
Abstract
Understanding spatial and temporal changes in species composition has long demanded the attention of ecologists. However, only recently questions related to changes in species functional traits have been investigated. We explored patterns of species and functional dissimilarity of periphytic algae at six lakes sampled over a year across a subtropical floodplain. We evaluated the importance of turnover and nestedness components across space and time; the influence of environmental dissimilarity, spatial distance, and time on species and functional dissimilarity; and whether functional dissimilarity results from non-stochastic assembly processes. We used six functional traits to describe 155 species. Functional dissimilarity was estimated by functional dendrograms, and stochasticity was evaluated using null models. The turnover component was greater than nestedness for species and functional dissimilarity. Environmental dissimilarity, spatial distance, or time did not significantly explain species or functional dissimilarity. However, functional dissimilarity was significantly greater than expected given the observed species dissimilarity. The main finding of this study is that community assembly was deterministic with respect to traits. Further, each lake contributed similarly to the overall species and traits pool. These results highlight the importance of comparing species and functional dissimilarities to reach a better understanding of the organization of periphytic algal communities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A central question in Ecology is to understand the patterns and processes related to the organization and establishment of communities as well as their implications for the maintenance of biodiversity and ecosystem processes (McGill et al., 2006). Traditionally, studies on biodiversity focused on the patterns and mechanisms that explain species richness (e.g. Rosenzweig, 1995). Nonetheless, two separated communities in space or time may present similar species richness but show distinct species composition (beta diversity, Legendre et al., 2005; Melo et al., 2011). Dissimilarities in species composition may be related to environmental dissimilarity between habitats such that dissimilar communities may occur in environmentally dissimilar habitats (Weiher et al., 2011). Community similarity may also decline with increasing geographic distance among sites (‘distance decay’; Soininen et al., 2007) due to decreasing similarity in environmental features and to dispersion limitation of the organisms (Soininen et al., 2007). Finally, community similarity tends to decrease over time due to temporal environmental variability that affects local colonization and extinction processes and to temporal dynamics related to the dispersion of organisms across sites (Korhonen et al., 2010). Studies that focused on periphytic algal species showed that environmental dissimilarity (Schneck et al., 2011a) and spatial distance (Wetzel et al., 2012) may be important in structuring algal communities.
Further, there is strong evidence of the importance of species functional traits to determine the organization of communities (Petchey & Gaston, 2006; Mason & De Bello, 2013). For instance, studies that analyzed different aspects related to algal biological traits showed that algal life form, adherence, and locomotion are key traits that determine algae response to light (Lange et al., 2011), nutrients (Passy, 2007; Piggott et al., 2012), sediment deposition (Piggott et al., 2012), disturbance (Passy, 2007; Schneck & Melo, 2012), substrate roughness (Schneck et al., 2011b), and dispersal patterns (Wetzel et al., 2012). In this sense, it is important to understand not only the patterns regarding the dissimilarity in species composition but also the patterns associated with functional dissimilarity. Functional dissimilarity among communities assesses the changes in species functional traits in space or time and thus takes into account the similarity in ecological and evolutionary characteristics of species (Swenson, 2011; Swenson et al., 2012). Considering species and functional dissimilarity, it could be expected that high species dissimilarity could result in either high functional dissimilarity (low functional redundancy) or low functional dissimilarity (high functional redundancy) (Swenson et al., 2012).
Dissimilarities in species composition between communities may result from two distinct patterns: turnover and nestedness (Baselga, 2010; Baselga & Orme, 2012). Turnover refers to the replacement of species in one community by different species in the other community, that is, two communities with similar species richness share just a few species due to environmental or historical spatial restrictions or barriers (Baselga, 2010; Villéger et al., 2013). In turn, the nestedness-resultant dissimilarity reflects the increasing dissimilarity between nested communities produced by the increasing differences in the number of species. A nested pattern may result from different factors that lead to species loss or gain by selective extinction or colonization, such as habitat quality, area and isolation, or species tolerance to abiotic factors (Patterson & Atmar, 1986; Wright et al., 1998). This approach was extended to functional dissimilarity, enabling its partition in functional turnover and nestedness-resultant components (Melo, 2013; Villéger et al., 2013).
Revealing the patterns related to species and functional dissimilarity is an important step to understand how communities are assembled. However, as stated by Villéger et al. (2013), we also need to investigate whether functional dissimilarity differs from expected given the levels of species dissimilarity. Dissimilarity may be non-random regarding the functional characteristics of the species, since deterministic processes, such as limiting similarity and environmental filters, would produce communities that are functionally more divergent or more similar (i.e., greater or lower functional dissimilarity) than expected given species dissimilarity. Alternatively, stochastic processes are expected to produce random variation in the functional strategies of the species given the species pool (Swenson et al., 2011; Swenson et al., 2012).
