
Abstract
This study investigates whether (1) structural configuration (considering substrate composition, wood debris, marginal and riparian vegetation) of streams suggests habitat simplification on a basin scale and (2) fish assemblage structure follows stream habitat configurations. Ninety-five sites in two pasture basins in southeastern Brazil were randomly selected. Two passes of electro-fishing were done in a 75-m reach at each site. Ten habitat descriptors related to in-stream and riparian physical structures were evaluated at each stream reach. Two groups of streams were identified by principal component analysis: a grassy and a non-grassy group. In comparison to non-grassy sites, grass-dominated streams exhibited a more homogenous fish fauna due to lower species diversity and higher dominance, especially by the guppy Poecilia reticulata, a highly generalist exotic species. The grassy group had not only the most simplified habitat but also the most simplified fish fauna, and the combination of grass-dominated, completely deforested, heavily silted conditions with recent land use conversion may cause a future habitat homogenization followed by a fish fauna homogenization on a regional scale.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The importance of in-stream habitat complexity as a determinant of lotic fish diversity was pioneered in the studies of temperate and tropical systems (e.g., Gorman & Karr, 1978; Schlosser, 1982; Angermeier & Karr, 1983, 1984), and such relevance has been continuously reported in several regions (e.g., Meffe & Sheldon, 1988; Pusey et al., 1993; Reyes-Gavilán et al., 1996; Martin-Smith, 1998; Gerhard et al., 2004; Mendonça et al., 2005). Habitat complexity influences movements of prey and predators through hazardous sites (Gilliam & Fraser, 2001) with higher fish vagility in areas where habitat heterogeneity is low, such as channelized and lowland streams (Gorman, 1986). Indeed, reduced habitat complexity may create an environment less favorable for large-sized juveniles and reinforce negative impacts of degradation of stream habitats, as Finstad et al. (2007) observed for salmonid populations.
Interaction among substrate, woody debris, and marginal grasses often increases in-stream habitat complexity and creates a combination of microhabitats usually explored by fish species with different needs (e.g., Gorman & Karr, 1978; Angermeier & Karr, 1984; Collier et al., 1999; Bührnheim & Cox-Fernandes, 2003; Growns et al., 2003; Torgersen & Close, 2004; Schneider & Winemiller, 2008). Removal of riparian vegetation represents one of the several mechanisms that can alter the quantity and quality of these elements, resulting in grass proliferation, increased sedimentation, and depleted woody debris (Pusey & Arthington, 2003; King & Warbuton, 2007 and references therein), which leads to a less complex in-stream habitat.
In-stream and riparian habitat alterations may influence several aspects of fish biology at local, regional, and global scales. At global, inter-basin, basin, and local scales, modification of habitats with elimination of biogeographic filters has contributed to the increasing local homogenization of the global freshwater fauna (Rahel, 2007). At reach and basin scales, for example, biomass of periphyton feeders in open canopy sites was shown to be higher in comparison to forested sites (Burcham, 1988; Esteves et al., 2008); in contrast, banks that have been cleared of trees and shrubs and become revegetated with grasses support fish assemblages with larger components of small fish, mostly microphagic carnivores and omnivores, when compared with banks covered with trees, which exhibited great number of herbivores (Growns et al., 2003). After environmental changes, biotic interaction among native species decreases, reducing their populations, and increases the vulnerability of their assemblages to invasion by non-native species (Baltz & Moyle, 1993) which are often tolerant and may become dominant in streams with degraded habitat and poor water quality (Paul & Meyer, 2001).
Brazil is notable for its high potential of natural renewable water resources, freshwater seafood production, and quantity of freshwater fish species, which represents more than 50% of the total registered for South America (Earthtrends, 2009). However, such potential and richness has been threatened due to years of pollution and eutrophication, silting, impounding and flood control, fisheries, and species introductions (Agostinho et al., 2005). An interesting example of such impairment is the northwestern region of São Paulo (Casatti et al., 2006) in which over 77% of the original forests were replaced by pasture, characterizing this area as the most degraded of the State, where only 4% of the original vegetation remains (SMA/IF, 2005). Using two pasture river basins of this region as models, we made the prediction that, in critically logged basins with high susceptibility to erosive processes, low-order streams must exhibit high resemblance regarding physical structure and fish fauna. Specifically, in this study our purpose was to answer the following questions: does structural configuration (considering substrate composition, wood debris, marginal and riparian vegetation) of streams suggest habitat simplification on a basin scale? Does fish assemblage structure follow stream habitat configurations?
