
Abstract
Osteoblasts play a major role in bone remodeling and are regulated by transcription factors. GATA4, a zinc finger transcription factor from the GATA family, has an unclear role in osteoblast differentiation. In this study, the role of GATA4 in osteoblast differentiation was studied both in vitro and in vivo by GATA4 knockdown. GATA4 expression increased during osteoblast differentiation. GATA4 knockdown in osteoblast precursor cells reduced alkaline phosphatase activity and decreased the formation of calcified nodule in an osteogenic-induced cell culture system. In vivo, micro-CT showed that local injection of lentivirus-delivered GATA4 shRNA caused reduced new bone formation during tooth movement. Histological analyses such as total collagen and Goldner’s trichrome staining confirmed these results. In vivo immunohistochemical analysis showed reduced expression of osterix (OSX), osteopontin (OPN), and osteocalcin (OCN) in the shGATA4 group (P < 0.05). Consistently, both western blotting and quantitative reverse-transcription PCR proved that expression of osteogenesis-related genes, including OSX, OPN, and OCN, was significantly repressed in the shGATA4 group in vitro (P < 0.01). For further analysis of the pathways involved in this process, we examined the MAPK signaling pathway, and found knockdown of GATA4, downregulated p38 signaling pathways (P < 0.01). Collectively, these results imply GATA4 is a regulator of osteoblastic differentiation via the p38 signaling pathways.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Physiologically, the bone remodeling is a complex and a regulated process. The two type of cells involved in bone remodeling: osteoclasts that modulate bone resorption and are derived from hematopoietic cells and osteoblasts that mediate new bone formation and are derived from mesenchymal cells (Pittenger et al. 1999; Shoji et al. 2010; Tripuwabhrut et al. 2013; Yokoya et al. 1997). Balance between osteoclasts and osteoblasts is vital for bone homeostasis, whereas an imbalance may lead to bone diseases such as osteoporosis, hyperostosis, and periodontal disease (Teitelbaum 2000).
Proliferation, differentiation, and maturation of osteoblasts are critical events in bone formation and remodeling. Osteoblast differentiation is a complex process, consisting of maturation of preosteoblasts into matrix-secreting mature osteoblasts (Pittenger 2008; Rodriguez-Lozano et al. 2011). Osteoblasts synthesize and release various osteoblast-specific marker proteins such as alkaline phosphatase (ALP), osterix (OSX), osteopontin (OPN), and osteocalcin (OCN), which not only function as structural support, but also play a role in osteoblast maturation and function with regard to cell interaction, matrix deposition, and mineralization (de Oliveira et al. 2003; Wu et al. 2015). Therefore, differentiation of osteoblasts is controlled by orchestration of diverse transcription factors.
GATA4, a transcription factor, and a member of the GATA zinc finger family, proves to be a cardiac-specific transcriptional activator. Previous studies have pointed out that GATA4 is not only necessary for stabilization of postnatal cardiac function and avoiding stress-induced heart failure but also controls pathological cardiac hypertrophy, which may give rise to serious cardiac diseases. In addition to its important role in cardiac pathology, GATA4 has also been reported to regulate transcription in the gut, lungs, and ovaries (Mazaud et al. 2007; Molkentin et al. 1998; Viger et al. 2008). Mitogen-activated protein kinases (MAPKs) cascades are evolutionarily conserved and are the fundamental mechanism for cellular responses to a broad range of extracellular signals, especially many of the extracellular ligands related to osteoblasts, such as PTH, BMPs, noncanonical WNTs, FGFs, and TNF (Suzuki et al. 2002; Xiao et al. 2000; Zhou et al. 2006). Previous studies have shown that GATA4 is a vital transcription factor that induces/regulates the hypertrophy partly by receiving signals from MAPKs in the heart (van Berlo et al. 2011). Nevertheless, the synergistic effect of GATA4 and MAPK in osteoblast differentiation is still unclear.
In this study, the role of GATA4 in the regulation of the osteoblast differentiation and bone formation process was examined. The study showed that knockdown of GATA4 in osteoblast precursor cells and in mouse tooth movement (TM) model inhibited osteoblast differentiation and decreased expression of bone-related genes both in vitro and in vivo. Additionally, the study showed that p38 signaling mediates these processes under the influence of GATA4. These results imply GATA4 is a vital regulator of osteoblast differentiation, which may provide a new understanding to osteogenesis and lead to better treatment alternatives for bone diseases.
Materials and methods
Experimental animals
Mice (C57BL/6J) were housed in the Nanjing Medical University animal facility and had free access to mouse water and chow. All animals were handled in compliance with protocols approved by the Experimental Animal Care and Use Committee of Nanjing Medical University (permit no. 2015-03-40).
Osteoblast precursor cells isolation and culture
Primary osteoblast precursor cells were acquired from calvarial bones of 3-day-old mice. Initially, calvarial bones were separated and placed in phosphate-buffered saline (PBS) to remove the surrounding soft tissues. Calvarial pieces were then digested with 0.25% trypsin–EDTA for 10 min, then 0.2% collagenase in Hank’s solution for 30 min, and, finally, with 0.2% collagenase in Hank’s solution for 60 min, all at 37 °C. Following the third digestion, cells were collected and resuspended in complete medium with Minimum Essential Medium Alpha (α-MEM, Gibco, Invitrogen, Grand Island, USA), 15% fetal bovine serum (FBS, Invitrogen), 1% penicillin–streptomycin, and cultured at 37 °C with 5% CO2 (Zhong et al. 2016).