Patterns associated with species and functional dissimilarity are poorly explored for periphytic algae in general, and particularly in floodplain ecosystems. In this context, we assessed the patterns of species and functional dissimilarity and their respective turnover and nestedness-resultant components of periphytic algal communities in subtropical floodplain lakes. We tested whether (i) dissimilarity is mostly related to the turnover or nestedness-resultant components at spatial and temporal scales; (ii) functional dissimilarity is positively related to species dissimilarity; (iii) species and functional dissimilarity are positively related to environmental dissimilarity, geographic distance, or time; (iv) functional dissimilarity is not a stochastic process, differing from random expectations given the levels of species dissimilarity.
Materials and methods
Study area
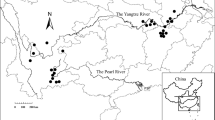
We used data from six lakes belonging to the Upper Paraná River floodplain (22º54′ 30.3″S; 53º38′ 24.3″W; 22º44′50.76″S; 53º15′11.16″W) (Fig. 1). The region has a hydrological regime characterized by a wet period (October to February) and a dry period (June to September). In flooded periods (high water levels), the average of Paraná River hydrometric level is around 4.5 m, flooding the entire floodplain area and thus increasing the connectivity between the environments (Souza-Filho & Stevaux, 2004). More recently, dam regulation changed the frequency, duration, and amplitude of the floods (Agostinho et al., 2004). All the six lakes are connected to the Ivinhema River and during the high water phase to the Paraná River (Souza-Filho & Stevaux, 2004). The greatest distance between lakes is roughly 20 km. All lakes were sampled four times in 2011 (S1-March, S2-June, S3-September and, S4-December). Samplings S1 and S4 were conducted in months that are characteristically considered as high-water period, despite the low water level observed at S4 (Fig. 2); while S2 and S3 samplings occurred during the low-water period (Fig. 2).

Location of the Upper Parana River floodplain, Brazil, and the six sampled lakes: 1 Peroba, 2 Capivara, 3 Patos, 4 Jacaré, 5 Cervo, 6 Sumida. Indicative arrow shows direction of flow

Hydrometric level of the Paraná River in 2011. Arrows indicate the months of sampling; dashed line indicates the level of flooding the plain
Sampling
The sampling of environmental variables was carried out in the subsurface of the limnetic zone of each lake concurrently with the sampling of biotic variables. The following variables were measured in the field: Secchi depth (m), conductivity (µS cm−1) and pH (Digimed digital potentiometers), turbidity (NTU), total solids (mg l−1), and total alkalinity (mEq l−1). Water samples were filtered through Whatman GF/F filters, under low pressure (<0.5 atm) and stored at −20°C for later determination of orthophosphate (µg l−1), total phosphorus (µg l−1), and total nitrogen (µg l−1, Bergamin et al., 1978). Analyses of environment variables were done following Roberto et al. (2009).
Periphytic algae were collected from the sixth or seventh plant internode of three submerged mature petioles of Eichhornia azurea (Sw.) Kunth, totaling three sub-samples per lake. The plants were sampled on the marginal region of different macrophyte patches located in the littoral zone of the lakes. E. azurea is the most abundant macrophyte in this floodplain and is present in all the studied lakes (Bini et al., 2001). The periphytic material was taken by scraping part of petioles of E. azurea, using a stainless steel blade wrapped in aluminum foil and jets of distilled water (Algarte et al., 2014). The area scraped from the substrate (cm2) was calculated from measurements of the length and width of each petiole. The material removed was preserved with 0.5% acetic acid Lugol’s solution for later counting.
The periphytic algal samples were quantified using sedimentation chambers in an inverted microscope (Utermöhl, 1958) and sedimentation time followed Lund et al. (1958). The counts were carried out in random fields until reaching at least 100 individuals (cells, colonies, or filaments) from the most common species in each sub-sample and according to the species accumulation curve (Ferragut & Bicudo, 2012). We counted an average of 813 individuals per sub-sample (minimum = 241, maximum = 3288, standard deviation = 674). The classification system adopted was that proposed by Round (1971).
Periphytic algal functional niche was characterized for main facets as growth, reproduction, life form, locomotion, substrate adherence, and disturbance resistance (Biggs et al., 1998), using traits commonly included in studies on periphytic algae that use a trait-based approach (e.g. Ferragut & Bicudo, 2010; Passy & Larson, 2011; Schneck & Melo, 2012; Dunck et al., 2013; Hart et al., 2013; Algarte et al., 2014). The selection of functional traits was conducted considering the traits that represent the ecological niche or that provide the most satisfactory establishment in the habitats. The species functional matrix was composed of six algal functional traits distributed in 17 categories and one quantitative trait: life form (non-motile unicelular, filamentous, flagellate, or colonial), adherence form (motile, entangled, prostrate, stalked, or heterotrichous-differentiated basal cell), reproduction (predominantly episodic sporulation, predominantly filament fragmentation, or predominantly mitosis), disturbance resistance (high, medium, or low) and nitrogen fixation (yes or not) according to Biggs et al. (1998), and size (quantitative, mean length of at least five individuals, in µm). Life form, adherence form, and size were directly analyzed on the individuals of each species; the other functional traits were analyzed using information from species identification specialized bibliography (e.g., Komárek & Anagnostidis, 1986; Komárek & Anagnostidis, 1989; Biggs et al., 1998; Komárek & Anagnostidis, 1998; John et al., 2002; Wehr & Sheath, 2003). We are aware that functional traits such as reproduction and nitrogen fixation are plastic or facultative in nature. However, the use of specialized bibliography to assign these functional traits is the best way to proceed, since it is very difficult, if not impossible, to evaluate such traits for each individual while counting is been conducted.
