
Abstract
We aimed to establish the effect of seasonal drought on leaf litter breakdown and invertebrate communities. Differences in breakdown rates of Nothofagus pumilio were experimentally compared using the litter bag method (coarse and fine mesh size bags) in two first-order streams, one intermittent and one perennial, during two different hydrological periods. Colonizing fauna found in coarse mesh bags was quantified, identified and compared with benthic biota from the same streams. Leaf litter decay rates in low flow conditions revealed that breakdown was principally a consequence of microbial action in the intermittent stream. In contrast, breakdown in high flow conditions was caused by invertebrate feeding in both streams. Collector–gatherers constituted most of the abundance and biomass in bags from the intermittent stream, due to their rapid benthic recolonization. Shredders peaked at approximately 50% remaining leaf litter mass in both streams only during high flow, which coincided with general models of detritus breakdown in streams. Considering global warming scenarios, with drought and water temperature increases expected for many regions of the world, these studies on the consequences for the biota and ecological processes of small streams will allow the prediction of negative effects on such vulnerable ecosystems.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Headwater streams have the lowest flows relative to other parts of the stream network, and are therefore highly vulnerable to drought disturbance. While permanent streams essentially flow during the entire year, intermittent streams are characterized by lower flow conditions and a dry phase, making these systems challenging habitats for aquatic biota (Richardson & Danehy, 2007). Alterations in flow regimes may have multiple impacts on biodiversity, firstly as a determinant of the physical habitat, and also by bringing about changes in the natural patterns of lateral and longitudinal connectivity which are typical of fluvial systems (Bunn & Arthington, 2002). Therefore, the community composition of permanent streams may undergo considerable change, developing non-perennial stream biota adapted to either withstanding (by resistance mechanisms) or recovering from (i.e., being resilient to) drought (Lake, 2000; Chessman, 2015).
Stream functioning may also be altered by drought. In particular, leaf litter breakdown is strongly dependent on water availability and is thus expected to progress more slowly in intermittent streams (Kirby et al., 1983; Maamri et al., 1997). There are several reasons for this: leaching proceeds more rapidly in aquatic environments (Treplin & Zimmer, 2012); microbial decomposer biomass and activity is usually higher in submerged leaves (Bruder et al., 2011), although this may depend on leaf species (Treplin & Zimmer, 2012) and water temperature; and finally, because shredders are more abundant in permanent streams (Kirby et al., 1983; Datry et al., 2011).
As for other regions of the world, global change scenarios anticipate decreasing precipitations for Patagonia (Nuñez et al., 2009; Dai, 2013), which would increase the number of intermittent streams within stream networks and the length of their dry phase. In addition, global tendencies of increasing temperatures should result in less snow deposition and earlier melting, anticipating peak discharges in winter-spring and intensifying intermittency of small streams during the dry season (Breshears et al., 2005). Considering these predictions, it is remarkable that organic matter (OM) breakdown has received little attention in intermittent lotic systems and has not been explicitly included in general models of OM fluxes in fluvial systems (Maamri et al., 1997; Treplin & Zimmer, 2012).
In deciduous forests the effect of drought on breakdown would differ according to the timing of leaf abscission. If leaf fall coincides with cessation of the surface flow, leaves may accumulate in residual pools and/or the surface of the dry streambed (Acuña et al., 2005; Dieter et al., 2011). But if flow resumes quickly after leaf abscission, drought is unlikely to have an effect on breakdown (Corti et al., 2011). However, Datry et al. (2011) found that temporary streams remain persistently affected by drying events that have occurred throughout the previous summer season. They suggested that the absence of water before leaf litter input in the stream affects structural characteristics such as invertebrate assemblage abundance and composition, which determine breakdown rates. Temperature also affects breakdown rates (Ferreira & Canhoto, 2014), and it may differ between intermittent and perennial streams, and also temporally. Therefore, the comparison of decay rates corrected by cumulative degrees over experimental days takes into account the potentially important differences in temperature between stream conditions (Benfield, 2011).
One main driver of leaf litter breakdown in streams is the shredding invertebrate component. Studies in perennial low-order streams of the Patagonian Andes revealed that the main primary consumers in terms of density and biomass are benthic invertebrates belonging to shredder, collector–gatherer, and scraper functional feeding groups (FFG) (Albariño & Díaz Villanueva, 2006; Miserendino, 2007) and that temporal variations in benthic invertebrate community dynamics are synchronized with basal resource abundance (Díaz Villanueva et al., 2010). Patch resource colonization by shredder and collector–gatherer invertebrates plays an important role in stream ecosystem functioning (Cheshire et al., 2005; Boyero et al., 2011) as they cycle and transfer OM and nutrients both up the food web and spatially across the lateral, longitudinal, and vertical axes of such ecosystems. Cummins et al. (1989) proposed a model that describes leaf litter colonization by these FFGs. The model predicts that shredders will peak when leaf litter mass reaches 50% of initial mass and collector–gatherers will do so when leaf litter breakdown is advanced (leaf mass loss > 50%), as proportionally more fine particulate OM is expected to accumulate.
In order to compare the effects of flow intermittency in two contrasting phases of the hydroperiod, we conducted two leaf litter breakdown experiments in each of two first order, forested streams: one intermittent and one perennial. The first experiment began in autumn, before initiation of the hydroperiod (i.e., when the lowest flow is observed in perennial streams), and the second experiment started in winter, when the flow had completely recovered. We expected litter breakdown rates to differ between streams in both experiments due to the “drying memory” of the system as described by Datry et al. (2011). Also, we expected litter breakdown rates to vary with time (i.e., between experiments) because of differences in stream conditions (e.g., thermal regimes, invertebrate abundance and composition dynamics, natural litter availability, and conditioning). We tested the following specific hypotheses: (1) litter breakdown rates are higher in the perennial stream in both hydroperiods, as a consequence of a fully established benthic invertebrate assemblage and higher shredder abundances compared to the intermittent stream; (2) shredder abundance and biomass in litter bags peak when litter remaining mass is 50% of initial mass, while collector–gatherer abundance and biomass peak when litter breakdown is advanced (litter mass loss > 50%) and (3) shredder density is similar in litter bags and natural leaf packs in early winter but their densities are higher in litter bags than in stream leaf packs in early spring because litter stock is temporally depleted after the autumnal allochthonous pulse input; in other words, leaf litter availability in stream beds becomes temporally limiting. The outcomes of testing these hypotheses will serve as a primer on the knowledge of first-order stream processes under drought events and will make predictions possible as to changes in the structure and functioning of these vulnerable ecosystems.
