
Abstract
Extremely low frequency magnetic field (ELF-MF), as a ubiquitous ecophysiological factor, has been determined to influence the health of human and animals worldwide. In this study, we focused on the effects of ELF-MF on growth performance and digestive enzyme activity in juvenile tilapia, Oreochromis niloticus. Totally 450 fish were exposed to ELF-MF (50 Hz; 0, 30, 100, 150 and 200 μT) for 30 days, respectively. The effects on growth and digestion were monitored on the 10th, 20th, and 30th day after induction (immediate effect), as well as 20th day after cessation of exposure (delayed effect). The activities of pepsin and intestinal protease of tilapia were significantly decreased after exposure in certain intensities of ELF-MF, but recovered when the ELF-MF exposure was removed 20 days later. Similar effects were also observed in the growth parameters, moreover interestingly, the specific growth rate (SGR) even represented a compensatory growth. To our knowledge, the present research provides the first determination that a certain extent of ELF-MF exposure could inhibit the growth and digestion of fish, and cause compensatory restorations after being removed upon a time.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The extremely low frequency (ELF, <300 Hz) magnetic field (MF) is produced by residential electric-equipment, nearby power and overhead high voltages transmission lines and domestic installations; as for fish underwater, there are also underwater cables, offshore energy devices, hydroelectric dams constructed in fish habitats (Hartwig et al. 2009). Basically all the power-line frequency MFs are at about 50 Hz (in Europe and Asia) or 60 Hz (in North America) (Fatigoni et al. 2005). As is reported, general public exposure to power-line frequency MF is on average <0.01 μT (Kaune et al. 2002a, b; Legros et al. 2012). Several reports have demonstrated the ELF-MF around us will affect animal behavior and also the circadian rhythm of mammals (Kavaliers et al. 1984; Salzinger et al. 1990; Chung et al. 2003; Janac et al. 2012). Fransson et al. (2001) even showed that changes of MF around European songbirds in migratory condition could result in an increasing body mass. There also have been many articles concerning adverse influences of MFs in fish (Cameron et al. 1985; Zimmerman et al. 1990; Formicki and Winnicki 1998), which imply a high sensitivity of fish against MFs. However, the details and mechanism of the ELF-MF on fish are still confusing (Davis 1962; Juutilainen 2005). Fish are relatively sensitive to the changes in their surrounding environment and Tilapia (Oreochromis niloticus) is a freshwater fish with the characteristic of rapid growth, short breeding circle and low cost. Therefore, we selected Tilapia as the objects to indicate exposed effects of ELF-MF on growth performances in living system.
Furthermore, digestive enzymes present in various locations along the digestive tract and their activities have direct and evident relationship with biological growth and development (Kolkovski et al. 1993). Thus, we studied the effects of a 30-day exposure to ELF-MF (50 Hz; 0, 30, 100, 150 and 200 μT) on growth performance (morphological characteristics, specific growth rate, and condition factor) and activities of digestive enzymes (pepsin and intestinal protease) in tilapia. Then we collected the changes induced by ELF-MF on the 10th, 20th, and 30th day (immediate effect), as well as the 20th day after cessation of exposure (delayed effect). The results would provide us new insights about the functions of ELF-MF exposure on the growth and digestion of tilapia.
Materials and methods
Source of ELF-MF
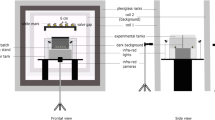
The ELF-MF exposure system used in this experiment was generated by a device designed by the China Electric Power Research Institute (Wuhan, China). The double-wrapped device was composed of a pair of Helmholtz coils (2 m in diameter, with a coil separation of 1 m) in a Magnet Lab. And the coils were insulated with a soft copper wire 1.5 mm in diameter. The experimental fish were placed in the relatively homogeneous ELF-MF produced by the equal and parallel 50 Hz alternating current while the sham exposure was conducted using the equal and opposite 50 Hz alternating current (Fig. 1a). During the exposure, the fish were all placed in the center of the coils and the alternating-current MF exposure was conducted repeatedly all day long for 30 days.