A hundred and fifty-five species of periphytic algae were found in the six studied lakes (electronic supplementary material—ESM1). Bacillariophyceae, Zygnemaphyceae, and Chlorophyceae were the most species-rich classes of algae at all sampling periods. Bacillariophyceae was the dominant group, accounting for 34% of total density, followed by Zygnemaphyceae with 22%. Regarding species traits, non-motile unicellular life form was the dominant one (91 species), followed by the filamentous life form (41 species). Entangled (78 species) and prostrate (36 species) were the dominant adherence forms. Most species reproduce by mitosis (86 species); the disturbance resistance of species was predominantly low (102 species), and there was predominance of non-nitrogen-fixing species (148 species) (electronic supplementary material—ESM1).
Data analysis
A principal component analysis (PCA) was applied to summarize environmental variables (Legendre & Legendre, 1998) with variables previously log-transformed (except pH) and standardized. Axis retention was evaluated under the Broken-Stick criterion (Jackson, 1993).
We constructed a presence–absence matrix derived from a quantitative matrix that contained 155 species and 24 samples (six lakes × four sampling periods). We used the Sørensen index (βsor; Legendre & Legendre, 1998) to calculate pairwise dissimilarity in species composition between all samples (276 dissimilarity values). The distance matrix among species functional traits was calculated using a modification of Gower distance, the coefficient of distance for mixed variables proposed by Pavoine et al. (2009). This distance matrix was transformed into a dendrogram using the method of average linkage clustering (UPGMA). This dendrogram was used along with the species presence–absence matrix to calculate pairwise functional dissimilarity between samples through the Sørensen index (βsor) adapted to functional traits (Melo, 2013).
Pairwise environmental dissimilarity was calculated using Euclidean distance after centering each variable by its mean and scaling each variable by its standard deviation (Legendre & Legendre, 1998). The spatial distance between all pairs of samples was obtained by calculating the Euclidean distance using a matrix of geographical coordinates. A Euclidean distance matrix between sampling periods (i.e., number of months apart) was generated to take into account the temporal variation in the dataset.
The partition of βsor (total dissimilarity, calculated as specified above) in two components that account for the turnover-resultant dissimilarity (βsim) and for the dissimilarity due to differences in richness (nestedness-resultant component, βnes) was carried out according to Baselga & Orme (2012) for species dissimilarity and according to Melo (2013) for functional dissimilarity. In this method, the turnover component is obtained by calculating the Simpson dissimilarity index, while the nestedness-resultant component is the difference between βsor and βsim. We conducted separated partitions to analyze patterns for either spatial or temporal scales. To decompose dissimilarities in space, we calculated βsim and βnes for all pairs of lakes within the same sampling period, totaling 60 βsim and βnes values (15 pairwise comparisons between lakes × four sampling periods). We also calculated βsim and βnes at the temporal scale for all pairs of sampling periods within the same lake, totaling 36 βsim and βnes values (six pairwise comparisons between sampling periods × six lakes). Two-tailed paired t tests were used to test for differences between the turnover and the nestedness-resultant components separately at the spatial and temporal scales for each type of dissimilarity (species and functional). Size effects of t tests were calculated using Cohen’s d measure for paired samples t test (Cohen, 1988).
The relationship between species and functional dissimilarity was accessed through a Mantel test (1000 permutations) on the Sørensen dissimilarity matrices. We used multiple regressions on distance matrices (MRM; Legendre et al., 1994; see also Lichstein, 2007) to evaluate whether total dissimilarity (βsor) in species or functional composition is explained by environmental dissimilarity, geographic distance, and time. The significances of regression coefficients and coefficients of determination were evaluated with 10,000 permutations. Before MRMs, we selected the environmental dissimilarity matrix with maximum Spearman’s rank correlation with each biotic dissimilarity matrix among all possible combinations of subsets of environmental variables (BioEnv approach; Clarke & Ainsworth, 1993). The selected variables were total nitrogen and orthophosphate for both species (r = 0.26) and functional (r = 0.19) dissimilarity.