Methods
Study area
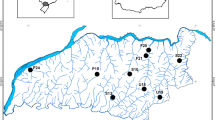
This study was conducted in two low-order streams belonging to Ñireco river basin (San Carlos de Bariloche—Nahuel Huapi National Park, Argentina) (41°14′S 71°20′W). This basin drains an area densely covered with Nothofagus pumilio (Poepp. et Endl.) Krasser (common name: “lenga”), the deciduous mountain beech, which forms monospecific forests constituting the highest mountain tree belt of the temperate rainforest up to the timberline along the Patagonian Andes. Streams are characterized by pebble–cobble bottoms and low nutrient concentrations (associated with the igneous origin of the geological landscape) (Modenutti et al., 2010). In other low-order streams from the same catchment area, nutrient concentrations were reported to be at the limit of detection (Buria et al., 2010). Allochthonous leaf litter inputs into streams are dominated by N. pumilio leaves which represent more than 95% of total leaf litter standing stock (Albariño et al., 2009). The area receives ~2000 mm year−1 as rainfall and snow, following a bimodal hydrological regime with high discharges in late autumn and spring (rainfall and snowmelt, respectively) and the lowest discharge in summer–early autumn.
The streams were selected by field inspection and satellite photography to ensure that they were comparable in general characteristics, such as catchment and riparian vegetation, catchment area, altitude, nearness, gradient, and orientation. Although no geological and hydrological studies have been done in the area, differences in stream hydrology (i.e., perennial vs. intermittent condition) are probably driven by differences in subsurface catchment volume that result in different phreatic water accumulations.
The experiments were run in one perennial and one intermittent (hydroperiod: 6–8 months) neighboring unnamed streams, and were repeated at two different times within the 2011 hydroperiod. The first experiment started in April, before the intermittent stream had surface running water, and continued for 129 days. The second experiment was conducted after the hydroperiod had begun in the intermittent (June) and was run for 136 days (Fig. 1a). During a previous inspection (summer 2011) in a dry phase of the intermittent stream channel, we found empty sand cases of a caddisfly species. This finding suggests that the hydroperiod in this stream was long enough to allow some invertebrate species to fulfill their aquatic life cycles.

a Water level, extraction dates of leaf litter bags, and discharge values for extraction dates in the two streams during the study period. Circles (experiment 1) and cross (experiment 2) correspond to t 0–t 4 litter bag retrieval dates. Arrows indicate benthic survey dates. b Temperatures in the intermittent and perennial streams, and also in the riparian area during study. Lines show water depth fluctuations, inverted triangles show discharge values obtained during extraction dates
Physicochemical and morphometric variables
Prior to the setting of the first experiment, two water level and temperature data loggers (HOBO U20, Onset Co.) were launched in each stream, recording these variables every 10 min during the study. An additional logger deployed at the margin of the intermittent stream recorded air temperature, enabling us to correct the air barometric pressure from the stream surroundings, in order to obtain water depth fluctuations in each stream. In addition, conductivity was measured on every sampling date with a multiprobe instrument (YSI 85, YSI Inc.). Discharge was measured by the addition of a conservative tracer to the stream, and recording conductivity changes over time (Gordon et al., 2004). Water samples were collected in August 2011 (between experiments 1 and 2) in acid-washed bottles and transported on ice to the laboratory where they were analyzed for dissolved nutrients (ammonium, nitrates, and inorganic P). Ammonium was measured with the indophenol blue method (APHA, 2005) and NO3 − was measured as NO2 − after reduction through a cadmium–copper column (Wood et al., 1967), and quantified spectrophotometrically. Soluble reactive P was determined by the ascorbate-reduced molybdenum blue method (APHA, 2005).
Breakdown experiment
We used the litter bag methodology (Graça et al., 2005) to test breakdown rates and invertebrate colonization in both streams. Undamaged freshly fallen leaves of N. pumilio were collected in the same study area during the previous season from the riparian floor during leaf abscission and dried at room temperature. Experimental units consisted of plastic leaf bags (20 × 12 cm) of two different mesh sizes, containing 3.00 g fresh mass of lenga leaf litter. The 5 mm coarse mesh bags allowed invertebrate colonization and shredder activity and the 500 μm fine mesh bags excluded invertebrate driven decay but permitted microbial decomposition (i.e., bacterial and fungal activity). The design of our experiments was bi-factorial, considering bag mesh size (two levels: coarse and fine), and stream (two levels: perennial and intermittent) as factors.
The first experiment was set on 8 April (autumn in the Southern Hemisphere), and the second started on 17 June (just before the onset of winter). On each date, 20 experimental units were placed in the streambed along a 30 m reach at each site and secured with metal rods along the stream bottom. Five pairs of bags were removed after 18, 39, 81, and 129 days of exposure in experiment 1 and after 18, 59, 101, and 136 days in experiment 2 (i.e., t 1, t 2, t 3, and t 4 for each experiment). Scheduled days were set to span more than 4 months of experimental exposure and were adjusted in accordance with weather conditions and previous data. Mass loss due to leaching was determined in the laboratory, by preparing two sets of five bags with previously weighed leaf litter submerged in tap water. After 24 or 48 h, the samples were dried in the furnace (60°C, 48 h) to estimate mass loss after one and 2 days of aquatic conditions, and these values were considered for all decay models.
In the laboratory invertebrates were separated from remaining leaf material in a well-illuminated white tray and were fixed and preserved in 70% ethanol for further processing. Remaining leaf litter (i.e., leaves and leaf fragments > 5 mm) from each bag was collected, weighed after drying at 60°C for 48 h, incinerated at 400°C for 1 h and weighed to the nearest 0.01 g. The ash weight was subtracted from the dry weight to obtain ash-free dry mass (AFDM).