Schematic of a the tilapia exposed to ELF-MF and b the experimental procedure
Animals and treatments
The experiment was carried out between June and July 2012 in Wuhan High Voltage Institute, China Electric Power Research Institute (Wuhan, China). Tilapia (Oreochromis niloticus) with initial weight of 12.0 ± 0.8 g were acquired from an aquatic seed field (Yingshan, Huanggang, Hubei, China). Before onset of experiments, fish were weighed in batches first and then acclimated to the experimental conditions for 2 weeks. Then, 450 tilapia in good conditions were randomly distributed in 15 aquariums (n = 30) and divided into five different groups, each consisted of three individual replicate aquariums. The five groups respectively underwent ELF-MF exposure at 0, 30, 100, 150 and 200 μT. All the fish were maintained in a flow-through, dechlorinated water system of 15 glass aquariums (1.2 m × 0.6 m × 0.7 m, 360 L) which were kept at 27–29 °C with dissolved oxygen greater than 5 mg/L, under a Light/Dark 14:10 photoperiod. The experimental fish were fed twice daily (10:00 and 16:00) with approximate 0.5 g/fish, equivalently at a feeding rate of 3–5 % body weight of fish per day. Food leftover was removed by siphon. The fish were not sexed as the gonads had not yet been formed. The fish protocol for this study was approved by the Animal Care and Use Committee of Hubei Province in China and by the ethics committee of Huazhong University of Science and Technology.
Detection of growth performances and digestive enzyme activities
Sample preparation
The growth indicators and digestive enzyme activity parameters of tilapia were separately examined on the 10th, 20th, 30th day after induction based on the data of preliminary experiment and the exposure was ceased on the 30th day as the immediate effect of ELF-MF. Moreover, we also detected the indicators mentioned above on the 20th day after exposure ceased as the delayed effect. Before each sampling, 9 fish selected from every group were starved for approximated 24 h and then evaluated in growth performances. Subsequently they were killed on ice and dissected immediately, with stomachs and intestines separated for the following experiments. After that, the stomachs and intestines were rinsed, weighed and kept by groups before homogenizing separately in 9 volumes of ice-cold saline. The homogenates were then centrifuged at 6,000 rpm for 20 min at 4 °C. The supernatants containing the enzymes were stored at−70 °C until used.
Growth performances
Morphological characteristics were reflected totally in the changes of body length, body weight and body height. Changes of the morphological characteristics were examined in the afternoon (15:00–17:00) of the 10th, 20th, 30th exposure day and the 20th after exposure ceased, respectively.
Weight gain was recorded as the difference between mean initial weight and mean final weight by using a digital balance. Specific growth rate (SGR) and the condition factor (K) were used to demonstrate the growth performances of young fish.
where W1 is the final body weight (g), W0 is the initial body weight (g), and t is the experimental period in days (Brown 1957).
where W is the mean body weight (g) of each sample, L is the mean body length (cm) of each sample.
Activities of digestive enzyme
The proteolytic activities of crude enzyme extract was determined using the folin-phenol reagent method, which is based on the casein hydrolysis assay of Kunitz (Kunitz 1947) and immediately modified by Walter (Walter 1984).
Statistical analysis
Statistical analysis was performed with SPSS 13.0 package (SPSS Inc., Chicago, IL, USA). All data were presented as mean ± SD (n = 9 fish per group). The differences between exposed and sham-exposed were tested using ANOVA. LSD tests were used for multiple comparisons. Results were considered statistically significant if P < 0.05.