To evaluate whether functional dissimilarity is greater or less than expected given the species dissimilarity, we generated a null model in which species labels were randomized across tips of the dendrogram. We randomly selected 1000 iterations from the trait dendrogram and, for each iteration, we calculated the Sørensen index (as described above) to create a null distribution for the expected functional dissimilarity. This null model maintains intact the sample-by-species matrix (i.e., community composition, species richness, and species dissimilarity are maintained) as it only randomizes the functional similarity among species (a similar null model was used by Swenson et al., 2011). We calculated the observed functional dissimilarity for each of the 276 pairs of samples (using a trait dendrogram and the Sørensen index, as described above) and the equivalent average of 1000 values of functional dissimilarity generated by the null model to obtain one set of 276 pairs of functional dissimilarity (the observed value and its respective expected value under the null model). Then, we conducted a two-tailed paired t test to test whether observed functional dissimilarity was greater or less than expected given species dissimilarity. Here, instead of comparing each observed value to the distribution of the simulated functional dissimilarity separately for all the 276 pairwise comparisons, we opted to conduct a paired t test so that the results would highlight the pattern for the complete dataset and not for each pairwise comparison separately. A similar approach was used by Schneck et al. (2011b) and by Both & Melo (2015) for other randomization tests.
Results were similar between the full dataset and after excluding rare species (less than 1% abundance in one sample), and thus we are showing only results from analyses including the full dataset. All analyses were carried out using R platform (R Core Team, 2014). Packages ade4 (Chessel et al., 2004) and picante (Kembel et al., 2010) were used to construct the distance functional matrix and dendrogram, betapart (Baselga et al., 2013) and CommEcol (Melo, 2013) to partition species and functional dissimilarity, respectively, vegan (Oksanen et al., 2013) for Mantel test and BioEnv, ecodist (Goslee & Urban, 2007) for MRM, and stats for t tests.
Results
The first axis explained 36% of the total variation in the limnological data and had pH (r = −0.69), turbidity (r = −0.53) and total phosphorus (r = −0.33) as the most-related environmental variables. The second axis explained 23% of the variation and was positively correlated with pH (r = 0.62) and negatively correlated with turbidity (r = −0.47), orthophosphate (r = −0.35), and total phosphorus (r = −0.34). In general, the lakes were ordinated according to the sampling period (Fig. 3). In S1 and S4, the lakes were characterized by higher values of nutrients, turbidity, and conductivity than in S2 and S3 (Fig. 3). In S2, all lakes had higher values of pH and total solids, while at S3 the lakes had more transparent waters (Fig. 3).

Site scores derived from a principal component analysis applied to the environmental variables. Arrows indicate the Pearson correlations between original variables and ordination scores. (Lakes L1 Peroba, L2 Capivara, L3 Patos, L4 Jacaré, L5 Cervo, L6 Sumida, S1 March, S2 June, S3 September and, S4 December, pH, CON conductivity, SEC Secchi depth, TUR turbidity, MST total solids, ALK total alkalinity, NT total nitrogen, PO4 orthophosphate, PT total phosphorus)
At the spatial scale, mean total species dissimilarity (βsor) was 0.54 ± 0.14 and mean total functional dissimilarity was 0.26 ± 0.07. At the temporal scale, mean total species dissimilarity was 0.47 ± 0.13 and mean total functional dissimilarity was 0.23 ± 0.09. The turnover component was significantly greater than the nestedness-resultant component at the spatial scale for both species (t = 16.90, P < 0.001, size effect = 2.18) and functional dissimilarity (t = 5.77, P < 0.001, size effect = 0.74) (Fig. 4) and at the temporal scale also for both species (t = 12.87, P < 0.001, size effect = 2.15) and functional dissimilarity (t = 2.74, P = 0.009, size effect = 0.46), being responsible for 83.2, 66.4, 82.1, and 61.7% of the mean total dissimilarity, respectively (Fig. 4).

Box-and-whisker plots of species dissimilarity (1) and functional dissimilarity (2) of periphytic algal communities partitioned into its turnover and nestedness-resultant components at the spatial scale (A) and the temporal scale (B) at four sampling periods. The heavy line shows the median, box ends show quartiles, whiskers show 95% confidence interval, and open circles show outliers
Species and functional dissimilarities showed a positive but weak correlation (Mantel test, r = 0.627, P = 0.001). However, both species dissimilarity and functional dissimilarity were not significantly explained (R 2 = 0.07, P = 0.184; R 2 = 0.02, P = 0.649, respectively) by either environmental dissimilarity (standardized partial regression coefficient b = 0.23, b = 0.13, respectively), spatial distance (standardized partial regression coefficient b = −0.09, b = 0.03, respectively), or time (standardized partial regression coefficient b = 0.03, b = 0.01, respectively).
Functional dissimilarity differed from random expectations given the observed species dissimilarity (t = 12.55, P < 0.001). Functional dissimilarity (mean = 0.26 ± 0.07) was greater than expected by the null model (mean = 0.22 ± 0.05), with a mean of the differences of 0.04 (Fig. 5).

