
Abstract
The influence of Potamogeton pectinatus colonisation on benthic nitrogen dynamics was studied in the littoral zone of a lowland pit lake with high nitrate concentration (~200 μM). Our hypothesis was that in aquatic environments where nitrogen availability is not limiting, colonisation by rooted macrophytes changes the dynamics of the benthic nitrogen cycle, stimulating N assimilation and denitrification and increasing the system capacity to take up external nitrogen loads. To test this hypothesis, we quantified and compared seasonal variations of light and dark benthic metabolism, dissolved inorganic nitrogen (DIN) fluxes, denitrification and N assimilation rates in an area colonised by P. pectinatus and a reference site colonised by microphytobenthos. In both areas, the benthic system was net autotrophic and a sink for DIN (2,241–2,644 mmol m−2 y−1). Plant colonisation increased nitrogen losses via denitrification by 30% compared to the unvegetated area. In contrast to what is generally observed in coastal marine systems, where the presence of rooted macrophytes limits denitrification rates, under the very high nitrate concentrations in the studied lake, both denitrification (1,237–1,570 mmol m−2 y−1) and N assimilation (1,039–1,095 mmol m−2 y−1) played important and comparable roles in the removal of DIN from the water column.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nitrogen is recognised as a major pollutant in aquatic environments mainly due to sewage disposal, run-off of fertiliser and/or manure applied in areas exploited for intensive agriculture and livestock rearing (Vitousek et al., 1997; Galloway et al., 2003). In addition, widespread reclamation of riparian wetlands originally present along rivers and lakes, and the deterioration of the residual areas contributes to enhanced nitrogen pollution of aquatic ecosystems. In agricultural landscapes, shallow aquatic environments and transitional zones are considered ‘hot spots’ of biogeochemical activity that transform and reduce the N load in the terrestrial to aquatic continuum (Wetzel, 1990; den Heyer & Kalff, 1998; Saunders & Kalff, 2001; David et al., 2006; Seitzinger et al., 2006). This situation can be, at least partially, counteracted by landscape approaches to nitrogen management, through the restoration or creation of ‘filter’ wetland ecosystems. However, this approach is still limited by our current poor understanding of the factors which regulate nitrogen cycling in these environments (Megonigal et al., 2004; Verhoeven et al., 2006; Palmer & Filoso, 2009; Burgin et al., 2012).
In shallow aquatic ecosystems, several biogeochemical processes in the surficial sediment contribute to the transformation and elimination of nitrogen. These processes largely depend on prokaryotic activities: oxidative (mineralisation of organic matter and nitrification) or reductive (denitrification, ANAMMOX and DNRA) pathways, which transform organic and inorganic nitrogen into various inorganic forms (Burgin & Hamilton, 2007). The rooted macrophytes that colonise illuminated sediments in pristine aquatic environments are known to play a role as biogeochemical regulators that influence nitrogen cycling due to their photosynthetic activity and their high nitrogen demand for primary productivity (Reddy et al., 1989; Caffrey & Kemp, 1992; Risgaard-Petersen & Jensen, 1997). Nitrogen assimilation, organic matter enrichment of the sediment and radial oxygen loss by roots are some examples of processes that alter environmental characteristics, with potentially both positive and negative effects on microbially mediated N transformations. Dissolved organic carbon excretion from macrophyte roots for example provides bacteria with energy sources that can fuel nitrogen fixation and/or denitrification, while radial oxygen loss in the rhizosphere can provide O2 to aerobic nitrifiers that in turn generates nitrate for denitrification, DNRA and ANAMMOX (Welsh et al., 1996; Risgaard-Petersen & Jensen, 1997; Ottosen et al., 1999; Welsh, 2000; Karjalainen et al., 2001; Nielsen et al., 2001). In addition, plant leaves and stems increase the colonisable surface available to periphyton communities that also sustain high nitrogen transformation rates (Eriksson & Weisner, 1999; Toet et al., 2003). Conversely, N assimilation by the plant leaves and roots regulates dissolved inorganic nitrogen (DIN) availability in the water and sediment, and competes with nitrifiers and denitrifiers for DIN species (Caffrey & Kemp, 1992; Welsh et al., 2000; Risgaard-Petersen, 2004; Bartoli et al., 2008; Racchetti et al., 2010; Veraart & de Bruijne, 2011). In diffusion dominated systems like aquatic sediments, interactions between benthic macrophytes and bacterial processes are complex and influence the function of the so called ‘benthic filter’, and ultimately the role of these aquatic ecosystems as nitrogen sinks (Seitzinger et al., 2006). In fact, depending on the relative importance of the abovementioned processes, mineralised nitrogen may exit the sediment as DIN (NH4 + + NO2 − + NO3 −), which is still available to primary producers, or as nitrogen gas (N2) which is permanently lost from the system, or be temporally assimilated into plant biomass. Thus, with respect to excess nitrogen removal, the balance between assimilatory and dissimilatory pathways is a key feature.
The majority of the studies on the influence of aquatic macrophytes on N dynamics have focused on coastal marine ecosystems (Nielsen et al., 2004 and references therein). In these environments, where nitrogen availability generally limits primary productivity, the presence of benthic primary producers depresses nitrification and denitrification and plant assimilation dominates benthic nitrogen transformations (Ottosen et al., 1999; Welsh et al., 2000; Risgaard-Petersen, 2004). Under these circumstances, the benthic filter is mainly due to temporal nitrogen storage in plant biomass and eventually the export or permanent burial of refractory organic nitrogen (Nielsen et al., 2004 and references therein). In contrast, the relatively few studies of shallow freshwater systems show variable effects dependent upon the vegetation type and environmental conditions (Veraart & de Bruijne, 2011). For example direct comparison studies have failed to demonstrate differences in denitrification rates between vegetated and unvegetated sediments in lakes (Saunders & Kalff, 2001), wetlands (Toet et al., 2003) and laboratory microcosms (Veraart & de Bruijne, 2011) or report a positive effect of different aquatic plants on both nitrification and denitrification (Christensen & Sørensen, 1986; Caffrey & Kemp, 1992; Risgaard-Petersen & Jensen, 1997; Ottosen et al., 1999; Pinardi et al., 2009). In addition, a recent attempt to summarise the results of different studies failed to demonstrate a significant effect of aquatic plants on denitrification rates, although there was a tendency towards lower rates in the presence of plants under illuminated conditions (Piña-Ochoa & Álvarez-Cobelas, 2006).
Thus, the net effect of rooted macrophytes on denitrification and on the relative contribution of heterotrophic and autotrophic processes in shallow freshwater environments remains unclear. However, an understanding of how ecosystem modification alters the benthic nitrogen cycle is essential information for the recovery of aquatic ecosystems and related ecological functions in agricultural landscapes (McClain et al., 2003; Burgin et al., 2012). These ecosystems are subject to high and often increasing inputs of nitrate, which would be expected to favour nitrate removal via denitrification (David et al., 2006; Nizzoli et al., 2010), by reducing competition between plants and microbes. Under these conditions, we hypothesise that the presence of rooted macrophytes would change the dynamics of the nitrogen cycle and enhance denitrification rates compared to uncolonised sites, and ultimately improve the capacity of the system to eliminate external nitrogen loads.
In order to evaluate this hypothesis we quantified and compared seasonal variations of benthic metabolism, DIN fluxes, denitrification and nitrogen assimilation rates in a pit lake, at a site undergoing colonisation by the rooted macrophyte Potamogeton pectinatus and at an unvegetated reference site, under light and dark conditions. Pit lakes are becoming a common feature in agricultural floodplains and offer the possibility to analyse the effect of rooted macrophytes on N dynamics during sediment colonisation.
