
Abstract
Different regional patterns of glaciation are expected to have brought about a differential effect on the present genetic structure of natural tree populations in the temperate regions. The aim of the present study is to test this hypothesis in Nothofagus antarctica, a key tree species of the temperate forests of southern South America. An almost continuous ice layer characterized the region of the Andes south of 41°S, while towards northern latitudes the pattern was more fragmented. Therefore, a higher chance for the location of larger or more numerous glacial refuges in the north of the Argentinean range, leads us to predict a higher genetic diversity in this region. Twelve natural populations of N. antarctica were sampled along the northern half of its Argentinean range, including six above 41°S and six below that latitude. Sampled populations were genetically characterized through cpDNA and isozyme gene markers. Both groups of populations were compared by means of several diversity and differentiation parameters. A genetic structure analysis was conducted with isozyme data through clustering and Bayesian approaches. Based on three polymorphic chloroplast regions, only two haplotypes were distinguished, one corresponding to the nine northernmost sampled populations and the other to the two southernmost ones. Only the population located between those two groups resulted polymorphic. AMOVA analyses also revealed a latitudinal genetic structure for the populations surveyed, and higher levels of genetic variation were recognized in the northern populations.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The last glaciation has acted as a major force responsible for modelling the current distribution and genetic structure of natural tree populations in the temperate regions. Numerous studies account for this evolutionary relation; most of them correspond to the northern hemisphere, where glaciation has been much more extensive and continuous than in the south.
Donat (1933) was the first to suggest differentiation processes determined by glacial influence in southern South America. He postulated the existence of a glacial barrier between 48°S and 52°S in order to explain the bicentric distribution of several taxa. Other latitudinal differences in the Patagonian biota were more recently proposed based on the reconstruction of paleo-climates by Markgraf et al. (1996), who suggested a latitudinal division for the evolutionary history of Patagonian vegetation into three areas: south of 51°S, between 51°S and 43°S and north of 43°S.
Different regional patterns of glaciation are expected to have brought about a differential effect on the ecosystems under their influence. A single key species distributed over different glaciation patterns could give the possibility to test this hypothesis. Such a scenario can be found in southern South America.
Based on geological and geomorphologic observations, Flint and Fidalgo (1964, 1969) delineated maps showing the eastern limit of the ice sheet during Last Glacial Maximum (LGM) between 39°10′ S and 43°10′ S. Glaciated and ice-free areas are distinguished in detail; this allows us to recognize two different glaciation patterns in the north of Argentine Patagonia, roughly delimited by the 41°S latitude.
As described by Flint and Fidalgo (1964, 1969), north of 41°S the water divide follows a clear north–south direction coinciding with a high mountain axis, which forms the political division between Chile and Argentina. The mountain ice sheet nourished from the west by the Pacific humidity exceeded the Andes range through the transverse valleys as single tongues that extended eastward from the high mountains down main valleys, now occupied by lakes. These ice tongues became more sharp-pointed northward, leaving in this direction more ice-free areas between them.
South of 41°S, instead, the water divide is constituted by several mountain ranges with north–south orientation located from 25 to 75 km east of the high mountain axis, thus constituting a broader Cordillera. Those eastern mountains acted as barriers to eastward flow of the mountain ice sheet, which was not thick enough as to surmount most of them. Consequently, the eastern border of the ice followed mainly a north–south direction determined by the water divide. Beyond it, the dry steppe probably did not offer great suitable places for the establishment of forests. On the contrary, the ice-free areas between the “glacial tongues” in the northern region were not as dry as the arid steppe, thus providing better chances for the localization of bigger or more numerous glacial refuges.
Nothofagus antarctica (G. Forster) Oersted, locally called Ñire, is a tree species of the temperate forests of southern South America that ranges along both glaciation patterns. It has a broad ecological range, covering almost 20 latitudinal degrees, in altitudes from the timberline to the sea level, and in extremely opposite humidity conditions, from peat bogs to semiarid areas with less than 500 mm annual precipitation (Ramírez et al. 1985).
Particularly the northern Argentinean range of N. antarctica is fragmentary, with pure forest patches of variable dimension scattered in a steppe matrix or in combination with patches of other forest tree species. Mixed patches with Austrocedrus chilensis (D.Don.) Pic. Ser. et Bizzarri, Araucaria araucana (Molina) K. Koch. and other Nothofagus species are common in this region. In contrast, a more continuous pure forest characterizes the region southward of 41°S.
The reconstruction of the recent history of natural populations of N. antarctica is one of the objectives of the present study, and palynological antecedents are crucial for this purpose. Unfortunately, such information is scarce and present only in specific studies for scattered locations in Patagonia. In addition, pollen morphology makes it difficult to draw a distinction among the Nothofagus species (Heusser 1984). However, some authors can clearly identify six South American Nothofagus, among them N. antarctica (Markgraf and D’Antoni 1978).
The scattered pollen records suggest the persistence of Nothofagus forests in several locations west of the Andes during the last glaciation, mainly along the Pacific Coastal Range (Villagrán 1991), but also along the Central Valley and foothills of the Andes north of the Lake District (42°S) (Heusser 1983, 1984; Villagrán 1991), and even at latitudes between 43°S and 51°S (Markgraf et al. 1996; Ashworth et al. 1991, cit. in Markgraf et al. 1996).