Materials and methods
Study area and site selection
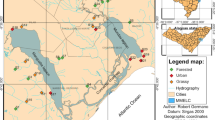
The study area is located in the northwestern portion of São Paulo State, in southeastern Brazil (Fig. 1), and belongs to the Serra Geral geological formation, composed of basaltic and sedimentary rocks of the Cauiá and Bauru groups. The sandy Botucatu and Pirambóia formations (São Bento group) are the aquifers of the region (IPT, 2000). According to the slope classification, the region is flat or smoothly rolling (Silva et al., 2007a), basins are highly susceptible to erosive processes, and pasture is the dominant land-cover class in general (occurring in 78.1% of the area) as well as in the 30 m riparian buffer (Silva et al., 2007b). The climate is hot tropical (Nimer, 1989), with average annual rainfall about 1,250 mm (Silva et al., 2007a). The rainy season lasts from October to March (January and February are the wettest months, with 54% of the annual rainfall) and the dry season from April to September; the maximum mean temperature (31°C) occurs in January and the minimum average (13°C) occurs in July (IPT, 2000). Human population density is about 61–84 inhabitants/km2 (Silva et al., 2007a).

Location of the study area with the 95 sites in the northwestern region of the São Paulo State, Brazil. Open squares refer to sites in the Rio São José dos Dourados basin (sampled in 2003), triangles and circles to Rio Turvo-Grande basin (triangles sampled in 2004 and circles in 2005)
Sites for sampling were selected from 1:50,000 topographic maps in the São José dos Dourados and Turvo-Grande watersheds, excluding urbanized areas. The number of sites was approximately proportional to the total length in kilometer for first-, second-, and third-order reaches (Roth et al., 1999). One site was randomly selected for each 100 km of a length order; to account for landowner refusals or unsampleable conditions, extra sites were selected as backups, following Kasyak (2001). After ensuring that the chosen sites could be effectively sampled, a 75-m stream reach was selected to include all available mesohabitats (pools, riffles, and runs) (Kasyak, 2001). Ninety-five reaches (Fig. 1, Appendix—see Supplementary material), comprising 56 of first order, 29 of second, and 10 of third, were sampled. For logistical reasons, each stream reach was sampled once during the dry seasons from 2003 to 2005 to reduce seasonal effected differences to a minimum (2003: sites 1–35; 2004: sites 36–56; 2005: sites 57–95).
Habitat evaluation and fish sampling
Ten habitat descriptors related to in-stream and riparian physical structures were estimated visually in broad categories at each stream reach as follows: proportions of bottom occupied by (1) clay, (2) sand, (3) gravel, (4) rock, (5) boulder, and (6) bedrock; proportion of internal habitat occupied by (7) woody debris; proportion of banks occupied by (8) marginal grasses (mostly weed Brachiaria spp. banks proliferated from pasture); proportion of (9) trees, and (10) shrubs across a riparian stripe of 10 m width.
Fishing effort was standardized across all collecting sites with each stream reach blocked at both up- and downstream by 5-mm-mesh stop nets and then submitted to two electric fishing passes for a total of 40–50 min. Electro-fishing equipment consists of a stationary generator of alternating current (220 V, 50–60 Hz, 3.4–4.1 A, 1,000 W), as detailed by Castro et al. (2003). Captured specimens were fixed in 10% formalin solution and, after 48 h, transferred to a 70% EtOH solution. Fishes were identified to species and counted. All specimens were deposited at the fish collection of the Departamento de Zoologia e Botânica da Universidade Estadual Paulista (DZSJRP), São José do Rio Preto, São Paulo, Brazil.
Analysis
In order to investigate the habitat simplification hypothesis, a principal component analysis (PCA) was performed by using a covariance matrix with the values of 10 habitat descriptors of the 95 sites, employing software PC-Ord version 4 (McCune & Mefford, 1999). In order to decide which components (axes) would be retained for interpretation, broken-stick eigenvalues were compared to actual eigenvalues for each axes; all axis with eigenvalues greater than the broken-stick eigenvalues were selected for interpretation (McCune & Mefford, 1999). Following a similar approach to MacRae & Jackson (2001), standard deviation of site scores for axis 1 and 2 produced by PCA was calculated to assess if, within predefined stream groups, habitat could be considered more or less simplified.
