
Abstract
Eight species of the genus Asparagus, members of the group of European species closely related to A. officinalis, were analysed using internal transcribed spacer (ITS), and expressed sequence tag-derived simple sequence repeat (EST-SSR) markers, as well as cytological observations of their hybrids, to study their phylogenetic relationships and the possibility of broadening the narrow genetic base of cultivated varieties. Phylogenetic analysis using ITS data revealed two major clades: clade I consisting of A. acutifolius and clade II (referred to in this study as the ‘officinalis group’) comprised of sequences derived from species closely related to A. officinalis; but the different species within the ‘officinalis group’ could not be clearly separated. In contrast, cluster analysis of EST-SSR marker data showed six major clades and clearly separated each population, grouping most of the genotypes from each population together. That is, EST-SSR markers were found to be more informative than ITS markers about the relationships within the ‘officinalis group’, indicating that EST-SSR markers are more useful than ITS sequences for establishing phylogenetic relationships at the intrageneric level. All the crosses carried out at the same ploidy level were successful. The high crossability, together with the regular meiotic behaviour and high pollen and seed fertility observed in the interspecific hybrids analysed, suggest relatively close relationships between the species studied. We conclude that the group of species classified in the ‘officinalis group’ are in the primary gene pool, indicating that these species could be used to increase the genetic diversity of the cultivated species. In addition, the tetraploid landrace “Morado de Huétor” could be employed as a bridge to generate new cultivated germplasm.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The genus Asparagus comprises three subgenera Asparagus, Protasparagus and Myrsiphyllum (Clifford and Conran 1987) which include 100–300 species distributed in the arid and subarid regions of Europe, Asia, Africa and Australia (Kubota et al. 2012). The species of the first subgenera are dioecious, with unisexual flowers, whereas the second and the third subgenera include only hermaphroditic plants. It has a remarkably consistent basic chromosome number of x = 10, across species of different ploidy levels. Polyploidization seems to be common in asparagus, favoured by its perennial nature, and may have played an important role in the evolution of this genus.
Asparagus officinalis L. (2n = 2x = 10) is the most economically important Asparagus species and the only one cultivated worldwide as a vegetable crop. Current cultivars of this species have a narrow genetic base probably as a consequence of their common origin from the Dutch population ‘Violet Dutch’ (s XVIII) (Knaflewsky 1996). Hence, it would be interesting to introgress agronomically important traits from wild relatives thereby enlarging the gene pool for breeding. Within the Asparagus subgenera there is a group of wild growing Euroasiatic species closely related to A. officinalis that could be used for this purpose. The European species include A. prostratus Dumort., A. maritimus (L.) Mill., A. pseudoscaber Grecescu, A. brachyphyllus Turcz., A. kasakstanicus Iljin and A. tenuifolius Lam. (Komarov et al. 1935; Valdes 1980). In addition, there are other European wild species (A. acutifolius L., A. horridus L., A. aphyllus L. and A. albus L.), distantly related to the cultivated one, that are distributed across the Mediterranean area. These species are important because their young spears are collected for human consumption in Mediterranean countries where they can even be found in some local markets (Pieroni 2005). All of them belong to the Asparagus subgenus except A. albus that is in the subgenus Protasparagus. Also from the Protasparagus subgenus, there is A. densiflorus (Kunth) Jessop, a southern African ornamental species that is highly resistant to Fusarium spp. (Ellison 1986; Marcellan and Camadro 1996; Stephens et al. 1989), one of the major limiting factors in asparagus production worldwide. Resistance to other important diseases in cultivated asparagus such as rust (Puccinia asparagi DC.) and purple spot [Stemphylium vesicarium (Wallr.) E. Simmons (teleomorph Pleospora herbarum)] has also been reported. For example, A. maritimus was found to be resistant to rust while resistance to both rust and purple spot diseases was observed in A. acutifolius, A. albus, A. horridus and A. aphyllus (Alberti et al. 2004; Falavigna et al. 2008). Furthermore, tolerance to various types of abiotic stress such as salinity, drought and soil acidity has been observed in A. maritimus, A. acutifolius and A. tenuifolius, respectively (Venezia et al. 1993). To date, few wild populations have been evaluated and it will be necessary to explore a wide range in order to identify agronomically important traits.
There are not many reports of crosses between A. officinalis and its wild relatives, or of their use in asparagus breeding. Exceptions include successful crosses between A. officinalis and the wild species A. tenuifolius (Bozzini 1963), A. prostratus (McCollum 1988) and A. maritimus (Alberti et al. 2004; Falavigna et al. 2008). Falavigna et al. (2008) reported a F1 plant derived from the cross between a tetraploid hybrid plant (“Violetto d’Albenga” × A. maritimus) and A. acutifolius. This hybrid plant showed a high level of sterility and is resistant to rust and purple spot. It was backcrossed with a tetraploid A. officinalis plant and eight BC1 plants were obtained (Falavigna et al. 2008). Marcellan and Camadro (1999) tried to generate viable A. officinalis and A. densiflorus cv. ‘Sprengeri’ interspecific hybrids in order to introduce resistance to Fusarium from the latter species. These attempts were, however, unsuccessful due to the failure of endosperm development. According to the classification proposed by Harlan and de Wet (1971) for cultivated species, the group of species that are close to A. officinalis are likely to be in the primary gene pool and the remaining wild species members of the secondary and tertiary gene pools. Nevertheless, more studies are necessary to determine the gene pool of each species. Specifically, more data are needed on the taxonomic relationships within the genus Asparagus to guide the choice of wild germplasm for increasing the genetic basis of cultivated asparagus.