Box-and-whisker plots of the null model results for observed and expected functional dissimilarity, given the observed species dissimilarity of periphytic algal communities. The heavy line shows the median, box ends show quartiles, whiskers show 95% confidence interval, and open circles show outliers
Discussion
Our results showed similar patterns in species and functional dissimilarities. First, we found a greater contribution of the turnover component than of the nestedness-resultant component for both species and functional dissimilarities at space and time. This indicates that each lake contributes similarly to the overall species and traits pool, since the biotic dissimilarity between lakes and between sampling periods is mainly caused by species and traits substitution and not by differences in richness (e.g., Villéger et al., 2013). Moreover, the positive correlation between species and functional dissimilarities indicates a low functional redundancy among species from the regional pool. Some studies on lotic ecosystems have already shown that the covariation between trait-based and taxonomic patterns may have similar geographical variation (Heino et al., 2007; Heino et al., 2013), depending of the spatial extent of the study area, environmental conditions, and species distribution (Heino et al., 2013). Here, we lack evidence of which factors are responsible for this positive correlation since our results did not show any significant relationship between species/functional dissimilarity and environmental dissimilarity, spatial distance, or time.
The lack of evidence of environmental, spatial, or temporal effects on periphytic algal communities for both species and functional dissimilarity was not an expected result (but see Nabout et al., 2009 for a similar result). Among all results, the lack of a spatial effect is the most expected to occur, since microorganisms have high dispersal abilities and thus a low rate of distance decay in similarity is expected (Nekola & White, 1999; but see Wetzel et al., 2012). On the other hand, the relevance of environmental heterogeneity in shaping the structure of communities of microorganisms has been shown for various aquatic environments (De Bie et al., 2012; Algarte et al., 2014; Padial et al., 2014; Dunck et al., 2015). Environmental filters may restrict the occurrence of species to the environmental conditions of the habitats, and higher variety of environmental conditions provides more variety of niches, leading to variation in species composition among localities (Chase & Leibold, 2003; Leibold et al., 2004).
Our results could be explained by the small spatial extent of the study (the two most distant lakes are ~20 km away) and by the high connectivity among environments in the floodplain such that the lakes are environmentally homogeneous and the landscape has no limitation to the dispersal of algae. However, temporal differences in water level and limnological characteristics of the lakes occurred within the 9 months of this study (as highlighted by the PCA in Fig. 3), but such temporal variations had no effects on the biotic dissimilarity. This result contradicts the expectation that temporal variation strongly affects the structure of communities at floodplain ecosystems trough seasonal flood pulses (Thomaz et al., 2007). Thus, it could be suggested that environmental selective factors, such as seasonal changes on water level and on limnological characteristics, may not be important for understanding dissimilarities in algal communities in the Upper Paraná River floodplain, at least in the spatial scale of this study and under the regulated water discharge regime imposed by hydroelectric dams. In this floodplain, floods present some stochasticity because of regulation by dams, which change the frequency, duration, and amplitude of the floods, the connectivity between the main river and the marginal lakes, and the depths of these environments (Agostinho et al., 2004; Thomaz et al., 2007). The water regulation by reservoirs causes large daily fluctuations on the discharge and water level (up to 1.0 m) of Paraná River (Souza-Filho & Stevaux, 2004). Such unpredictable disturbances by floods and water level fluctuations may be important factors driving local colonization and extinction events in this floodplain, being thus the driving forces behind the diversity patterns observed in this study. Evidence from other studies suggests that the pervasive effects of floods may obscure the effects of other environmental factors. For instance, it has been shown that the disturbance caused by a managed flood regime at a Swiss floodplain river was not uniform among floods, causing high variation in biomass patterns among years (Uehlinger et al., 2003). Moreover, recent studies on the Upper Paraná River floodplain also suggested that the regulated water discharge regime may override the expected effect of natural flood pulse regimes on different biological communities (e.g., Padial et al., 2014).
Important evidence that the assembly of the studied periphytic algal communities is regulated by deterministic processes is that functional dissimilarity was not random given species dissimilarity. According to Swenson et al. (2011), if the replacement of species between two communities is functionally not random such that the species in the communities are functionally more divergent than expected, dissimilarity could not be explained solely by dispersal limitation but by environmental gradients that strongly influence the replacement of species functional strategies and the assembly of communities. Thus, the fact that periphytic algal communities are functionally more dissimilar in space and time than expected given the observed species dissimilarity indicates that deterministic processes related to environmental gradients are shaping the functional organization of these algal communities. However, the environmental variables included in our analyses were not able to highlight the underlying environmental gradient, probably caused by stochastic events of the flood pulse, indicating that future studies should try to control effects of stochastic events in an attempt to elucidate whether such approach could be able to enhance the explanation of the variation in the community composition of periphytic algae from floodplain lakes.
In summary, our main finding is that functional dissimilarity of periphytic algal communities differs from null expectations, that is, community assembly was deterministic with respect to traits. This result emphasizes the importance of evaluating both species and functional dissimilarities. Moreover, we found that each lake contributes similarly to the overall species and traits pool (predominance of the turnover component over the nestedness-resultant component), such that conservation efforts aiming just a few lakes may not be effective in protecting all the regional algal species pool. These results highlight the importance of studies comparing species and functional dissimilarities at both spatial and temporal scales to reach a better understanding of the organization of periphytic algal communities.
References
Agostinho, A. A., L. C. Gomes, S. Veríssimo & E. K. Okada, 2004. Flood regime, dam regulation and fish in the Upper Parana River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish Biology and Fisheries 14: 11–19.