Materials and methods
Study site
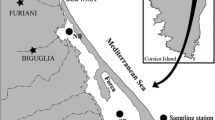
The study was carried out in a pit lake, formed by sand and gravel extraction, located in the Po river floodplain near the city of Piacenza, Northern Italy (Fig. 1), approximately 2 km from the main course of the river. The lake has a maximum depth of 20 m and the littoral area (defined as the surface with a maximum depth of 3 m) accounts for ~10% of the lake surface. Sand extraction at the site ceased in 2004 and presently the littoral zone is undergoing natural colonisation by both riparian and aquatic vegetation, especially P. pectinatus. The lake has no discrete inlets or outlets and water inputs are from groundwater and overland run-off; the surrounding area is mainly used for livestock farming and agriculture. Average DIN concentrations are high (up to 200 μM) but the lake can be regarded as oligo-mesotrophic with respect to total phosphorus (<0.3 μM) and chlorophyll-a (Chl-a) concentrations (1–14 μg l−1). The water is rich in dissolved ions (conductivity 550 μS cm−1, 25°C) with an average dissolved ionic composition dominated by Ca2+, Mg2+, HCO3 − and SO4 2− that account for 38 and 53%, and 60 and 22% of total cations and anions, respectively (Tavernini et al., 2009; Nizzoli et al., 2010). Sampling was conducted at two sites in the littoral area of the lake with similar water depth (Fig. 1): site P was located within the area colonised by P. pectinatus, while site S was located in an adjacent (approximately 20 m away) uncolonised area.

Map showing the location of the studied lake in Northern Italy and the main morphometric and chemical features of the lake. The shaded area represents the zone colonised by P. pectinatus, and the letters P and S indicate the locations of the P. pectinatus and bare sediment sampling sites, respectively
Sediment collection and experimental set-up
Bare (site S) and P. pectinatus colonised (site P) sediments were sampled approximately bimonthly in March, May, July, October and December 2007. On each occasion, three sediment cores (inner diameter 5 cm, height 30 cm) were collected for sediment characterisation, six sediment cores (inner diameter 8 cm, height 40 cm) for flux and denitrification measurements in bare sediments and six sediment cores (inner diameter 20 cm, height 40 cm) for flux and denitrification measurements in P. pectinatus colonised sediment. All cores were collected by hand using Plexiglass core tubes at a water depth of approximately 1.5 m and the height of the sediment inside cores was adjusted to approx. 15 cm. Cores were returned to the laboratory within 3 h for further processing and incubation. Approximately 200 l of site water was also collected for core maintenance and incubation in the laboratory.
Water temperature and dissolved oxygen were determined in situ with a multiparameter probe (YSI 556) and 50 ml water was filtered (Whatman GF/F) for DIN determinations.
In the laboratory, cores for sediment characterisation were extruded and the upper 0–1 cm horizon retained for porosity, density, organic matter and organic carbon and nitrogen determinations. Microphytobenthos (MPB) biomass at site S was also determined as Chl-a concentration after extraction with 90% acetone according to Lorenzen (1967). Sediment density, porosity and organic matter content as loss on ignition at 550°C (LOI) were determined as described in Nizzoli et al. (2005). Dry sediments were also analysed for organic carbon and nitrogen content using a CHN elemental analyser (CHNS-O EA 1108 Carlo Erba).
Determination of sediment–water oxygen and DIN fluxes and denitrification rates
In the laboratory, the cores were submersed with the top open in a tank containing in situ aerated water for 24 h at in situ temperature, under a light/dark cycle close to that of the sampling period. Prior to incubations, the water within the cores was exchanged with the water in the incubation tank. Water mixing inside the cores was ensured by small aquarium pumps (flow ~ 2.5 l min−1) in the vegetated cores and by magnetic bars driven by an external motor at 60 rpm in bare sediment cores. Sets of three cores per site were incubated to determine oxygen and DIN fluxes under light and dark conditions, as described in Bartoli et al. (2008). All incubations were conducted at in situ oxygen and nutrient concentrations, in a constant temperate room at in situ temperature (±1°C). Light incubations were performed at the average irradiance of the sampling period. Incubation times ranged from 1.5 to 5 h depending upon season in order to keep oxygen concentration at the end of the incubation within 20% of the initial value. Water samples were collected from cores at the beginning and at the end of the incubations using plastic syringes as preliminary tests demonstrated that changes in solute concentrations where linear over time. Samples for O2 determinations were transferred to glass vials (Exetainers, Labco, High Wycombe, UK) and fixed with Winkler reagents (APHA, 1998). Samples for NH4 +, and NO2 − + NO3 − (hereafter referred to as NO x ) determinations were filtered through Whatman GF/F glass fibre filters, transferred to plastic vials, stored at 4°C and analysed within 24 h.
Following flux incubations, the incubation tanks were topped up with site water and the cores were left uncapped and submerged in the tanks for 2 h to equilibrate prior to determination of denitrification rates by the isotope pairing technique (IPT; Nielsen, 1992). To initiate incubations, the water in the tanks was lowered to below the core tops and a 15NO3 − solution (98 at.%, Sigma-Aldrich) was added to give a final concentration of 50–100 µM (20–60% enrichment). The actual 15N addition to each core was calculated by difference from water samples collected before and 10 min after the 15N addition. Following the addition of labelled nitrate cores were pre-incubated from 15 min (summer) to 30 min (winter) before the start of the incubation to allow the tracer to diffuse to the sediment nitrate reduction zone (Nielsen, 1992) and were then closed using floating plexiglass lids. Incubation times were set to ensure that oxygen consumption was less than 20% of the initial O2 concentration, which is a prerequisite for the isotope pairing method (Nielsen, 1992).
At the end of the incubations, 7 M ZnCl2 was added at each core to a final concentration of 10 mM, in order to prevent further denitrification, and dissolved N2 pools in the water column and sediment porewater were homogenised by gently stirring the sediment with a metal bar. Subsamples of the final slurry were transferred to 12-ml Exetainer vials (Exetainer, Labco, High Wycombe, UK), fixed with 150 μl 7 M ZnCl2 and stored at 4°C until analysed.
Thereafter, the contents of the vegetated cores were sieved with a 1 mm mesh and the collected material sorted into living leaves, roots + rhizomes and detritus (dead leaves, roots and rhizomes). Biomass was rinsed to remove epiphytes and dried at 70°C until constant weight. Dried biomass was powdered and analysed for C and N content using a CHNS-O EA 1108 Elemental Analyzer (Carlo Erba).
Calculation of flux and denitrification rates and theoretical nitrogen demand
Oxygen and DIN fluxes were calculated from the difference between final and initial concentration of the target compound in the water column according to Eq. (1):
F x is the flux of the x species (μmol m−2 h−1), C f is the final concentration of x (μmol l−1), C i is the initial concentration of x (μmol l−1), V is the volume of the water column (l), t is the incubation time (h) and A is the sediment surface area inside the core (m2).
Total denitrification rates (DE), denitrification of nitrate diffusing to the sediment from the water column (D W) and denitrification of nitrate produced within the sediment by nitrification (D N) were calculated following the equations proposed by Nielsen (1992). Independence of denitrification rates from added 15NO3 − was also checked to exclude significant overestimation due to ANAMMOX (Risgaard-Petersen et al., 2003). D N rates measured at the site colonised by P. pectinatus should be regarded as minimum rates, as due to root oxygen release, this site fails to meet the criteria for accurate determination of coupled nitrification–denitrification using the IPT (Nielsen, 1992; see ‘Discussion’ for details).
Net daily sediment–water column fluxes and denitrification rates were calculated as the sum of the light and dark rates multiplied by the average light and dark periods at the time of sampling. Annual rates were estimated by integrating daily rates over the sampling year assuming a constant change between two subsequent sampling periods using the following equation:
AR is the annual rate (mmol m−2 y−1), DR n is the daily rates calculated for sampling day n and for the following sampling day (n + 1) and Δdays is the number of days between sampling days n and n + 1.