Studies of phylogenetic relationships within the genus Asparagus may also be useful to investigate evolution in this genus. Robust DNA molecular tools for analysis of both chloroplast DNA (cpDNA) and nuclear ribosomal DNA (nrDNA) sequences are widely used in systematic plant molecular studies to infer phylogenetic relationships (Peterson et al. 2004). Nuclear ribosomal DNA is organised into one or several chromosomal loci and consists of tandem repeats of three ribosomal DNA genes, and intergenic and internal transcribed spacers. Internal transcribed spacers (ITS) are conserved regions, which are very variable, and hence tend to be useful for obtaining phylogenetic information in closely related taxa (Baldwin et al. 1995). In Asparagus, phylogenetic studies have been performed on the basis of restriction fragment length polymorphism (RFLP) analysis of nrDNA by Štajner et al. (2002). These authors used only two species closely related to A. officinalis (A. tenuifolius and A. maritimus) in their study and considered only a small number of characteristics; indeed, this is a limitation of restriction fragment analysis. Similarly, in our group, RFLP analysis of nrDNA (ITS1-5.8S-ITS2) was applied to study the phylogeny of a Spanish cultivated tetraploid landrace and it was found to be a hybrid between A. officinalis and A. maritimus (Moreno et al. 2008). Fukuda et al. (2005) carried out a wider study using cpDNA sequences applied to a larger number of species that gave a complete picture of the taxonomy and evolution of the genus Asparagus considering the three subgenera Asparagus, Protasparagus and Myrsiphyllum.
Simple sequence repeats (SSRs) or microsatellites markers derived from genomic DNA are widely used in cultivar fingerprinting, genetic diversity assessment, molecular mapping, and marker-assisted breeding. High rates of transferability of genomic-SSR loci across species within a genus have been reported (Eujayl et al. 2004; Thiel et al. 2003). However, the transferability of genomic-SSR loci across genera appears to be low (Roa et al. 2000; Thiel et al. 2003). On the other hand, expressed sequence tag-derived SSRs (EST-SSRs), derived from the transcriptome, represent the most conserved portion of the genome and are therefore more transferable between species than genomic-SSR markers (Scott et al. 2000). Hence, EST-SSR markers are particularly useful for cross-species genetic studies (Eujayl et al. 2004; Thiel et al. 2003). Specifically in asparagus, EST-SSR markers were developed and used by Caruso et al. (2008) to study the genetic relationships within currently cultivated varieties of asparagus.
The aim of the current study was to explore the phylogenetic relationships within the European group of species closely related to A. officinalis by alignments of DNA sequences from the ITS1-5.8S-ITS2 region and EST-SSR markers. In addition, to evaluate the possibility of transferring genes from wild to cultivated species and their potential exploitation in breeding programmes, we also assessed the crossability between the different asparagus species.
Materials and methods
Plant material
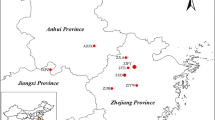
A total of 15 individuals from eight species of Asparagus and both cultivated and wild landraces were used in this study (Table 1). These individuals included diploid and polyploid species, while the individuals of A. officinalis included both current cultivars and wild populations.
Isolation of genomic DNA
Total genomic DNA of each plant was extracted from 1 g of tips from young spears following a modified CTAB extraction protocol described by Torres et al. (1993). The quality and concentration of extracted DNA was measured by using a NanoDrop ND-1000 spectrophotometer (Thermo Scientific, Waltham, MA).
Amplification of the rDNA ITS region
The region including the two ITSs (ITS-1 and ITS-2) and the 5.8 S subunit of nuclear ribosomal DNA were amplified using the universal primers ITSL (5′-TCGTAACAAGGTTTCCGTAGGTG-3′) and ITS4 (5′- TCCTCCGCTTATTGATATGC-3′) reported by Štajner et al. (2002). Amplification was carried out as in Štajner et al. (2002). PCR products were separated by gel electrophoresis on gels composed of a mixture of 1 % Seakem agarose and 1 % NuSieve agarose in 1× TBE buffer and stained with ethidium bromide.
Cloning, sequencing and phylogenetic analysis of ITS regions
PCR products containing the desired band corresponding to the ITS region (ITS1-5.8S-ITS2) were purified using a SureClean kit (Bioline, UK). Purified products were cloned into the pGEM-T vector system (Promega, Madison, WI) and transformed into competent Escherichia coli JM109 cells, in accordance with the manufacturer’s protocol. Variation in this region within individuals was found using RFLP analysis in previous studies (Moreno et al. 2008) and, therefore, at least two clones representative of each accession were selected for sequencing. Clones were sequenced in an automated ABI Prism sequencer (Applied Biosystems, Madrid, Spain) in the Genomics Unit of the Central Research Support Service at the University of Córdoba (Spain).
