkeys varied daily and between groups. All forest fragments (n = 5) and continuous forest sites (n = 2) used by bearded saki monkeys were included in the analyses of day range. We compared each dependent variable among forest size classes using ANOVA, with Student’s t post hoc analyses. We examined area used per hour across cycles via repeated measures ANOVA with a Greenhouse-Geisser correction for data not meeting sphericity assumptions. Because there were differences among forest size classes in overall analyses, we also included fragment size here as a between-subjects factor in repeated-measures analyses. We determined the seasonal comparisons only with data from study sites (n = 4; nos. 1202, 2206, 2303, and Km41) where bearded saki monkeys were present during all of the study cycles.
We classified the spatial distribution (clumped, random, or uniform pattern) for each bearded saki group using a nearest-neighbor analysis in the Animal Movement extension (Hooge and Eichenlaub 1997) for ArcView. We calculated an R statistic, the ratio of the actual average distance between the nearest-neighbor points and the expected average distance between points, and a z-score indicated whether the deviation from randomness was significant at α = 0.05 (Lee and Wong 2001). We followed this same procedure for the trees that were used for food by bearded saki monkeys.
Spatial Analysis: Travel Patterns
We calculated the daily distance traveled (km) and distance traveled per hour (km/h) for each day of data. We compared travel distance among forest size classes using ANOVA. We tested to see if there were differences in the distance traveled per hour across data cycles using repeated measures ANOVA with a Greenhouse-Geisser correction. Forest size class was again included as a between-subjects factor because distance traveled varied with size class in overall analyses. The seasonal analysis included only study sites (n = 4) where bearded saki monkeys were present for all of the data cycles. To compare the distance traveled and area of forest used daily, we divided distance (km) by area (ha) to provide a measurement in the amount of space used versus distance traveled. We compared forest size classes via ANOVA.
We scored daily travel patterns on a continuum from straight-line paths to circular paths based on the distance between the start and stop endpoints for the day, divided by the total distance traveled. A value of 1 indicated a straight line from start to finish, and a value of 0 indicated that the group had fully returned to its starting point. We log-transformed the data to meet the assumption of normality. We compared size classes using ANOVA. We also calculated the percentage of the feeding trees that were revisited throughout the day, and compared the results between forest class sizes using ANOVA.
Results
Presence in Forest Fragments
Northern bearded saki monkeys were present in 5 forest fragments (nos. 3304, 2303, 2206, 1202, and 2107) and 2 areas of continuous forest during the study; however, the bearded saki monkeys were continuously present in only 2 of the BDFFP forest fragments (nos. 1202 and 2206, both in the 10-ha fragment size class) during each of the study cycles in 2003, 2005, and 2006. Only 1 group was present in each 10-ha fragment, and group size was 4 for both fragments (2 males and 2 females) until October 2005, when a female disappeared from fragment no. 1202. Bearded saki monkeys were present in both 100-ha fragments (nos. 3304 and 2303), but groups left and reentered both sites frequently, and fragment no. 3304 hosted bearded sakis for only 2 of the 6 data collection cycles. There was only 1 group in each of the 100-ha fragments, and both groups were multimale and multifemale. Group size was 8–12 in no. 2303 and 13–14 in no. 3304. One individual was present in 1-ha fragment no. 2107 in 2003. Group size in the continuous forest was 22–35 at Cabo Frio and 20–33 at Km41.
We never encountered bearded saki monkeys in 4 of the 9 forest fragments. The presence of the other 5 primate species varied, and there was no indication that any primates were ever present during the study in 1-ha fragments nos. 2108 and 3114 (Table I).
Home and Day Ranges
Bearded saki home range in continuous forest was 559 ha at the Km41 study area and 300 ha at the Cabo Frio study area. The home ranges of the groups living in the 10-ha forest fragments (no. 2206: actual area 13.96 ha; and no. 1202: actual area 13.67 ha, includes 0.57 ha of tall secondary forest buffer strip) were 12.05 ha and 13.67 ha, respectively.