East of the Andes the information between 43°S and 51°S is scarce and, according to one study at Perito Moreno Glacier (50°S) (Mercer and Ager 1983, cit. in Markgraf et al. 1996) apparently there were no trees. On the other hand, local studies from north of 41°30′ S showed evidence to support the presence of N. antarctica in the eastern side of the Andes shortly after glaciation, about 14,000 years BP (Markgraf 1984, 1987; Markgraf et al. 1986; Markgraf and Bianchi 1999). Accordingly, several genetic studies support the existence of multiple ice-age refuges east and west of the Andes of different native tree species [e.g. Nothofagus nervosa (Phil.) Dim. et Mil. (Marchelli et al. 1998; Marchelli and Gallo 2006), Fitzroya cupressoides (Mol.) Johnst. (Premoli et al. 2000), Austrocedrus chilensis (Pastorino and Gallo 2002), Pilgerodendron uviferum (Don.) Flor. (Premoli et al. 2002)].
The hypothesis of the present study is that the forest ecosystems northward and southward of 41°S have differentially been affected by Last Glacial Maximum, and our prediction is that the degree and distribution of the genetic variation of N. antarctica will show a different pattern on either side of this line. We additionally expect a higher genetic diversity and differentiation among the northern populations due to the likely occurrence of multiple glacial refuges there. On the contrary, the southern populations are supposed to be less variable due to post-glacial re-colonization from northern refuges and consequent loss of genetic variants after several founder events.
Our first approach will be based on the screening of polymorphisms on the chloroplast genome through PCR-RFLP. Due to their low mutation rate and maternal inheritance in most angiosperms (Birky 1995), cpDNA markers have been utilized to reconstruct recent life history of several forest tree species, particularly to determine the most likely localization of glacial refuges and to infer post-glaciation migration routes (e.g. Petit et al. 2002). Different cpDNA lineages in natural populations are evidence of their different origins. Nuclear genes will also be surveyed by using isozyme markers, which due to their bi-parental inheritance, higher variability and higher mutation rate, are expected to reflect the impact of processes suffered during the last generations after deglaciation. The bi-parental inheritance implies that the influence of pollen-mediated gene flow, which is usually much more extended than the exchange of genes through seeds, can be detected in the patterns of the genetic variation observed with these markers.
Materials and methods
Sampling
Twelve natural Nothofagus antarctica populations of the northern half of its Argentinean range were sampled (Table 1), six located northward and six southward of 41°S. Buds from a total of 674 trees equally distributed between both groups were collected in winter. In each population, trees were chosen keeping a minimum distance of 30 m among them in order to reduce the risk of sampling related individuals. Samples were kept at −18°C until laboratory analyses.
Chloroplast DNA
DNA was extracted from buds following the protocol described by Dumolin et al. (1995) with slight modifications as mentioned in Marchelli and Gallo (2006). A total of 60 individuals was analysed (five from each of the 12 populations). Twenty trees originating from 10 populations (two individuals per population) were employed to screen chloroplast DNA for polymorphisms. Different combinations of universal primers (Heinze 2007) were used to amplify 11 intergenic non-coding regions: ycf3-ccmp6 (Heinze 2007; Weising and Gardner 1999), atpH-atpI (Grivet et al. 2001), trnD-trnT (Demesure et al. 1995); trnS-rps4 (Demesure et al. 1995; Souza-Chies et al. 1997), ucp-a/trnT-ucp-b/trnL (Taberlet et al. 1991), rpoC1-f5-rpoC1 exon2 (Grivet et al. 2001; Liston 1992), atpI-ccmp5 (Grivet et al. 2001; Weising and Gardner 1999), trnK1-trnK2 (Demesure et al. 1995), matKf2-ccmp1 (Heinze 2007; Weising and Gardner 1999), trnK2-trnQ (Dumolin-Lapègue et al. 1997), ccmp5-rpoC2-r5 (Weising and Gardner 1999; Grivet et al. 2001). Ten chloroplast DNA microsatellites (cpSSRs) were additionally tested: ccmp1, ccmp2, ccmp5, ccmp6, ccmp7 (Weising and Gardner 1999), mdt1, mdt3, mdt4, mcd4, mcd5 (Deguilloux et al. 2003). Amplifications were carried out in a total volume of 25 μl containing about 15 ng of template DNA, 1.6 mM of MgCl2, 100 μM of each dNTP, 0.2 μM of each primer and 1 U of Taq polymerase (Invitrogen-Gibco) with the respective 1× PCR buffer. A common PCR program (Heinze 2007) was used for all the intergenic non-coding regions and for five of the cpDNA SSRs (Weising and Gardner 1999): 3 min at 94°C, followed by 10 cycles of 50 s at 94°C and 1 min at 70°C, then 35 cycles of 50 s at 94°C, 50 s at 55°C and 2 min at 70°C, with a final extension step of 10 min at 70°C. For the remaining cpDNA SSRs the PCR conditions were as proposed by Deguilloux et al. (2003) and the following cycling temperatures were employed: 5 min at 94°C, 25 cycles of 1 min 94°C, 1 min at 45°C and 1 min at 72°C, with a final extension step of 8 min at 72°C. The reactions were performed either in a Biometra Uno-Thermo Block or in a MJResearch PT-200 thermo cycler. In those cases where amplification of the intergenic regions was positive, 8 μl of PCR product were digested by 5 U of restriction endonuclease HinfI or TaqI in a total volume of 23 μl. Digestion was performed at 65°C for 3 h in the case of TaqI and at 37°C overnight in the case of HinfI. The total digestion volume was loaded into 8% non-denaturating polyacrylamide gels. Electrophoresis was carried out in TRIS borate EDTA (1×) at 300 v for 4 h. Visualization of the fragments was done under UV light after staining with ethidium bromide. Gel documentation was obtained with a digital camera, and images were analysed with the BioDoc Analyse version 2.0 (Biometra).