An exponential decay model was adjusted in both experiments for percentage of remaining leaf litter AFDM as a function of exposure time. Breakdown rates k (d−1) were estimated assuming the following model (Graça et al., 2005):
where M t is the final remaining leaf litter AFDM, M 0 is the initial AFDM, k is the breakdown rate and t is the time in days.
Since experiment 1 was set in the intermittent stream when there was no surface water, we first explored a decay model that considered the initial dry phase (i.e., t 0, t leach24, t leach48, t 1, and t 2) and the final wet phase (t 0′, t 3, and t 4; t 0′ being defined as the estimated breakdown of the first day of continuous running water) independently. In the dry phase, this model overestimated mass loss due to leaching and both curves had poorly adjusted r 2 values (<0.60). Alternatively, a model integrating both phases was tested. However, as an isolated water flow pulse took place thirteen days after t 0, leaching mass loss to fit this model was assumed to occur at this recorded water pulse instead of the usual 24 and 48 h.
Breakdown rates (k) were additionally estimated as a function of total degree days by summing mean daily water temperature (obtained from data logger temperature series) accumulated from the beginning of the experiments to the day of leaf litter bag retrieval from streams.
Benthic invertebrates
Benthic field surveys were conducted in June and September 2011 in both streams to quantify benthic invertebrate abundance and biomass at each experimental site and compare species composition and abundance in the benthos and in the bags. The survey dates were chosen as they coincided with t 3, around 90 days of leaf conditioning, which is considered ideal in this species for invertebrate consumption. Samples were taken with a Surber sampler (mesh size = 200 µm, surface area = 0.06 m2) in erosional areas (riffles, n = 3) and with a modified corer sampler (surface area = 0.03 m2) in depositional reaches (pools, n = 3) to include both types of habitat in the comparison. Samples were transported to the laboratory, where they were fixed and preserved in 5% formalin until processing. Samples were washed through 500 and 1000 µm sieves to facilitate invertebrate sorting. Invertebrates >500 µm were preserved in 70% ethanol for counting and identification.
Invertebrates from experimental units and benthic surveys were observed under stereomicroscope and identified to the lowest possible taxonomic level following keys for South American taxa (Domínguez et al., 2009). We also assigned invertebrates to their corresponding FFGs according to Merritt & Cummins (1996): shredder, scraper, collector–filterer, collector–gatherer, and predator, following previous studies in North Patagonian streams (Albariño & Díaz Villanueva, 2006; Miserendino, 2007; Díaz Villanueva et al., 2010). In this study, only the abundance and biomass of the deposition detritivore fraction, i.e., shredders and collector–gatherers, were analyzed. All taxa were quantified in number and biomass, obtained by drying organisms at 60°C for 24 h and weighing to the nearest 0.01 mg. Relative values were referred to as number of individuals per bag or area unit, and grams of dry mass for each experimental or sample unit.
We calculated a “crowdedness” index for the comparison of shredder abundance in leaf litter coarse mesh bags with their abundance in “ambient” leaf litter standing stock. For this calculation only values for t 3 were analyzed, as it was the time when remaining leaf litter in bags was near 50% of its initial mass and because benthic sampling took place near those dates. Shredder abundance in benthic samples was expressed for the same surface area as bags (240 cm2). Finally, mean values for bags were plotted against the mean values for benthic samples of each stream (i.e., shredder abundance in bags of experiment 1 vs. June survey and those of experiment 2 vs. September survey). If values fell far from a 1:1 line, shredders were colonizing leaf litter at different densities to those in the stream bottom.
Statistical analysis
Two-way analysis of covariance (ANCOVA) was used to test (i) differences in breakdown rates between experiments (i.e., between hydroperiods) and streams, in coarse mesh bags, and (ii) differences between mesh types and streams in each experiment (Hypothesis 1). In the models, the natural logarithm of remaining AFDM was the dependent variable, experiment and mesh size were fixed factors, stream was random factor and days or cumulated degree days were covariates. In particular, comparisons of breakdown rates between experiments and streams (i) were carried out using only coarse mesh bag treatment with k′ values, to eliminate the effect of potential differences in temperature between hydroperiods. By doing this, we focused the comparison on the effect of shredder assemblages and simultaneously avoiding the indirect effect of different temperatures between stream types and hydroperiods. ANCOVAs were performed with STATISTICA 11 software package.
To assess differences in total invertebrate abundance and biomass, and abundance and biomass of detritivore FFGs colonizing coarse mesh bags (Hypothesis 2) between streams (subject factor) and extraction dates (treatment factor), we used two-way repeated measures ANOVA. Repeated Measures ANOVAs were performed with SPSS software package. Finally, shredder “crowdedness” in bags and benthos was compared using Mann–Whitney U test (Hypothesis 3), with SIGMA PLOT 11.0.
Results
During experiment 1 surface flow was registered in the intermittent stream on only 79 out of 129 days (60%); 4 days between t 0 and t 1 (from day 13 to 16), associated with a single, isolated storm, and from 8 June (after t 2, Fig. 1a). The first 4 days of running water in the intermittent stream were probably due to the accumulation of precipitations in the previous weeks, which were gradually recharging ground water in the catchments of this region. Water depth changes in the perennial stream during the same period reflected stream flow variation associated with precipitations in the area. During experiment 2, both streams had surface water, but the perennial was always deeper and had higher discharge than the intermittent (Fig. 1a). Except for the days when the intermittent stream was dry, water temperature was lower in the perennial stream during both experiments (low discharge and high discharge conditions) (Fig. 1b). In both cases, the difference in mean temperature between streams was nearly 1°C, but during the whole study period, it was more variable in the perennial (CV = 85%) than in the intermittent (CV = 55%). Water depth increased in both streams from the first to the second experiment, but this increase was proportionally lower in the perennial than in the intermittent (Fig. 1a; Table 1). Discharge also increased in both streams throughout this experimentation period, but it was always higher in perennial stream (Fig. 1a). Nutrients were low and close to the limit of detection in both streams (Table 1).