Bearded saki monkeys occupied a larger average daily area in the continuous forest than in the 10-ha and 1-ha forest fragments (F 3,3 = 10.61, p = 0.042; Table II). There was an even greater difference when we compared average area used per hour among forest size classes (F 3,3 = 52.20, p = 0.0043). Both of these results remained significant when we excluded data from the 1-ha fragment (average daily area: F 2,3 = 11.21, p = 0.041; average area used per hour: F 2,3 = 59.53, p = 0.0039).
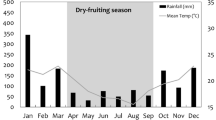
When we compared data across the 5 2005–2006 study cycles for the 4 bearded saki groups, there was a difference between cycles (F 1,1 = 356.31, p = 0.0034; Fig. 2), with area use increasing in cycle 2, the April–June 2005 time period immediately preceding the dry season. The interaction between cycle and size was marginally significant (F 2,1 = 186.06, p = 0.052), with the seasonal effect being most pronounced in the continuous forest.

Average area of forest used per hour across cycles (± standard error bars) for all bearded saki groups present for all 5 data collection cycles from 2005 to 2006.
Spatial Use of the Forest Fragments
In 2005–2006, bearded saki monkeys used 83% of the available area in the 100-ha fragment no. 2303, 54% of the 100-ha fragment no. 3304, 100% of the 10-ha fragment no. 1202, and 86% of the 10-ha fragment no. 2206. Bearded saki monkey groups occupied their forested habitat in a clumped pattern in the continuous forest sites (n = 2) and in the 100-ha fragments (n = 2), and in a uniform pattern in the 10-ha fragments (n = 2; Table III). The spatial distribution of the feeding trees was clumped in all study sites, except for 10-ha fragment no. 1202.
When we examined the areas of frequent and infrequent use by the bearded saki monkeys in the 10-ha and 100-ha forest fragments, we found that the subjects concentrated the majority of their activities away from low-lying riparian areas that contained an abundance of palms (Arecaceae), which were rarely utilized as a food resource by the monkeys. The monkeys traveled through these areas, but they did not spend a considerable amount of time there.
Travel Patterns
Bearded saki monkeys traveled greater daily distances in the continuous forest and 100-ha fragments than in the smaller fragments (all data: F 3,3 = 58.97, p = 0.0036; 1-ha data excluded: F 2,3 = 29.77, p = 0.011; Table II). When we standardized comparisons by distance traveled per hour, differences between the forest size classes were even greater (all data: F 3,3 = 155.13, p = 0.0009; 1-ha data excluded: F 2,3 = 99.70, p = 0.0018). There was no difference in distance traveled per hour between cycles (F 1,1 = 1.44, p = 0.44) or in the interaction between cycle and size (F 2,1 = 2.62, p = 0.40). The monkeys traveled greater distances per ha in the smaller fragments (all data: F 3,3 = 431.78, p = 0.0002; 1-ha data excluded: F 2,3 = 133.73, p = 0.0012).
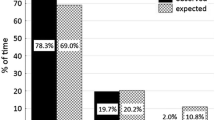
Overall, monkeys in the continuous forest traveled more in a straight-line path (value close to 1) than the monkeys in the 100-ha and 10-ha fragments, which followed more circular (value close to 0) daily routes (F 2,3 = 14.37, p = 0.029). When we considered the individual in the 1-ha fragment with the other bearded saki groups, there was still an overall difference between size classes (F 3,3 = 9.58, p = 0.048; Fig. 3a), but the 1-ha fragment values (from 2 days of data) were not statistically different from the values of the other size classes. Further, bearded saki monkeys revisited a smaller percentage of feeding trees in continuous forest and 100-ha fragments than in the smaller fragments (F 3,3 = 103.34, p = 0.0016; Fig. 3b), and this pattern held when we omitted data from the 1-ha fragment (F 2,3 = 15.72, p = 0.026).

Daily travel patterns. (a) Values close to 0 represent a circular route, and values close to 1 represent a straight-line route. (b) Percentage of feeding trees revisited each day. Standard error bars are presented in the graphs, and A, B, and C represent differences among the groups determined by Student’s t post hoc analyses.