Results
Effects of ELF-MF on the morphological characteristics in tilapia
The effect of ELF-MF exposure on the morphological characteristics is shown in Fig. 2. As is shown in Fig. 2a, body length was significantly decreased after exposed in 30, 100, 150 and 200 μT ELF-MF on the 20th, 30th and 50th days in comparison with the sham group (20th day: p = 0.001, 0.001, 0.001 and 0.003; 30th day: p = 0.001, 0.000, 0.000 and 0.000; 50th day p = 0.006, 0.001, 0.000 and 0.024, respectively). Similarly, the body weights of all exposed groups were significantly lower than those of sham exposure (20th day: p = 0.000, 0.000, 0.000 and 0.000; 30th day: p = 0.000, 0.000, 0.000 and 0.000; 50th day p = 0.002, 0.000, 0.000 and 0.000, shown in Fig. 2b). However, the significant decline of body height happened in 150 μT group (20th, p = 0.005) and 30, 100, 150 μT groups (30th day: p = 0.016, 0.004, 0.001 and 50th day: p = 0.032, 0.009, 0.002, Fig. 2c). For the duration of the experiment, the average of body length, body weight and body height in 150 μT exposed group seemed to be markedly decreased compared with other groups. Interestingly, the 200 μT exposed group revealed a compensatory growth without the influence of ELF-MF, which led to slight rise in the average of morphological characteristics.

The effects of continuous 30-day exposure to ELF-MF (50 Hz; 0, 30, 100, 150 and 200 μT) on the morphological characteristics of tilapia, including a body length, b body weight, c body height. The ELF-MF exposure was ceased on the 30th exposure day, and the 50th day in the figure meant 20 days after exposure. Each bar represents the mean ± SD (n = 9 fish per group). *p < 0.05 and **p < 0.01 indicate significant and highly significant differences compared with the control group, respectively (ANOVA and LSD range tests)
Effects of ELF-MF on SGR and K in tilapia
A significant interaction between ELF-MF induction and growth was observed, as revealed by the SGR and K values (Fig. 3).

The effects of continuous 30-day exposure to ELF-MF (50 Hz; 0, 30, 100, 150 and 200 μT) on a specific growth rate (SGR) and b condition factor (K) of tilapia. The ELF-MF exposure was ceased on the 30th exposure day, and the 50th day in the figure meant 20 days after exposure. Each bar represents the mean ± SD. *p < 0.05 and **p < 0.01 indicate significant and highly significant differences compared with the control group, respectively (ANOVA and LSD range tests)
The decrease in SGR existed in an earlier period (from the10th day) than the changes of morphological characteristics (Fig. 3a). The SGR indicators of all exposed groups were significantly lower than the sham exposed group on the 10th, 20th and 50th days except the 200 μT on the 10th day (10th day: p = 0.018, 0.006, 0.004 and 0.081; 20th day: p = 0.000, 0.000, 0.000 and 0.000; 50th p = 0.000, 0.000, 0.000 and 0.000). Moreover, on the 30th day, only the 150 μT group got a significant descending in SGR (p = 0.542, 0.291, 0.000 and 0.542). On the contrary, the K indicator revealed a significant increase on the 20th exposure day (p = 0.000, 0.010, 0.003 and 0.019) while no obvious difference was discovered on the 10th exposure day compared with the sham group (p = 0.782, 0.756, 0.809 and 0.849; Fig. 3b). On the 30th day, the K indicator of 100, 150 and 200 μT groups were significantly higher than the sham group (p = 0.000, 0.002 and 0.005). Meanwhile, a significant increase in K indicator was also observed in the 30, 100 and 150 μT exposed groups on the 50th day (p = 0.011, 0.023 and 0.001).
Effects of ELF-MF on digestive enzyme activities in tilapia
The effect of ELF-MF on the enzyme activity of tilapia largely depended on the intensity of magnetic induction. During the ELF-MF exposure, pepsin activities of all exposed groups were significantly reduced (10th day: p = 0.000, 0.000, 0.000 and 0.008; 20th day: p = 0.004, 0.000, 0.000 and 0.001; 30th day p = 0.001, 0.000, 0.000 and 0.003, respectively). As for the intestinal protease activities, significant decreases were also observed in all the treated groups on the 10th, 20th and 30th exposure days (10th day: p = 0.003, 0.001, 0.000 and 0.009; 20th day: p = 0.004, 0.002, 0.001 and 0.004; 30th day p = 0.004, 0.002, 0.000 and 0.000, respectively). But on the 50th day, only the decrease in intestinal protease activity of 150 μT group was found a significantly difference (p = 0.041) while no significant variation was observed in pepsin activities of all exposed tilapias in comparison with the sham-exposed tilapias (p = 0.551, 0.157, 0.092 and 0.161; Fig. 4).