When comparing breakdown rates (k′) between stream types and experimental periods in order to assess the seasonal effects of hydroperiod conditions on invertebrate community efficiency, we found that leaf litter decomposed faster in the perennial stream (ANCOVAStream: F 1,144 = 20.4; P < 0.001) and also faster in experiment 2 (ANCOVAExperiment: F 1,144 = 29.9; P < 0.001).
In experiment 1, decay rates differed depending on mesh size and stream (ANCOVAStream × Mesh size, Table 2). In the intermittent stream leaf litter decomposed similarly regardless of the bag mesh size; mass loss after 129 days (t 4) was 32 and 34% (of initial mass) for the coarse and fine mesh, respectively (Fig. 2a), while in the perennial stream, leaf litter decomposed twice faster in coarse mesh bags than in fine mesh bags (mass loss at t 4 = 55 vs. 27%; Fig. 2b) and was also two-fold faster than in bags from the intermittent stream (Tables 2, 3). In experiment 2, leaf litter decomposed significantly faster in coarse mesh bags, without differences between streams (Table 3; Fig. 2c, d).

Percentage of leaf litter mass remaining during experiments in the intermittent and perennial streams. Dots are mean ± 1 SE. Coarse and fine mesh bags (empty and filled symbols, respectively). Solid and dashed lines (fine and coarse mesh bags, respectively) represent the exponential leaf litter decay models estimated for each dataset in experiment 1 (a, b) and experiment 2 (c, d) against days elapsed. Shaded area in panel a corresponds to drought conditions
The patterns of decay rates calculated in degree days instead of days were similar in experiment 1, i.e., leaf litter in the perennial stream decomposed faster in coarse mesh bags than in fine mesh bags, and also faster in this stream than in both treatments in the intermittent one (Table 3). In contrast, in experiment 2 cumulated degree days were 85% higher than in experiment 1 in the intermittent stream but temperature-corrected decay rates (k′) were significantly higher in the perennial one (Table 2). Regarding mesh size, k′ was significantly higher in coarse mesh bags from both streams, i.e., the same pattern as for k decay rate (Table 2).
Total invertebrate abundance and biomass were similar in coarse mesh bags from both streams in experiment 1 (Table 4). Temporal colonization was different between streams, increasing with time in the intermittent stream (in terms of biomass) but peaking at t 2 and then decreasing in the perennial stream (Fig. 3a, b). Mean invertebrate size was 70% higher in the perennial stream and in both streams it increased over time (Table 4; Fig. 3c), except at the third extraction when it abruptly decreased in the perennial stream (Fig. 3c). In experiment 2, temporal colonization (both abundance and biomass) differed between extraction dates in both streams (Table 4). Abundance and biomass increased over time in the intermittent stream and both peaked at t 3, while in the perennial stream abundance and biomass peaked at t 4 (Fig. 3d, e). Mean invertebrate size also showed this pattern, peaking at t 3 in the intermittent stream and t 4 in the perennial stream, and as in experiment 1, it was 95% higher in the perennial stream (Table 4; Fig. 3f).

Total abundance of benthic invertebrates colonizing leaf litter bags in experiment 1 (a) and 2 (d), and total biomass of benthic invertebrates (b, e). Mean invertebrate size in both streams along extraction dates, experiment 1 (c) and experiment 2 (f). Bars and error bars are mean ± 1 SE. Colors in bars represent relative abundance of shredders, collector–gatherers, and other FFGs
A total of 24 invertebrate taxa were collected in the benthic surveys of both streams (Table 5). They belonged to three phyla (Annelida, Arthropoda, and Platyhelminthes) but aquatic insects dominated the assemblages. Twenty-three taxa were found in the perennial stream, ten of which were detritivores (seven shredders and three collector–gatherers), eight were predators, three scrapers, and two collector–filterers (Table 5). Seven species were exclusive to this stream, i.e., not found in the intermittent, and 87% of overall species were found in experimental bags. On the other hand, the intermittent stream had a simpler benthic community, total richness was 15, and all FFGs had fewer representatives than in the perennial one, for instance detritivores were represented by seven taxa (four shredders and three collector–gatherers), five predators, two scrapers, and one collector–filterer. Eleven taxa (73% of overall species) were found in experimental units from this stream. The shredder caddisfly Verger capillatus (Ulmer) was the only taxa found exclusively in the intermittent stream (Table 5).
Shredder abundance did not differ between streams or extraction dates. Shredder biomass was different between streams only in experiment 1, being in average seven-fold higher in the permanent stream (Table 4; Fig. 3a, b). Collector–gatherers were more abundant in the intermittent stream, increasing over time in terms of abundance and biomass in this stream and peaking at t 2 in the perennial (Table 4; Fig. 3a, b). In experiment 2, shredder and collector–gatherer abundance and biomass increased over time in both streams (Table 4; Fig. 3d, e), although in the intermittent, collector–gatherers peaked at t 3 (Fig. 3d, e) and biomass was higher than in the perennial (Table 4).
Leaf litter in the benthos was 13.1 and 23.9 g m−2 in June and 0.5 and 2.8 g m−2 in September in the intermittent and perennial streams, respectively. Therefore, it reflected 96 and 89% reductions in benthic leaf litter abundance between dates. The comparison of shredder “crowdedness” per unit area between litter bags and Surber samples showed no differences in any of the streams in both experiments (Fig. 4). However, in experiment 2 shredders were slightly more abundant in litter bags than in the benthic samples of the intermittent stream (i.e., values fell outside the 1:1 line, Fig. 4). The opposite trend was observed in the perennial stream: shredders tended to be more abundant at the stream bottom.