Sequence editing and alignment were conducted using Genious software (version 5.0.3, Drummond et al. 2010). Phylogenetic analysis of the ITS aligned sequence data was performed by parsimony analysis using PHYLIP software (version 3.69, Felsenstein 1993) and a sequence of A. albus was used as the out-group. Confidence in resulting nodes was assessed using the bootstrap procedure (Felsenstein 1985) with 100 replicates.
Amplification of EST-SSR markers and phylogenetic analysis
Around ten genotypes from each accession of species related to A. officinalis were analysed using EST-SSR markers, except in the case of A. tenuifolius for which only two plants were available. A set of four EST-SSR markers (AG8, TC1, TC3 and TC7) previously developed by Caruso et al. (2008) was employed in this study. Forward primers were synthesized with fluorescent dyes 6FAM or HEX (Applied Biosystems) at the 5′ ends. Amplification of these markers was performed as in Caruso et al. (2008). The PCR products were separated using an automated capillary sequencer (ABI 3130 Genetic Analyzer; Applied Biosystems/HITACHI, Madrid, Spain) in the Unit of Genomics of the Central Research Support Service at the University of Córdoba. The size of the amplified bands was calculated based on an internal DNA standard (400HD-ROX) with GeneScan software (version 3.x) and the results were interpreted using the Genotyper program (version 3.7) all from Applied Biosystems.
Due to the polyploid nature of many accessions, for the phylogenetic study, alleles were scored as present (1) or absent (0) for each marker and a binary data matrix was created. The estimation of the genetic similarity and distance between polyploids organisms remains problematic. However, Bruvo et al. (2004) developed a measure of genetic distance for polyploids that is similar to band-sharing indices, but takes the mutational distance between alleles into account. The Bruvo distance was calculated for this study using the POLYSAT package of Clark and Jasieniuk (2011). The species studied were all members of the Asparagus genus, hence the assumption of a molecular clock was acceptable. Therefore, grouping of the genotypes was determined by using the unweighted pair group method with arithmetic mean (UPGMA) and a dendrogram was produced using the PHYLIP software (Felsenstein 1993).
Production of interspecific hybrids
In order to assess the crossability between the asparagus species, crosses between cultivated asparagus and its wild relatives were performed. Crosses were made at the same ploidy level. Tetraploid and hexaploid plants from “Morado de Huétor” landrace were used in the crosses (Moreno et al. 2006). The ploidy level was determined by estimating the relative DNA content by flow cytometry (Ploidy Analyser PA-I; Partec, Münster, Germany). The analysis was carried out as described in Moreno et al. (2008).
To produce F1 hybrids, all female plants chosen as pollen recipients were bagged to prevent pollination from unknown pollen sources. During spring, fresh pollen from the male parents was collected and employed for hand pollination of females. The viability of each cross was calculated as the number of the seeds obtained relative to the number of pollinations performed. The hybrid plants and their parental species were grown in a field plot at the University of Córdoba (Spain).
Cytological observations
Meiotic analysis and pollen fertility studies were carried out in all parental species and six interspecific F1 hybrids. For meiotic studies, flower buds with pollen mother cells at meiotic stages were fixed in a freshly prepared 3 parts absolute ethanol to 1 part glacial acetic acid (v/v) mixture and stored at 4 °C. After fixation, the buds were treated in 2 N HCl for 5 min at room temperature and stained with 45 % acetocarmine for 10 min. The anthers were then squashed in 45 % acetocarmine stain solution. Around 100 cells in Anaphase I from one plant of each parental species and their interspecific F1 hybrids were analysed for chromosome distribution. Pollen stainability was used to indicate pollen fertility and was estimated using fresh pollen grains sampled from mature anthers at flowering. Over 100 pollen grains from each of three flowers per parent and hybrids were analysed. Pollen grains were treated with 45 % acetocarmine for several seconds in order to visualize the viable and non-viable grains. Darkly stained pollen grains were considered to be fertile and viable, and unstained or very lightly stained ones sterile or non-viable. Pollen fertility was then scored as the percentage of viable stained pollen grains with respect to the total number of grains counted for each flower. One-way analysis of variance was applied to determine whether the fertility differed significantly between the accessions studied.
Results
Alignment of ITS region and phylogenetic analysis
The primer pair ITSL-ITS4 amplified a fragment of around 750 bp in all 15 Asparagus accessions. Positive clones of a 0.75 kb fragment covering the ITS region were sequenced and aligned, starting with ITS1. At least two clones from each of the 15 accessions used in this study were completely sequenced in both directions. This amounted to a total of 23 sequences (Table 2). The lengths of ITS1, 5.8S and ITS2 regions varied little in the Asparagus species. Sequences of 756, 757 and 760 bp length were found, 756 bp being the most frequently observed in most of the species. The analysis of the sequence alignments across the accessions identified 118 single nucleotide polymorphisms. These alignments are available on request from the authors. The number of sequences per accession ranged from 1 (A. albus and A. maritimus from Cartagena) to 3 (“Violetto d’Albenga”) and some sequences were detected in two or three species (Table 2).