Discussion
The purpose of our study was to compare a variety of bearded saki groups living in forest fragments of different sizes and continuous forest. As a result, sampling effort was lower for each group than it would have been had we followed only 1 group. Although sampling effort was lower per group, there were extreme spatial variations between bearded saki monkeys living in small forest fragments and monkeys living in continuous forest. Bearded sakis living in 10-ha fragments occupied smaller daily areas, traveled shorter distances daily, revisited a larger percentage of feeding trees throughout the day, followed more circular paths, and moved in a more uniform pattern throughout their habitat than bearded saki monkeys living in 100-ha fragments and continuous forest. Further, in the majority of the study areas, the monkeys did not obtain food resources from all areas of available forest. The results indicate that forest fragment size influences the spatial use of the patch by bearded saki monkeys, but other characteristics of the fragment, e.g., tree composition, resource availability, and canopy openness, may also affect a forest fragment’s potential to provide adequate resources for the species.
Both continuous-forest home range estimates from this study, 300 ha at Cabo Frio and 559 at Km41, exceeded the previously stated home ranges for bearded saki monkeys of 200–250 ha (Ayres 1981; van Roosmalen et al. 1981). However, van Roosmalen et al. (1981) stated that their home range estimate was conservative and not accurately calculated. Further, their day range estimates were based on only 5 days of data. It is likely that the bearded sakis in our study had home ranges >559 ha in the continuous forest. We had fewer contact hours with the bearded saki monkeys at Cabo Frio than at Km41. Because average daily area used and distance traveled were similar at the 2 sites, it is possible that the bearded saki monkeys at Cabo Frio had a home range that approached the size of the home range of the monkeys at Km41. There were no outlier points in the data sets that would have influenced home range estimates.
Our data did not fit the notion that primates minimize the daily distance traveled in order to locate food resources (Milton 1984) because the bearded saki monkeys in the small forest fragments traveled long daily distances per area of forest available (km/ha). The monkeys in the small forest fragments also revisited a greater percentage of feeding trees daily than the bearded sakis living in the larger forested areas. We can assume that the monkeys did not deplete their resources after each feeding bout, as they were able to return hours later and feed again, but the additive effect of these revisits could indicate a greater pressure on the resources in the forest fragments. Even though group size was smaller in the 10-ha forest fragments than in the continuous forest, bearded saki monkeys lived at greater densities in the small forest fragments than in the continuous forest (Boyle 2008a), which suggests that resource competition may have been greater in the small fragments. Our data for feeding tree revisits by the monkeys in continuous forest and 100-ha fragments were in accordance with Norconk and Kinzey (1994).
Overall bearded sakis used a greater area of forest per hour before the start of the dry season. Seasonal differences were most pronounced in the continuous forest, with little fluctuation in the 10-ha fragments. The lack of seasonal fluctuations in the 10-ha fragments may indicate that the monkeys were maximizing their resources throughout the year.
The matrix surrounding the forest fragments was heterogeneous during the study, and has varied since the 1980s (Bierregaard Jr. and Stouffer 1997; Boyle 2008a). There was no evidence that bearded saki monkeys traveled in or out of any of the forest fragments that were surrounded by a matrix of pasture and low secondary growth (Boyle 2008a). The exact date of when the monkeys arrived in the 2 10-ha fragments is not known, but it is possible that the monkeys entered these fragments when tall secondary growth was more prevalent in certain areas of the matrix. Bearded saki monkeys did, however, travel in and out of the 100-ha fragments throughout the study period, and both of these forest fragments were connected to continuous forest by forested corridors. Had the monkeys been isolated in the 100-ha fragments, it is likely that they would have used a greater percentage of the fragments.
Bearded saki group size in the forest fragments was not typical of group size in the continuous forest. This was evident even in the 100-ha fragments. Because the bearded sakis in the 100-ha fragments left and re-entered the fragments, it is possible that the monkeys were part of a larger group; however, there were never >14 individuals in a 100-ha forest fragment at one time.