Shredder abundance per surface in litter bags at t 4 (benthic experiment) and in Surber samples (benthic survey) from June to September 2011. Int and Per are the acronyms for the intermittent and perennial streams. Abundance in litter bags of experiment 1 are compared with abundance in Surber samples from June survey. Abundance in litter bags of experiment 2 are compared with September samples. Values represent mean ± 1 SE. Dashed line is a 1:1 abundance for reference
Discussion
Our experimental design made possible the comparison of leaf litter breakdown in two first-order mountain forest streams with contrasting hydroperiods at two different moments of the hydrological cycle. Overall, leaf litter breakdown was controlled in different ways by detritivores and decomposers, modulated by differences in surface water flow and temperature. Our first hypothesis of different decay rates in streams during both experiments was partially rejected, as leaf litter decay rates differed between intermittent and perennial streams only at the beginning of the hydroperiod (experiment 1, early autumn). Breakdown was faster in the perennial stream only in experiment 1, showing resilient functioning in the intermittent stream. Our results indicated that in experiment 1, invertebrates in the intermittent streams had no measurable effect on breakdown while in the perennial stream this process was faster in the litter bags with invertebrate access.
Detritivore colonization in litter bags differed from the proposed model in low discharge conditions, but was confirmed in high discharge conditions when shredders peaked near 50% remaining leaf litter in both streams (Hypothesis 2). However, shredders did not display the expected pattern, since they did not prefer litter bags to natural benthic leaf accumulations as food resource patches when litter abundance was seasonally declining (experiment 2) (Hypothesis 3). Although this result was counterintuitive, it supports our measured N. pumilio decay values in coarse mesh bags, which would otherwise be overestimated regarding benthic shredder crowdedness if higher shredder abundance had been feeding inside the bags than in the ambient condition.
Leaf litter breakdown
Breakdown of N. pumilio leaf litter measured in this study might classify as a slow-medium decay species (k ~ 0.005 in most cases, though slightly higher in coarse mesh bags) according to Webster & Benfield (1986) categorization. This coincides with previous studies on litter decomposition of such species in higher order streams of the same basin (k = 0.0036–0.0049), with water temperatures of 2°C (Albariño et al., 2009). In a stream in central Chile, Valdovinos (2001) reported a lenga leaf litter k rate slightly higher (k = 0.0072) than the ones obtained in the present study. Interestingly, that stream system was characterized by fewer shredders and higher water temperatures in winter–spring (up to 14°C) than our streams. As the studies on lenga leaf litter breakdown have been carried out in streams located at its Northern distribution, and lenga forms a monospecific tree belt up to the timberline along the Patagonian Andes (from 39 to 55°C), it would be interesting to test whether leaf litter breakdown is even slower at higher and harsher latitudes.
Leaf litter breakdown is usually faster in aquatic than in terrestrial environments, due to more efficient litter conditioning: i.e., leaching of organic compounds, higher fungal and bacterial biomass, and invertebrate consumption under constant water conditions (Richardson et al., 2009; Bruder et al., 2011; Treplin & Zimmer, 2012). However, in low discharge conditions, leaf litter decomposition by microorganisms (fine mesh bag treatment) was similar in both streams, although litter in the intermittent stream was exposed to aquatic conditions for only 60% of the total experiment length. It is likely that frequent rains during the dry phase (12 out of 50 days, source AIC: Intergovernmental Water Bureau of the Río Negro Basin, Argentina) might have maintained moisture conditions high enough to sustain microbial activity within leaf packs. Although some studies have proven that the effect of water intermittency on leaf litter decay strongly reduced k values (Bruder et al., 2011; Corti et al., 2011), showing the importance of aquatic conditions in microbial driven decomposition process, we suggest that low temperature and low nutrient concentrations determine low microbial activity in our mountain streams, regardless of their hydroperiod.
The slower leaf litter breakdown rate found in coarse mesh bags from the intermittent stream in low discharge conditions could also be attributed to differences in shredder activity (reflected by lower biomass and densities than those in the perennial stream), as previously stated (Kirby et al., 1983; Richardson, 1990; Maamri et al., 1997). This pattern seems intimately related to detritivore life cycles which need to be synchronized with the hydroperiod of intermittent streams. Among the many possible strategies used by invertebrates to elude drought (Robson et al., 2011), desiccation-resistant eggs represent an important difference when compared with the life cycles of biota from perennial streams. This would lead to an initial (rewetting) phase when these eggs may hatch almost simultaneously, and the incipient benthic community becomes dominated by first colonizers, mostly early instars of aquatic insect larvae. As the highest leaf litter consumption by detritivore insects is usually attained at highest individual biomass (González & Graca, 2003), and the early instars of some shredders may behave as collector–gatherers (López-Rodríguez et al., 2012), it seems reasonable that during experiment 1 decay rates in coarse and fine mesh bags in the intermittent stream were similar, and were both lower than in the perennial one where a benthic community was fully established.
During high discharge conditions, water was never a limiting factor and the invertebrate community had already developed in the intermittent stream. Interestingly, higher water temperatures in the intermittent stream were a determining factor, reflected by the differences between streams when temperature effect on decomposition was eliminated (k′). Decay rate (k) in the fine mesh bags was similar between streams because temperature stimulated decomposition in the intermittent stream. It is well documented that temperature has an effect on decay (Ferreira & Chauvet, 2011a, b), by stimulating biological activity, mainly controlling the enzymatic reactions of microbes (Bärlocher & Kendrick, 1975; Ferreira & Canhoto, 2014).
Finally, considering the adjusted decay models and the estimated dates for 50 and 90% mass disappearance (Table 3), it seems plausible that OM dynamics between intermittent and perennial streams will be markedly different as ruled by water presence. Lenga leaf litter is a low-quality food resource with slow breakdown, which may take more than 1 year to decompose completely. Therefore, water presence for periods of 6–8 months, as in our study stream, would determine that OM is not cycled within the aquatic system. It is unclear whether during the dry phase OM accumulates in the stream channel until the next hydroperiod to reenter the water cycling pathway, or whether it is substantially recycled by terrestrial decomposers.
Detritivore colonization
Detritivore invertebrates function as key consumers in headwater forested streams, as they process huge amounts of allochthonous OM from the surrounding riparian forest. This is a consequence, in part, of the low quality of this OM resource (i.e., high C:N ratios) and the low growth efficiencies of many shredders (Balseiro & Albariño, 2006; Graça & Canhoto, 2006). Therefore, these basal consumers substantially connect terrestrial organic inputs with top predators in stream and riparian food webs (Wesner, 2010).