Flow cytometric analysis
Osteoblast precursor cells from the second passage were used to characterize stem cell surface markers by flow cytometric (FCM) analysis. These osteoblast precursor cells were harvested by 0.25% tryptase without EDTA, then washed with PBS 3 times, and resuspended. Cell suspensions (2 × 106 cells) were gained and incubated for 20 min at 4 °C in the dark with CD29-APC, CD34-FITC, CD45-PE, and CD105-PE antibodies (BD Biosciences, San Jose, USA). After staining, cells were washed twice in PBS and then analyzed with flow cytometry (Ge et al. 2016; Yu et al. 2012).
Osteogenic differentiation
Cells were plated in 6-well plates at a density of 1.5 × 105 cells/well with osteogenic differentiation medium including 10% FBS, 10 mM β-glycerolphosphate (Sigma, St. Louis, USA), 10 nM dexamethasone (Sigma), and 50 mg/mL ascorbate phosphate (Sigma).
Lentiviral transfection
Knockdown of GATA4 in osteoblast precursor cells was conducted by lentiviral transduction of short-hairpin RNAs (shRNAs; GenePharma, Shanghai, China). The target sequence of shRNA with a green fluorescent protein (GFP) tag to knockdown mouse GATA4 (shGATA4) was 5′-CCAAGCAGGACTCTTGGAA-3′. A scrambled sequence that was also tagged with GFP (5′-TTCTCCGAACGTGTCACGT-3′) was taken as the negative control (shCTRL). The multiplicity of infection (MOI) was estimated basing on a published protocol (Maranga et al. 2004). Then, recombinant lentivirus was applied to infect osteoblast precursor cells with an MOI of 50 in a medium supplemented with 5 μg/mL Polybrene (GenePharma), the medium was switched to a normal medium 24 h after transfection. After 3-day infection, fluorescence microscopy (Leica Microsystems Inc, Ontario, Canada) was chosen to measure the efficiency of lentiviral infection, and the knockdown efficiency of GATA4 was assessed by Western blotting after both 3-day and 14-day infection. To perform the mineralization induction experiment, the medium was switched to an osteogenic differentiation medium 24 h after transfection, with medium replacement every 3 days.
Cell counting kit-8 assay
Osteoblast precursor cells that were transfected with shGATA4 or shCTRL lentivirus were cultured in 96-well plates at a density of 1.0 × 103 cells/well in α-MEM with 10% FBS 24 h after transfection. For four subsequent days, a cell counting kit-8 (CCK8) solution (Dojindo, Tokyo, Japan) was added to each well, cultured for 30 min at 37 °C according to the manufacturer’s instructions, and then, the cell proliferation rate was quantified by a microplate reader at 450 nm.
Alkaline phosphatase (ALP) staining
Cells transfected with shCTRL or shGATA4 were seeded at a density of 5 × 104 cells/well in 12-well plates in osteogenic medium and ALP staining was conducted 5 days after inducing ossification with the BCIP/NBT Alkaline Phosphatase Color Development Kit (Beyotime Institute of Biotechnology, Shanghai, China) according to the manufacturer’s protocol.
Alizarin Red S (ARS) staining
ARS staining was undertaken 2 weeks after inducing ossification with 2% alizarin red (Beyotime Institute of Biotechnology) according to the manufacturer’s instructions to visualize mineralized nodules. Micrographs of these mineralized nodules were captured prior to dissolution of these nodules in each well by addition of 600 μL 10% cetylpyridinium chloride (CPC) at room temperature for 30 min. Then, 100 μL of the supernatant was transferred into 96-well plates and an automatic microplate reader was employed to measure absorbance at 562 nm.
Tooth movement model protocol
Induction of tooth movement (TM) was done as described in previous studies (Guo et al. 2016; Taddei et al. 2012). In brief, an orthodontic appliance consisting of a 0.25 × 0.76 mm Ni–Ti sentalloy closed coil spring (Grikin, Beijing, China) was applied to ligate the region between the upper incisors and upper left first molar using a dental adhesive resin (3M Unitek, Monrovia, USA), delivering a mesial direction force of 0.35 N. After 14 days of force application, the mice were sacrificed. Eight-week-old male mice were randomly divided into control (n = 5) and experimental (n = 5) groups.
Lentivirus injection
Mice in the experimental group received 5 μL concentrated lentiviral supernatant (shGATA4; 1 × 109 TU/mL) injected under the buccal periosteum of the left upper first molar in the TM model on days 3–10 after induction of tooth movement, whereas control group mice received the same dose of lentivirus delivering a nonspecific sequence (shCTRL).
Micro-CT analysis
To analyze the extent of bone formation, we separated the maxilla free from soft tissue, then fixed it in 70% ethanol overnight at 4 °C, and scanned it at a resolution of 9 μm/pixel with a micro-CT system (Skyscan 1176, Kontich, Belgium) at 50 kV and 456 μA. NRecon v1.6 and CTAn v1.13.8.1 were used to reconstruct and analyze the images (Zhou et al. 2016). The area of alveolar bone between the mesial and distal roots of the first maxillary molar in the TM model was defined as the region of interest (ROI). Six continuous images from this ROI were used for 3-dimensional reconstruction and analysis. Quantitative parameters analyzed were: trabecular bone volume over total volume (BV/TV), trabecular number (Tb.N), trabecular separation (Tb.Sp), and trabecular thickness (Tb.Th). The scan analysis operator was blinded to the treatments linked to each sample.