Three chloroplast regions showed polymorphism in the subsample: atpH-atpI; trnD-trnT, both after digestion with HinfI, and ycf3-ccmp6 digested with TaqI. These regions were then screened in all the individuals (N = 60). One individual of each variant for each chloroplast region (six in total) was sequenced in an ABI3100 facility (Applied Biosystems) in order to confirm the polymorphisms. The regions were amplified as mentioned above and purified using the Wizard Kit of Promega. Products were sequenced using the Big Dye 3.1 terminator cycle sequencing kit (Applied Biosystems) and analysed with MEGA 3.1 (Kumar et al. 2004).
Polymorphic fragments were labelled by decreasing order of fragment size as visualized in the polyacrylamide gels. Point mutations in the restriction endonuclease motif were denoted with a 9. Haplotypes were defined according to different combinations of length variants, following the procedure of Demesure et al. (1996). The average within-populations gene diversity (h S), the total gene diversity (h T) and the gene differentiation over all populations (G ST) were estimated according to Pons and Petit (1995) using the program HAPLODIV available at http://www.pierroton.inra.fr/genetics/labo/Software/.
Isozymes
Isozyme genotypes were determined by subjecting buds of each of the 674 sampled trees to horizontal starch gel electrophoresis. Laboratory procedures followed those described by Stecconi et al. (2004) with slight modifications. Vegetative extraction buffer I from Cheliak and Pitel (1984) and a discontinuous buffer system was utilized on routine: 0.3 M boric acid/0.06 M NaOH till pH 8.2 (Poulik 1959); gel: 0.07 M Tris/0.008 M citric acid till pH 8.5; starch 10.5% w/v, sucrose 2% w/v; for 6 h at 80 mA. Two isozymes with their genetic control previously determined (Pastorino et al. 2008) were revealed with the staining solutions proposed by Cheliak and Pitel (1984): phosphoglucose isomerase (PGI, E.C.5.3.1.9) and phosphoglucomutase (PGM, E.C.2.7.5.1).
Allelic and genotypic frequencies were calculated for each population by means of GSED 1.1d (Gillet 1997), and subsequently the following diversity parameters were estimated: allelic richness r g, with CONTRIB 1.02, available at http://www.pierroton.inra.fr/genetics/labo/Software (Petit et al. 1998), effective number of alleles A e, with GSED 1.1d, expected heterozygosity H e and observed heterozygosity H o, using Arlequin 3.0 (Excoffier et al. 2005). Allelic richness was calculated after rarefaction to a common sample size (g, the smallest) in order to standardize and avoid the bias of the uneven sample sizes of the different populations. A corrected value was expressed by subtracting one from that calculated after the Hulbert (1971) formula (El Mousadik and Petit 1996).
Differences in allelic frequencies among populations were tested by means of an exact test with Arlequin 3.0. Once homogeneity was denied, differentiation was expressed by means of Gregorius’ (1985) D j and δ, calculated with GSED 1.1d and considering all the populations as infinite, and with the more widely-known F ST of Wright (1978) obtained with Arlequin 3.0.
Genetic structure of the sampled populations was analysed through different approaches. First, a cluster analysis based on Gregorius’ (1974) genetic distances between populations (d 0) was made using the UPGMA linkage procedure (Sneath and Sokal 1973) with SAS software (1989). A second approach was achieved by utilizing the software package BAPS 4.14 (Corander et al. 2003), based on Bayesian inference. Geographic coordinates of each population were added in this case. In order to transform geodesic in geocentric coordinates, the program Transcoord TS was utilized (Olondriz and Brunini 1998).
Subsequently, a partitioning of the molecular variance among and within groups (Excoffier et al. 1992) was performed in each of the three groupings proposed [i.e. the original hypothesis (northern and southern groups), the result of the cluster analysis based on d 0 genetic distance, and the structure resulting from the Bayesian approach], by means of locus-by-locus AMOVA analyses using Arlequin 3.0.
In order to test the hypothesis that northern populations are more variable than southern ones, two groups were formed by pooling the trees of the populations northward and southward of 41°S in each of them. Thus, each group contained a similar number of trees from six populations. A homogeneity G test (Sokal and Rohlf 1981) of the allelic frequencies was performed between these two groups with the help of GSED 1.1d, and their diversities were characterized through the parameters described above.
However, this procedure implied a certain bias since sample sizes of each population were neither equal nor proportional to the corresponding population sizes. Thus the over-represented populations brought about a greater weight on the calculated diversity parameters. As an alternative, the average value among the populations of each group was calculated for each of the parameters.