Resilience has proven to be more important than resistance in terms of recovery from disturbances associated with flow regimes (drought and floods) in headwater streams, i.e., the community recovers faster by means of colonization than by tolerance mechanisms (Caruso, 2002; Fritz & Dodds, 2004). In our intermittent stream, the benthic community was poor compared to the perennial stream (15 vs. 23 taxa for total richness) even after the water cycle had been appropriately reestablished. This difference was mirrored in the detritivore guilds (considering both shredders and collector–gatherers) accounting for 7 versus 10 taxa, respectively. In particular, shredder richness clearly differed between streams (4 species in the intermittent vs. 7 in the perennial, respectively). These values are consistent with results for other small streams in the area. In two perennial streams (first and second order) in the same forested catchment, Albariño et al. (2009) described invertebrate benthic communities with the same seven shredder species (except for the only species characteristic of the intermittent stream, Table 5). Regardless of their different taxonomic richness, the high abundance and biomass of detritivores recorded in our study systems emphasizes the existence of detritus-based communities in small, forested mountain streams, similar to the results of other studies (Dieterich & Anderson, 1995; Cheshire et al., 2005; Richardson et al., 2009).
In this work, the patterns of invertebrate colonization in experimental bags, especially detritivore dynamics, were different in the two streams. In low discharge conditions, abundance was similar between streams, but invertebrate mean size was lower in the intermittent one. This was due to the small individual size of early larval stages of aquatic biota that began to colonize the stream bottom after approximately 5 months without surface flowing water. In this case, the first extraction was dominated by small individuals of the shredder caddisfly V. capillatus which provided most of the numerical abundance (~90%), but it seems they moved from bags to wetter microhabitats, since they abruptly decreased at the second extraction date (performed during drought conditions) and increased again at the last two extractions. Other taxa found at this early stage were mainly oligochaetes and chironomids, which are reported as common post-drought first colonizers (Otermin et al., 2002; García et al., 2008). Both groups are collector–gatherers and this explains why this FFG contributed overall higher relative abundance and biomass in litter bags in the intermittent stream rather than in the perennial one. Although it was predicted that this guild would increase when leaf litter breakdown was advanced (Cummins et al., 1989), in our intermittent stream litter bag colonization reflected the colonizing pattern of the stream bottom, and this could be the reason for the high abundance of collector–gatherers during this part of the hydroperiod. On the other hand, in the perennial stream, shredders were more important colonizers than collector–gatherers in both conditions, reflecting the fact that perennial benthic communities do not undergo such a pronounced recolonization stage (i.e., dominated by chironomids and oligochaetes) and that litter bags represented an attractive resource for the shredder guild. Overall, collector–gatherers did not show a clear increase in abundance or biomass as leaf litter mass decayed, because total mass loss in experiments was slightly higher than 50% and this precluded us from testing our hypothesis on this FFG.
Shredders prefer leaf litter that has been colonized and biochemically enriched and softened by fungi and bacteria, which usually occurs when leaf litter mass reaches 50% of its initial mass (Graça & Canhoto, 2006), thus their abundance and biomass should have peaked at this stage in experimental bags. In both conditions (low and high discharge), shredder abundance and biomass tended to increase in leaf litter bags as leaf mass disappeared. Only during experiment 2 did shredders peak near 50% remaining leaf litter mass in both streams, as predicted in Cummins et al. (1989), showing a regular pattern for both individual numbers and biomass. Regarding experiment 1, leaf litter loss in the intermittent stream was just 25% of total mass, preventing us from reaching any conclusion on this topic. In the perennial stream, where final leaf litter losses were 55%, shredder abundance and biomass as well as invertebrate size were high at t 2, but then they increased again at t 4, in partial agreement with the model. This result is in accordance with the finding that discharge conditions are needed to ensure the breakdown process, as shredder efficiency is enhanced by full motility and accessibility to litter packs when totally submerged (Leberfinger et al., 2010).
We had suggested that experimental leaf litter packs would be a more attractive resource than ambient benthic leaf litter only in spring because, this resource is expected to: (i) be abundant and of assorted quality in streambeds only at the time of leaf shedding (experiment 1) and (ii) become limiting as the annual OM breakdown proceeds. Our results did not support expectations, as only our ‘shredder crowdedness index’ in litter bags was slightly higher than in the stream bottom. However, crowdedness was higher in experiment 2 than in 1 showing a trend to have shredders concentrated in litter accumulations both in bags and stream bottoms. Leaf litter detritus may represent a limiting resource for shredder guilds at certain moments of the year (Richardson, 1991; Albariño et al., 2009). Several studies have found temporal confluence of leaf litter inputs and shredder life cycles in streams (Cummins et al., 1989; Richardson & Danehy, 2007; Boyero et al., 2011), and such a link could be intensified in monospecific forests like those of N. pumilio in Northwest Patagonia, where allochthonous plant detritus comes almost exclusively from this species.
Among the biota found in these first-order streams, it is important to notice the presence of one caddisfly species (V. capillatus) found exclusively in the intermittent one. Case remains and small populations of this species have already been found along the driest border of the Andean rain forest in the region, in several small-sized temporary streams (Albariño and Mariluan, pers. obs.). We suggest that this species may function as an indicator of water intermittency in this catchment and possibly in areas with similar environmental conditions and forest canopy. The role of V. capillatus in intermittent systems of North Patagonia as an OM cycler and as a biotic indicator of stream ecological integrity are subjects that deserve further attention.
Conclusions
In this work, we focused principally on a key ecosystem process, i.e., detritus breakdown, and invertebrate colonization, aiming to contribute to the general understanding of the ecological functioning of small stream ecosystems subject to drought. Our study revealed resilient functioning of leaf litter breakdown and invertebrate benthic recolonization in the intermittent stream after surface flow had been restored. At the same time, it suggests that OM dynamics differ markedly between intermittent and perennial streams in an annual basis. In forested regions where small streams are characteristically supplied by low-quality leaf litter (i.e., slow breakdown rates), which takes many months to decompose completely, detritus processing is likely to be disrupted after channels dry up, mainly due to the absence of both surface water and aquatic shredders. Low-order headwater streams are vulnerable to anthropogenic alterations due to their size, e.g., alterations in the surrounding landscape, riparian zones, stream channels, and water flow. In a global change context with increasing human population putting urbanization pressure on natural systems, and temperatures rising globally at the same time, the thermal and hydrological regimes of these systems could undergo profound changes. Our results suggest that the impact on headwater systems will depend on the intensity and length of the disturbance, since water availability is a key factor in faunal distribution and ecosystem processes.