Histological analysis
Maxillary halves from the operated side of each animal were separated and fixed in 4% paraformaldehyde (PFA) for 24 h, decalcified with 10% EDTA (pH 7.4) for 3 weeks at 4 °C, embedded in paraffin, and cut into 5 μm-thick sections. The sections were stained with total collagen and Goldner’s trichrome, and stained sections were observed using light microscopy (Leica Microsystems Inc.) and photographed using Leica imaging software at ×200 the original magnification (Chen et al. 2014; Ma et al. 2013). Histological analysis of bone parameters were scaled by consecutively analyzing the images with Image J, as reported previously (Egan et al. 2012).
Immunofluorescence staining
Tissue sections of alveolar bone were stained with 4′,6-diamidino-2-phenylindole (DAPI) (Beyotime Institute of Biotechnology) at 1:1000 in PBS and then observed with fluorescence microscopy (Leica Microsystems Inc.) and analyzed using Leica imaging software at ×400 the original magnification.
Immunohistochemistry
Antigens from stained sections were retrieved by boiling in sodium citrate buffer solution and, then, endogenous peroxidase activity was quenched with 3% hydrogen peroxide (H2O2). After blocking with 1% goat serum for 30 min at 37 °C, sections were incubated with primary antibodies overnight at 4 °C (Sun et al. 2016). The following primary antibodies were employed: anti-GATA4, anti-osterix, anti-osteopontin, and anti-osteocalcin (ab84593, ab22552, ab63856, and ab93876, respectively, Abcam, Cambridge, UK). Antibodies were detected with goat anti-rabbit immunoglobulin G (IgG) and visualized by developing with a diaminobenzidine (DAB) substrate–chromogen system (Maxim, Fuzhou, China). Sections were observed at ×400 the original magnification. The percentage of positive cells compared to the total osteoblasts at the bone boundary or positive areas in each sample were counted using Image-Pro Plus 5.0 (Media Cybernetics, Silver Spring, USA) (Yu et al. 2014).
Western blotting
Total proteins obtained through cell lysis buffer, resolved by SDS–PAGE, and transferred onto polyvinylidene fluoride (PVDF) membranes. Methods of western blotting protocol used in this study were as described by Li et al. (2015). Antibodies against GATA4, OSX, OPN, and OCN (Abcam), and antibodies against p-p38, p38, p-ERK, ERK (Cell Signaling Technology, Boston, USA) and antibody against GAPDH (Bioworld, Nanjing, China) were used as primary antibodies. Semi-quantitative measurements were carried out using Image J.
Quantitative reverse-transcription PCR
Total RNA was isolated with a RNA isolation kit (Takara, Dalian, China) and a quantitative RT-PCR was carried out as described by Guo et al. (2016). Primers included: GAPDH 5′-ACCACAGTCCATGCCATCAC-3′ (forward) and 5′-TCCACCACCCTGTTGCTGTA-3′ (reverse); OSX 5′-GCTGGGTAGAGGAAGGTC-3′ (forward) and 5′-CAAAGAGAGAAGAACTGAGGAG-3′ (reverse); OPN 5′-ACCATGCAGAGAGCGAGGATT-3′ (forward) and 5′- GGGACATCGACTGTAGGGACG-3′ (reverse); and OCN 5′-ATCTTTCTGCTCACTCTGCT-3′ (forward) and 5′-CTACCTTATTGCCCTCCTG-3′ (reverse).
Statistical analysis
All values are displayed as mean ± SD. Experiments were conducted at least three times separately. Statistical analysis was conducted using Student’s t test (SPSS 19.0), and P values <0.05 were regarded as indicative of statistical significance.
Results
High-level expression of GATA4 in mouse osteoblast precursor cells
Osteoblast precursor cells from the second passage were characterized by FCM analysis. As displayed in Fig. 1a, osteoblast precursor cells showed positive expression of mesenchymal stem cell surface markers CD29 and CD105, whereas expression of hematopoietic stem cell markers CD34 and CD45 were absent, indicating that osteoblast precursor cells may be a stem cells population, which makes them suitable for studying osteoblast differentiation. To further determine the association between GATA4 and osteogenic differentiation, osteoblast precursor cells were cultured in osteogenic medium for 0, 3, 7, and 14 days. Then, western blotting was carried out to examine the time-dependent expression mode of GATA4 and osteoblast-specific marker proteins such as OSX, OPN, and OCN in mouse osteoblast precursor cells. As shown in Fig. 1b–f, expression of GATA4 and osteoblast-specific marker proteins transiently increased during osteoblast differentiation, implying GATA4 may be crucial for the regulation of osteoblast differentiation.