The theoretical nitrogen demands for growth of P. pectinatus and MPB were calculated from daily net benthic production (NBP), which was calculated as the sum of the light and dark rates multiplied by the average light and dark periods at the time of sampling. Daily NBP was then converted into carbon equivalents assuming a photosynthetic quotient of 1.2 (Schramm et al., 1984; Wetzel & Likens, 1991). Nitrogen demands to support this production were calculated from the measured C:N ratios of leaves for P. pectinatus and assuming a molar C:N ratio of 9 for MPB (Sundbäck & Miles, 2000). Daily rates were then scaled up to annual assimilation as previously described for benthic fluxes. The standing stock of MPB as C m−2 was calculated using a ratio of 30:1 C:Chl-a (De Jonge, 1980) and converted to N m−2 assuming a molar C:N ratio of 9 (Sundbäck & Miles, 2000).
Analytical methods
Dissolved oxygen was determined by the Winkler method (APHA, 1998); ammonium concentrations were determined by the indophenol blue method according to Bower & Holm-Hansen (1980). NO x was determined by diazotation, after reduction to nitrite over cadmium columns (APHA, 1998). The abundance of 29N2 and 30N2 in the dissolved N2 pool was determined by mass spectrometry at the National Environmental Research Agency, Silkeborg, Denmark, as previously described by Risgaard-Petersen & Rysgaard (1995).
Statistical methods
Normality and homoscedasticity of data were examined and when required data were square root transformed and retested. Flux data and denitrification rates were analysed with a three-way ANOVA (site, light and month as fixed variables; interaction included), with an a posteriori comparison of the means performed using a post-hoc SNK test (Sokal and Rohlf, 1995).
Results
Water and sediment characteristics, primary producer biomass and elemental composition
Water physical and chemical characteristics were similar at both sampling sites (Table 1). Temperature ranged between 9°C (December) and 27°C (July). Water column NH4 + concentrations varied seasonally between 16 μM in winter and 2 μM in summer, whereas NO x concentrations were consistently high (153–182 μM) throughout the year. The sediment at site S was relatively homogeneous and mainly composed of sand and silt alternated by layers of clay with a low average organic matter (LOI) content of 3.0 ± 2.2% (Table 1). The sediment surface was colonised by MPB, with Chl-a concentrations comprised between 48 ± 25 (March) mg m−2 and 73 ± 15 (July) mg m−2 (Table 1). The sediment at the P. pectinatus colonised site P was more heterogeneous and contained a significant amount of partially decomposed plant detritus (20–158 gdw m−2; Table 2), with the sediment LOI and Org-C content in surficial sediment being significantly higher (ANOVA, P < 0.01) on four of five occasions compared to that of site S (Table 1).
The P. pectinatus stand extended from 0.5 m to about 2 m depth. Aboveground biomass varied seasonally with the lowest values (38 ± 19 gdw m−2) measured in March, when coverage was minimum and the bed was constituted by a few sparse individuals, and the highest in July (156 ± 59 gdw m−2). Thereafter, there was a steady decline in biomass, down to 65 ± 35 gdw m−2 in December (Table 2). Belowground biomass increased from March (20 ± 12 gdw m−2) to October (40 ± 13 gdw m−2) with above- to belowground biomass ratios minimum in winter (~1.6–2.0) and maximum in summer (~4.4). The C content of the above- and belowground biomass was similar (35%) and relatively constant between seasons (Table 2). By contrast, the nitrogen content was consistently higher in above (annual mean 2.3%) compared to below ground (annual mean 1.6%) biomass. The N contents of both above- and belowground biomass showed clear seasonal trends with maximum values measured in winter and minimal values in summer (Table 2). Consequently, the C to N ratios of both the above- and belowground biomass were also the highest in summer (25 and 87, respectively) and the lowest in winter (15 and 21, respectively).
The average nitrogen content in the surficial sediment was similar at both sites (Tables 1, 3). The estimated N pool in MPB biomass was relatively low (13–20 mmol N m−2) compared to that in the P. pectinatus biomass (108–191 mmol N m−2) (Fig. 2a; Table 3).

Seasonal evolution of a N standing stocks, b N retention (turnover) time in the primary producer biomass, c estimated net community productivity and d theoretical community N assimilation at the P. pectinatus (black symbols and bars) and MPB (white symbols and bars) colonised areas. Data are reported as mean ± standard deviation (nitrogen pools n = 6; net community productivity, theoretical nitrogen uptake and N retention time n = 3)
Oxygen fluxes, primary productivity and related theoretical nitrogen demand
Oxygen fluxes differed significantly between light and dark conditions (ANOVA, P < 0.001) at both sampling sites (Table 4). During light incubations, net oxygen production was measured in both vegetated and bare benthic systems on all the sampling dates (Fig. 3a, b), but was significantly higher (ANOVA, P < 0.01; Table 4) at the P. pectinatus colonised site (annual mean 5.8 ± 3.0 vs 2.1 ± 2.0 mmol m−2 h−1). Net oxygen production at this site showed a clear seasonal trend with the highest rates measured in May (9.6 ± 3.7 mmol m−2 h−1) and the lowest in October (3.7 ± 0.9 mmol m−2 h−1) (Fig. 3a). A similar pattern was observed in the MPB colonised sediments at site S where the highest net O2 production occurred in July (4.6 ± 2.3 mmol m−2 h−1) (Fig. 3b).

Seasonal evolution of benthic light and dark oxygen fluxes (O2, upper panels) and gross primary production (GPP, black lines upper panels), and denitrification rates (DE, lower panels) measured at the P. pectinatus (a, c) and MPB (b, d) colonised areas. In denitrification panels, the hatched portion of the bars represents denitrification coupled to nitrification (D N) and the unhatched portion denitrification of nitrate diffusing from the water column (D W). All data are reported as mean ± standard deviations (n = 3)
Benthic dark oxygen consumption was also significantly higher at site P (range −1.5 ± 0.3 to −7.0 ± 3.4 mmol m−2 h−1) compared to site S where respiration varied between −0.8 ± 0.2 and −2.5 ± 0.2 mmol O2 m−2 h−1 (ANOVA, P < 0.01; Table 4). At both sites, the lowest dark oxygen demands were measured in March and the highest in July (Fig. 3a, b). Gross community primary production (dark respiration + light production) was 1.6–7 times higher at site P compared to site S and peaked in May (15.5 ± 3.8 mmol O2 m−2 h−1) at site P and in July (7.0 ± 2.3 mmol O2 m−2 h−1) at site S (Fig. 3a, b).
Mean daily net C fixation rates (Fig. 2c) calculated by multiplying net oxygen production rates by the average monthly hours of insolation per day and converting to C using a photosynthetic quotient of 1.2 (Schramm et al., 1984; Wetzel & Likens, 1991) were 103 ± 10 mmol C m−2 d−1 (range 34 ± 8–112 ± 43 mmol C m−2 d−1) at the P. pectinatus colonised site and 41 ± 5 mmol C m−2 d−1 at the MPB colonised site (range 10 ± 6 to 57 ± 29 mmol C m−2 d−1). Annual net community production (Table 3) was positive at both sites but ~2 times higher at site P (22,218 ± 5,845 mmol m−2) compared to site S (9,355 ± 3,655 mmol m−2).