References
Acuña, V., I. Muñoz, A. Giorgi, M. Omella, F. Sabater & S. Sabater, 2005. Drought and postdrought recovery cycles in an intermittent Mediterranean stream: structural and functional aspects. Journal of the North American Benthological Society 24: 919–933.
Albariño, R. & V. Díaz Villanueva, 2006. Feeding ecology of two plecopterans in low order Andean-Patagonian streams. International Review Hydrobiology 91: 122–135.
Albariño, R., V. Díaz Villanueva & L. Buria, 2009. Leaf Litter Dynamics in a Forested Small Andean Catchment, Northern Patagonia, Argentina Ecological Advances on Chilean Temperate Rainforests. Academia Press, Gent: 183–211.
APHA, 2005. Standard Methods for the Examination of Water, Sewage, and Wastewater. American Public Health Association, Washington, DC.
Balseiro, E. & R. Albariño, 2006. C-N mismatch in the leaf litter–shredder relationship of an Andean Patagonian stream detritivore. Journal of the North American Benthological Society 25: 607–615.
Bärlocher, F. & B. Kendrick, 1975. Leaf-conditioning by microorganisms. Oecologia 20: 359–362.
Benfield, E. F., 2011. Decomposition of Leaf Material. In Hauer, F. R. & G. A. Lamberti (eds), Methods in Stream Ecology. Academic Press, California: 711–720.
Boyero, L., R. G. Pearson, D. Dudgeon, M. A. S. Graça, M. O. Gessner, R. J. Albariño, V. Ferreira, C. M. Yule, A. J. Boulton, M. Arunachalam, M. Callisto, E. Chauvet, A. Ramirez, J. Chará, M. S. Moretti, J. F. Gonçalves Jr, J. E. Helson, A. M. Chará-Serna, A. Encalada, J. N. Davies, S. Lamothe, A. Cornejo, J. Castela, A. O. Li, L. M. Buria, V. Díaz Villanueva, M. C. Zúñiga & C. M. Pringle, 2011. Global distribution of a key trophic guild contrasts with common latitudinal diversity patterns. Ecology 92: 1839–1848.
Breshears, D. D., N. S. Cobb, P. M. Rich, K. P. Price, C. D. Allen, R. G. Balice, W. H. Romme, J. H. Kastens, M. L. Floyd & J. Belnap, 2005. Regional vegetation die-off in response to global-change-type drought. Proceedings of the National Academy of Sciences of the United States of America 102: 15144–15148.
Bruder, A., E. Chauvet & M. O. Gessner, 2011. Litter diversity, fungal decomposers and litter decomposition under simulated stream intermittency. Functional Ecology 25: 1269–1277.
Bunn, S. E. & A. H. Arthington, 2002. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environmental Management 30: 492–507.
Buria, L., R. Albarino, V. D. Villanueva, B. Modenutti & E. Balseiro, 2010. Does predation by the introduced rainbow trout cascade down to detritus and algae in a forested small stream in Patagonia? Hydrobiologia 651: 161–172.
Caruso, B., 2002. Temporal and spatial patterns of extreme low flows and effects on stream ecosystems in Otago, New Zealand. Journal of Hydrology 257: 115–133.
Corti, R., S. T. Larned, T. Datry & L. Drummond, 2011. Natural variation in immersion and emersion affects breakdown and invertebrate colonization of leaf litter in a temporary river. Aquatic Sciences 73: 537–550.
Cummins, K. W., M. A. Wilzbach, D. M. Gates, J. B. Perry & W. B. Taliaferro, 1989. Shredders and riparian vegetation. BioScience 39: 24–30.
Cheshire, K., L. Boyero & R. G. Pearson, 2005. Food webs in tropical Australian streams: shredders are not scarce. Freshwater Biology 50: 748–769.
Chessman, B. C., 2015. Relationships between lotic macroinvertebrate traits and responses to extreme drought. Freshwater Biology 60: 50–63.
Dai, A., 2013. Increasing drought under global warming in observations and models. Nature Climate Change 3: 52–58.
Datry, T., R. Corti, C. Claret & M. Philippe, 2011. Flow intermittence controls leaf litter breakdown in a French temporary alluvial river: the “drying memory”. Aquatic Sciences 73: 471–483.
Díaz Villanueva, V., L. Buria & R. Albariño, 2010. Primary consumers and resources: annual variation in two contrasting reaches of a Patagonian mountain stream. International Journal of Limnology 46: 21–28.
Dieter, D., D. von Schiller, E. M. García-Roger, M. M. Sánchez-Montoya, R. Gómez, J. Mora-Gómez, F. Sangiorgio, J. Gelbrecht & K. Tockner, 2011. Preconditioning effects of intermittent stream flow on leaf litter decomposition. Aquatic Sciences 73: 599–609.
Dieterich, M. & N. Anderson, 1995. Life cycles and food habits of mayflies and stoneflies from temporary streams in western Oregon. Freshwater Biology 34: 47–60.
Domínguez, E., H. R. Fernández & F. M. Lillo, 2009. Macroinvertebrados bentónicos sudamericanos: Sistemática y biología. Fundación Miguel Lillo Tucumán, Tucumán.
Ferreira, V. & C. Canhoto, 2014. Effect of experimental and seasonal warming on litter decomposition in a temperate stream. Aquatic Sciences. doi:10.1007/s00027-013-0322-7.
Ferreira, V. & E. Chauvet, 2011a. Future increase in temperature more than decrease in litter quality can affect microbial litter decomposition in streams. Oecologia 167: 279–291.
Ferreira, V. & E. Chauvet, 2011b. Synergistic effects of water temperature and dissolved nutrients on litter decomposition and associated fungi. Global Change Biology 17: 551–564.