The effects of continuous 30-day exposure to ELF-MF (50 Hz; 0, 30, 100, 150 and 200 μT) on the digestive enzyme activities of a pepsin and b intestinal protease in tilapia. The ELF-MF exposure was ceased on the 30th exposure day, and the 50th day in the figure meant 20 days after exposure. Each bar represents the mean ± SD. *p < 0.05 and **p < 0.01 indicate significant and highly significant differences compared with the control group, respectively (ANOVA and LSD range tests)
Discussion
Under the experimental conditions, the results of the present study indicate that the exposed levels of ELF-MF significantly affect the growth performance and digestive enzyme activities of tilapia. In the current study, tilapias experienced exposure for 30 days and restoration for 20 days, which can demonstrate the immediate effects and the delayed effects of ELF-MF well.
Janac et al. (2012) have reported that the biological effects of ELF-MF depend on the characteristics (type, frequency, intensity, and exposure duration) of the applied MF and other factors (Janac et al. 2012). Prior studies point out the importance of exposure duration in the expression of biological responses (Lai and Carino 1999; Janac et al. 2012). A study showed that the intensity and duration of alternating-current MF exposure interact; increasing the duration of exposure results in effects observed at lower intensities (Lai and Carino 1999). Therefore, the intensity and exposure duration of ELF-MF play important roles in the biological effects production; proper intensity and exposure duration can lead to the changes in living bodies. Furthermore, alternating-current MF exposure contributes to the delayed hatching of zebrafish (Skauli et al. 2000). Moreover, exposure to MFs can delay the development of fertilized Medaka fish embryo at 60 Hz and 100 μT, as well as the sea urchin embryo (Cameron et al. 1985; Zimmerman et al. 1990). Similarly in our study, ELF-MF induction can decrease the body length, body weight and body height in lower intensities, and morphological characteristics and pepsin activity both performed dose-independent feature in lower intensities groups (30, 100 and 150 μT). In other words, MF exposure could slow down the growth of fish.
Furthermore, interesting recoveries of growth indicators after continuous ELF-MF exposure ceased were observed in the present experiment. However, we have not compared the differences between the following points: (i) removal of ELF-MF at 30th day and (ii) extension the ELF-MF to 50th day, therefore, whether the growth indicators will restore or not because of the removal of ELF-MF cannot be decided. However, all the growth indicators are still in a lower level than the sham group that confirms the inhibitory damage by ELF-MF. Therefore ELF-MF can suppress growth of tilapias. This is consistent with other published studies to date (Cameron et al. 1985; Zimmerman et al. 1990). However, whether the removal of ELF-MF will cause the recovery of growth indicators need further study.
Besides, digestive enzyme activity is an important index of digestive function, which determines the nutritional digestion and absorption ability of organisms and thus corresponds to biological growth and development rate (Suzer et al. 2008). In the current study, we discover that the inhibition of digestive enzyme activities caused by ELF-MF is ahead of that on growth performance. Moreover, after cessation of ELF-MF exposure, the digestive enzyme activities recovered rapidly and the SGR levels of the exposed fish were even higher than those of the sham group. The fact implies that the slower growth rate may be due to the inhibition in digestive enzyme activities. Therefore, when the exposure was stopped, the inhibiting effect caused by ELF-MF also disappeared, and then the growth rate and the digestive enzyme activities could recover rapidly towards the original levels, though slightly lower than normal. Therefore, ELF-MF can influence the activity of enzymes in animals. Similarly to our hypothesis, previous studies have shown that zebrafish embryos were monitored the asynchronous hatching when exposure to 1,000 μT ELF-MF, which was also determined to be resulted from the effect of ELF-MF on the secretion of embryonic hatching enzyme (Iuchi and Yamagami 1976; Janac et al. 2012). Moreover, it has been shown that separate MFs exposure during the early ontogenesis decreased amyloytic activity and enzyme-substrate affinity in the intestine of roach (Rutilus rutilus) underyearlings (Golovanova and Talikina 2006). Recently research also indicates that the ELF-MF exposure affect the enzyme-substrate affinity (Golovanova et al. 2013). These facts could give us answers about the catch-up growth or restoration of digestive enzyme activities when the exposure has been ceased after the 30th day. However, further experiments with appropriate methods are still needed to determine the mechanism of the effect of ELF-MF on digestive enzyme activities.