Results and discussion
Chloroplast DNA
The three polymorphic cpDNA fragments scored in all the populations consisted in one insertion/deletion in fragment ycf3-ccmp6 and two restriction site mutations, one in each of the other two analysed fragments. These polymorphisms allowed the identification of two haplotypes separated by three mutations. The analysis of the DNA sequences confirmed the presence of an insertion/deletion of 14 bp in fragment ycf3-ccmp6.
The distribution of these two haplotypes was geographically structured in a latitudinal way, since the nine northernmost sampled populations bore one haplotype and the two southernmost ones held the other. The only polymorphic population was Cholila (42°32′ S), which is situated at a latitude between those two groups of populations (Fig. 1). Consequently, the genetic diversity calculated at the species level resulted obviously very low (h s = 0.033, s.e. = 0.033; h t = 0.324, s.e. = 0.141), and the differentiation among populations very high (G ST = 0.897, s.e. = 0.113). This was also the case for other Nothofagus species of these forests [G ST = 0.93 for N. nervosa (Marchelli and Gallo 2006); G ST = 0.757 for N. obliqua (Azpilicueta et al., submitted.)].

Snowline at Last Glacial Maximum (LGM) according to Hollin and Schilling (1981) and Nothofagus antarctica sampled populations showing the distribution of the two cpDNA haplotypes found (full circle corresponds to the northern haplotype, empty circle to the southern haplotype)
Taking into consideration the very low mutation rate of the chloroplast genome (Clegg et al. 1994), the existence of two groups of populations whose haplotypes are differentiated by three mutations suggests a long lasting isolation in at least two different glacial refuges from which the analysed populations of N. antarctica could have spread out after the last glacial period. The larger distribution of the northern haplotype might be related with the two different glaciation types. The northern, less glaciated region could have been re-colonized first, since glacial retreat could be assumed to have been faster. In agreement with this suggestion, the few, local registers of fossil pollen reported that forest expansion was delayed by about 3,000 years east of the Andes at latitudes south of 43°S (Markgraf et al. 1996). Thus, a refuge located in the northern region between the “glacial tongues”, as suggested for other studied species (Marchelli and Gallo 2006; Azpilicueta et al., submitted), would have expanded after glaciation retreat by re-colonizing the valleys now free of ice. Southward of 41°S, instead, a refuge could only have been situated in the steppe, just in front of the glacier. This idea has been already proposed for other three drought tolerant tree species of Patagonia: Austrocedrus chilensis (Pastorino and Gallo 2002), Nothofagus obliqua (Azpilicueta et al., submitted) and Araucaria araucana (Marchelli et al., submitted).
The presence of the two haplotypes at Cholila could be indicating a secondary contact zone with the admixture of migratory routes. Meeting points usually result in transition zones of highly diverse populations (e.g. Vendramin et al. 1998; Mátyás and Sperisen 2001). This possible admixture of two migratory routes seems to have been relatively recent, since only one population turned out to be polymorphic, which is precisely located between the two groups of monomorphic populations.
Latitudinal variation in the distribution of haplotype variants and low levels of polymorphisms were also observed in other Nothofagus species (Marchelli and Gallo 2006; Millerón et al. 2008; Azpilicueta et al., submitted) suggesting a general trend in these forests. The low levels of cpDNA variation observed for the Nothofagus species analysed so far contrast with those of other forest trees where several haplotypes were usually identified (see review in Petit et al. 2005). Regarding our samples, Cholila is the only population which presented evidence of seed-mediated gene flow among populations of both groups.
Latitudinal differentiation among cpDNA variants was also observed in plant species from North America (Soltis et al. 1997; Magni et al. 2005). Moreover, a marked difference in the historical colonization between oak species from North America and Europe was interpreted as the response of the species to different glacial scenarios. Whereas in Europe oaks were separated from the ice sheet by a widespread tundra, in North America oaks grew close to the ice sheet and therefore movement after LGM was moderate (Magni et al. 2005; Grivet et al. 2006). Evidence in South America seems to support a scenario similar to that of North America.
The lack of detailed palynological information in southern South America is a constraint to further argue the possible location of glacial refuges. Moreover, the low levels of polymorphisms detected at the chloroplast DNA avoid further speculations on the recolonization history with this sole source of information.
Isozymes
Allelic frequencies and the genetic variation parameters of the analysed populations are shown in Table 2. Regarding the mean values of those parameters among the 12 populations, the intra-population variation should be considered as moderate at the species level. Quilanlahue population presented the lowest values of intra-population variation (Table 2). An effective number of alleles per locus (A e) of only 1.028 is evidence of virtual monomorphism in the two sampled genes. Contrasting, Correntoso population, which is near to it, is the most diverse, as seen through A e (1.446) and H e (0.307).
Significant differences in allelic frequencies were observed between the northern and the southern groups of pooled populations for both surveyed loci (P PGI = 0.0018 and P PGM < 0.0001). Both the pooling and the average-among-populations alternatives to compare northern and southern populations showed a higher genetic diversity in the north regarding the number of effective alleles (A e) and expected (H e) and observed (H o) heterozygosities (Table 2). However, the results diverged between both alternatives when the allelic richness was considered. The pooling procedure threw a higher richness in the north but the average-among-populations showed the opposite. This result is mainly caused by the unexpectedly rather frequent presence of the rare allele Pgi2-120 in the southern population El Foyel (9%), and the virtual monomorphism of both markers in the northern population Quilanlahue. In any case, one should consider that all alleles present in the southern group are also present in the northern group, while one of the alleles of the northern group could not be found in the south (Pgm2-96).