One of the goals in many conservation studies is to determine the minimum fragment size needed to sustain a species (Lovejoy and Oren 1981). Although it is important to understand a species’ spatial and habitat needs, the size of the fragment is not the only consideration. In this study, bearded saki monkeys did not always utilize the entire area available to them. Sites that had many tree falls, and areas dominated by riparian or variably flooded habitats, were avoided. Only in fragment no. 1202, where there were no streams present, and the habitat was fairly homogeneous, did the monkeys use all parts of the forest fragment. Therefore, it is important to take into consideration habitat characteristics, particularly if the individuals are living in a forest fragment. Forest fragments experience greater tree mortality, mortality of large trees near forest edges, changes in tree species composition, and changes in microclimate relative to interior, continuous forest (Laurance et al. 2000, 2006). If there are stochastic events, such as increased tree falls and tree mortality due to heavy wind or storms, even less of the fragment may be available as habitat. Further, in years of low resource productivity, some small patches may not provide the minimum amount of resources necessary for the monkeys. This is especially of concern with the monkeys at BDFFP because 1) the bearded saki monkeys appeared to be uniformly maximizing their use of both 10-ha fragments; and 2) no bearded saki infants were observed in the 10-ha fragments during this study, raising the possibility that the nutritional contents of such fragments may not be sufficient for the reproduction of the inhabiting individuals (Boyle 2008a). It is also possible that bearded saki groups in the 10-ha fragments were not reproducing due to other demographic variables that were present in these small forest fragments.
Our data suggest that further decreases in the size of the forest available to bearded saki monkeys, and the accompanying changes in group size and spatial patterns, could negatively affect bearded saki populations. In continuous forest, bearded sakis live in large groups that split into smaller parties and then regroup together several times during the day (Norconk and Kinzey 1994; van Roosmalen et al. 1988), yet we never spotted bearded sakis leaving the 2 10-ha fragments, and we never noted bearded sakis entering these forest fragments. Thus, at least for the duration of our study, the monkeys in the 10-ha fragment did not appear to belong to a larger group and did not come in physical contact with other bearded saki monkeys. Further study of the bearded saki monkeys at BDFFP is critical in order to understand how the isolated groups in the 10-ha fragments fare, and whether the monkeys leave their fragment and interact with conspecifics from the continuous forest.
We conclude that 1) species with large home ranges are capable of inhabiting forest fragments that are considerably smaller that the species’ typical home range, but living in such forest fragments may alter the monkeys’ behavior, demographics, and ecology; 2) the conservation of large tracts of forest is essential for the future status of species with large home ranges; 3) the habitat type of the area should be a priority in determining future reserves; and 4) the development of tall secondary growth forest could connect populations that are isolated in forest fragments.
References
Achard, F., Eva, H. D., Stibig, H. J., Mayaux, P., Gallego, J., Richards, T., et al. (2002). Determination of deforestation rates of the world’s humid tropical forests. Science, 297, 999–1002.
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 48, 227–265.
Anderson, J., Rowcliffe, J. M., & Colinshaw, G. (2007). Does the matrix matter? A forest primate in a complex agricultural landscape. Biological Conservation, 135, 212–222.
Attuquayefio, D. K., Gorman, M. L., & Wolton, R. J. (1986). Home range sizes in the wood mouse Apodmus sylvaticus: Habitat, sex and seasonal differences. Journal of Zoology, 210, 45–53.
Ayres, J. M. (1981). Observações sobre a ecologia e o comportamento dos cuxiús (Chiropotes albinasus e Chiropotes satanas, Cebidae: Primates). M.S. thesis. Manaus, Amazonas: Insituto Nacional de Pesquisas da Amazônia and Fundação Universidade do Amazonas.
Bicca-Marques, J. C. (2003). How do howler monkeys cope with habitat fragmentation? In L. K. Marsh (Ed.), Primates in fragments: Ecology and conservation (pp. 283–303). New York: Kluwer Academic/Plenum.
Bierregaard, R. O., Jr., Lovejoy, T. E., Kapos, V., Dossantos, A. A., & Hutchings, R. W. (1992). The biological dynamics of tropical rain-forest fragments. Bioscience, 42, 859–866.
Bierregaard, R. O., Jr., & Stouffer, P. C. (1997). Understory birds and dynamic habitat mosaics in Amazonian rainforests. In W. F. Laurance & R. O. Bierregaard Jr. (Eds.), Tropical forest remnants: Ecology, management, and conservation of fragmented communities (pp. 138–155). Chicago: University of Chicago Press.