The theoretical N demand to support primary production at the vegetated site, based on net C fixation and the measured C:N ratios in the aboveground biomass, ranged from 1.8 ± 0.4 to 4.5 ± 1.7 mmol m−2 d−1 (average 3.0 mmol m−2 d−1) (Fig. 2d). The highest values were estimated for May and the lowest in October, with the net annual N demand estimated to be 1,095 ± 282 mmol m−2 (Table 3). On average these rates were only slightly higher compared to the estimated N demand of MPB which ranged from 1.1 ± 0.6 to 6.4 ± 3.2 mmol m−2 d−1 (average 2.7 mmol m−2 d−1). As rates of N assimilation by MPB are based on net primary productivity and a fixed C:N = 9, they show the same seasonal pattern as net productivity (Fig. 2c) and account for an estimated annual N assimilation of 1,039 ± 406 mmol m−2 (Table 3).
Denitrification rates and dissolved inorganic nitrogen fluxes
At both sites, total denitrification (DE) followed a significant seasonal pattern (ANOVA, P < 0.001; Table 4) with the highest rates measured in July (Fig. 3c, d). On average daily DE was 1.3 times higher at site P (4,427 ± 588 μmol m−2 d−1) compared to site S (3,474 ± 513 μmol m−2 d−1). The significant interaction between light and site (ANOVA site * light, P < 0.001; Table 4) indicates that the observed difference was largely due to the different regulation of denitrification by light environment at the two sites. At the MPB colonised site S (Fig. 3d), average denitrification rates measured in the light were ~50% lower than rates measured during dark incubations (annual mean 103 ± 18 μmol m−2 h−1 vs 200 ± 38 μmol m−2 h−1). By contrast, at site P light and dark rates of denitrification were not statistically different (ANOVA, P > 0.05). Thus, although dark rates of denitrification were similar at both sites on all sampling occasions, light rates were on average 70% higher at site P compared to site S (Fig. 3c, d). Overall measured rates, adjusted for the photoperiod on each sampling occasion, equate to net annual nitrogen removal rates via denitrification of 1,237 ± 65 and 1,570 ± 84 mmol m−2 y−1 at sites S and P, respectively (Table 3).
Denitrification was mainly fuelled by nitrate diffusing from the water column (D W) which accounted for 90 and 86% of total denitrification at sites P and S, respectively. D W rates varied between 2 and 497 μmol m−2 h−1 and showed the same seasonal patterns and light/dark shifts as DE. Average daily D W rates were higher at site P (4,013 ± 614 μmol m−2 d−1) compared to site S (3,069 ± 491 μmol m−2 d−1) with this difference again being primarily due to differences in the light measured rates (Table 4; Fig. 3c, d).
Nitrification was only a minor source of nitrate for denitrification (D N) with rates ranging from 3 to 30 μmol m−2 h−1. Average D N was higher under dark (20 ± 21 μmol m−2 h−1) compared to light (13 ± 16 μmol m−2 h−1) (ANOVA, P > 0.001), although D N showed seasonal variations at both sites, the differences between vegetated and bare sediments did not follow any clear pattern. In the MPB colonised sediment of site S, coupled denitrification rates increased from spring to autumn, whereas at site P minimum rates were measured in summer and the highest rates in spring and autumn (Fig. 3c, d).
At both sampling sites on all sampling dates, the benthic compartment was a net sink for DIN during both light and dark incubations (Fig. 4a, b) with annual DIN consumptions of 2,644 ± 366 and 2,241 ± 212 mmol N m−2 y−1 measured at sites P and S, respectively. DIN fluxes were mainly driven by NO x consumption (−10 to −611 μmol m−2 h−1) reflecting the relative abundances of NH4 + and NO x in the water column (Table 1). NO x fluxes were similar at both sampling sites and showed significant seasonal trends (ANOVA, P < 0.01; Table 4) with the highest uptakes recorded in May and July (−460 and −418 μmol m−2 h−1 in light and dark, respectively) and the lowest in December (Fig. 4c, d).

Seasonal evolution of light and dark benthic fluxes of dissolved inorganic nitrogen (DIN; a, b), NO x (c, d) and NH4 + (e, f) measured at the P. pectinatus and MPB colonised sites determined during light (white bars) and dark (black bars) incubations. Data are reported as mean ± standard deviations (n = 3)
Ammonium fluxes (Fig. 4e, f) were of quantitatively minor importance compared to those of NO x , but were significantly influenced by light environment at both sites (ANOVA, P < 0.001; Table 4). Generally, NH4 + consumption was measured under illuminated conditions, especially at site P, whilst reduced consumption or small effluxes were measured in the dark at both sites, although both light and dark fluxes were significantly greater at site P compared to site S (annual average −363 ± 100 vs −138 ± 52 mmol m−2 y−1 at sites P and S, respectively) (ANOVA, P < 0.01; Table 4).
Discussion
Benthic oxygen metabolism and nitrogen fluxes
Despite the recent origin of the studied pit lake, the littoral sediments were metabolically highly active under both light and dark conditions. The relatively high surface sediment concentrations of Chl-a at site S indicate that the sediment was heavily colonised by a community of MPB. Chl-a concentrations were similar on all the sampling dates suggesting that this MPB community was present throughout the year at a relatively stable biomass as has been observed in other shallow fresh and brackish water aquatic environments (Sundbäck & Miles, 2000; Wetzel, 2001). P. pectinatus, a common angiosperm in hardwater environments (Vestergaard & Sand-Jensen, 2000), is actively colonising these sediments. In the meadow, P. pectinatus biomasses were comparable with values reported in the literature (van Wijk, 1988) and exhibited seasonal variations and an active growth phase from March to July, consistent with the data reported by Kantrud (1990). Colonisation by P. pectinatus has had a marked effect on benthic features increasing sediment heterogeneity, organic matter content, metabolism and nitrogen cycling.
Based on the measured light and dark benthic oxygen fluxes, both the P. pectinatus and MPB colonised sites would be classified as net to totally autotrophic systems on all the sampling dates (Viaroli & Christian, 2003). Benthic metabolism followed a clear seasonal pattern in relation to temperature and primary producer biomass, with the highest rates of both dark respiration and gross and net oxygen production in the light measured in summer. However, colonisation by P. pectinatus increased benthic autotrophy with estimated gross oxygen production at site P being on average ~2.8 times higher compared to site S.
The gross community primary production to respiration ratio above 1 indicates that during the sampling period, the produced biomass was not completely remineralised at the site of production. Part of the annual production may have been accumulated as living biomass, buried in the sediment or exported from the site of production by wind action or transferred to higher trophic levels. Our data did not allow to discriminate the fate of the produced biomass, however, at the vegetated site on average, 40% of the total C stored in biomass (leaves + roots + detritus) was present as detritus in the sediment suggesting that burial may be an important sink for the organic matter produced at this site. In addition, transfer to higher trophic levels may also be an important pathway for P. pectinatus biomass control and export from the system (van Wijk, 1988; Søndergaard et al., 1996).