Genetic differentiation was moderate at the species level (δ = 0.082; F ST = 0.109), being the northern populations the most differentiated (Table 2). The average value of D j among northern populations (0.106) is quite larger than among the southern ones (0.065), in agreement with the fragmentary condition of this part of the range. Restricted gene flow among fragments would explain larger differentiation. In the south portion of the region sampled, instead, lower differentiation seems to respond to the expectation for a widespread species (Hamrick et al. 1992), in which pollen-mediated gene flow is typically extensive.
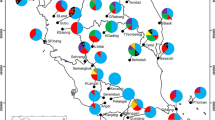
The northern population Caviahue was the most differentiated one, as measured by D j (0.189), probably due to the good allele frequency balance in Pgm2 locus (it is the population with the most even structure in this marker), and the presence of four alleles in Pgi2. The southern population El Foyel followed (D j = 0.141), in this case due to the relative high frequency of the rare allele Pgi2-120 (Fig. 2).

Allelic differentiation snail for the gene pool in Nothofagus antarctica sampled populations. Length of the radii of each “pie portion” is proportional to D j , and radius of the circle is proportional to δ
The region surveyed roughly coincides with that of a population genetic study of another native tree species: A. chilensis, also based on isozyme markers (Pastorino et al. 2004). In the case of this conifer, the northern populations also turned out to be the most variable.
Through the homogeneity exact test among populations significant differences were revealed (P < 0.0001). Two clear groups can be recognized in the UPGMA dendrogram: the smaller one constituted by four of the northern populations, and the other by Quilanlahue, Córdoba and those of the southern half of the sampled region (Fig. 3). A single population clearly separated in each group: Caviahue and Foyel, in the northern and southern groups respectively.

UPGMA dendrogram with gene pool distances at two isozyme loci between 12 Nothofagus antarctica natural populations
The best structure from the Bayesian analysis resulted in three groups: one with the northern populations Roblecillos, Tromen and Caviahue, other with Foyel solely, and the third one constituted by the rest [log (marginal likelihood) of the optimal partitioning = −868.044]. Thus, besides the explicit separation of the northern and the southern populations of the original hypothesis, in the other two structures proposed (those resulting from the dendrogram and the Bayesian approach), a northern group could also clearly be seen. This evidenced a general latitudinal trend.
According to the AMOVA analyses, the structure revealed by the UPGMA dendrogram based on Gregorius’ genetic distance appeared to be the best. It showed that almost all of the variation not explained by the intrapopulation variation lay on the variation among groups (15.59%), and additionally revealed the lowest percentage of variation among individuals within populations (81.81%) (Table 3).
The AMOVA analyses showed a good structure for the three proposed groupings, since the variation among-groups resulted in the three cases higher than the variation among-populations (all covariance components associated with the different possible levels of genetic structure were significant, see Table 3). Namely, the AMOVA analyses serve as additional evidence to support a latitudinal structure of the sampled populations.
Final considerations
The combination of two marker types characterized by very different evolutionary dynamics, such as cpDNA and isozymes, is desirable in order to get a complete picture of the genetic pattern of the natural populations of a species. The results of both marker types are not necessarily coincident but complementary (Avise 1994), and both sources of information have been recommended in order to identify evolutionary significant units (Moritz 1994).
Two groups of populations with a latitudinal distribution were identified by the cpDNA markers, which could be assumed as being of different origin. A merging region resulting from seed-mediated gene flow was also detected. This result rules out the hypothesis of the southern populations deriving from a northern refuge. However, it serves to corroborate a latitudinal pattern of genetic variation. Likewise, the isozyme approach allowed the identification of other latitudinal groups undetected with chloroplast markers, therefore providing additional insights. These nuclear markers were also useful to recognize the higher genetic variation of the northern populations. Different genetic variation patterns between north and south are hence evident, although not with a strict division at 41°S. We can nevertheless confidently sustain a general latitudinal trend in the genetic structure of N. antarctica, in agreement with the latitudinal pattern of glaciation.
To sum up, in the eastern foothills of the Andes range, along the northern half of its current distribution area, N. antarctica would have survived last glaciation in at least two refuges, one situated somewhere north of 42°30′ S, and the other somewhere south of that latitude. On the other hand, the higher differentiation among the northern populations as revealed by the nuclear markers, would be evidence of several patches with restricted gene flow among them (even pollen-mediated gene flow). However, those patches would have a unique origin (due to be carrier of the same cpDNA haplotype), and thus two alternative hypotheses arise as possible: either (1) migration effectively occurred after glaciation from a unique refuge located in any of the transversal valleys, as to re-colonize the previously glaciated areas, and thus the isolation among those patches is only a feature of the last generations, or (2) the patches located in different valleys derived through a fragmentation process caused by glaciation from a somehow continuous forest existing before LGM, thus creating several refuges but with a unique haplotype. The second alternative matches the already proposed multiple refuges model (Marchelli and Gallo 2006), and sounds more plausible. In any of both scenarios, and closer to the present days, the northern refuge (or refuges) would have extended its (their) influence up to 42°30′ S, where the northern genes would have merged with the southern ones.