Bingham, B. B., & Noon, B. R. (1997). Mitigation of habitat “take”: Application to habitat conservation planning. Conservation Biology, 11, 127–139.
Bonvicino, C. R., Boubli, J. P., Otazú, I. B., Almeida, F. C., Nascimento, F. F., Coura, J. R., et al. (2003). Morphologic, karyotypic, and molecular evidence of a new form of Chiropotes (Primates, Pitheciinae). American Journal of Primatology, 61, 123–133.
Börger, L., Franconi, N., Ferretti, F., Meschi, F., de Michele, G., Gantz, A., et al. (2006). An integrated approach to indentify spatiotemporal and individual-level determinants of animal home range size. The American Naturalist, 168, 471–485.
Boyle, S. A. (2008a). The effects of forest fragmentation on primates in the Brazilian Amazon. Ph.D. dissertation. Tempe, AZ: Arizona State University.
Boyle, S. A. (2008b). Human impacts on primate conservation in central Amazonia. Tropical Conservation Science, 1, 6–17.
Boyle, S. A., Lourenço, W. A., da Silva, L. R., & Smith, A. T. (2009). Home range estimates vary with sample size and methods. Folia Primatologica, 80, 33–42.
Bull, E. L., & Holthausen, R. S. (1993). Habitat use and management of pileated woodpeckers in northeastern Oregon. Journal of Wildlife Management, 57, 335–345.
Burt, W. H. (1943). Territoriality and home range concepts as applied to mammals. Journal of Mammalogy, 24, 346–352.
Cederlund, G. N., & Okarma, H. (1988). Home range and habitat use of adult female moose. Journal of Wildlife Management, 52, 336–343.
Chapman, C. A., & Onderdonk, D. A. (1998). Forests without primates: Primate/plant codependency. American Journal of Primatology, 45, 127–141.
Clutton-Brock, T. H., & Harvey, P. H. (1977). Species differences in feeding and ranging behaviour in primates. In T. H. Clutton-Brock (Ed.), Primate ecology: Studies of feeding and ranging behaviour in lemurs, monkeys and apes (pp. 557–584). London: Academic Press.
Cristóbal-Azkarate, J., & Arroyo-Rodríguez, V. (2007). Diet and activity pattern of howler monkeys (Alouatta palliata) in Los Tuxtlas, Mexico: Effects of habitat fragmentation and implications for conservation. American Journal of Primatology, 69, 1–17.
Estrada, A., & Coates-Estrada, R. (1996). Tropical rain forest fragmentation and wild populations of primates at Los Tuxtlas, Mexico. International Journal of Primatology, 17, 759–783.
Estrada, A., Anzures, A., & Coates-Estrada, R. (1999). Tropical rain forest fragmentation, howler monkeys (Alouatta palliata), and dung beetles at Los Tuxtlas, Mexico. American Journal of Primatology, 48, 253–262.
Fedigan, L. M., Fedigan, L., Chapman, C., & Glander, K. E. (1988). Spider monkey home ranges: A comparison of radio telemetry and direct observation. American Journal of Primatology, 16, 19–29.
Ford, S. M., & Davis, L. C. (1992). Systematics and body size: Implications for feeding adaptation in New World monkeys. American Journal of Physical Anthropology, 88, 415–468.
Gascon, C., Williamson, G. B., & da Fonseca, G. A. B. (2000). Ecology — Receding forest edges and vanishing reserves. Science, 288, 1356–1358.
Gascon, C., & Bierregaard, R. O., Jr. (2001). The biological dynamics of forest fragments project: The study site, experimental design, and research activity. In R. O. Bierregaard Jr., C. Gascon, T. E. Lovejoy & R. Mesquita (Eds.), Lessons from Amazonia: The ecology and conservation of a fragmented forest (pp. 31–42). New Haven: Yale University Press.
Gascon, C., Bierregaard, R. O., Jr., Laurance, W. F., & Rankin-De Mérona, J. M. (2001). Deforestation and forest fragmentation in the Amazon. In R. O. Bierregaard Jr., C. Gascon, T. E. Lovejoy & R. Mesquita (Eds.), Lessons from Amazonia: The ecology and conservation of a fragmented forest (pp. 22–30). New Haven, CT: Yale University Press.