The theoretical annual nitrogen assimilation required to support net primary production at the P. pectinatus colonised site was similar to that calculated for the MPB colonised area despite the large difference measured in primary production between the two sites. However, the calculated assimilation by P. pectinatus should be considered to represent a minimum value as it likely underestimates the true value due to the fact that it is calculated from community light and dark oxygen fluxes and the N content of the P. pectinatus biomass. First, this method would underestimate actual primary production as any photosynthetically generated oxygen, which was lost to the sediment via the roots, would not be measured using our flux method and therefore not accounted for in the estimates of primary production. Second, our measurements of oxygen fluxes actually measure community metabolism not just that of P. pectinatus and therefore include the production of both MPB and epiphytes which have higher N contents, lower C:N ratios and therefore higher specific N demands than P. pectinatus. Previous studies suggest that radial oxygen release by P. pectinatus roots accounts for only ~4% of total oxygen production (Sand-Jensen et al., 1982) and therefore any error in our calculations due to this effect would be small compared to other aquatic plants which have higher rates of radial oxygen loss from their root systems (Pinardi et al., 2009). Our calculations did not explicitly consider the proportions of community production which were due to the epiphytes and benthic microalgae. Previous studies have shown that epiphyte to macrophyte productivity varies greatly depending on plant morphology, light and nutrient availability and wind action, and can represent from 1 to 50% of total production (Cattaneo & Kalff, 1980; Nõges et al., 2009). Consequently, if epiphytes and MPB had contributed a substantial proportion of total community productivity, our estimates would significantly underestimate N assimilation at the P. pectinatus colonised site, as epiphyte and MPB biomass have much lower C:N ratios and therefore a much higher N demand per unit of growth than P. pectinatus. However, visual observations during sampling and incubation suggest a low epiphyte load at our site and, despite these potential sources of uncertainty, our estimates of the N demand for growth of P. pectinatus of between 0.02 and 0.09 mmol N gdw d−1 agree with previously reported N assimilation rates for macrophytes in autotrophic shallow aquatic environments (Caffrey & Kemp, 1992; Risgaard-Petersen et al., 1998; Hansen et al., 2000; Pinardi et al., 2009; Kreiling et al., 2011). In contrast, the difference in the N content and C:N ratio between P. pectinatus and MPB biomass is the most probable explanation for the similar calculated theoretical N assimilation at the two sampling sites, despite the large differences in the rates of primary production measured. As the much lower C:N ratios of the microalgae and cyanobacteria that comprise the microphytobenthic community means that the N demand for growth per unit of carbon fixed is very much greater than that of macrophytes.
Despite the similar N assimilation at both study sites, P. pectinatus colonisation greatly influences N turnover. At the P. pectinatus colonised site, the standing stock of N represented by the living and dead (detrital) macrophyte biomass was almost 20 times greater than that calculated for the MPB biomass at site S and therefore, both the living and dead plant biomass represent important nitrogen reservoirs at this site. The annual average N retained in P. pectinatus biomass was 159 mmol m−2 and was approximately 10 times greater than that retained in MPB biomass at site S. Indeed, the biomass of both living plants and detritus, each accounted for ~15% of the total N pool at site P. Consequently, the presence of P. pectinatus increased the average retention time of N bound within the plant biomass, calculated as the ratio between N standing stock in the biomass and theoretical N assimilation (Sundbäck & Miles, 2000), to 53 days (Fig. 2b). This value is almost 9 times greater than the mean retention time estimated in the same manner for MPB biomass at site S (6 days). Thus, the continued colonisation of the littoral zone of this recently formed pit lake by P. pectinatus would be expected to significantly reduce nitrogen mobility within the system.
The high nitrogen demand for P. pectinatus and MPB production also influenced sediment–water column DIN fluxes and the benthic compartment was a net sink for DIN on all sampling occasions. Assimilation of this DIN by primary producers played an important role in regulating NH4 + fluxes across the sediment–water interface. During light incubations, NH4 + was consistently consumed by the benthic system, while in dark incubations NH4 + was generally released to the water column. However, at both sites DIN influxes were mainly sustained by NO x consumption as a consequence of its very much higher concentration in the water column compared to NH4 +. NO x fluxes were strongly directed towards the sediment under both light and dark conditions, and this and the fact that the estimated N demand to sustain primary production accounted for on average only 50–60% of benthic DIN consumption, suggests not only that nitrogen availability was not limiting for primary productivity, but that other factors also influenced DIN and particularly NO x fluxes.
Denitrification
In autotrophic shallow coastal marine aquatic environments colonised by benthic macrophytes or MPB, denitrification rates are generally low, especially in macrophyte colonised sediments (Table 5) because of competition between the primary producers, and nitrifying and denitrifying bacteria for DIN, and increased oxygen availability due to photosynthesis (Sundbäck & Miles, 2000; Welsh et al., 2000; Risgaard-Petersen, 2003), although this is not always the case (Dunn et al., 2012 and references therein). In contrast, denitrification rates measured in freshwater sediments including those measured in this study are on average approximately an order of magnitude higher than those measured in coastal ecosystems and there was little difference between MPB and macrophyte colonised sediments (Table 5). The overall annual denitrification rates of 1,570 and 1,237 mmol m−2 y−1 measured at sites P and S, respectively, fall within the upper range of values (100–3,700 mmol m−2 y−1) measured in other freshwater environments (Piña-Ochoa & Álvarez-Cobelas, 2006; Seitzinger et al., 2006), reflecting the consistently high nitrate availability in the water column throughout the year. These data demonstrate that shallow aquatic environments characterised by high NO3 − loads can sustain high denitrification rates, despite their highly autotrophic status and thereby ameliorate the impacts of external nitrate loads.
Colonisation of the littoral areas of the lake by P. pectinatus increased denitrification rates compared to MPB colonised areas and thus enhanced nitrogen elimination from the lake. The observed difference in denitrification rates between the P. pectinatus and MPB colonised areas was largely due to decreased denitrification rates at the MPB site during light incubations and can largely be explained by the mode by which the primary producers influence the diffusion of water column nitrate to the sediment denitrification zone, as direct denitrification (D W) was by far the dominant component of total denitrification. At site S the benthic microalgae inhibited D W rates measured in the light. Whilst this could be due to the competition between MPB assimilation and denitrifying bacteria for nitrate in the light, this is unlikely given the high availability of DIN in the studied system and it is probable that this inhibition was an indirect effect of photosynthesis due to oxygen evolution. MPB photosynthesis is known to increase the thickness of the surface oxic layer of the sediment (Revsbech et al., 2005). This augments the mean diffusional path length of NO3 − to the sediment denitrification zone, which occurs just below the sediment oxic–anoxic boundary and thereby reduces the diffusion rate of NO3 − from the water column to the denitrification zone to fuel D W (Risgaard-Petersen et al., 1994; Rysgaard et al., 1995; Dunn et al., 2012). Conversely, during dark incubations in the absence of photosynthetic oxygen production, the surface oxic layer of sediment is thinner, the diffusional path length for water column NO3 − is shorter and increased rates of NO3 − diffusion to the sediment increase its availability and therefore rates of D W.
In contrast, at the site colonised by P. pectinatus rates of D W were similar during both light and dark incubations, indicating that the photosynthetic activity of the plants did not directly influence the supply of NO3 − from the water column or its availability to denitrifiers in the sediment. As discussed earlier, it has been estimated that P. pectinatus releases only ~4% of the oxygen produced by photosynthesis to the sediment (Sand-Jensen et al., 1982) and much of this oxygen may be released to the deeper sediments where most of the roots occur, rather than the near surface sediments to which water column NO3 − diffuses and is consumed by denitrification. Thus, P. pectinatus photosynthesis may have little or no effect on the thickness of the surface oxic layer of sediment and therefore diffusion of NO3 − from the water column to underlying anoxic denitrification zone, resulting in nitrate dynamics and denitrification rates being similar under both light and dark conditions.
Nitrification was only a minor source of nitrate for denitrification (D N) at both sampling sites, although this was mainly due to the high rates of D W, rather than low rates of D N per se, as actual rates were within the typical range for freshwater sediments (Canfield et al., 2005). Previous studies of MPB colonised sediments have typically found that despite the increased availability of oxygen under illuminated conditions, rates of nitrification and therefore D N are typically inhibited due to competition between nitrifiers and MPB for NH4 + (Risgaard-Petersen, 2003). However, more rarely in ammonium-rich sediments, oxygen rather than ammonium is the limiting factor for nitrification and photosynthetic oxygen evolution has been found to stimulate nitrification rates and as a consequence D N (Dunn et al., 2012 and references therein). Our results at site S showed little effect of light environment on rates of D N and rates were similar under light and dark conditions, although the tendency for somewhat higher rates under dark conditions indicates that competition for NH4 + rather than oxygen may be the more important factor regulating rates of nitrification and D N.