The rarefaction method (Simberloff, 1972) in the software Biodiversity Pro (McAleece et al. 1997) was used to compare species richness of the groups. Species diversity in each stream reach was evaluated by the Shannon–Wiener index with base 10 logarithm and dominance by the Simpson index (Magurran, 2004). Univariate comparisons of species diversity and dominance between stream groups were further investigated using T-test for independent samples (0.05 of significance level). After using the Bray–Curtis similarity index based on presence/absence in the Primer software (Clarke & Gorley, 2006) to produce a similarity matrix for each predefined stream group, the average similarity of fish faunas was calculated to assess the ichthyofaunistic simplification within each stream group.
Results
From the PCA, only axis 1 demonstrated an eigenvalue (154,168) greater than the broken-stick value (74,977), being retained for interpretation. The positive loadings were calculated for grass and sand, and the negative loadings for remaining descriptors, which defined two groups of streams (Table 1, Fig. 2): a grassy and a non-grassy group. After removing sites 3, 4, and 66, which were not closed assigned to any group, 47 streams were included in the grassy group and 45 streams in the non-grassy group. The standard deviation of PCA site scores for the grassy group (axis 1 = 11.62, axis 2 = 7.39) was lower than for the non-grassy (axis 1 = 17.96, axis 2 = 19.08), indicating a more simplified habitat within the grassy group.

Projection of the two-first principal components of 95 sites and ten habitat descriptors. Numbers refer to the stream reach identification. Cumulative variance from axis 1 and 2 was 76%
In these 92 sites, 15,627 fish belonging to 64 species, distributed in 19 families and six orders (Table 2), were registered. Of these, 49 species and 9,989 specimens were associated with the grassy group and 56 species and 5,638 specimens with the non-grassy group. Rarefaction curves confirmed the highest species richness in the non-grassy group (Fig. 3). Poecilia reticulata, Astyanax altiparanae, Knodus moenkhausii, and Aspidoras fuscoguttatus were the most representative species in the overall abundance. Eight species were present in more than 50% of the sampled reaches (A. altiparanae, Oligosarcus pintoi, Hoplias malabaricus, A. fuscoguttatus, Hypostomus ancistroides, Rhamdia quelen, Gymnotus carapo, and P. reticulata). The non-grassy group encompassed 67% of the overall rare species richness (n = 21), that is, rare species having <10 specimens, against 52% registered in the grassy group. Among rare species, Planaltina britskii, Apareiodon piracicabae, Cetopsorhamdia iheringi, Phenacorhamdia tenebrosa, and Pseudopimelodus pulcher occurred in reaches where P. reticulata was represented by no more than two individuals.

Rarefaction curves based on abundance of fish fauna from grassy and non-grassy stream groups
The T-test for ecological descriptors revealed that, in comparison to non-grassy sites, grassy sites had lower diversity (P = 0.026) and higher dominance (P = 0.017), attributed to the guppy P. reticulata, which contributed to 31.98% of the total abundance in the grassy group and 11.33% in the non-grassy (Table 2). Average Bray–Curtis similarity calculated on presence/absence data was 45% in the grassy group and 37% in the non-grassy group, indicating that grassy sites have more simplified fish faunas than non-grassy sites.
Discussion
Two stream groups were determined, representing, on a macro scale, the stream types prevalent in the studied region. Considering the pasture predominance, it is not surprising that almost half of the randomly selected streams belonged to the grassy group. This particular kind of stream physiognomy is characterized by extensive proliferation of Poaceae grasses (mostly Brachiaria spp.) on the stream banks. Despite serving to stabilize stream banks in the absence of riparian vegetation, exacerbated proliferation of grasses might lead to a suite of changes in habitat structure, water quality, food web structure, flow diversity into the channel, and in the fish assemblages (Pusey & Arthington, 2003 and references therein). Thus, an intermediate condition of grass proliferation, rather than as homogenous growths across the channel, would contribute to providing structural complexity to stream habitat (Collier et al., 1999) but, when dominant, may have negative consequences for the aquatic biota (Pusey & Arthington, 2003).