Algarte, V. M., L. Rodrigues, V. L. Landeiro, T. Siqueira & L. M. Bini, 2014. Variance partitioning of deconstructed periphyton communities: does the use of biological traits matter? Hydrobiologia 722: 279–290.
Baselga, A., 2010. Partitioning the turnover and nestedness components of beta diversity. Global Ecology and Biogeography 19: 134–143.
Baselga, A. & D. Orme, 2012. Betapart: an R package for the study of beta diversity. Methods in Ecology and Evolution 3: 808–812.
Baselga, A., D. Orme & S. Villeger, 2013. Betapart: partitioning beta diversity into turnover and nestedness components. R package version 1.2. http://cran.r-project.org/package=betapart.
Bergamin, H., B. F. Reis & E. A. G. Zagato, 1978. A new device for improving sensitivity and stabilization in flow injection analysis. Analytica Chimica Acta 97: 427–431.
Biggs, J. F., R. J. Stevenson & R. L. Lowe, 1998. A habitat matrix conceptual model for stream periphyton. Archiv für Hydrobiologie 143: 21–56.
Bini, L. M., S. M. Thomaz & D. C. Souza, 2001. Species richness and b-diversity of aquatic macrophytes in the Upper Paraná River floodplain. Archiv für Hydrobiologie 151: 511–525.
Both, C. & A. S. Melo, 2015. Diversity of anuran communities facing bullfrog invasion in Atlantic Forest ponds. Biological Invasions 17: 1137–1147.
Chase, J. M. & M. A. Leibold, 2003. Ecological Niches. Linking Classical and Contemporary Approaches. University of Chicago Press, Chicago Press.
Chessel, D., A. B. Dufour & J. Thioulouse, 2004. The ade4 package-I- One-table methods. R News 4: 5–10. http://www.r-project.org/doc/Rnews/bib/Rnewsbib.html.
Clarke, K. R. & M. Ainsworth, 1993. A method of linking multivariate community structure to environmental variables. Marine Ecology-Progress Series 92: 205–219.
Cohen, J., 1988. Statistical power analysis for the behavioral sciences. Lawrence Erlbaum Associates, Hillsdale.
De Bie, T., L. De Meester, L. Brendonck, K. Martens, B. Goddeeris, D. Ercken, H. Hampel, L. Denys, L. Vanhecke, K. Van der Gucht, J. Van Wichelen, W. Vyverman & S. A. J. Declerck, 2012. Body size and dispersal mode as key traits determining metacommunity structure of aquatic organisms. Ecology Letters 15: 740–747.
Dunck, B., J. C. Bortolini, L. C. Rodrigues, S. Jati, S. Train & L. Rodrigues, 2013. Flood pulse drives functional diversity and adaptative strategies of planktonic and periphytic algae in isolated tropical floodplain lake (Brazil). Brazilian Journal of Botany 36: 257–266.
Dunck, B., E. Lima-Fernandes, F. Cássio, A. Cunha, L. Rodrigues & C. Pascoal, 2015. Responses of primary production, leaf litter decomposition and associated communities to stream eutrophication. Environmental Pollution 202: 32–40.
Ferragut, C. & D. C. Bicudo, 2010. Periphytic algal community adaptive strategies in N and P enriched experiments in a tropical oligotrophic reservoir. Hydrobiologia 646: 295–309.
Ferragut, C. & D. C. Bicudo, 2012. Effect of N and P enrichment on periphytic algal community succession in a tropical oligotrophic reservoir. Limnology 13: 131–141.
Goslee, S. C. & D. L. Urban, 2007. The ecodist package for dissimilarity-based analysis of ecological data. Journal of Statistical Software 22: 1–19.
Hart, D. D., B. J. F. Biggs, V. I. Nikora & C. A. Flindes, 2013. Flow effects on periphyton patches and their ecological consequences in a New Zealand river. Freshwater Biology 58: 1588–1602.
Heino, J., H. Mykrä, J. Kotanen & T. Muotka, 2007. Ecological filters and variability in stream macroinvertebrate communities: do taxonomic and functional structure follow the same path? Ecography 30: 217–230.
Heino, J., D. Schmera & T. Eros, 2013. A macroecological perspective of trait patterns in stream communities. Freshwater Biology 58: 1539–1555.
Jackson, D. A., 1993. Sttoping rules in principal components analysis: a comparison of heuristical and statistical approaches. Ecology 74: 2204–2214.
John, D. M., B. A. Whitton & A. J. Brook, 2002. The Freshwater Algal Flora of the British Isles. An Identification Guide to Freshwater and Terrestrial Algae. University Press, Cambridge.
Kembel, S. W., P. D. Cowan, M. R. Helmus, W. K. Cornwell, H. Morlon, D. D. Ackerly, S. P. Blomberg & C. O. Webb, 2010. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 26: 1463–1464.
Komárek, J. & K. Anagnostidis, 1986. Modern approach to the classification system of cyanophytes 2 – Chroococcales. Algological Studies 43: 157–226.