Gese, E. M., Orrin, J. R., & Mytton, W. R. (1988). Home range and habitat use of coyotes in southeastern Colorado. Journal of Wildlife Management, 52, 640–646.
Gilbert, K. A. (2003). Primates and fragmentation of the Amazon forest. In L. K. Marsh (Ed.), Primates in fragments: Ecology and conservation (pp. 145–157). New York: Kluwer Academic.
Gilbert, K. A., & Setz, E. A. (2001). Primates in a fragmented landscape: Six species in Central Amazonia. In R. O. Bierregaard Jr., C. Gascon, T. E. Lovejoy & R. Mesquita (Eds.), Lessons from Amazonia: The ecology and conservation of a fragmented forest (pp. 262–270). New Haven, CT: Yale University Press.
Harcourt, A. H., & Doherty, D. A. (2005). Species-area relationships of primates in tropical forest fragments: A global analysis. Journal of Applied Ecology, 42, 630–637.
Harris, S., Cresswell, W. J., Forde, P. G., Trewhella, W. J., Woollard, T., & Wray, S. (1990). Home-range analysis using radio-tracing data—a review of problems and techniques particularly as applied to the study of mammals. Mammal Review, 20, 97–123.
Harvey, P. H., & Clutton-Brock, T. H. (1981). Primate home-range size and metabolic needs. Behavioral Ecology and Sociobiology, 8, 151–155.
Haskell, J. P., Ritchie, M. E., & Olff, H. (2002). Fractal geometry predicts varying body size scaling relationships for mammal and bird home ranges. Nature, 418, 527–530.
Hershkovitz, P. (1985). A preliminary taxonomic review of the South American bearded saki monkeys genus Chiropotes (Cebidae, Platyrrhini), with the description of a new subspecies. Fieldiana, 27, 1–46.
Hooge, P. N., & Eichenlaub, B. (1997). Animal Movement Extension to ArcView. Anchorage, AK: Alaska Biological Science Center, U.S. Geological Survey.
Irwin, M. T. (2007). Living in forest fragments reduces group cohesion in diademed sifakas (Propithecus diadema) in eastern Madagascar by reducting food patch size. American Journal of Primatology, 69, 434–447.
Irwin, M. T. (2008). Diademed sifaka (Propithecus diadema) ranging and habitat use in continuous and fragmented forest: higher density but lower viability in fragments? Biotropica, 40, 231–240.
Johns, A. D., & Skorupa, J. P. (1987). Responses of rain-forest primates to habitat disturbance — a review. International Journal of Primatology, 8, 157–191.
Kinzey, W. G., & Norconk, M. A. (1990). Hardness as a basis of fruit choice in two sympatric primates. American Journal of Physical Anthropology, 81, 5–15.
Laurance, W. F., Delamonica, P., Laurance, S. G., Vasconcelos, H. L., & Lovejoy, T. E. (2000). Rainforest fragmentation kills big trees. Nature, 404, 836–836.
Laurance, W. F., Nascimento, H. E. M., Laurance, S. G., Andrade, A. C., Fearnside, P. M., Ribeiro, J. E. L., et al. (2006). Rain forest fragmentation and the proliferation of successional trees. Ecology, 87, 469–482.
Lee, J., & Wong, D. W. S. (2001). Statistical analysis with ArcView GIS. New York: John Wiley & Sons.
Lehman, S. M., Rajaonson, A., & Day, S. (2006). Lemur responses to edge effects in the Vohibola III Classified Forest, Madagascar. American Journal of Primatology, 68, 293–299.
Li, B., Chen, C., Ji, W., & Ren, B. (2000). Seasonal home range changes of the Sichuan snub-nosed monkey (Rhinopithecus roxellana) in the Qinling Mountains of China. Folia Primatologica, 71, 375–386.
Linnell, J. D. C., Andersen, R., Kvam, T., Andrén, H., Liberg, O., Odden, P., et al. (2001). Home range size and choice of management strategy for lynx in Scandinavia. Environmental Management, 27, 869–879.