Streams classified in the non-grassy group, in contrast, exhibited a reasonable substrate diversification, as indicated by habitat descriptors values and PCA scores. In fact, this group was closely assigned to more complex riparian conditions (presence of wood debris, trees, and shrubs), reinforcing the linkage between in-stream and riparian habitats. In the studied region, as has occurred worldwide, riparian clearing is critical and a land-cover study using satellite scenes from 1996 and 1997 showed only 25% of riparian buffer zone preservation (Silva et al., 2007a). The low number of reaches associated with wood debris and trees reflects this critical situation. Despite some level of physical habitat diversification, which indicates that stream habitats are not completely simplified at basin scale, the fact that more than half of the streams are grass-dominated, completed logged, heavily silted, and bottom-simplified deserves attention. In accordance with Bunn et al. (1997), we do not advocate eliminating grasses, or other invasive vegetation, from stream reaches because, as stated by those authors, it may mobilize vast quantities of sediment currently trapped among vegetation roots, and this will increase silting downstream. Re-establishment of the riparian canopy is of particular importance because it represents the most cost-effective source of a long-term control of invasive plants on the stream channel (Bunn et al., 1997), also buffering sediment inputs and improving in-stream habitat quality for aquatic biota.
Grassy reaches have more simplified fish faunas than non-grassy, a feature mainly attributed to the dominance of the exotic guppy P. reticulata. Guppies are small live-bearing fish native to Venezuela, Barbados, Trinidad, Northern Brazil, and the Guyanas (Welcomme, 1988). Through accidental or deliberate release, mostly for mosquito control, this species has successfully colonized at least 54 countries worldwide (FAO, 2009). Individuals are able to survive—and even to become dominant—in conditions of abrupt temperature changes (Chung, 2001), poor habitat quality (Casatti et al., 2006), and hypoxia (Kramer & Mehegan, 1981; Welcomme, 1988). In fact, guppies’ dominance is indicative of possible anthropogenic environmental impacts (Kennard et al., 2005; Cunico et al., 2006). A closer relationship between guppies and grasses was already mentioned by Pusey & Arthington (2003) and may represent a case of “invasional meltdown,” characterized by Simberloff (2006) as a community-level phenomenon in which a non-indigenous species facilitates another’s invasion, increasing its likelihood of survival and/or magnitude of impact. Native riparian forest replacement by pasture sometimes contributes to the establishment of non-native grass stands at open-canopy banks, which represents a shallow and warm microhabitat, difficult to access for predators, and widely used by guppies.
Influences of guppies on studied fish fauna were not evaluated at all, though the occurrence of rare species mostly in the non-grassy group (for example, A. piracicabae, Hypostomus sp., and P. pulcher) and the occurrence of some rare species in reaches with few P. reticulata specimens seem to mirror the fact that invading fishes in altered habitats are often associated with indirect changes in species composition and local extinction of native species. For example, altered abiotic conditions allowed non-native fishes to establish in California streams, and these new species henceforth eliminated native ones through competition or predation (Baltz & Moyle, 1993). In certain abiotic conditions or even at high human disturbance levels, exotic species presumably have a high potential to become successful invaders, regardless of the biota already present (Moyle & Light, 1996).
Although some species have broad distribution across the studied sites, fish assemblages were quite different between stream groups. Higher species diversity in the non-grassy group reflects a clear relationship between substrate diversity and more microhabitats mediating the coexistence of distinct species, especially bottom dwellers. A direct relationship between substrate diversity and fish diversity was reported in Indiana and Panama streams, being considered an important predictor of riffle-guild diversity (Gorman & Karr, 1978). In addition, streams with hard substrates presumably can harbor fish communities having more trophic modes. For instance, I. schubarti, C. zebra, Hypostomus sp., and P. nasus are all bottom-dwelling species associated with hard substrates (Casatti & Castro, 1998; Casatti et al., 2005); the first one is a grubber which excavates while moving (sensu Sazima, 1986) and consumes mostly aquatic insects; the second one is a sit-and-wait predator that ambushes its prey by staying motionless and dashing at it from close quarters (sensu Sazima, 1986); and the two last ones are grazers (sensu Keenleyside, 1979) that feed on rock-attached periphyton (Casatti & Castro, 1998; Casatti et al., 2005). Thus, elimination of hard substrates may result in low representativeness of species with similar feeding behaviur.