Characterization of osteoblast precursor cells and GATA4 expression in osteoblast precursor cells in mice. a Surface markers of osteoblast precursor cells were identified by flow cytometric analysis. Osteoblast precursor cells were stained with stem cell markers (CD29 and CD105) and hematopoietic markers (CD34 and CD45). b Western blotting results confirmed that mineralization-induced osteoblast precursor cells at all stages had higher GATA4, OSX, OPN and OCN expression than undifferentiated cells. c–f Western blotting bands from b were quantitatively calculated as the ratio of GATA4, OSX, OPN and OCN to GAPDH. *P < 0.05, **P < 0.01. All data are shown as mean ± SD
GATA4 knockdown inhibits osteoblast differentiation
To investigate the possible role of GATA4 in osteoblast differentiation, we knocked down the expression of endogenous GATA4 by lentivirus-mediated infection with a specific shRNA. Seventy-two hours after osteoblast precursor cells were transfected by lentiviral particles (MOI 50) in the presence of Polybrene, distinct green fluorescence expression was observed on fluorescence microscopy (Fig. 2a). Transfection efficiency was >80%. To confirm the effect of transfection of the shGATA4 lentiviral vector on GATA4 knockdown in osteoblast precursor cells, protein extraction was done on days 3 and 14 after transfection in normal media, followed by western blotting to quantify protein expression levels of GATA4 in osteoblast precursor cells. Western blotting analysis indicated that GATA4 expression was efficiently and stably knocked down in the experimental group, compared to the control group (Fig. 2b–e). To determine the effect of GATA4 on proliferative ability of osteoblast precursor cells, we quantified cell number with a CCK-8 kit. No significant differences were observed in the numbers of cells, indicating that GATA4 did not affect the proliferative ability of osteoblast precursor cells (Fig. 2f).

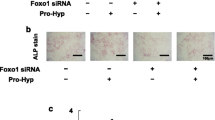
GATA4 knockdown reduced osteogenic differentiation of osteoblast precursor cells. a Transfection efficiency of osteoblast precursor cells was observed under fluorescence microscopy 72 h after lentiviral infection. The up row of the panel shows the bright field of the cells. Transfected osteoblast precursor cells displayed green fluorescence (low). Scale bar 50 μm. b, c GATA4 expression was tested by western blotting at 3 and 14 days after transfection, respectively. d, e Western blotting bands from b and c were quantitatively calculated as the ratio of GATA4 to GAPDH, respectively. f Cell proliferation of osteoblast precursor cells was analyzed at 0–4 days following transfection with lentivirus by a cell counting kit-8 (CCK-8) assay. g 5 days after mineralization, ALP staining was performed and observed with a scanner. Alizarin Red S staining was conducted 14 days after mineralization and then observed with a scanner and microscope. Scale bar 50 μm. h Calcium semiquantitative test was performed. *P < 0.05, **P < 0.01. All data are shown as mean ± SD
When osteoblast precursor cells were cultured with mineral medium for 5 days, ALP activity was examined by ALP kit, the shGATA4 transfected cells presented weaker ALP activity compared to the shCTRL group. In addition, ARS staining was conducted for 14 days after osteogenic induction, and it presented that the density of calcium nodes observably decreased in shGATA4 group (Fig. 2g). Meanwhile, the units of calcium reduced significantly, as indicated by the calcium semiquantitative test in GATA4 knocked-down osteoblast precursor cells (Fig. 2h). All of these findings suggest that GATA4 may be an essential factor for osteoblast differentiation.
Local silencing of GATA4 decreases bone formation during TM
To examine the role of GATA4 in bone formation in vivo, a TM animal model was chosen, and alveolar bone remodeling in shCTRL- and shGATA4-injected mice was observed. The selected shRNA vectors contained a GFP tag; thus, cells infected with the lentivirus would express GFP. Indeed, strong GFP expression was seen in experimental and control mice, whereas the groups not injected, displayed only background fluorescence, indicating that lentivirus was transfected as expected (Fig. 3a). Then, in order to visualize and verify the knockdown efficiency of GATA4 in vivo, immunohistochemistry of sections of alveolar bone using the rabbit polyclonal anti-GATA4 antibody was performed. As expected, GATA4 was abundantly available in the alveolar tissue between the roots of the first maxillary molars of shCTRL-injected mice at day 14. Conversely, much lesser GATA4-labeled cells were detected in shGATA4-injected mice at day 14 (Fig. 3b, c). Micro-CT carried out to quantitatively observe bone constitution and analyze correlative parameters of alveolar bone showed that, in the TM model, shGATA4-injected mice exhibited remarkable reduction of alveolar bone within the roots of the first upper molar at day 14, whereas there was smaller influence on the alveolar bone in the control group (Fig. 3d). Furthermore, shGATA4-injected mice showed decreased BV/TV (P < 0.01; Fig. 3e) and Tb.N (P < 0.05; Fig. 3f) values, and increased Tb.Sp value (P < 0.01; Fig. 3g). However, no significant difference of Tb.Th values was seen between experimental and control mice (Fig. 3h). Therefore, it can be speculated that GATA4 knockdown might play an inhibitory role in bone formation. To further explore our hypothesis, the sections were stained with total collagen or Goldner’s trichrome to examine histological differences between control and experimental groups and to determine whether GATA4 knockdown may influence bone remodeling. The results showed that bone formation within the area between the roots of the first maxillary molar was decreased significantly in the shGATA4-injected group compared with controls (Fig. 3i–l), which was consistent with results from the micro-CT analysis.