Komárek, J. & K. Anagnostidis, 1989. Modern approach to the classification system of cyanophytes 4 – Nostocales. Algological Studies 56: 247–345.
Komárek, J. & K. Anagnostidis, 1998. Cyanoprokaryota 1. Teil: Chroococcales. In Ettl, H., G. Gärtner, H. Heynig & D. Mollenhauer (eds), Süsswasserflora von Mitteleuropa 19/1. Gustav Fischer, Jena-Stuttgart-Lübeck-Ulm.
Korhonen, J. J., J. Soininen & H. Hillebrand, 2010. A quantitative analysis of temporal turnover in aquatic species assemblages across ecosystems. Ecology 91: 508–517.
Lange, K., A. Liess, J. J. Piggott, C. R. Townsend & C. D. Matthaei, 2011. Light, nutrients and grazing interact to determine stream diatom community composition and functional group structure. Freshwater Biology 56: 264–278.
Legendre, P. & L. Legendre, 1998. Numerical Ecology, 2nd ed. Elsevier, Amsterdam.
Legendre, P., F. Lapointe & P. Casgrain, 1994. Modeling brain evolution from behavior: a permutational regression approach. Evolution 48: 1487–1499.
Legendre, P., D. Borcard & P. R. Peres-Neto, 2005. Analyzing beta diversity: partitioning the spatial variation of community composition data. Ecological Monographs 75: 435–450.
Leibold, M. A., M. Holyoak, N. Mouquet, P. Amarasekare, J. M. Chase, M. F. Hoopes, R. D. Holt, J. B. Shurin, R. Law, D. Tilman, M. Loreau & A. Gonzalez, 2004. The metacommunity concept: a framework for multi-scale community ecology. Ecology Letters 7: 601–613.
Lichstein, J., 2007. Multiple regression on distance matrices: a multivariate spatial analysis tool. Plant Ecology 188: 117–131.
Lund, J. W. G., C. Kipling & E. D. LeCren, 1958. The inverted microscope method of estimating algal numbers and statistical basis of estimation by counting. Hydrobiologia 11: 143–170.
Mason, N. W. H. & F. De Bello, 2013. Functional diversity: a tool for answering challenging ecological questions. Journal of Vegetation Science 24: 777–780.
McGill, B. J., B. J. Enquist, E. Weiher & M. Westoby, 2006. Rebuilding community ecology from functional traits. Trends in Ecology and Evolution 21: 178–185.
Melo, A. S., 2013. CommEcol: Community Ecology Analyses. R package version 1.5.8/r24. http://R-Forge.Rproject.org/projects/commecol.
Melo, A. S., F. Schneck, L. U. Hepp, N. R. Simões, T. Siqueira & L. M. Bini, 2011. Focusing on variation: methods and applications of the concept of beta diversity in aquatic ecosystems. Acta Limnologica Brasiliensia 23: 318–331.
Nabout, J. C., T. Siqueira, L. M. Bini & I. S. Nogueira, 2009. No evidence for environmental and spatial processes in structuring phytoplankton communities. Acta Oecologica 35: 720–726.
Nekola, J. C. & P. S. White, 1999. The distance decay of similarity in biogeography and ecology. Journal of Biogeography 26: 867–878.
Oksanen, J., F. G. Blanchet, R. Kindt, P. Legendre, P. R. Minchin, R. B. O’Hara, G. L. Simpson, M. P. Solymos, H. H. Stevens & H. Wagner, 2013. Vegan: community ecology package. R package version 2.0-9. http://cran.r-project.org/package=vegan.
Padial, A. P., F. Ceschin, S. A. J. Declerck, L. De Meester, C. C. Bonecker, F. A. Lansac-Tôha, L. Rodrigues, L. C. Rodrigues, S. Train, L. F. M. Velho & L. M. Bini, 2014. Dispersal ability determines the role of environmental, spatial and temporal drivers of metacommunity structure. PLoS One 9: e111227.
Passy, S. I., 2007. Diatom ecological guilds display distinct and predictable behavior along nutrient and disturbance gradients in running waters. Aquatic Botany 86: 171–178.
Passy, S. & C. A. Larson, 2011. Succession in stream biofilms is an environmentally driven gradient of stress tolerance. Microbial Ecology 62: 414–424.
Patterson, B. D. & W. Atmar, 1986. Nested subsets and the structure of insular mammalian faunas and archipelagos. Biological Journal of the Linnean Society 28: 65–82.
Pavoine, S., J. Vallet, A. B. Dufour, S. Gachet & H. Daniel, 2009. On the challenge of treating various types of variables: application for improving the measurement of functional diversity. Oikos 118: 391–402.
Petchey, O. L. & K. J. Gaston, 2006. Functional diversity: back to basics and looking forward. Ecology Letters 9: 741–758.
Piggott, J. J., K. Lange, C. R. Townsend & C. D. Matthaei, 2012. Multiple stressors in agricultural streams: a mesocosm study of interactions among raised water temperature, sediment addition and nutrient enrichment. PLoS One 7: e49873.