A parsimony analysis with A. albus as the out-group resulted in 100 equally parsimonious trees. The consensus phylogenetic tree showed two major clades (I and II) that are supported by bootstrap values of 100 and 82 %, respectively (Fig. 1). Clade I is composed of two sequences of A. acutifolius and clade II includes sequences derived from the species closely related to A. officinalis. We propose to name this second clade the ‘officinalis group’. In turn, clade II can be divided into three subgroups (IIa, IIb and IIc) supported by bootstrap values of 26, 14 and 72 %, respectively. It is not, however, possible to associate these subgroups with specific species, A. maritimus and “Morado de Huétor” being present in all three, A. brachyphyllus in two (IIb and IIc), and the other species studied (A. officinalis, A. prostratus, A pseudoscaber, A. tenuifolius and “Violetto d’Albenga”) in only one (Fig. 1). These results indicate that A. acutifolius is more distant to the other species, but the different species of the ‘officinalis group’ could not be clearly separated by using ITS region sequences.

Unrooted consensus parsimony tree of Asparagus species based on ITS region sequence alignment. Numbers S1-S22 are the names of the different sequences employed in this study as listed in Table 2. Asparagus albus was used as outgroup. Numbers at the nodes represent bootstrap values (%) based on 100 replicates
Phylogenetic analysis using EST-SSR markers
In order to clarify the relationships within the ‘officinalis group’, EST-SSR markers were used. In this case, A. acutifolius was not employed because in the ITS analysis was clearly separate from ‘officinalis group’. A total of 143 different alleles were detected with the four EST-SSR markers across all 102 genotypes representing seven species of Asparagus and the “Morado de Huétor” landrace population studied. The number of alleles per EST-SSR locus ranged from 30 to 43, with an average of 35.7 alleles per marker. The primer pair TC3 amplified the most alleles and their sizes ranged from 133 to 195 bp, while the fewest alleles were observed at both TC1 and TC7 loci, with sizes ranging from 199 to 251 bp and from 190 to 245, respectively. All the markers, except TC7 that did not work in A. albus, amplified in all seven species with different levels of ploidy (Table 3). These results demonstrate the transferability of these EST-SSRs between Asparagus species. There was variation among species in the number of alleles per primer pair. One or two alleles were detected in each of the diploid species (A. officinalis, A. albus and A. tenuifolius), indicating in all cases the amplification of a single locus. In the polyploid species, the number of alleles per marker in each individual varied, depending on their ploidy level. For example, in hexaploid species there were one to six alleles per marker, while up to 11 alleles were identified with the marker TC1 in an individual of the dodecaploid A. maritimus population.
The relationships among the species were explored by cluster analysis. The tree based on Bruvo distances separated all populations except “Morado de Huétor” and A. maritimus (Cartagena) that were split into two different groups. Most of the genotypes were also clustered together by their species of origin (Fig. 2). Six major clusters were defined. The first one is composed of diploid A. tenuifolius. The second main cluster contains the diploid A. officinalis, both wild and cultivated, and tetraploid A. postratus. The third cluster is composed of tetraploid landrace “Morado de Huétor” and hexaploid A. maritimus populations from Albania, Venice (Italy), Padova (Italy) and Cartagena (Spain). Most of the individuals from the dodecaploid A. maritimus population from Cartagena (Spain) are differentiated from these three populations, despite being a member of the same species. The fourth cluster contains just hexaploid A. brachyphyllus. The fifth cluster contains dodecaploid A. maritimus from Cartagena and the tetraploid landrace “Morado de Huétor”. Finally, hexaploid A. pseudoscaber formed a separate cluster (VI). The Bruvo distance coefficient mean within populations ranged from 0.63 for “Morado de Huétor” to 0.47 for A. pseudoscaber indicating a greater diversity in this landrace.

Consensus dendrogram of 102 Asparagus individuals obtained using 4 EST-SSR markers. The accessions codes are: AMc, A. maritimus (from Cartagena); AMp, A. maritimus (from Padova); AMv, A. maritimus (from Venice); AMa, A. maritimus (from Albania); APS, A. pseudoscaber; AB, A. brachyphyllus; APb, A. prostratus (from Bares); APo, A. prostratus (from Ortigueira); AOw, A. officinalis (wild); AOc, A. officinalis (cultivated); HT, “Morado de Huétor”; AT, A. tenuifolius; and AA, A. albus. Numbers in brackets indicate the number of individuals grouped in each branch
Production of interspecific hybrids
All the crosses carried out at the same ploidy level between the species employed in this study were successful. The viability of each cross is shown in Table 4.
Cytological observations
From the seeds obtained in the crosses, around 10 F1 plants were developed and their hybrid nature was confirmed employing EST-SSR. Specific alleles from both parental species were found together in the hybrids (data not shown).
Studies on meiotic chromosome segregation were carried out in a male plant of each interspecific F1 hybrids along with their parental species. The frequencies of anaphase I cells showing regular chromosome disjunction and of cells with laggards are presented in Table 3. In the majority of the pollen mother cells evaluated the chromosomes were located at the poles indicating that chromosome disjunction at anaphase I was essentially normal in all the species studied. Nevertheless, in all the genotypes analysed, except in A. officinalis and “Morado de Huétor” (6x) both with 0 %, a low frequency of cells with laggards were also observed. In general, all the interspecific hybrids had a slightly higher percentage of laggards than their respective parental species (Table 4).