The higher species dominance in the grassy group must mirror the in-stream habitat simplification, leading to an environment in which only a few species are favored, and presumably with little diversity of trophic modes. In fact, the dominant species (K. moenkhausii, A. altiparanae, A. fuscoguttatus, and P. reticulata) in this set of streams are opportunistic omnivores with a tendency to insectivory which do not depend at all on diversified and hard bottom substrates (Ceneviva-Bastos & Casatti, 2007; pers. obs.). These findings are in accordance with Pereira et al. (2007), who studied the trophic structure of the ichthyofauna associated with riverine sandbanks and reported a simplified habitat, with little shelter and food available, and predominance of feeding generalist species. Indeed, the frequency and types of feeding interactions between species may follow habitat simplification. For example, an increase of interactions between nuclear and followers species could be expected in silted environments, as demonstrated in the case of the nuclear A. fuscoguttatus, whose individuals dig in the soft bottoms during feeding, promoting a cloud of suspended particles which are caught by the followers K. moenkhausii, P. reticulata, and A. altiparanae (Teresa & Carvalho, 2008).
The grassy group has not only the most simplified habitat but also the most simplified fish fauna—in accordance with the higher average Bray–Curtis similarity—which would suggest a condition of habitat homogenization followed by faunal homogenization. The presumed local consequence of extensive habitat homogenization is the widespread colonization of streams by P. reticulata, thus characterizing a type of biotic homogenization that conforms to the scenario 1 proposed by Rahel (2002), in which the invasive colonization of an exotic species may increase the similarity among sites.
The catchments of the studied area have been extensively used for pasture over the last decades, showing high levels of deforestation with intense use of the riparian zones (Silva et al., 2007a). Since 2005, however, livestock farmers have been progressively replacing pasture with sugar cane crops aiming to provide ethanol as an alternative to fossil fuels (Novaes, 2007), with strong expectations of an invasive input of this vegetation across riparian zones (see Silva et al., 2007a). We have little knowledge about the effects of such changes on the regional aquatic biota (but see Corbi & Trivinho-Strixino, 2008; Corbi et al., 2008); however, considering some studies conducted in other productive sugar cane areas of the world (Bunn et al., 1997; Fanning & Bohl, 2002; Kishimba et al., 2004), the scenarios are unpleasant.
In conclusion, in comparison to non-grassy sites, grass-dominated streams exhibited a more homogenous fish fauna due to lower species diversity and higher dominance, especially by the guppy, a highly generalist exotic species. Indeed, it is presumed that, on a regional scale, the combination of grass-dominated, completely deforested, heavily silted conditions with recent land use conversion may produce a future habitat homogenization followed by a fish fauna homogenization.
References
Agostinho, A. A., S. M. Thomaz & L. C. Gomes, 2005. Conservation of the biodiversity of Brazil’s inland waters. Conservation Biology 19: 646–652.
Angermeier, P. L. & J. R. Karr, 1983. Fish communities along gradients in a system of tropical streams. Environmental Biology of Fishes 9: 117–135.
Angermeier, P. L. & J. R. Karr, 1984. Relationships between woody debris and fish habitat in a small warmwater stream. Transactions of the American Fisheries Society 113: 716–726.
Baltz, D. M. & P. B. Moyle, 1993. Invasion resistance to introduced species by a native assemblage of California stream fishes. Ecological Applications 3: 246–255.
Buckup, P. A., N. A. Menezes & M. S. Ghazzi, 2007. Catálogo das espécies de peixes de água doce do Brasil. Série Livros, Museu Nacional, Rio de Janeiro.
Bührnheim, C. M. & C. Cox-Fernandes, 2003. Structure of fish assemblages in Amazonian rain-forest streams: effects of habitats and locality. Copeia 2003(2): 255–262.
Bunn, S. E., P. M. Davies & D. M. Kellaway, 1997. Contributions of sugar cane and invasive pasture grass to the aquatic food web of a tropical lowland stream. Marine and Freshwater Research 48: 173–179.
Burcham, J., 1988. Fish communities and environmental characteristics of two lowland streams in Costa Rica. Revista de Biología Tropical 36: 273–285.