R Core Team, 2014. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.r-project.org/.
Roberto, M. C., N. F. Santana & S. M. Thomaz, 2009. Limnology in the Upper Paraná River floodplain: large-scale spatial and temporal patterns, and the influence of reservoirs. Brazilian Journal of Biology 69: 717–725.
Rosenzweig, M. L., 1995. Species diversity in space and time. Cambridge University Press, Cambridge.
Round, F. E., 1971. The taxonomy of the Chlorophyta, 2. Journal of the British Phycological Society 6: 235–264.
Schneck, F., A. Schwarzbold, S. C. Rodrigues & A. S. Melo, 2011a. Environmental variability drives phytoplankton assemblage persistence in a subtropical reservoir. Austral Ecology 36: 839–848.
Schneck, F., A. Schwarzbold & A. S. Melo, 2011b. Substrate roughness affects stream benthic algal diversity, assemblage composition, and nestedness. Journal of the North American Benthological Society 30: 1049–1056.
Schneck, F. & A. S. Melo, 2012. Hydrological disturbance overrides the effect of substratum roughness on the resistance and resilience of stream benthic algae. Freshwater Biology 57: 1678–1688.
Soininen, J., R. McDonald & H. Hillebrand, 2007. The distance decay of similarity in ecological communities. Ecography 30: 3–12.
Souza-Filho, E. E. & J. C. Stevaux, 2004. Geology and geomorphology of the Baía Curutuba-Ivinhema River complex. In Thomaz, S. M., A. A. Agostinho & N. S. Hahn (eds), The upper Paraná River and its floodplain: physical aspects, ecology and conservation. Backhuys Publishers, Leiden: 1–29.
Swenson, N. G., 2011. The role of evolutionary processes in producing biodiversity patterns, and the interrelationships between taxonomic, functional and phylogenetic biodiversity. American Journal of Botany 98: 472–480.
Swenson, N. G., P. Anglada-Cordero & J. A. Barone, 2011. Deterministic tropical tree community turnover: evidence from patterns of functional beta diversity along an elevational gradient. Proceedings of the Royal Society B: Biological Sciences 278: 877–884.
Swenson, N. G., J. C. Stegen, S. J. Davies, D. L. Erickson, J. Forero-Montana, A. H. Hurlbert, W. J. Kress, J. Thompson, M. Uriarte, S. J. Wright & J. K. Zimmerman, 2012. Temporal turnover in the composition of tropical tree communities: functional determinism and phylogenetic stochasticity. Ecology 93: 490–499.
Thomaz, S. M., L. M. Bini & R. L. Bozelli, 2007. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579: 1–13.
Uehlinger, U., B. Kawecka & C. T. Robinson, 2003. Effects of experimental floods on periphyton and stream metabolism below a high dam in the Swiss Alps (River Spöl). Aquatic Sciences 65: 199–209.
Utermöhl, H., 1958. Zur Vervollkommnung der quantitativen phytoplankton-methodic. Internationale Vereinigung für Theoretische und Amgewandte Limnologie 9: 1–39.
Villéger, S. S., G. Grenouillet & S. Brosse, 2013. Decomposing functional beta-diversity reveals that low functional beta-diversity is driven by low functional turnover in European fish assemblages. Global Ecology and Biogeography 22: 671–681.
Wehr, J. D. & R. G. Sheath, 2003. Freshwater Algae of North America. Ecology and Classification. Academic Press, San Diego.
Weiher, E., D. Freund, T. Bunton, A. Stefanski, T. Lee & S. Bentivenga, 2011. Advances, challenges and a developing synthesis of ecological community assembly theory. Philosophical Transactions of the Royal Society B: Biological Sciences 366: 2403–2413.
Wetzel, C. E., D. C. Bicudo, L. Ector, E. A. Lobo, J. Soininen, L. Victor & L. M. Bini, 2012. Distance decay of similarity in neotropical diatom communities. PLoS One 7: e45071.
Wright, D. H., B. D. Patterson, G. M. Mikkelson, A. Cutler & W. Atmar, 1998. A comparative analysis of nested subset patterns of species composition. Oecologia 113: 1–20.
Acknowledgments
We acknowledge Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for a doctoral scholarship granted for Bárbara Dunck; Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for a productivity grant to Liliana Rodrigues and a research grant to Fabiana Schneck; Núcleo de Pesquisa em Limnologia, Ictiologia e Aquicultura (Nupélia) for technical and logistical support; Laboratory of Limnology of Nupélia for analysis of environmental variables; and Sidinei Magela Thomaz for comments on an earlier version of this manuscript. This study was supported by the Long Term Ecological Research Program (PELD-CNPq).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Stefano Amalfitano
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Dunck, B., Schneck, F. & Rodrigues, L. Patterns in species and functional dissimilarity: insights from periphytic algae in subtropical floodplain lakes. Hydrobiologia 763, 237–247 (2016). https://doi.org/10.1007/s10750-015-2379-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-015-2379-x