shRNA-mediated knockdown of GATA4 in vivo and decreased bone formation in shGATA4-injected mice. a Double-labeled fluorescent immunostaining of GFP (green, marker of transduction) in the first column, 4′,6-diamidino-2-phenylindole (DAPI)-stained cell nuclei (blue) in the second column, and merged images of the first and second column (the third column) in alveolar bone at day 14 after the operation for inducing tooth movement. Scale bar 50 μm. b GATA4-expressing cells determined by anti-GATA4 immunohistochemistry of alveolar bone at day 14 after TM. Scale bar 50 μm. c Percentages of GATA4-positive cells in experimental and control groups at day 14 after TM. d Micro-CT sagittal view images of alveolar bone at day 14 after TM. Red arrows indicate altered bone formation of alveolar bone. Scale bar 500 μm. e–h Quantitative analysis of bone volume over total volume (BV/TV), number (Tb.N), trabecular separation (Tb.Sp), and thickness (Tb.Th) of alveolar bone by micro-CT. i Representative photographs of alveolar sections stained histochemically for total collagen. Scale bar 100 μm. j Tissue sections were stained by Goldner’s trichrome staining. Mineralized matrix was stained green. Scale bar 100 μm. k, l Collagen area and mineralized bone area from sections of two different stains compared to total area, respectively. *P < 0.05, **P < 0.01. All data are shown as mean ± SD. (Color figure online)
GATA4 knockdown inhibits osteoblast-specific gene expression
Local knockdown of GATA4 in mice resulted in decreased expression level of osteogenic markers such as OSX, OPN, and OCN in osteoblasts on the bone surface on day 14 after the TM operation when tested by immunohistochemistry (Fig. 4a–f). Both shGATA4- and shCTRL-transfected cells were cultured in mineralization medium for 14 days. Expression levels of selected representative osteogenic markers were analyzed by Western blotting and quantitative RT-PCR. As expected, the expression levels of GATA4, OSX, OPN, and OCN were also reduced in shGATA4-transfected cells (Fig. 4g–n), in concordance with in vivo results.

Decreased expression of osteogenic markers in GATA4 knocked-down mice or cells compared with control group. a, c, e Immunohistochemically determined levels of representative osteogenic markers such as OSX, OPN, and OCN, were reduced in the alveolar bone on day 14 after the tooth movement operation in mice. Scale bar 50 μm. b, d Percentages of OSX- or OPN-positive cells in experimental and control groups on day 14 after TM were calculated, respectively. f Percentage of OCN-positive area was calculated in experimental and control groups on day 14 after TM. g Protein expression levels of GATA4 and several osteogenic markers (OSX, OPN, and OCN) in shGATA4-transfected osteoblast precursor cells were reduced as compared with control cells after 14 days following mineralization induction. h–k Western blotting bands from g were quantitatively calculated as the ratio of GATA4, OSX, OPN, and OCN to GAPDH, respectively. l–n 14 days after mineralization, expression of osteogenesis-related genes (OSX, OPN, and OCN) were tested by RT-PCR. *P < 0.05, **P < 0.01. All data are shown as mean ± SD
GATA4 knockdown decreases p38 activation
Osteogenic differentiation is regulated by different signaling pathways including MAPK. To elucidate whether GATA4 is involved in MAPK signaling pathway, we explored the reaction of the MAPK signaling pathway to osteogenic stimulation in control and shGATA4-transfected osteoblast precursor cells. The expression of phosphorylated p38 was markedly increased in control group undergoing osteogenic differentiation at day 7, whereas p38 phosphorylation in shGATA4-transfected cells were blunted compared to control group cells following induction of mineralization, although levels of phosphorylated ERK were unaffected (Fig. 5a–d). In addition, as shown in Fig. 5e–f, the expression of phosphorylated p38 transiently increased during osteoblast differentiation, indicating MAPK–p38 signaling may play a role in the regulation of osteoblast differentiation. Interestingly, the expression of GATA4 also showed an increasing tendency during osteoblast differentiation (Fig. 1b), consistent with the expression mode of phosphorylated p38, suggesting GATA4 may regulate osteoblastic differentiation and bone remodeling via p38-mediated signaling.

GATA4 knockdown decreases p38 activation. a Levels of GATA4, p-p38, p38, p-ERK, and ERK were detected by western blotting 7 days after induction of mineralization. b–d Quantification of western blotting experiments was displayed with the mean level of GATA4 over GAPDH, p-p38 over p38 and p-ERK over ERK in control and experimental groups. e Western blotting results indicated that mineralization-induced osteoblast precursor cells at all stages had higher phosphorylated p38 expression than undifferentiated cells. f Western blotting bands from e were quantitatively calculated as the ratio of p-p38 to p38.*P < 0.05, **P < 0.01. All data are shown as mean ± SD
Discussion
Osteoblast differentiation is essential for bone rebuilding, many transcription factors take part in the osteoblast differentiation process. GATA4, a transcription factor, has been reported to regulate transcription in many organs (Mazaud et al. 2007; Molkentin et al. 1998; Viger et al. 2008). However, the role of GATA4 in osteoblast differentiation is still unclear.
In the study, calvarial cells, which are considered osteoblast precursor cells, were employed for in vitro experiments and a TM in vivo model was established to observe the role of GATA4 in osteoblast differentiation. As GATA4 expression was up regulated during the osteoblast mineralization process as well as osteoblast-specific marker proteins such as OSX, OPN, and OCN, we proposed a hypothesis that GATA4 might be involved in osteoblastic differentiation. In order to confirm this hypothesis, we established GATA4 knockdown system to examine expression levels of osteogenesis-related genes, and found the expression of mineralization markers (OSX, OPN, and OCN) to be significantly decreased by GATA4 knockdown. Downregulation of GATA4 expression also weakened ALP activity, which is an early indicator of mineralization. Furthermore, findings from ARS staining and calcium semiquantitative analysis support the key role of GATA4 in osteoblastic differentiation.