These results, although matching the previous information on other Patagonian tree species, should be confirmed with additional information. First of all, more nuclear genes are desirable to support the derived conclusions. Likewise, additional polymorphic cpDNA fragments could be the key to identify different haplotypes among the northern populations of N. antarctica. Also the screening of more southern populations, and even those of Tierra del Fuego Island, will cast light on the evolutionary relations between LGM and the genetic patterns of the main forest trees of Patagonia. The consideration of the widespread Nothofagus pumilio (Poepp. et Endl.) Krasser, which has a roughly coincident latitudinal range with our object of study, would be relevant in this regard. Ongoing research lines will give insights into the evolutionary history of these widespread species.
References
Ashworth AC, Markgraf V, Villagrán C (1991) Late quaternary climatic history of the Chilean Channels based on fossil pollen and beetle analyses, with an analysis of the modern vegetation and pollen rain. J Quat Sci 6:279–291. doi:10.1002/jqs.3390060403
Avise JC (1994) Molecular markers. Natural History and Evolution, Chapman and Hall, New York
Birky CW (1995) Uniparental inheritance of mithocondrial and chloroplast genes: mechanisms and evolution. Proc Natl Acad Sci USA 92:11331–11338. doi:10.1073/pnas.92.25.11331
Cheliak WM, Pitel JA (1984) Techniques for starch gel electrophoresis of enzymes of forest tree species. Information Report PI-X-42. Petawawa National Forestry Institute, Canadian Forestry Service, p 49
Clegg MT, Gaut BS Learn GH Jr, Morton BR (1994) Rates and patterns of chloroplast DNA evolution. Proc Natl Acad Sci USA 91:6795–6801
Corander J, Waldmann P, Sillanpää MJ (2003) Bayesian analysis of genetic differentiation between populations. Genetics 163:367–374
Deguilloux MF, Dumolin-Lapègue L, Gielly L, Grivet D, Petit JR (2003) A set of primers for the amplification of chloroplast microsatellites in Quercus. Primer Note Mol Ecol Notes 3:24–27. doi:10.1046/j.1471-8286.2003.00339.x
Demesure B, Sodzi N, Petit JR (1995) A set of universal primers for amplification of polymorphic non-coding regions of mitochondrial and chloroplast DNA in plants. Mol Ecol 4:129–131. doi:10.1111/j.1365-294X.1995.tb00201.x
Demesure B, Comps B, Petit JR (1996) Chloroplast DNA phylogeography of the common beech (Fagus sylvatica L.) in Europe Evolution. Int J Org Evol 50:2515–2520. doi:10.2307/2410719
Donat A (1933) Sind Drosera uniflora und Pinguicula antarctica bizentrische Typen? Ber Dtsch Bot Ges 2:67–77
Dumolin S, Demesure B, Petit RJ (1995) Inheritance of chloroplast and mitochondrial genomes in pedunculate oak investigated with an efficient PCR method. Theor Appl Genet 91:1253–1256. doi:10.1007/BF00220937
Dumolin-Lapègue S, Demesure B, Fineschi S, Le Corre V, Petit RJ (1997) Phylogeographic structure of white oaks throughout the European continent. Genetics 146:1475–1487
El Mousadik A, Petit RJ (1996) High level of genetic differentiation for allelic richness among populations of the argan trees [Argania spinosa (L.) Skeels] endemic of Morocco. Theor Appl Genet 92:832–839. doi:10.1007/BF00221895
Excoffier L, Smouse P, Quattro JM (1992) Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131:479–491
Excoffier L, Laval G, Schneider S (2005) Arlequin ver. 3.0: an integrated software package for population genetics data analysis. Evol Bioinform Online 1:47–50
Flint RF, Fidalgo F (1964) Glacial geology of the east flank of the Argentine Andes between latitude 39°10′ S and latitude 41°20′ S. Geol Soc Am Bull 75:335–352. doi:10.1130/0016-7606(1964)75[335:GGOTEF]2.0.CO;2
Flint RF, Fidalgo F (1969) Glacial drift in the eastern Argentine Andes between latitude 41°10′ S and latitude 43°10′ S. Geol Soc Am Bull 80:1043–1052. doi:10.1130/0016-7606(1969)80[1043:GDITEA]2.0.CO;2
Gillet E (1997) GSED Genetic Structures from Electrophoresis Data, version 1.1d, program and user’s manual. Institut für Forstgenetik und Forstpflanzenzüchtung, Faculty of Forest Genetics and Forest Ecology, University of Göttingen, URL: http://www.uni-forst.gwdg.de/forst/fg/index.htm
Gregorius H-R (1974) On the concept of genetic distance between populations based on gene frequencies. IUFRO joint meeting of working parties on population and ecological genetics, breeding theory and progeny testing. Department of Forest Genetics, Royal College of Forestry, Stockholm, pp 17–26
Gregorius H-R (1985) Measurement of genetic differentiation in plant populations. In: Gregorius H-R (ed) Population genetics in forestry (Lecture Notes in Biomathematics), vol 60, pp 276–285