Cambray, J. A., 2003. Impact on indigenous species biodiversity caused by the globalization of alien recreational freshwater fisheries. Hydrobiologia 500: 217–230.
Casatti, L. & R. M. C. Castro, 1998. A fish community of the São Francisco river headwater riffles, southeastern Brazil. Ichthyological Exploration of Freshwaters 9: 229–242.
Casatti, L., F. C. Rocha & D. C. Pereira, 2005. Habitat use by two species of Hypostomus (Pisces, Loricariidae) in southeastern Brazilian streams. Biota Neotropica 5: 157–165.
Casatti, L., F. Langeani, A. M. Silva & R. M. C. Castro, 2006. Stream fish, water and habitat quality in pasture dominated basin, Southeastern Brazil. Brazilian Journal of Biology 66: 681–696.
Castro, R. M. C., L. Casatti, H. F. Santos, K. M. Ferreira, A. C. Ribeiro, R. C. Benine, G. Z. P. Dardis, A. L. A. Melo, R. Stopiglia, T. X. Abreu, F. A. Bockmann, M. Carvalho, F. Z. Gibran & F. C. T. Lima, 2003. Estrutura e composição da ictiofauna de riachos do Rio Paranapanema, sudeste e sul do Brasil. Biota Neotropica 3: 1–31.
Ceneviva-Bastos, M. & L. Casatti, 2007. Oportunismo alimentar de Knodus moenkhausii (Teleostei, Characidae): uma espécie abundante em riachos do noroeste do Estado de São Paulo, Brasil. Iheringia 97: 7–15.
Chung, K. S., 2001. Critical thermal maxima and acclimation rate of the tropical guppy Poecilia reticulata. Hydrobiologia 462: 253–257.
Clarke, K. R. & R. N. Gorley, 2006. Primer v6: User Manual/Tutorial. Plymouth Marine Laboratory, Plymouth.
Collier, K. J., P. D. Champion & G. F. Crocker, 1999. Patch- and reach-scale dynamics of a macrophyte-invertebrate system in a New Zealand lowland stream. Hydrobiologia 392: 89–97.
Corbi, J. J. & S. Trivinho-Strixino, 2008. Relationship between sugar cane cultivation and stream macroinvertebrate communities. Brazilian Archives of Biology and Technology 51: 769–779.
Corbi, J. J., S. Trivinho-Strixino & A. Santos, 2008. Environmental evaluation of metals in sediments and dragonflies due to sugar cane cultivation in Neotropical streams. Water, Air, and Soil Pollution 195: 325–333.
Cunico, A. M., A. A. Agostinho & J. D. Latini, 2006. Influência da urbanização sobre as assembléias de peixes em três córregos de Maringá, Paraná. Revista Brasileira de Zoologia 23: 1101–1110.
Earthtrends, 2009. Earthtrends: the environmental information portal. World Resources Institute [available on internet at http://earthtrends.wri.org/country_profiles/index.php?theme=2]. Accessed 25 February 2009.
Esteves, K. E., A. V. P. Lobo & M. D. R. Faria, 2008. Trophic structure of a fish community along environmental gradients of a subtropical river (Paraitinga River, Upper Tietê River Basin, Brazil). Hydrobiologia 598: 373–387.
Fanning, D. J. & H. P. Bohl, 2002. Oxygen Depletion in Waterways in Sugarcane Growing Areas in North Queensland, 1999-2002: Methods for Stream Monitoring and Runoff Experiments. Technical Report 31/02, CSIRO Land and Water, Townsville.
FAO, 2009. Database on introduction of aquatic species (DIAS) [available on internet at http://www.fao.org/fishery/introsp/search]. Accessed 18 March 2009.
Finstad, A. G., S. Einum, T. Forseth & O. Ugedal, 2007. Shelter availability affects behaviour, size-dependent and mean growth of juvenile Atlantic salmon. Freshwater Biology 52: 1710–1718.
Gerhard, P., R. Moraes & S. Molander, 2004. Stream fish communities and their associations to habitat variables in a rain forest reserve in southeastern Brazil. Environmental Biology of Fishes 71: 321–340.
Gilliam, J. F. & D. F. Fraser, 2001. Movements in corridors: enhancement by predation threat, disturbance, and habitat structure. Ecology 82: 258–273.