In addition to the in vitro experiment, an in vivo experiment was conducted to explore the relation between GATA4 and osteoblast differentiation. Results of micro-CT and histological analysis showed clearly that bone formation in the experimental group was significantly decreased, compared to the control group. Meanwhile, expression of mineralization-related markers such as OSX, OPN, and OCN was also reduced, in alveolar bone on day 14 after the TM operation. Our in vivo data is consistent with results from the in vitro experiment, further confirming our assumption that GATA4 plays a critical role in osteoblast differentiation.
ALP and OSX are early-stage markers of osteoblast differentiation while OPN and OCN are closely related to the late-stage osteoblast differentiation (Li et al. 2016; Yusa et al. 2016). ALP is an established marker of osteoblast activity and a functional marker of bone formation (Mojarad et al. 2016). OSX plays a critical role in osteoblastic differentiation and is expressed in functional osteoblasts (Nakashima et al. 2002). OPN is an extracellular matrix protein expressed in many cell types especially osteoblasts, in which it plays multiple roles in diverse biological and pathological processes (Schlafer et al. 2012). OCN has close relations with bone maturation, which is identified as a late-stage marker of bone formation (Li et al. 2014a, b). However, the relationship between GATA4 and osteoblastic differentiation is still unknown. In this study, we demonstrated that GATA4 knockdown in osteoblast precursor cells significantly reduced the expression of both early and late stage osteogenic markers, implying the long-term effects of GATA4 on the differentiation of osteoblasts.
Recently, the role of GATA4 in osteoblast differentiation gained prominence when Miranda-Carboni et al. hypothesized that GATA4 is a possible E2 direct target in osteoblasts and could regulate osteoblast-specific genes by recruiting ERα to DNA in osteoblasts, Güemes et al. showed that GATA4, in osteoblasts, is essential for the survival and bone development, and GATA4 is a regulator of osteoblast commitment via E2-dependent and E2-independent pathways during early development (Miranda-Carboni et al. 2011; Güemes et al. 2014). However, Song et al. (2014) discovered that GATA4 may work to negatively modulate osteoblast differentiation by controlling Dlx5 binding ability to the runt-related transcription factor 2 (RUNX2) promoter. The signaling pathway involved in how GATA4 regulates osteoblast differentiation remains unclear. Increasing evidence suggests that MAPK pathways play a vital role in osteoblast differentiation. Some studies have proved ERK and p38 MAPKs are significant for early osteoblast differentiation and ERK or p38 inhibition suppresses osteoblast-specific gene expression (Suzuki et al. 2002; Xiao et al. 2000). Earlier literature suggested GATA4 as one of several p38 target proteins that may affect gonadal somatic cell development (Gierl et al. 2012). Overexpression of gelsolin could cause cardiac hypertrophy, which is due to the crosstalk of GATA4 and p38 signaling (Hu et al. 2014). Previous studies have also pointed out that the GATA4 protein has several possible MAPK phosphorylation sites within the N- and C-terminal domains, and p38 can directly interact with GATA4 within the N-terminal domain at several particular sites such as T100 and S105 (Charron et al. 2001). However, the specific interaction of GATA4 with MAPK signaling pathways during osteoblast differentiation has not been elucidated. In the study, incubating GATA4 knocked down cells in osteogenic-induced medium lead to decreased phosphorylation of p38, hence suggesting GATA4 downregulation may inhibit the sustained activation of p38, although levels of phosphorylated ERK were unaffected. In addition, the expression of phosphorylated p38 was upregulated during osteoblast differentiation, in agreement with the increase tendency of GATA4, indicating GATA4 may regulate osteoblastic differentiation and bone remodeling via p38-mediated signaling, and then loss-of-function of GATA4 in osteoblasts may influence osteoblast differentiation through downregulation of osteoblast-specific gene markers such as OSX, OPN, and OCN. Our study indicates that GATA4 can act as an activator in osteoblastic differentiation, consistent with results from Miranda-Carboni’s et al. (2011) and Güemes’ et al. (2014) study, and our data reveals that GATA4 plays a vital role in osteogenic differentiation and bone remodeling through p38-mediated signaling. Although MAPK activation often promotes cell proliferation, in the study downregulation of p-p38 in MAPK by GATA4 knockdown did not alter proliferation of osteoblasts without osteogenic induction. It is the first study elucidating a relationship between GATA4 and p38 signaling in osteoblast differentiation.
In summary, based on the presented results, it is reasonable to hypothesize that GATA4 may play a positive role in osteoblast differentiation, which may be affected through p38-mediated signaling. These findings improve our understanding of the regulatory mechanism of bone formation and may help to provide novel therapeutic approaches through regulation of osteoblast differentiation to increase bone formation. Thus, GATA4 may be an effective target to treat bone-related disorders. Therefore, detailed mechanisms underlying GATA4 gene regulation should be further elucidated to facilitate the development of additional therapeutic approaches in various bone diseases such as osteoporosis, hyperostosis, and periodontal disease.