The net effect of rooted macrophytes on D N rates is still contradictory. Several studies have reported higher rates of coupled nitrification–denitrification in freshwater sediments colonised by rooted macrophytes due to stimulation of nitrification in oxic microniches in the rhizosphere as a consequence of radial oxygen release from the plant roots (Christensen & Sørensen, 1986; Caffrey & Kemp, 1992; Risgaard-Petersen & Jensen, 1997); others report little or no effect (Saunders & Kalff, 2001; Toet et al., 2003; Veraart & de Bruijne, 2011). Our data suggest that D N rates were not influenced by either the presence of P. pectinatus or its photosynthetic activity as rates were highly similar between sites S and P, and under both light and dark conditions in the P. pectinatus colonised sediments. However, this absence of stimulation of D N rates by P. pectinatus may be at least partially explained by the method used to determine denitrification rates in this study. The IPT assumes uniform mixing by diffusion of the labelled NO3 − added to the water column with NO3 − produced in the sediment by nitrification, as D N is only measured if part of the 14NO3 − produced by nitrification is ‘captured’ during denitrification by the labelled 15NO3 − in the measured 29N2 species (Nielsen, 1992). This assumption is not met in the rhizosphere of rooted aquatic plants as nitrate production by nitrification can occur in oxic microzones around individual roots that are diffusionally isolated from the labelled 15NO3 − added to the water column (Risgaard-Petersen & Jensen, 1997; Welsh et al., 2000). Therefore, our experimental approach could have underestimated rates of D N at site P, as essentially it would only measure D N occurring in the near surface sediments that are accessible to the added label and not D N occurring deeper in the rhizosphere in zones diffusionally isolated from the added 15NO3 −. However, the degree of this potential underestimation would depend largely on the rates and distribution of nitrification and D N occurring in the rhizosphere. In our P. pectinatus colonised sediments this underestimation may in fact be relatively small, since as previously discussed, relatively little of the oxygen produced by P. pectinatus is released from the roots to the rhizosphere sediment (Sand-Jensen et al., 1982) and a previous study found that rates of D N in the rhizosphere of P. pectinatus were low compared to other aquatic plants (Ottosen et al., 1999). However, although we cannot exclude that our measured rates of D N somewhat underestimate the actual rates, this would be only a minor influence on the measured overall rates of denitrification, as nitrification was only a minor NO3 − source for denitrification in this NO3 −-rich lake. At both the MPB and P. pectinatus colonised sites, DE was mainly sustained by NO3 − diffusing from the overlying water column (D W). Such a high contribution of D W to DE is in agreement with the general model relating water column nitrate availability to the relative contributions of D W and D N to overall denitrification rates (Seitzinger et al., 2006). According to this model, D W would account for more than the 80% of DE when water column NO x concentrations exceed 100 μM, which is consistent with the 70% to almost 100% contributions of D W to DE that we measured in the P. pectinatus colonised sediments. Even if we considered a 30% underestimation of D N at the vegetated site (Ottosen et al., 1999), this would only increase the annual average rate of D N from 17 ± 4 to 22 ± 5 μmol m−2 h−1, which would be an insignificant influence on the average rate of DE of 186 ± 24 μmol m−2 h−1.
Relative contribution of denitrification and DIN assimilation to N dynamics
In order to evaluate the relative contribution of assimilatory and dissimilatory pathways to N dynamics we compared the theoretical nitrogen assimilation and measured denitrification rates (Fig. 5). The total benthic nitrogen demand calculated as the sum of theoretical assimilation and total denitrification strongly correlated with measured benthic DIN consumption with a ratio close to 1 for both the MPB and macrophyte colonised sites (Fig. 6), indicating that the measurements of denitrification and estimates of community N assimilation were reasonably accurate. These results also demonstrate that in this system, essentially all of the DIN consumed by the benthic compartment at both the MPB and P. pectinatus colonised sites can be accounted for by community N assimilation and denitrification, which are consequently the major processes driving benthic N dynamics in the lake. As discussed in previous sections, in both the MPB and P. pectinatus colonised areas denitrification based on water column nitrate was an important N sink and water column nitrate was also a major source of nitrogen for plant growth. Previous studies of aquatic macrophytes have shown a high degree of variability in the relative contributions of root and shoot assimilation of DIN to the overall N demand with the availability of DIN in the water column and the root to shoot ratio of the plant biomass cited as important controlling factors (Caffrey & Kemp, 1992). In the studied lake, due to the consistently high NO x concentrations in the water column and the relatively low root:shoot ratios (0.2–0.5) of the P. pectinatus population it would be expected that the water column would be the major source of DIN for the growth of P. pectinatus.

Seasonal evolution of total denitrification rates and theoretical community N assimilation at the P. pectinatus and MPB colonised sites. Data are reported as mean ± standard deviation (n = 3)

Relationship between community total benthic DIN consumption and the sum of denitrification and calculated community theoretical N assimilation rates at the P. pectinus (closed circles) and MPB (open circles) colonised sites. Data are reported as mean ± standard deviation (n = 3), the hatched line indicates a 1:1 relationship
On the annual scale, the ratio of community N assimilation to denitrification was a little below 1 indicating a slight dominance of denitrification over N assimilation as a sink for N. On average ~53% of the DIN consumed at both sampling sites was eliminated as N2 but this percentage varied between sampling dates especially at the P. pectinatus site (Fig. 5). At this site, N assimilation dominated during winter while in summer denitrification accounted for ~70% of benthic N consumption. Overall, our data support the growing body of evidence that the contribution of denitrification compared to plant N assimilation to the N dynamics in freshwater systems is systematically higher compared to that measured in coastal marine environments (Table 5), where denitrification plays a relatively minor role (e.g. Risgaard-Petersen et al., 1998; Risgaard-Petersen & Ottosen, 2000; Welsh et al., 2000; Bartoli et al., 2008; Dunn et al., 2012). The calculated mean annual contribution of denitrification to benthic DIN consumption also agrees well with the contributions calculated in other fresh water aquatic systems where denitrification accounted for 40–80% of total benthic DIN consumption (Matheson et al., 2002; Pinardi et al., 2009; Kreiling et al., 2011).
Overall, our results indicate that in contrast to what is generally observed in shallow water marine environments (Table 5) both denitrification and N assimilation play important and comparable roles in DIN removal in freshwater systems and suggest that shallow aquatic environments in agricultural landscapes may act as efficient N traps. In addition, our results suggest that the continued P. pectinatus colonisation of the littoral zone of the studied pit lake will modify the N dynamics of the lake by (1) reducing N mobility due to the longer retention time of assimilated N in the biomass, (2) potentially enhancing N-burial in the sediment due to the more refractory nature of the biomass produced and (3) enhancing N removal via denitrification.
References
APHA, 1998. Standard Methods for the Examination of Water and Wastewaters, 20th ed. APHA, Washington, DC.
Bartoli, M., D. Nizzoli, G. Castaldelli & P. Viaroli, 2008. Community metabolism and buffering capacity of nitrogen in a Ruppia cirrhosa meadow. Journal of Experimental Marine Biology and Ecology 360: 21–30.
Bower, C. E. & T. H. Hansen, 1980. A salicylate–hypochlorite method for determining ammonia in seawater. Canadian Journal of Fisheries and Aquatic Science 37: 794–798.
Burgin, A. J. & S. K. Hamilton, 2007. Have we overemphasized the role of denitrification in aquatic ecosystems? A review of nitrate removal pathways. Frontiers in Ecology and the Environment 5: 89–96.
Burgin, A. J., J. G. Lazar, P. M. Groffman, A. J. Gold & D. Q. Kellogg, 2012. Balancing nitrogen retention ecosystem services and greenhouse gas disservices at the landscape scale. Ecological Engineering. doi:10.1016/j.ecoleng.2012.05.003.