Pollen fertility results from the parental genotypes and their interspecific F1 hybrids are presented in Table 4. A high percentage of stained pollen (>90.26 %) was observed for all parental species. These data also reflect the meiotic regularity. The high pollen viability indicates that the irregularities observed at meiosis are not significant in terms of species fertility. All the interspecific hybrids analysed also showed high pollen fertility and uniform pollen grains. Average pollen fertility ranged from 85.17 ± 2.08 % for the hybrid derived from “Morado de Huétor” (6x) × A. maritimus to 96.66 ± 1.13 % for the hybrid derived from A. maritimus × A. brachyphyllus. There were no significant differences in the frequency of viable pollen among genotypes, or between the interspecific F1 hybrids and their parental lines. All interspecific F1 hybrids are currently growing under field conditions in Córdoba and good fruit development was observed in female F1 hybrids.
Discussion
It is commonly accepted that the centre of origin of cultivated asparagus or garden asparagus (A. officinalis) is eastern Europe or western Asia, probably in the area of the Caucasus (Ellison 1986; Geoffriau et al. 1992; Sturtevant 1890). On the other hand, this species is widely naturalised as an escape from cultivation and its native distribution is uncertain (Kay et al. 2001). Its current native distribution may be related to its aforementioned region of origin. Geoffriau et al. (1992) and Sarabi et al. (2010) found current cultivars to be well differentiated from wild populations of A. officinalis from Turkey and Iran, at the isoenzymatic level and in terms of RAPD markers, respectively. The other European wild species studied, which are closely related to A. officinalis and which have different ploidy levels, have different distributions. A. prostratus (4x) is distributed along the Atlantic coast from the north of Spain to the south-east of Ireland and north-west of Germany, while A. maritimus (4x, 6x, 12x) has been reported to be growing around the Mediterranean coast (Morocco, Spain, France, Italy, Albania) (Valdes 1975). On the other hand, A. pseudoscaber (6x) has been found mainly in the Danube valley and A. tenuifolius has a wide distribution to the north of the Mediterranean Sea and in central Europe. Lastly, A. kasakstanicus (of unknown ploidy level) and A. brachyphyllus (4x, 6x) are both widely distributed in Asia and present on the northern coast of the Black Sea (Komarov et al. 1935; Valdes 1975). Polyploid wild species could have evolved (1) from a diploid species by autopolyploidy or (2) from hybridisation between different species by allopolyploidy. Unreduced gametes could have played an important role in this process; tetraploid asparagus plants have been found in fields of diploid garden asparagus through the formation of unreduced gametes (Marcellan and Camadro 1996). Unreduced gametes could also explain the presence of some hexaploid and octoploid plants in the tetraploid landrace “Morado de Huétor” (Moreno et al. 2006).
Nuclear ribosomal DNA sequences are widely used in plant molecular systematic studies to infer phylogenetic relationships (Peterson et al. 2004). The ITS1-5.8S-ITS2 sequence of nrDNA used in this study showed polymorphism both within and among the Asparagus species studied. However, only the sequences of A. acutifolius were clearly separated from the sequences of the group of species closely related to cultivated asparagus (the ‘officinalis group’), among which it was not possible to infer phylogenetic relationships. Despite the near-universal usage of ITS sequence data in plant phylogenetic studies, the complex and unpredictable evolutionary changes in this type of sequence may reduce their usefulness for phylogenetic analysis because the assumption of orthology could be affected (Álvarez and Wendel 2003). Specifically, in this study, within-genome variability for ITS sequences was found and it may be related to different NOR (nucleolus organizer regions) loci. Three pairs of NORs have been reported in diploid cultivated asparagus (De Melo and Guerra 2001; Reamon-Büttner et al. 1999). This could affect the criterion for phylogenetic inference that the sequences compared should be orthologous. It has been suggested that clearer insights are likely to emerge from the use of single-copy or low-copy nuclear genes (Álvarez and Wendel 2003).
Phylogenetic relationships at intraspecific level or between closely related species can be inferred using SSRs (Simpson 2006). In our study, EST-SSR markers were more informative than ITS sequences concerning the relationships within the ‘officinalis group’. The diploid wild species A. tenuifolius separated early from the remaining species of the group, suggesting that polyploid species may have arisen from A. officinalis or from a common diploid ancestor. In a previous study, the nuclear DNA content (1C) from A. tenuifolius (1.71 pg) was found to be higher than that for both A. officinalis (1.39–1.46 pg) and A. maritimus (1.28–1.31 pg) (Štajner et al. 2002). According to our results, the crossability among the species grouped into the ‘officinalis group’ and both the regular meiosis and high pollen fertility of the interspecific F1 hybrids indicate a relatively close genetic relationship between them. This result suggests a recent speciation. With respect to A. acutifolius, it was not included in the ‘officinalis group’ in this study. Incompatibility crosses with both A. officinalis and A. maritimus have been reported (Falavigna et al. 2008). However, this incompatibility barrier was overcome when a tetraploid hybrid of A. officinalis × A. maritimus was employed obtaining a hybrid plant after about 2,000 crosses (Falavigna et al. 2008). In the polyploids species, post-zygotic barriers to the gene flow could be related to the appearance of individuals with odd numbers of their basic chromosome (e.g. 3x, 5x), to the failure of the normal development of the endosperm or to the abortion of the embryo after the fertilization (Marcellan and Camadro 1996). On the other hand, the gene flow among the hexaploid species (A. maritimus, A. pseudoscaber and A. brachyphyllus) can be limited, probably because of ecological and geographic separation. In our study, it was not possible to cross A. tenuifolius with other Asparagus species because it did not grow well under our environmental conditions and the plants did not flower. Some time ago, Bozzini (1963) reported hybrid plants from crosses between A. tenuifolius and A. officinalis with regular meiotic metaphase but with low pollen fertility (45–70 %) compared to parental species (95–99 %). Considering its aforementioned current geographical distribution, A. officinalis or the common diploid ancestor could be expected to be more widely distributed across Europe. These facts suggest that the natural distribution of A. officinalis may be in decline in Europe. Other wild species do seem to be in decline, for example, A. prostratus in Germany, Belgium and the south of Great Britain (Kay et al. 2001; Rich et al. 2002). In Spain, this species was considered locally abundant along the north coast (Rich and Rich 1998, 1999). However, in our fieldwork in this region we were unable to find some previously cited populations.