Conclusion
The objective of this study was to evaluate the possible role of GATA4 in the differentiation of osteoblasts. Knockdown of GATA4 in calvarial cells and in the mouse TM model inhibited osteoblast differentiation, and decreased expression of bone-related genes both in vitro and in vivo, as these may be modulated via p38-mediated signaling. These results imply GATA4 is a vital regulator of osteoblast differentiation. Understanding the importance of the role of GATA4 in the differentiation of osteoblasts may facilitate the development of additional therapeutic approaches in various bone diseases.
References
Charron F, Tsimiklis G, Arcand M et al (2001) Tissue-specific GATA factors are transcriptional effectors of the small GTPase RhoA. Genes Dev 15:2702–2719
Chen W, Ma J, Zhu G et al (2014) Cbfβ deletion in mice recapitulates cleidocranial dysplasia and reveals multiple functions of Cbfβ required for skeletal development. Proc Natl Acad Sci USA 111:8482–8487. doi:10.1073/pnas.1310617111
de Oliveira PT, Zalzal SF, Irie K et al (2003) Early expression of bonematrix proteins in osteogenic cell cultures. J Histochem Cytochem 51:633–641
Egan KP, Brennan TA, Pignolo RJ (2012) Bone histomorphometry using free and commonly available software. Histopathology 61:1168–1173. doi:10.1111/j.1365-2559.2012.04333.x
Ge T, Yu Q, Liu W et al (2016) Characterization of bone marrow-derived mesenchymal stem cells from dimethyloxallyl glycine-preconditioned mice: evaluation of the feasibility of dimethyloxallyl glycine as a mobilization agent. Mol Med Rep 13:3498–3506. doi:10.3892/mmr.2016.4945
Gierl MS, Gruhn WH, von Seggern A et al (2012) GADD45G functions in male sex determination by promoting p38 signaling and Sry expression. Dev Cell 23:1032–1042. doi:10.1016/j.devcel.2012.09.014
Güemes M, Garcia AJ, Rigueur D et al (2014) GATA4 is essential for bone mineralization via ERα and TGFβ/BMP pathways. J Bone Miner Res 29:2676–2687. doi:10.1002/jbmr.2296
Guo S, Ni Y, Ben J et al (2016) Class A scavenger receptor exacerbates osteoclastogenesis by an interleukin-6-mediated mechanism through ERK and JNK signaling pathways. Int J Biol Sci 12:1155–1167
Hu WS, Ho TJ, Pai P et al (2014) Gelsolin (GSN) induces cardiomyocyte hypertrophy and BNP expression via p38 signaling and GATA-4 transcriptional factor activation. Mol Cell Biochem 390:263–270. doi:10.1007/s11010-014-1977-7
Li Y, Yan M, Wang Z et al (2014a) 17beta-estradiol promotes the odonto/osteogenic differentiation of stem cells from apical papilla via mitogen-activated protein kinase pathway. Stem Cell Res Ther 5:125. doi:10.1186/scrt515
Li J, Yan M, Wang Z et al (2014b) Effects of canonical NF-κB signaling pathway on the proliferation and odonto/osteogenic differentiation of human stem cells from apical papilla. Biomed Res Int 2014:319651. doi:10.1155/2014/319651
Li F, Zhao ZH, Cai ZJ et al (2015) Oxidized Low-density lipoprotein promotes osteoblastic differentiation of valvular interstitial cells through RAGE/MAPK. Cardiology 130:55–61. doi:10.1159/000369126
Li M, He P, Wu Y et al (2016) Stimulatory effects of the degradation products from Mg-Ca-Sr alloy on the osteogenesis through regulating ERK signaling pathway. Sci Rep 6:32323. doi:10.1038/srep32323
Ma J, Chen W, Zhang L et al (2013) RNA interference-mediated silencing of Atp6i prevents both periapical bone erosion and inflammation in the mouse model of endodontic disease. Infect Immun 81:1021–1030
Maranga L, Cunha A, Clemente J et al (2004) Scale-up of virus-like particles production: effects of sparging, agitation and bioreactor scale on cell growth, infection kinetics productivity. J Biotechnol 107:55–64
Mazaud GS, Tetu A, Legault E et al (2007) The proximal Gata4 promoter directs reporter gene expression to sertoli cells during mouse gonadal development. Biol Reprod 76:85–95
Miranda-Carboni GA, Güemes M, Bailey S et al (2011) GATA4 regulates estrogen receptor-alpha-mediated osteoblast transcription. Mol Endocrinol 25:1126–1136. doi:10.1210/me.2010-0463
Mojarad F, Amiri I, Rafatjou R et al (2016) The effect of 1α,25(OH)2D3 on osteogenic differentiation of stem cells from dental pulp of exfoliated deciduous teeth. J Dent (Shiraz) 17:348–353
Molkentin JD, Lu JR, Antos CL et al (1998) A calcineurin-dependent transcriptional pathway for cardiac hypertrophy. Cell 93:215–228
Nakashima K, Zhou X, Kunkel G et al (2002) The novel zinc fingercontaining transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 108:17–29
Pittenger MF (2008) Mesenchymal stem cells from adult bone marrow. Methods Mol Biol 449:27–44. doi:10.1007/978-1-60327-169-1_2
Pittenger MF, Mackay AM, Beck SC et al (1999) Multilineage potential of adult human mesenchymal stem cells. Science 284:143–147
Rodriguez-Lozano FJ, Bueno C, Insausti CL et al (2011) Mesenchymal stem cells derived from dental tissues. Int Endod J 44:800–806. doi:10.1111/j.1365-2591.2011.01877.x