Caffrey, J. M. & W. M. Kemp, 1992. Influence of the submersed plant, Potamogeton perfoliatus, on nitrogen cycling in estuarine sediments. Limnology and Oceanography 37: 1483–1495.
Canfield, D. E., B. Thamdrup & E. Kristensen, 2005. Aquatic geomicrobiology. In Southward, A. J., P. A. Tyler, C. M. Young & L. A. Fuiman (eds), Advances in Marine Biology, Vol. 48. Elsevier Academic Press, Amsterdam: 640 pp.
Cattaneo, A. & J. Kalff, 1980. The relative contribution of aquatic macrophytes and their epiphytes to the production of macrophyte beds. Limnology and Oceanography 25: 280–289.
Christensen, P. B. & J. Sørensen, 1986. Temporal variation of denitrification activity in plant-covered, littoral sediment from Lake Hampen, Denmark. Applied and Environmental Microbiology 51: 1174–1179.
Dalsgaard, T., P. B. Christensen, S. Rysgaard & N. Risgaard-Petersen, 1999. Nitrogen removal in Danish coastal waters: importance and regulation (in Danish). In Lomstein B. A., (ed.), Marine environments into the millenium. Olsen & Olsen, Fredensborg, pp 102–118.
David, M. B., L. G. Wall, T. V. Royer & J. L. Tank, 2006. Denitrification and the nitrogen budget of a reservoir in an agricultural landscape. Ecological Applications 16: 2177–2190.
De Jonge, V. N., 1980. Fluctuations in the organic carbon to chlorophyll a ratios for estuarine benthic diatom populations. Marine Ecology Progress Series 2: 345–353.
den Heyer, C. & J. Kalff, 1998. Organic matter mineralization rates in sediments: a within- and among-lake study. Limnology and Oceanography 43: 695–705.
Dunn, R. J. K., D. T. Welsh, M. A. Jordan, N. J. Waltham, C. J. Lemckert & P. R. Teasdale, 2012. Benthic metabolism and nitrogen dynamics in a sub-tropical coastal lagoon: microphytobenthos stimulate nitrification and nitrate reduction through photosynthetic oxygen evolution. Estuarine Coastal and Shelf Science 113: 272–282.
Eriksson, P. G. & S. E. B. Weisner, 1999. An experimental study on effects of submersed macrophytes on nitrification and denitrification in ammonium-rich aquatic systems. Limnology and Oceanography 44: 1993–1999.
Galloway, J. N., J. D. Aber, J. W. Erisman, S. P. Seitzinger, R. W. Howarth, E. B. Cowling & B. J. Cosby, 2003. The nitrogen cascade. BioScience 53: 341–356.
Hansen, J. W., A. U. Pedersen, J. Berntsen, I. S. Rønbøg, L. S. Hansen & B. A. Lomstein, 2000. Photosynthesis, respiration, and nitrogen uptake by different compartments of a Zostera marina community. Aquatic Botany 66: 281–295.
Kantrud, H. A., 1990. Sago Pondweed (Potamogeton pectinatus L.): A Literature Review. U.S. Fish and Wildlife Service, Fish and Wildlife Resource Publication, 176, Washington, DC: 89 pp.
Karjalainen, H., G. Stefansdottir, L. Tuominen & T. Kairesalo, 2001. Do submersed plants enhance microbial activity in sediment? Aquatic Botany 69: 1–13.
Kreiling, R. M., W. B. Richardson, J. C. Cavanaugh & L. A. Bartsch, 2011. Summer nitrate uptake and denitrification in an upper Mississippi River Backwater Lake: the role of rooted aquatic vegetation. Biogeochemistry 104: 309–324.
Lorenzen, C. J., 1967. Determination of chlorophyll and phaeo-pigments: spectrophotometric equations. Limnology and Oceanography 12: 343–346.
Matheson, F. E., M. L. Nguyen, A. B. Cooper, T. P. Burt & D. C. Bull, 2002. Fate of 15N-nitrate in unplanted, planted and harvested riparian wetland soil microcosms. Ecological Engineering 19: 249–264.
McClain, M. E., E. W. Boyer, C. L. Dent, S. E. Gergel, N. B. Grimm, P. M. Groffman, S. C. Hart, J. W. Harvey, C. A. Johnston, E. Mayorga, W. H. McDowell & G. Pinay, 2003. Biogeochemical hot spots and hot moments at the interface of terrestrial and aquatic ecosystems. Ecosystems 6: 301–312.
Megonigal, J., M. Hines & P. Visscher, 2004. Anaerobic metabolism: linkages to trace gases and aerobic processes. In Schlesinger, W. H. (ed.), Biogeochemistry. Elsevier-Pergamon, Oxford: 317–424.
Nielsen, L. P., 1992. Denitrification in sediment determined from nitrogen isotope pairing. Federation of European Microbiological Societies, Microbiology Ecology 86: 357–362.
Nielsen, L. B., K. Finster, D. T. Welsh, A. P. Donnelly, R. A. Herbert, R. de Wit & B. A. Lomstein, 2001. Sulfate reduction and nitrogen fixation rates associated with roots, rhizomes and sediments from Zostera noltii and Spartina stricta meadows. Environmental Microbiology 3: 63–71.
Nielsen, S. L., G. T. Banta & M. F. Pedersen, 2004. Estuarine Nutrient Cycling: The Influence of Primary Producers, Vol. 2. Kluwer, Dordrecht: 263–280.
Nizzoli, D., D. T. Welsh, M. Bartoli & P. Viaroli, 2005. Impacts of mussel (Mytilis galloprovincialis) farming on oxygen consumption and nutrient recycling in a eutrophic coastal lagoon. Hydrobiologia 550: 183–198.
Nizzoli, D., E. Carraro, V. Nigro & P. Viaroli, 2010. Effect of organic enrichment and thermal regime on denitrification and dissimilatory nitrate reduction to ammonium (DNRA) in hypolimnetic sediments of two lowland lakes. Water Research 44: 2715–2724.
Nõges, T., H. Luup & T. Feldmann, 2009. Primary production of aquatic macrophytes and their epiphytes in two shallow lakes (Peipsi and Võrtsjärv) in Estonia. Aquatic Ecology 44: 83–92.
Ottosen, L. D. M., N. Risgaard-Petersen & L. P. Nielsen, 1999. Direct and indirect measurements of nitrification and denitrification in the rhizosphere of aquatic macrophytes. Aquatic Microbial Ecology 19: 81–91.
Palmer, M. A. & S. Filoso, 2009. Restoration of ecosystem services for environmental markets. Science 325: 575–576.
Piña-Ochoa, E. & M. Álvarez-Cobelas, 2006. Denitrification in aquatic environments: a cross-system analysis. Biogeochemistry 81: 111–130.
Pinardi, M., M. Bartoli, D. Longhi, U. Marzocchi, A. Laini, C. Ribaudo & P. Viaroli, 2009. Benthic metabolism and denitrification in a river reach: a comparison between vegetated and bare sediments. Journal of Limnology 68: 133–145.
Pind, A., N. Risgaard-Petersen & N. P. Revsbech, 1997. Denitrification and microphytobenthic NO3- consumption in a Danish lowland stream: diurnal and seasonal variation. Aquatic Microbial Ecology 12: 275–284.
Racchetti, E., M. Bartoli, C. Ribaudo, D. Longhi, L. Brito, M. Naldi, P. Iacumin & P. Viaroli, 2010. Short term changes in pore water chemistry in river sediments during the early colonization by Vallisneria spiralis. Hydrobiologia 652: 127–137.
Reddy, K. R., W. H. Patrick & C. W. Lindau, 1989. Nitrification–denitrification at the plant root–sediment interface in wetlands. Limnology and Oceanography 34: 1004–1013.