Lovejoy, T. E., & Oren, D. C. (1981). The minimum critical size of ecosystems. In R. L. Burgess & D. M. Sharp (Eds.), Forest island dynamics in man-dominated landscapes (pp. 7–12). New York: Springer-Verlag.
Mace, G. M., & Harvey, P. H. (1983). Energetic constraints on home-range size. The American Naturalist, 121, 120–132.
Malcolm, J. R. (1997). Biomass and diversity of small mammals in Amazonian forest fragments. In W. F. Laurance & R. O. Bierregaard Jr. (Eds.), Tropical forest remnants: Ecology, management, and conservation of fragmented communities (pp. 207–221). Chicago: University of Chicago Press.
Marsh, L. K. (2003). Primates in fragments: Ecology and conservation. New York: Kluwer Academic.
Martins, M. M., & Setz, E. Z. F. (2000). Diet of buffy tufted-eared marmosets (Callithrix aurita) in a forest fragment in southeastern Brazil. International Journal of Primatology, 21, 467–476.
Mbora, D. N. M., & Meikle, D. B. (2004). Forest fragmentation and the distribution, abundance and conservation of the Tana river red colobus (Procolobus rufomitratus). Biological Conservation, 118, 67–77.
Milton, K. (1984). Habitat, diet, and activity patterns of free-ranging woolly spider monkeys (Brachyteles arachnoides E. Geoffroy 1806). International Journal of Primatology, 5, 491–514.
Milton, K., & May, M. L. (1976). Body weight, diet and home range area in primates. Nature, 259, 459–462.
Norconk, M. A., & Kinzey, W. G. (1994). Challenge of neotropical frugivory: travel patterns of spider monkeys and bearded sakis. American Journal of Primatology, 34, 171–133.
Norconk, M. A., Raghanti, M. A., Martin, S. K., Grafton, B. W., Gregory, L. T., & De Dijn, B. P. E. (2003). Primates of Brownsberg Natuurpark, Suriname, with particular attention to the pitheciins. Neotropical Primates, 11, 94–100.
Odum, E. P., & Kuenzler, E. J. (1955). Measurement of territory and home range size in birds. Auk, 72, 128–137.
Onderdonk, D. A., & Chapman, C. A. (2000). Coping with forest fragmentation: the primates of Kibale National Park, Uganda. International Journal of Primatology, 21, 587–611.
Peetz, A. (2001). Ecology and social organization of the bearded saki Chiropotes satanas chiropotes (Primates: Pithecinae) in Venezuela. Ecotropical Monographs, 1, 1–170.
Powell, R. A. (2000). Animal home ranges and territories and home range estimators. In L. Boitani & T. K. Fuller (Eds.), Research techniques in animal ecology: Controversies and consequences (pp. 65–110). New York: Columbia University Press.
Rodgers, A. R., & Carr, A. P. (2002). HRE: The home range extension for ArcView. Ontario, Canada: Centre for Northern Forest Ecosystem Research, Ontario Ministry of Natural Resources.
Rylands, A. B., & Keuroghlian, A. (1988). Primate populations in continuous forest and forest fragments in central Amazonia. Acta Amazônica, 18, 291–307.
Schwarzkopf, L., & Rylands, A. B. (1989). Primate species richness in relation to habitat structure in Amazonian rainforest fragments. Biological Conservation, 48, 1–12.
Silva, Jr., J. S., & Figueiredo, W. M. B. (2002). Revisão sistemática dos cuxiús, gênero Chiropotes Lesson, 1840 (Primates, Pithecidae) In Livro de resumos: Xth Congresso Brasileira de Primatologia: Amazônia—A última fronteira. Belém, Pará: Sociedade Brasileira de Primatologia.
Silva, S. S. B. (2003). Comportamento alimentar do cuxiú-preto (Chiropotes satanas) na área de influência do resrvatório da usina hidreléctric de Tucuruí-Pará. M.S. thesis. Belém, Pará: Museu Paraenses Emílio Goeldi and Universidade Federal do Pará.
Skorupa, J. P. (1986). Responses of rainforest primates to selective logging in Kibale Forest, Uganda: A summary report. In K. Benirschke (Ed.), Primates: The road to self-sustaining populations (pp. 57–70). New York: Springer-Verlag.