On the other hand, the different area of distribution of polyploid species could be contributing to their speciation. Chromosome numbers of 2n = 4x = 40, 2n = 6x = 60 and 2n = 12x = 120 have been reported for A. maritimus (Bozzini 1959; Kay et al. 2001; Moreno et al. 2008; Štajner et al. 2002; Valdes 1980). Nowadays, only one dodecaploid population of A. maritimus has been reported and their plants have a different morphological appearance to the other A. maritimus populations studied (Moreno et al. 2008). The distribution of this A. maritimus (12x) population is restricted to a narrowly confined area of southeast of Spain (La Manga del Mar Menor) (Sánchez-Gómez et al. 2008). Following Stebbins (1971) nomenclature it could be considered an apoendemic population, that is: “narrowly restricted polyploids, usually at levels higher than tetraploid, which are related to and probably descendent from wide-spread diploid or having lowers levels of polyploidy” favoured by their perennial habit. A. brachyphyllus population has been reported to be tetraploid (Valdes 1980). However, the A. brachyphyllus population from Yoshkar-Ola (Russia) studied in this paper was found to be hexaploid, showing that populations with different ploidy levels are also present in this species. Hence, polyploidy seems to be playing an important role in the evolution of this group at present. In the asparagus ‘officinalis group’ the relationship between the geographical distribution of diploids and polyploids seems to be related to the adaptation of polyploidy to new ecological conditions (e.g., dodecaploid A. maritimus populations growing a few meters from the seashore). An increase in ploidy level with or without hybridization has often been associated with speciation and the origin of new adaptations (Levin 2002).
According to Moreno et al. (2008), the A.maritimus (12x) population was clearly separated from the A. maritimus (6x) populations. In the current EST-SSR study, one of the two groups formed in A. maritimus (12x) clustered with A. maritimus (6x) populations whereas the other group clustered separately from those ones. These results suggest that the evolutionary pathway in dodecaploid (Cartagena) and hexaploid populations (Albania, Padova and Venice) could be different. Nevertheless, further studies are required in order to clarify the taxonomic position and origin of the population from Cartagena. Moreover, three individuals from “Morado de Huétor” have also been clustered with the A. maritimus (6x) populations. This fact agrees with the interspecific origin suggested for this landrace (Moreno et al. 2008).
Current cultivars are thought to have a narrow genetic base, as indicated using both isozyme and RAPD markers (Brettin and Sink 1992; Geoffriau et al. 1992; Khandka et al. 1996; Knaflewsky 1996; Moreno et al. 2006). Therefore, the use of both asparagus landraces and wild related species in breeding programmes is necessary in order to increase the gene pool of asparagus cultivars. When the interspecific hybrid plant A. acutifolius × (A. officinalis × A. maritimus) obtanined by Falavigna et al. (2008) was backcrossed with a tetraploid plant of A. officinalis eight BC1 plants were obtained, showing that it is possible to broaden the genetic base of cultivated asparagus. In this study, the good crossability between the species closed to cultivated asparagus and both the regular meiosis and pollen fertility observed in their F1 show that it could be possible to introgress agronomically important traits in cultivated asparagus. As has been revealed in this study, the “Morado de Huétor” landrace could be used as bridge to transfer genes from the polyploid wild species studied to cultivated asparagus. The availability of plants with different ploidy levels within the “Morado de Huétor” landrace population (Moreno et al. 2006) allows crosses between the same ploidy levels. In our breeding programme, we have developed tetraploid plants by crossing hexaploids from “Morado de Huétor” with current diploid cultivars. Therefore, backcrossing of interspecific hexaploid hybrids with diploid cultivars could generate new tetraploid plants with germplasm from wild species. In order to increase the gene pool of diploid germplasm, in vitro anther culture of the new tetraploid plants could be a useful tool to obtain diploid plants with new germplasm. Recently, doubled haploid plants developed from diploid and di-haploid from interspecific tetraploid have been reported (Falavigna et at. 2008; Riccardi et al. 2011).
In conclusion, our results show that (1) EST-SSR markers are more informative for phylogenetic relationships at the intrageneric level than ITS sequences; (2) due the high fertility of the interspecific F1 hybrids, the group of species called the ‘officinalis group’ in this study can be considered to be in the primary gene pool, according to the classification of Harlan and de Wet (1971) for cultivated species. A. acutifolius could be included in the secondary gene pool because its gene pool is available to be utilized but more effort are required; and (3) a tetraploid landrace such as “Morado de Huétor” could play an important role as a bridge to transfer genes from the polyploid wild species to cultivated asparagus.