Revsbech, N., J. Jacobsen & L. P. Nielsen, 2005. Nitrogen transformations in microenvironments of river beds and riparian zones. Ecological Engineering 24: 447–455.
Risgaard-Petersen, N., 2003. Coupled nitrification–denitrification in autotrophic and heterotrophic estuarine sediments: on the influence of benthic microalgae. Limnology and Oceanography 48: 93–105.
Risgaard-Petersen, N., 2004. Denitrification. In Nielsen, S. L., G. T. Banta & M. F. Pedersen (eds), Estuarine Nutrient Cycling: The Influence of Primary Producers, Vol. 2. Kluwer, Dordrecht: 263–280.
Risgaard-Petersen, N. & K. Jensen, 1997. Nitrification and denitrification in the rhizosphere of the aquatic macrophyte Lobelia Dortmanna L. Limnology and Oceanography 42: 529–537.
Risgaard-Petersen, N. & L. D. M. Ottosen, 2000. Nitrogen cycling in two temperate Zostera marina beds: seasonal variation. Marine Ecology Progress Series 198: 93–107.
Risgaard-Petersen, N. & S. Rysgaard, 1995. Nitrate reduction in sediments and waterlogged soil measured by 15N techniques. In Alef, K. & P. Nannipieri (eds), Methods in Applied Soil Microbiology. Academic Press, London: 287–310.
Risgaard-Petersen, N., S. Rysgaard, L. P. Nielsen & N. P. Revsbech, 1994. Diurnal variation of denitrification in sediments colonised by benthic microphytes. Limnology and Oceanography 39: 573–579.
Risgaard-Petersen, N., T. Dalsgaard, S. Rysgaard, P. B. Christensen, J. Borum, K. McGlathery & L. P. Nielsen, 1998. Nitrogen balance of a temperate eelgrass Zostera marina bed. Marine Ecology Progress Series 174: 281–291.
Risgaard-Petersen, N., L. P. Nielsen, T. Dalsgaard & R. L. Meyer, 2003. Application of the isotope pairing technique in sediments where ANAMMOX and denitrification coexist. Limnology and Oceanography: Methods 1: 63–73.
Rysgaard, S., P. B. Christensen & L. P. Nielsen, 1995. Seasonal variation in nitrification and denitrification in estuarine sediment colonized by benthic microalgae and bioturbating infauna. Marine Ecology Progress Series 126: 111–121.
Sand-Jensen, K., C. Prahl & H. Stokolm, 1982. Oxygen release from roots of submerged aquatic macrophytes. Oikos 38: 349–354.
Saunders, D. L. & J. Kalff, 2001. Denitrification rates in the sediments of Lake Memphremagog, Canada–USA. Water Research 35: 1897–1904.
Schramm, W., E. Gualberto & C. Orosco, 1984. Release of dissolved organic matter from marine tropical reef plants: temperature and desiccation effects. Botanica Marina 27: 71–77.
Seitzinger, S., J. A. Harrison, J. K. Böhlke, A. F. Bouwman, R. Lowrance, B. Peterson, C. Tobias & G. Van Drecht, 2006. Denitrification across landscapes and waterscapes: a synthesis. Ecological Applications 16: 2064–2090.
Sokal, R. R. & F. J. Rohlf, 1995. Biometry. W.H. Freeman and Company, New York.
Søndergaard, M., L. Bruun, T. Lauridsen, E. Jeppesen & T. V. Madsen, 1996. The impact of grazing waterfowl on submerged macrophytes: in situ experiments in a shallow eutrophic lake. Aquatic Botany 53: 73–84.
Sundbäck, K. & A. Miles, 2000. Balance between denitrification and microalgal incorporation of nitrogen in microtidal sediment, NE Kattegat. Aquatic Microbial Ecology 22: 291–300.
Tavernini, S., D. Nizzoli, G. Rossetti & P. Viaroli, 2009. Trophic state and seasonal dynamics of phytoplankton communities in two sand-pit lakes at different successional stages. Journal of Limnology 68: 217–228.
Toet, S., L. H. F. A. Huibers, R. S. P. Van Logtestijn & J. T. A. Verhoeven, 2003. Denitrification in the periphyton associated with plant shoots and in the sediment of a wetland system supplied with sewage treatment plant effluent. Hydrobiologia 501: 29–44.
van Wijk, R. J., 1988. Ecological studies on Potamogeton pectinatus L. I. General characteristics, biomass production and life cycles under field conditions. Aquatic Botany 31: 211–258.
Veraart, A. J. & W. J. J. de Bruijne, 2011. Effects of aquatic vegetation type on denitrification. Biogeochemistry 104: 267–274.
Verhoeven, J. T. A., B. Arheimer, C. Yin & M. M. Hefting, 2006. Regional and global concerns over wetlands and water quality. Trends in Ecology & Evolution 21: 96–103.
Vestergaard, O. & K. Sand-Jensen, 2000. Alkalinity and trophic state regulate aquatic plant distribution in Danish lakes. Aquatic Botany 67: 85–107.
Viaroli, P. & R. R. Christian, 2003. Description of trophic status of an eutrophic coastal lagoon through potential oxygen production and consumption: defining hyperautotrophy and dystrophy. Ecological Indicators 3: 237–250.
Vitousek, P. M., J. D. Aber, R. W. Howarth, G. E. Likens, P. A. Matson, D. W. Schindler, W. H. Schlesinger & D. G. Tilman, 1997. Human alteration of the global nitrogen cycle: sources and consequences. Ecological Applications 5: 85.
Welsh, D. T., 2000. Nitrogen fixation in seagrass meadows: regulation, plant–bacteria interactions and significance to primary productivity. Ecology Letters 3: 58–71.
Welsh, D. T., P. K. Wellsbury, S. Bourguès, R. de Wit & R. A. Herbert, 1996. Relationship between porewater organic carbon content, sulphate reduction and nitrogen fixation (acetylene reduction) in the rhizosphere of Zostera noltii. Hydrobiologia 329: 175–183.
Welsh, D. T., M. Bartoli, D. Nizzoli, G. Castaldelli, S. A. Riou & P. Viaroli, 2000. Denitrification, nitrogen fixation, community primary productivity and inorganic-N and oxygen fluxes in an intertidal Zostera noltii meadow. Marine Ecology Progress Series 208: 65–77.
Wetzel, R. G., 1990. Land water interfaces: metabolic and limnological regulators, Verhein. International Verein Limnology 24: 6–24.
Wetzel, R. G., 2001. Limnology. Lake and River Ecosystems, 3rd ed. Academic Press, London: 1006 pp.
Wetzel, R. G. & G. E. Likens, 1991. Limnological Analyses, 2nd ed. Springer, New York: 391 pp.
Acknowledgments
This study was supported with funding from Italian Ministry of University and Research, Project PRIN 428 2008, ‘Nitrogen loads in the Po river basin: biogeochemical processes, transformations and effects in lowland reaches, transitional and coastal waters’ and by Bassanetti s.r.l. (Monticelli d’Ongina, Piacenza). The authors thank Emanuele Emani and Claudio Bassanetti that supported the organisation of this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: M. T. Ferreira, M. O’Hare, K. Szoszkiewicz & S. Hellsten / Plants in Hydrosystems: From Functional Ecology to Weed Research
Rights and permissions
About this article
Cite this article
Nizzoli, D., Welsh, D.T., Longhi, D. et al. Influence of Potamogeton pectinatus and microphytobenthos on benthic metabolism, nutrient fluxes and denitrification in a freshwater littoral sediment in an agricultural landscape: N assimilation versus N removal. Hydrobiologia 737, 183–200 (2014). https://doi.org/10.1007/s10750-013-1688-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-013-1688-1