Grivet D, Heinze B, Vendramin GG, Petit RJ (2001) Genome walking with consensus primers: application to the large single copy region of chloroplast DNA. Mol Ecol Notes 1:345–349. doi:10.1046/j.1471-8278.2001.00107.x
Grivet D, Deguilloux M-F, Petit RJ, Sork V (2006) Contrasting patterns of historical colonization in white oaks (Quercus spp.) in California and Europe. Mol Ecol 15:4085–4093. doi:10.1111/j.1365-294X.2006.03083.x
Hamrick JL, Godt MJW, Sherman-Broyles SL (1992) Factors influencing levels of genetic diversity in woody plant species. New For 6:95–124. doi:10.1007/BF00120641
Heinze B (2007) A data base of PCR primers for the chloroplast genomes of higher plants. Plant Methods 3:4. http://www.plantmethods.com/content/3/1/4. doi:10.1186/1746-4811-3-4
Heusser CJ (1983) Quaternary pollen record from Laguna Tagua Tagua, Chile. Science 219:1429–1432. doi:10.1126/science.219.4591.1429
Heusser CJ (1984) Late-glacial—Holocene climate of the Lake District of Chile. Quat Res 22:77–90. doi:10.1016/0033-5894(84)90008-5
Hollin JT, Schilling DH (1981) Late Wisconsin-Weichselian mountain glaciers and small ice caps. In: Denton GH, Hughes TJ (eds) Late Wisconsin-Weichselian mountain glaciers and small ice caps. The Last Great Ice Sheets. Wiley, New York, pp 179–206
Hulbert SH (1971) The nonconcept of species diversity: a critique and alternative parameters. Ecology 52:577–586. doi:10.2307/1934145
Kumar S, Tamura K, Nei M (2004) MEGA 3: integrated software for molecular evolutionary genetic analysis and sequence alignment. Brief Bioinform 5:150–163. doi:10.1093/bib/5.2.150
Liston A (1992) Variation in the chloroplast genes rpoC1 and rpoC2 of the genus Astralagus (Fabaceae): evidence from restriction site mapping of a PCR-amplified fragment. Am J Bot 79:953–961. doi:10.2307/2445007
Magni CR, Ducuoso A, Caron H, Petit RJ, Kremer A (2005) Chloroplast DNA variation of Quercus rubra Lin North America and comparison with other Fagaceae. Mol Ecol 14:513–524. doi:10.1111/j.1365-294X.2005.02400.x
Marchelli P, Gallo LA (2006) Multiple ice-age refuges in a southern beech of South America as evidenced by chloroplast DNA markers. Conserv Genet 7:591–603. doi:10.1007/s10592-005-9069-6
Marchelli P, Gallo LA, Scholz F, Ziegenhagen B (1998) Chloroplast DNA markers reveal a geographical divide across Argentinean southern beech Nothofagus nervosa (Phil.) Dim. et Mil. distribution area. Theor Appl Genet 97:642–646. doi:10.1007/s001220050940
Markgraf V (1984) Late pleistocene and holocene vegetation history of temperate Argentina: Lago Morenito, Bariloche. Diss Bot 72 (Festschrift Welten), pp 235–254
Markgraf V (1987) Paleoenvironmental changes at the northern limit of the subantarctic Nothofagus forest, lat 37°, Argentina. Quat Res 28:119–129. doi:10.1016/0033-5894(87)90037-8
Markgraf V, D’Antoni HL (1978) Pollen flora of Argentina. The University of Arizona Press, Tucson
Markgraf V, Bianchi MM (1999) Paleoenvironmental changes during the last 17, 000 years in western Patagonia: Mallín Aguado, Province of Neuquén, Argentina. Bamberger Geogr Schr 19:175–193
Markgraf V, Bradbury JP, Fernández J (1986) Bajada de Rahue, Province of Neuquén, Argentina: an interstadial deposit in northern Patagonia. Palaeogeogr Palaeoclimatol Palaeoecol 56:251–258. doi:10.1016/0031-0182(86)90097-0
Markgraf V, Romero EJ, Villagrán C (1996) The history and paleoecology of south American Nothofagus forests biogeography of Nothofagus Forests. In: Veblen TT (ed) The ecology and biogeography of Nothofagus Forests. Yale University Press, New Haven, pp 354–386
Mátyás G, Sperisen C (2001) Chloroplast DNA polymorphisms provide evidence for postglacial re-colonization of oaks (Quercus spp.) across the Swiss Alps. Theor Appl Genet 102:12–20. doi:10.1007/s001220051613
Millerón M, Gallo L, Marchelli P (2008) The effect of volcanism on postglacial migration and seed dispersal. A case study in southern South America. Tree Genet Genomes 4:435–443. doi:10.1007/s11295-007-0121-1
Moritz C (1994) Defining “Evolutionary Significant Units” for conservation. Trends Ecol Evol 9:373–375. doi:10.1016/0169-5347(94)90057-4
Olondriz J, Brunini C (1998) Transcoord: Manual del Usuario, Facultad de Ciencias Astronómicas y Geofísicas, Universidad Nacional de La Plata
Pastorino MJ, Gallo LA (2002) Quaternary evolutionary history of Austrocedrus chilensis, a cypress native to the Andean-Patagonian Forest. J Biogeogr 29:1167–1178. doi:10.1046/j.1365-2699.2002.00731.x