Schlafer S, Raarup MK, Wejse PL et al (2012) Osteopontin reduces biofilm formation in a multi-species model of dental biofilm. PLoS ONE 7:e41534. doi:10.1371/journal.pone.0041534
Shoji S, Tabuchi M, Miyazawa K et al (2010) Bisphosphonate inhibits bone turnover in OPG(−/−) mice via a depressive effect on both osteoclasts and osteoblasts. Calcif Tissue Int 87:181–192. doi:10.1007/s00223-010-9384-x
Song I, Kim K, Kim JH et al (2014) GATA4 negatively regulates osteoblast differentiation by downregulation of Runx2. BMB Rep 47:463–468
Sun J, Sun B, Wang W et al (2016) Histochemical examination of the effects of high-dose 1,25(OH)2D3 on bone remodeling in young growing rats. J Mol Histol 47:389–399. doi:10.1007/s10735-016-9681-4
Suzuki A, Guicheux J, Palmer G et al (2002) Evidence for a role of p38 MAP kinase in expression of alkaline phosphatase during osteoblastic cell differentiation. Bone 30:91–98
Taddei SR, Moura AP, Andrade I Jr et al (2012) Experimental model of tooth movement in mice: a standardized protocol for studying bone remodeling under compression and tensile strains. J Biomech 45:2729–2735. doi:10.1016/j.jbiomech.2012.09.006
Teitelbaum SL (2000) Bone resorption by osteoclasts. Science 289:1504–1508
Tripuwabhrut P, Mustafa M, Gjerde CG et al (2013) Effect of compressive force on human osteoblast-like cells and bone remodelling: an in vitro study. Arch Oral Biol 58:826–836. doi:10.1016/j.archoralbio.2013.01.004
van Berlo JH, Elrod JW, Aronow BJ et al (2011) Serine 105 phosphorylation of transcription factor GATA4 is necessary for stress-induced cardiac hypertrophy in vivo. Proc Natl Acad Sci USA 108:12331–12336. doi:10.1073/pnas.1104499108
Viger RS, Guittot SM, Anttonen M et al (2008) Role of the GATA family of transcription factors in endocrine development, function, and disease. Mol Endocrinol 22:781–798. doi:10.1210/me.2007-0513
Wu Y, Zhou J, Li Y et al (2015) Rap1A regulates osteoblastic differentiation via the ERK and p38 mediated signaling. PLoS ONE 10:e0143777. doi:10.1371/journal.pone.0143777
Xiao G, Jiang D, Thomas P et al (2000) MAPK pathways activate and phosphorylate the osteoblast-specific transcription factor, Cbfa1. J Biol Chem 275:4453–4459
Yokoya K, Sasaki T, Shibasaki Y (1997) Distributional changes of osteoclasts and pre-osteoclastic cells in periodontal tissues during experimental tooth movement as revealed by quantitative immunohistochemistry of H(+)-ATPase. J Dent Res 76:580–587
Yu J, Cao H, Yang J et al (2012) In vivo hepatic differentiation of mesenchymal stem cells from human umbilical cord blood after transplantation into mice with liver injury. Biochem Biophys Res Commun 422:539–545. doi:10.1016/j.bbrc.2012.04.156
Yu Y, Mu J, Fan Z et al (2014) Insulin-like growth factor 1 enhances the proliferation and osteogenic differentiation of human periodontal ligament stem cells via ERK and JNK MAPK pathways. Histochem Cell Biol 137:513–525. doi:10.1007/s00418-011-0908-x
Yusa K, Yamamoto O, Takano H et al (2016) Zinc-modified titanium surface enhances osteoblast differentiation of dental pulp stem cells in vitro. Sci Rep 6:29462. doi:10.1038/srep29462
Zhong ZA, Ethen NJ, Williams BO. (2016) Use of primary calvarial osteoblasts to evaluate the function of Wnt signaling in osteogenesis. Methods Mol Biol 1481:119–125. doi:10.1007/978-1-4939-6393-5$413
Zhou FH, Foster BK, Zhou XF et al (2006) TNF-alpha mediates p38 MAP kinase activation and negatively regulates bone formation at the injured growth plate in rats. J Bone Miner Res 21:1075–1088
Zhou Z, Yin Y, Jiang F et al (2016) CBX7 deficiency plays a positive role in dentin and alveolar bone development. J Mol Histol 47:401–411. doi:10.1007/s10735-016-9682-3
Acknowledgements
This work was supported by the National Natural Science Foundation of China (81371179), Natural Science Foundation of Jiangsu Province (BK20150048), and project funding by the Priority Academic Program Development of Jiangsu Higher Education Institutions (2014-037).
Author contributions
Conception and experimental design: JM, LW, and SG. Performing of experiments: TZ, SG, YZ. Data analysis: JM, TZ, SG and YZ. Contribution of reagents/materials/analysis tools: JM, TZ, SG, YZ and LW. Manuscript writing: JM, TZ, SG and YZ.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest with the contents of this article.
Additional information
Tingting Zhou, Shuyu Guo and Yuxin Zhang have contributed equally to this work.
Rights and permissions
About this article

Cite this article
Zhou, T., Guo, S., Zhang, Y. et al. GATA4 regulates osteoblastic differentiation and bone remodeling via p38-mediated signaling. J Mol Hist 48, 187–197 (2017). https://doi.org/10.1007/s10735-017-9719-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10735-017-9719-2