Strier, K. B. (2007). Conservation. In C. J. Campbell, A. Fuentes, K. C. MacKinnon, M. Panger & S. K. Bearder (Eds.), Primates in perspective (pp. 496–509). New York: Oxford University Press.
Tufto, J., Andersen, R., & Linnell, J. (1996). Habitat use and ecological correlates of home range size in a small cervid: the roe deer. Journal of Animal Ecology, 65, 715–724.
Tutin, C. E. G. (1999). Fragmented living: Behavioural ecology of primates in a forest fragment in the Lopé Reserve, Gabon. Primates, 40, 249–265.
Umapathy, G., & Kumar, A. (2003). Impacts of forest fragmentation on lion-tailed macaque and Nilgiri langur in Western Ghats, South India. In L. K. Marsh (Ed.), Primates in fragments: Ecology and conservation (pp. 163–189). New York: Kluwer Academic.
van Roosmalen, M. G. M., Mittermeier, R. A., & Milton, K. (1981). The bearded sakis, genus Chiropotes. In A. F. Coimbra-Filho & R. A. Mittermeier (Eds.), Ecology and behavior of neotropical primates (pp. 419–442). Rio de Janeiro: Academia Brasileira de Ciências.
van Roosmalen, M. G. M., Mittermeier, R. A., & Fleagle, J. G. (1988). Diet of the northern bearded saki (Chiropotes satanas chiropotes): A neotropical seed predator. American Journal of Primatology, 14, 11–35.
Veiga, L. M. (2006). A ecologia e o comportamento do cuxiú-preto (Chiropotes satanas) na paisagem fragmentada da Amazônia Oriental. Ph.D. dissertation. Belém, Pará: Universidade Federal do Pará.
Veiga, L. M., Silva, J., J. S., Mittermeier, R. A., & Boubli, J. P. (2008). Chiropotes chiropotes. In IUCN 2008. 2008 IUCN Red List of Threatened Species. www.iucnredlist.org. Downloaded on 21 December 2008.
Wieczkowski, J. (2004). Ecological correlates of abundance in the Tana mangabey (Cercocebus galeritus). American Journal of Primatology, 63, 125–138.
Wiktander, U., Olsson, O., & Nilsson, S. G. (2001). Seasonal variation in home-range size, and habitat area requirement of the lesser spotted woodpecker (Dendrocopos minor) in southern Sweden. Biological Conservation, 100, 387–395.
Wong, S. N. P., & Sicotte, P. (2007). Activity budget and ranging patterns of Colobus vellerosus in forest fragments in central Ghana. Folia Primatologica, 78, 245–254.
Worton, B. J. (1987). A review of models of home range for animal movement. Ecological Modelling, 38, 277–298.
Acknowledgments
We thank Brazil’s National Counsel of Technological and Scientific Development (Conselho Nacional de Desenvolvimento Científico e Tecnológico — CNPq), the Brazilian Institute of Environment and Renewable Natural Resources (Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis — IBAMA), and the National Institute for Amazonian Research (Instituto Nacional de Pesquisas da Amazônia — INPA) for support and permission to conduct this research in Brazil. The Biological Dynamics of Forest Fragments Project (BDFFP), Smithsonian Tropical Research Institute, and Arizona State University provided further logistical and financial support. This research was supported by grants and fellowships to SB from Fulbright/Institute of International Education, Margot Marsh Biodiversity Foundation, Providing Educational Opportunities (PEO), Primate Conservation, Inc., Organization for Tropical Studies, American Society of Primatologists, and IDEAWILD. We thank Alaercio Marajó dos Reis and Osmaildo Ferreira da Silva for valuable field assistance. Leanne Nash, David Kabelik, John Alcock, and 2 anonymous reviewers provided helpful feedback to improve the manuscript. This is publication number 529 in the BDFFP technical series.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Boyle, S.A., Lourenço, W.C., da Silva, L.R. et al. Travel and Spatial Patterns Change When Chiropotes satanas chiropotes Inhabit Forest Fragments. Int J Primatol 30, 515–531 (2009). https://doi.org/10.1007/s10764-009-9357-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-009-9357-y