Fritz, K. M. & W. K. Dodds, 2004. Resistance and resilience of macroinvertebrate assemblages to drying and flood in a tallgrass prairie stream system. Hydrobiologia 527: 99–112.
García, L., C. Delgado & I. Pardo, 2008. Seasonal changes of benthic communities in a temporary stream of Ibiza (Balearic Islands). Limnetica 27: 259–272.
González, J. M. & M. A. S. Graca, 2003. Conversion of leaf litter to secondary production by a shredding caddis-fly. Freshwater Biology 48: 1578–1592.
Gordon, N. D., T. A. McMahon, B. L. Finlayson, C. J. Gippel & R. J. Nathan, 2004. Stream Hydrology: An Introduction for Ecologists. Wiley, Chichester.
Graça, M. A. & C. Canhoto, 2006. Leaf litter processing in low order streams. Limnetica 25: 1–10.
Graça, M. A., F. Bärlocher & M. O. Gessner, 2005. Methods to Study Litter Decomposition: A Practical Guide. Springer, Berlin.
Kirby, J. M., J. R. Webster & E. F. Benfield, 1983. The Role of Shredders in Detrital Dynamics of Permanent and Temporary Streams. In Bartell, T. F. S. (ed.), Dynamics of Lotic Ecosystems. Ann Arbor Science Publishers, Michigan: 425–435.
Lake, P. S., 2000. Disturbance, patchiness, and diversity in streams. Journal of the North American Benthological Society 19: 573–592.
Leberfinger, K., I. Bohman & J. Herrmann, 2010. Drought impact on stream detritivores: experimental effects on leaf litter breakdown and life cycles. Hydrobiologia 652: 247–254.
López-Rodríguez, M. J., J. M. Tierno de Figueroa, T. Bo, A. Mogni & S. Fenoglio, 2012. Living apart together: on the biology of two sympatric Leuctra species (Plecoptera, Leuctridae) in an Apenninic Stream, Italy. International Review of Hydrobiology 97: 117–123.
Maamri, A., H. Chergui & E. Pattee, 1997. Leaf litter processing in a temporary northeastern Moroccan river. Archiv für Hydrobiologie 140: 513–531.
Merritt, R. W. & K. W. Cummins (eds), 1996. Aquatic insects of North America. Kendall/Hunt, Iowa.
Miserendino, M., 2007. Macroinvertebrate functional organization and water quality in a large arid river from Patagonia (Argentina). International Journal of Limnology 43: 133–145.
Modenutti, B., R. Albariño, M. Bastidas Navarro, V. Díaz Villanueva, M. S. Souza, C. Trochine, C. Laspoumaderes, F. Cuassolo, G. Mariluán, L. Buria & E. Balseiro, 2010. Structure and dynamic of food webs in Andean North Patagonian freshwater systems: organic matter, light and nutrient relationships. Ecologia Austral 20: 95–114.
Nuñez, M. N., S. A. Solman & M. F. Cabré, 2009. Regional climate change experiments over southern South America. II: climate change scenarios in the late twenty-first century. Climate Dynamics 32: 1081–1095.
Otermin, A., A. Basaguren & J. Pozo, 2002. Re-colonization by the macroinvertebrate community after a drought period in a first-order stream (Agüera Basin, Northern Spain). Limnetica 21: 117–128.
Richardson, J. S., 1991. Seasonal food limitation of detritivores in a montane stream: an experimental test. Ecology 72: 873–887.
Richardson, J. S. & R. J. Danehy, 2007. A synthesis of the ecology of headwater streams and their riparian zones in temperate forests. Forest Science 53: 131–147.
Richardson, J. S., T. M. Hoover & A. Lecerf, 2009. Coarse particulate organic matter dynamics in small streams: towards linking function to physical structure. Freshwater Biology 54: 2116–2126.
Richardson, W. B., 1990. A comparison of detritus processing between permanent and intermittent headwater streams. Journal of Freshwater Ecology 5: 341–357.
Robson, B. J., E. T. Chester & C. M. Austin, 2011. Why life history information matters: drought refuges and macroinvertebrate persistence in non-perennial streams subject to a drier climate. Marine and Freshwater Research 62: 801–810.
Treplin, M. & M. Zimmer, 2012. Drowned or dry: a cross-habitat comparison of detrital breakdown processes. Ecosystems 15: 477–491.
Valdovinos, C., 2001. Riparian leaf litter processing by benthic macroinvertebrates in a woodland stream of central Chile. Revista Chilena de Historia Natural 74: 445–453.
Webster, J. R. & E. F. Benfield, 1986. Vascular plant breakdown in freshwater ecosystems. Annual Review of Ecology, Evolution and Systematics 17: 567–594.
Wesner, J. S., 2010. Seasonal variation in the trophic structure of a spatial prey subsidy linking aquatic and terrestrial food webs: adult aquatic insects. Oikos 119: 170–178.
Wood, E., F. Armstrong & F. Richards, 1967. Determination of nitrate in sea water by cadmium-copper reduction to nitrite. Journal of Marine Biological Association 47: 23–31.
Acknowledgments
The final version of the manuscript was highly improved by the critical suggestions from two anonymous reviewers. We thank the Intergovernmental Water Bureau of the Río Negro Basin (AIC-Autoridad Interjurisdiccional de Cuencas de los Ríos Limay, Neuquén y Negro) for providing the precipitation records reported in this study. Special thanks to Nahuel Huapi Park Rangers of Zona Centro for kindly taking us to Chall-Huaco valley in winter fieldtrips. This research was supported by a Grant from the ANCYPT (National Agency of Scientific and Technical Promotion) PICT0793-2011. The first author has a CONICET (National Scientific and Technical Research Council) doctoral fellowship.
Conflict of interest
The authors declare not to have any conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Luz Boyero
Rights and permissions
About this article

Cite this article
Mariluan, G.D., Díaz Villanueva, V. & Albariño, R.J. Leaf litter breakdown and benthic invertebrate colonization affected by seasonal drought in headwater lotic systems of Andean Patagonia. Hydrobiologia 760, 171–187 (2015). https://doi.org/10.1007/s10750-015-2324-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-015-2324-z