References
Alberti P, Casali PE, Barbaglio E, Toppino L, Mennella G, Falavigna A (2004) Interspecific hybridization for Asparagus breeding. In: Proceedings of the XLVIII Italian society SIFV-SIGA joint meeting, pp 78–79
Álvarez I, Wendel JF (2003) Ribosomal ITS sequences and plant phylogenetic inference. Mol Phylogenet Evol 29:417–434
Baldwin BG, Sanderson MJ, Porter JM, Wojciechowski MF, Campbell CS, Donoghue MJ (1995) The ITS region of nuclear ribosomal DNA: a valuable source of evidence on angiosperm phylogeny. Ann Missouri Bot Gard 82:247–277
Bozzini A (1959) Revisione cito-systematica del genre Asparagus L. I. Le spezie di Asparagus della flora Italiana e chiave analitica per la loro determinazione. Caryologia 12:199–264
Bozzini A (1963) Interspecific hybridization and experimental mutagenesis in breeding Asparagus. Genet Agrar 16:212–218
Brettin TS, Sink KC (1992) Allozyme variation and genetics in asparagus. J Hered 83:383–385
Bruvo R, Michiels NK, D’Souza TG, Schulenburg H (2004) A simple method for the calculation of microsatellite genotype distances irrespective of ploidy level. Mol Ecol 13:2101–2106
Caruso M, Federici CT, Roose ML (2008) EST-SSR markers for asparagus genetic diversity evaluation and cultivar identification. Mol Breed 21:195–204
Clark LV, Jasieniuk M (2011) POLYSAT: an R package for polyploid microsatellite analysis. Mol Ecol Res 11:562–566
Clifford HT, Conran JG (1987) 2. Asparagus, 3. Protasparagus, 4. Myrsiphyllum. In: George AS (ed) Flora of Australia. Australian Government Publishing Service, Canberra, pp 159–164
De Melo NF, Guerra M (2001) Karyotypic stability in asparagus (Asparagus officinalis L.) cultivars revealed by rDNA in situ hybridization. Cytologia 66:127–131
Drummond AJ, Ashton B, Cheung M, Heled J, Kearse M, Moir R, Stones-Havas S, Thierer T, Wilson A (2010) Geneious v5.0, Available from http://www.geneious.com
Ellison JH (1986) Asparagus breeding. In: Breeding vegetable crops. AVI Publishing Co., pp 521–569
Eujayl I, Sledge MK, Wang L, May GD, Chekhovskiy K, Zwonitzer JC, Mian MAR (2004) Medicago truncatula EST-SSRs reveal cross-species genetic markers for Medicago spp. Theor Appl Genet 108:414–422
Falavigna A, Alberti P, Casali PE, Toppino L, Huaisong W, Mennella G (2008) Interspecific hybridization for asparagus breeding in Italy. Acta Hort (ISHS) 776:291–298
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783–791
Felsenstein J (1993) PHYLIP (Phylogeny Inference Package) version 35c Distributed by the author Department of Genetics. University of Washington, Seattle
Fukuda T, Ashizawa H, Suzuki R, Ochiai T, Nakamura T, Kanno A, Kameya T, Yokoyama J (2005) Molecular phylogeny of the genus Asparagus (Asparagaceae) inferred from plastid petB intron and petD-rpoA intergenic spacer sequences. Plant Spec Biol 20:121–132
Geoffriau E, Denoue D, Rameau C (1992) Assessment of genetic variation among asparagus (Asparagus officinalis L.) populations and cultivars: agromorphological and isozymic data. Euphytica 61:169–179
Harlan JR, de Wet JMJ (1971) Toward a rational classification of cultivated plants. Taxonomy 20:509–517
Kay QON, Davies EW, Rich TCG (2001) Taxonomy of the western European endemic Asparagus prostratus (A. officinalis subsp. prostratus) (Asparagaceae). Bot J Linn Soc 137:127–137
Khandka DK, Nejidat A, Golan-Goldhirsh A (1996) Polymorphism and DNA markers for asparagus cultivars identified by random amplified polymorphic DNA. Euphytica 87:39–44
Knaflewsky M (1996) Genealogy of asparagus cultivars. In: Proceedings VIII International Asparagus Symposium, pp 87–91
Komarov V, Shishkin B, Bobrov E (1935) Flora of the USSR. Akademiya Nauk SSSR, Moscow and Leningrad, pp 325–339
Kubota S, Konno I, Kanno A (2012) Molecular phylogeny of the genus Asparagus (Asparagaceae) explains interspecific crossability between the garden asparagus (A. officinalis) and other Asparagus species. Theor Appl Genet 124:345–354
Levin DA (2002) Polyploidy: incidence, types, and modes of establishment. The role of chromosomal change in plant evolution. Oxford University Press, Oxford, pp 84–133
Marcellan ON, Camadro EL (1996) Self- and cross-incompatibility in Asparagus officinalis and Asparagus densiflorus cv. Sprengeri. Can J Bot 74:1621–1625
Marcellan ON, Camadro EL (1999) Formation and development of embryo and endosperm in intra- and inter-specific crosses of Asparagus officinalis and A. densiflorus cv. Sprengeri. Sci Hortic 81:1–11
McCollum GD (1988) Asp 8271 and Asp 8284 Asparagus Germplasm. HortScience 23:641