Gorman, O. T., 1986. Assemblage organization of stream fishes: the effect of rivers in adventitious streams. American Naturalist 128: 611–616.
Gorman, O. T. & J. R. Karr, 1978. Habitat structure and stream fish communities. Ecology 59: 507–515.
Growns, I., P. C. Gerke, K. L. Astles & D. A. Pollard, 2003. A comparison of fish assemblages associated with different riparian vegetation types in the Hawkesbury–Nepean River system. Fisheries Management & Ecology 10: 209–220.
IPT (Instituto de Pesquisas Tecnológicas do Estado de São Paulo), 2000. Diagnóstico da situação atual dos Recursos Hídricos e estabelecimento de diretrizes técnicas para a elaboração do plano da Bacia Hidrográfica do São José dos Dourados. Relatório no 40675. Fundo Estadual de Recursos Hídricos, São Paulo.
Kasyak, P. F., 2001. Maryland biological stream survey: sampling manual. Maryland Department of Natural Resources, Monitoring and Non-tidal Assessment Division, Annapolis.
Keenleyside, M. H. A., 1979. Diversity and Adaptation in Fish Behaviour. Springer, Berlin.
Kennard, M. J., A. H. Arthington, B. J. Pusey & B. D. Harch, 2005. Are alien fish a reliable indicator of river health? Freshwater Biology 50: 174–193.
King, S. & K. Warbuton, 2007. The environmental preferences of three species of Australian freshwater fish in relation to the effects of riparian degradation. Environmental Biology of Fishes 78: 307–316.
Kishimba, M. A., L. Henry, H. Mwevura, A. J. Mmochi, M. Mihale & H. Hellar, 2004. The status of pesticide pollution in Tanzania. Talanta 64: 48–53.
Kramer, D. L. & J. P. Mehegan, 1981. Aquatic surface respiration, an adaptative response to hypoxia in the guppy, Poecilia reticulata (Pisces, Poeciliidae). Environmental Biology of Fishes 6: 299–313.
MacRae, P. S. D. & D. A. Jackson, 2001. Influence of smallmouth bass predation and habitat complexity on the structure of littoral zone fish assemblages. Canadian Journal of Fisheries and Aquatic Sciences 58: 342–351.
Magurran, A. E., 2004. Measuring Biological Diversity. Blackwell Publishing, Oxford.
Martin-Smith, K. M., 1998. Relationships between fishes and habitat in rainforest streams in Sabah, Malaysia. Journal of Fish Biology 52: 458–482.
McAleece, N., P. J. D. Lambshead, G. L. J. Paterson & J. G. Gage, 1997. BioDiversity Professional. Beta-Version. The Natural History Museum and Scottish Association for Marine Sciences.
McCune, B. & M. J. Mefford, 1999. PC-ORD. Multivariate Analysis of Ecological Data. Version 4. MjM Software Design, Gleneden Beach, Oregon.
Meffe, G. K. & A. L. Sheldon, 1988. The influence of habitat structure on fish assemblage composition in southeastern blackwater streams. The American Midland Naturalist 120: 225–240.
Mendonça, F. P., W. E. Magnusson & J. A. Zuanon, 2005. Relationships between habitat characteristics and fish assemblages in small streams of central Amazonia. Copeia 2005: 751–764.
Moyle, P. B. & T. Light, 1996. Fish invasions in California: do abiotic factors determine success? Ecology 77: 1666–1670.
Nimer, E., 1989. Climatologia do Brasil. IBGE, Rio de Janeiro.
Novaes, J. R. P., 2007. Campeões de produtividade: dores e febres nos canaviais paulistas. Estudos Avançados 21: 167–177.
Paul, M. J. & J. L. Meyer, 2001. Streams in the urban landscape. Annual Review of Ecology and Systematics 32: 333–365.
Pereira, P. R., C. S. Agostinho, R. J. Oliveira & E. E. Marques, 2007. Trophic guilds of fishes in sandbank habitats of a Neotropical river. Neotropical Ichthyology 5: 399–404.
Pusey, B. J. & A. H. Arthington, 2003. Importance of the riparian zone to the conservation and management of freshwater fish: a review. Marine and Freshwater Research 54: 1–16.