Pastorino MJ, Gallo LA, Hattemer HH (2004) Genetic variation in natural populations of Austrocedrus chilensis, a cypress of the Andean-Patagonian Forest. Biochem Syst Ecol 32:993–1008. doi:10.1016/j.bse.2004.03.002
Pastorino MJ, Marchelli P, Milleron M, Gallo LA (2008) Inheritance of isozyme variants in Nothofagus antarctica (G.Forster) Oersted. Basic Appl Genet (in press)
Petit RJ, El Mousadik A, Pons O (1998) Identifying populations for conservation on the basis of genetic markers. Conserv Biol 12:844–855. doi:10.1046/j.1523-1739.1998.96489.x
Petit RJ, Csaikl U, Bordács S et al (2002) Chloroplast DNA variation in European white oaks. Phylogeographyand patterns of diversity based on data from over 2600 populations. For Ecol Manage 156:5–26. doi:10.1016/S0378-1127(01)00645-4
Petit RJ, Duminil J, Fineschi S, Hampe A, Salvini D, Vendramin GG (2005) Comparative organization of chloroplast, mitochondrial and nuclear diversity in plant populations. Mol Ecol 14:689–701. doi:10.1111/j.1365-294X.2004.02410.x
Pons O, Petit RJ (1995) Estimation, variance and optimal sampling of gene diversityI. Haploid locus. Theor Appl Genet 90:462–470. doi:10.1007/BF00221991
Premoli AC, Kitzberger T, Veblen TT (2000) Isozyme variation and recent biogeographical history of the long-lived conifer Fitzroya cupressoides. J Biogeogr 27:251–260. doi:10.1046/j.1365-2699.2000.00402.x
Premoli AC, Souto CP, Rovere AE, Allnut TR, Newton AC (2002) Patterns of isozyme variation as indicators of biogeographic history in Pilgerodendron uviferum (D.Don) Florin. Divers Distrib 8:57–66. doi:10.1046/j.1472-4642.2002.00128.x
Poulik MD (1959) Starch gel electrophoresis in a discontinuous system of buffers. Nature 180:1477–1478. doi:10.1038/1801477a0
Ramírez C, Correa M, Figueroa H, San Martín J (1985) Variación del hábito y hábitat de Nothofagus antarctica en el centro sur de Chile. Bosque 6:55–73
SAS Institute Inc (1989) SAS/STAT® User’s Guide, Version 6, Fourth Edition, Volume 2, Cary, NC. SAS Institute Inc., 846 pp
Sneath PHA, Sokal RR (1973) Numerical taxonomy. W.H. Freeman, San Francisco
Sokal RR, Rohlf FJ (1981) Biometry: the principles and practice of statistics in biological research, 2nd edn. Freeman, San Francisco
Soltis DE, Gitzendanner MA, Strenge DD, Soltis PS (1997) Chloroplast DNA intraspecific phylogeography of plants from the Pacific Northwest of North America. Plant Syst Evol 206:353–373. doi:10.1007/BF00987957
Souza-Chies TT, Bittar G, Nadot S, Carter L, Besin E, Lejeune B (1997) Phylogenetic analysis of Iridaceae with parsimony and distance methods using the plastid gene rps4. Plant Syst Evol 204:109–123. doi:10.1007/BF00982535
Stecconi M, Marchelli P, Puntieri J, Picca P, Gallo LA (2004) Natural hybridization between a deciduous (Nothofagus antarctica, Nothofagaceae) and an evergreen (N. dombeyi) forest tree species as evidenced by morphological and isoenzymatic traits. Ann Bot (Lond) 94:775–786. doi:10.1093/aob/mch205
Taberlet P, Gielly L, Pautou G, Bouvet J (1991) Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol Biol 17:1105–1109. doi:10.1007/BF00037152
Vendramin GG, Anzidei M, Madaghiele A, Bucci G (1998) Distribution of genetic diversity in Pinus pinaster Aitas revealed by chloroplast microsatellites. Theor Appl Genet 97:456–463. doi:10.1007/s001220050917
Villagrán C (1991) Historia de los bosques templados del sur de Chile durante el Tardiglacial y Postglacial. Rev Chil Hist Nat 64:447–460
Weising K, Gardner RC (1999) A set of conserved PCR primer for the analysis of simple sequence repeat polymorphisms in chloroplast genomes of dicotyledonous angiosperms. Genome 42:9–19. doi:10.1139/gen-42-1-9
Wright S (1978) Evolution and the genetics of populations, vol. 4 Variability within and among natural populations. The University of Chicago Press, Chicago
Acknowledgements
The authors wish to thank A. Aparicio, M. M. Azpilicueta, M. Huentú, F. Izquierdo, A. Martínez, A. Martinez Meier and M. Sá for their help in the bud collection. This research was financed by Instituto Nacional de Tecnología Agropecuaria (INTA) through the project PATNO13 “Productividad y efectos ambientales en ñirantales”. The ABI3100 sequencer was acquired in the frame of the project PME70 (ANPCyT) between the Universidad Nacional del Comahue - CRUB Bariloche and INTA EEA Bariloche.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pastorino, M.J., Marchelli, P., Milleron, M. et al. The effect of different glaciation patterns over the current genetic structure of the southern beech Nothofagus antarctica . Genetica 136, 79–88 (2009). https://doi.org/10.1007/s10709-008-9314-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10709-008-9314-2