Moreno R, Espejo JA, Cabrera A, Millán T, Gil J (2006) Ploidic and molecular analysis of ‘Morado de Huetor’ asparagus (Asparagus officinalis L.) population; a Spanish tetraploid landrace. Genet Res Crop Evol 53:729–736
Moreno R, Espejo JA, Cabrera A, Gil J (2008) Origin of tetraploid cultivated asparagus landraces inferred from nuclear ribosomal DNA internal transcribed spacers’ polymorphisms. Ann Appl Biol 153:233–241
Peterson A, John H, Koch E, Peterson J (2004) A molecular phylogeny of the genus Gagea (Liliaceae) in Germany inferred from non-coding chloroplast and nuclear DNA sequences. Plant Syst Evol 245:145–162
Pieroni A (2005) Gathering food from the wild. Cultural history of plants. Routledge, New York, pp 29–44
Reamon-Büttner SM, Schmidt T, Jung C (1999) AFLPs represent highly repetitive sequences in Asparagus officinalis L. Chromosom Res 7:297–304
Riccardi P, Casali PE, Mercati F, Falavigna A, Sunseri F (2011) Genetic characterization of asparagus doubled haploids collection and wild relatives. Sci Hortic 130:691–700
Rich LK, Rich TCG (1998) Distribution and ecology of Asparagus prostratus. I. Cantabria, Northern Spain. Unpublished report, National Museum & Galleries
Rich LK, Rich TCG (1999) Distribution and ecology of Asparagus prostratus. V. Asturias, Northern Spain. Unpublished report, National Museum & Galleries
Rich TCG, Bennallick IJ, Cordrey L, Kay QON, Lockton A, Rich LK (2002) Distribution and population sizes of Asparagus prostratus Dumort. Wild asparagus in Britain. Watsonia, pp 183-192
Roa AC, Chavarriaga-Aguirre P, Duque MC, Maya MM, Bonierbale MW, Iglesias C, Tohme J (2000) Cross-species amplification of cassava (Manihot esculenta) (Euphorbiaceae) microsatellites: allelic polymorphism and degree of relationship. Am J Bot 87:1647–1655
Sánchez-Gómez P, Vera-Pérez JB, Jiménez-Martínez JF (2008) Liliaceae. Asparagus maritimus (L.) Mill. In: Bañares Á, BLanca G, Güemes J, Moreno JC, Ortiz S (eds) Atlas y Libro Rojo de la Flora Vascular Amenazada de España. Adenda. (Ministerio de Medio Ambiente, y Medio Rural y Marino)-Sociedad Española de Biología de la Conservación de Plantas. Madrid, pp 24–25
Sarabi B, Hassandokht MR, Hassani ME, Masoumi TR, Rich T (2010) Evaluation of genetic diversity among some Iranian wild asparagus populations using morphological characteristics and RAPD markers. Sci Hortic 126:1–7
Scott KD, Eggler P, Seaton G, Rossetto M, Ablett EM, Lee LS, Henry RJ (2000) Analysis of SSRs derived from grape ESTs. Theor Appl Genet 100:723–726
Simpson MG (2006) Plant molecular systematics. Plant systematics. Elsevier Academic Press, Amsterdam, pp 477–491
Štajner N, Bohanec B, Javornik B (2002) Genetic variability of economically important Asparagus species as revealed by genome size analysis and rDNA ITS polymorphisms. Plant Sci 162:931–937
Stebbins GL (1971) Chromosomal Evolution in Higher Plants. Addison-Wesley, London
Stephens CT, De Vries RM, Sink KC (1989) Evaluation of asparagus species for resistance to Fusarium oxysporum f. sp. asparagi and F. moniliforme. HortScience 24:365–368
Sturtevant EL (1890) History of garden vegetables. Am Nat 24:719–744
Thiel T, Michalek W, Varshney RK, Graner A (2003) Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor Appl Genet 106:411–422
Torres AM, Weeden NF, Martín A (1993) Linkage among isozyme, RFLP and RAPD markers in Vicia faba. Theor Appl Genet 85:937–945
Valdes B (1975) Notas sobre algunas especies europeaus de Asparagus (Liliaceae). Anales Instituto Botánico A J Cavanilles 32:1079–1092
Valdes B (1980) Asparagus L. In: Tutin TG, Heywood VH, Burges NA, Valentine D (eds) Flora Europaea. Cambridge University Press, Cambridge, pp 71–73
Venezia A, Soressi GP, Falavigna A (1993) Assessment of hybrids and varieties of Asparagus for resistance to Stemphylium vesicarium. Agric Ric 141:49–54
Acknowledgments
The first author acknowledges grant support from the Spanish program Torres Quevedo, financed in part by EU funds (ESF). This work was funded by the Spanish projects AGL2008-05229 and AGL2011-2395, financed in part by EU funds (ERDF). The authors are also grateful for support received from the University of Córdoba, member of the Agrifood Campus of International Excellence (ceiA3).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Castro, P., Gil, J., Cabrera, A. et al. Assessment of genetic diversity and phylogenetic relationships in Asparagus species related to Asparagus officinalis . Genet Resour Crop Evol 60, 1275–1288 (2013). https://doi.org/10.1007/s10722-012-9918-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-012-9918-3