Pusey, B. J., A. H. Arthington & M. G. Read, 1993. Spatial and temporal variation in fish assemblage structure in the Mary River, south-east Queensland: the influence of habitat structure. Environmental Biology of Fishes 37: 355–380.
Rahel, F. J., 2002. Homogenization of freshwater faunas. Annual Review of Ecology and Systematics 33: 291–315.
Rahel, F. J., 2007. Biogeographic barriers, connectivity and homogenization of freshwater faunas: it’s a small world after all. Freshwater Biology 52: 696–710.
Reyes-Gavilán, F. G., R. Garrido, A. G. Nicieza, M. M. Toledo & F. Braña, 1996. Fish community variation along physical gradients in short streams of northern Spain and the disruptive effect of dams. Hydrobiologia 321: 155–163.
Roth, N. E., M. T. Southerland, G. Mercurio, J. C. Chaillou, P. F. Kasyak, S. S. Stranko, A. P. Prochaska, D. G. Heimbuch & J. C. Seibel, 1999. State of Streams: 1995-1997 Maryland Biological Stream Survey Results. Prepared by Versar Inc. for the Maryland Department of Natural Resources, Monitoring and Non-Tidal Assessment Division, Annapolis, Maryland, CNWP-MANTA-EA-99-6.
Sazima, I., 1986. Similarities in feeding behaviour between some marine and freshwater fishes in two tropical communities. Journal of Fish Biology 29: 53–65.
Schlosser, I. J., 1982. Fish community structure and function along two habitat gradients in a headwater stream. Ecological Monographs 52: 395–414.
Schneider, K. N. & K. O. Winemiller, 2008. Structural complexity of woody debris patches influences fish and macroinvertebrate species richness in a temperate floodplain-river system. Hydrobiologia 610: 235–244.
Silva, A. M., M. A. Nalon, F. J. N. Kronka, C. A. Álvares, P. B. Camargo & L. A. Martinelli, 2007a. Historical land-cover/use in different slope and riparian buffer zones in watersheds of the State of São Paulo, Brazil. Scientia Agricola 64: 325–335.
Silva, A. M., L. Casatti, C. A. Álvares, A. M. Leite, L. A. Martinelli & S. F. Durrant, 2007b. Soil loss and habitat quality in streams of a meso-scale river basin. Scientia Agricola 64: 336–343.
Simberloff, D., 1972. Properties of the rarefaction diversity measurement. American Naturalist 106: 414–418.
Simberloff, D., 2006. Invasional meltdown 6 years later: important phenomenon, unfortunate metaphor, or both? Ecological Letters 9: 912–919.
SMA/IF (Secretaria do Meio Ambiente/Instituto Florestal), 2005. Inventário florestal da vegetação natural do Estado de São Paulo. Imprensa Oficial, São Paulo.
Teresa, F. B. & F. R. Carvalho, 2008. Feeding association between benthic and nektonic Neotropical stream fishes. Neotropical Ichthyology 6: 109–111.
Torgersen, C. E. & D. A. Close, 2004. Influence of habitat heterogeneity on the distribution of larval Pacific lamprey (Lampetra tridentata) at two spatial scales. Freshwater Biology 49: 614–630.
Welcomme, R. L., 1988. International Introductions of Inland Aquatic Species. FAO Fisheries Technical Paper T294, Food and Agriculture Organization of the United Nations, Rome.
Acknowledgements
We thank the colleagues at the Laboratório de Ictiologia for their help during field work and IBILCE-UNESP for facilities, IBAMA for collecting license (001/2003), landowners for allowing to conduct research on their properties, Mônica Ceneviva-Bastos and Luis H. Z. Branco for comments, anonymous reviewers for helpful suggestions, Francisco Langeani for fish identification, David R. Mercer for language revision, “BIOTA/FAPESP Program” (www.biota.org/br) of the Fundação de Amparo à Pesquisa do Estado de São Paulo and Conselho Nacional de Desenvolvimento Científico e Tecnológico for financial support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: J. A. Cambray
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Casatti, L., de Paula Ferreira, C. & Carvalho, F.R. Grass-dominated stream sites exhibit low fish species diversity and dominance by guppies: an assessment of two tropical pasture river basins. Hydrobiologia 632, 273–283 (2009). https://doi.org/10.1007/s10750-009-9849-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-009-9849-y