In this article, the influence of ELF-MF induction in feeding behavior was not systematically recorded in the current experiment. However, during the exposure, the exposed fish seemed to be less active than the sham group and tended to stay near the bottom of the aquarium. This phenomenon was consistent with the previous study conducted in fish (Mal 2005), even in rats and Mongolian gerbils exposed to magnetic fields at 50 Hz or 60 Hz (Salzinger et al. 1990; Janac et al. 2012).
It can be concluded that a certain extent of ELF-MF exposure may inhibit the growth and activities of digestive enzyme of tilapia, however, if ceased in time, the growth and digestion would recover rapidly to the native levels and cause compensatory growth in a period. In our present study, ELF-MF acts as an ecophysiological factor that regulates the immediate and delayed effects on growth performance and digestive enzyme activities. Nonetheless, due to the limited duration of our experiment, the subsequent, longer-term effects of ELF-MF must be examined in a future study.
References
Brown M (1957) Experimental studies on growth. In: Brown M (ed) The physiology of fishes. Academic Press, New York, pp 361–400
Cameron I, Hunter K, Winters W (1985) Retardation of embryogenesis by extremely low frequency 60 Hz electromagnetic fields. Physiol Chem Phys Med NMR 17(1):135–138
Chung MK, Kim JC, Myung SH, Lee DI (2003) Developmental toxicity evaluation of ELF magnetic fields in Sprague–Dawley rats. Bioelectromagnetics 24(4):231–240. doi:10.1002/bem.10114
Davis L (1962) Bibliography of biological effects of magnetic fields
Fatigoni C, Dominici L, Moretti M, Villarini M, Monarca S (2005) Genotoxic effects of extremely low frequency (ELF) magnetic fields (MF) evaluated by the Tradescantia-micronucleus assay. Environ Toxicol 20(6):585–591
Formicki K, Winnicki A (1998) Reactions of fish embryos and larvae to constant magnetic fields. Ital J Zool 65(S1):479–482
Fransson T, Jakobsson S, Johansson P, Kullberg C, Lind J, Vallin A (2001) Bird migration: magnetic cues trigger extensive refuelling. Nature 414(6859):35–36
Golovanova I, Talikina M (2006) On the impact of low concentrations of chlorophos in the period of early ontogenesis on digestive carbohydrases of underyearlings of roach Rutilus rutilus. J Ichthyol 46(5):404–408
Golovanova I, Filippov A, Krylov V, Chebotareva YV, Izyumov YG (2013) Effect of a magnetic field and copper upon activity of hydrolytic enzymes in roach (Rutilus rutilus) underyearlings. J Ichthyol 53(3):225–230
Hartwig V, Giovannetti G, Vanello N, Lombardi M, Landini L, Simi S (2009) Biological effects and safety in magnetic resonance imaging: a review. Int J Environ Res Public Health 6(6):1778–1798
Iuchi I, Yamagami K (1976) Induction of a precocious secretion of the hatching enzyme in the rainbow trout embryo by electric stimulation. Zool Mag 85(3):273–277
Janac B, Selakovic V, Raus S, Radenovic L, Zrnic M, Prolic Z (2012) Temporal patterns of extremely low frequency magnetic field-induced motor behavior changes in Mongolian gerbils of different age. Int J Radiat Biol 88(4):359–366. doi:10.3109/09553002.2012.652725
Juutilainen J (2005) Developmental effects of electromagnetic fields. Bioelectromagnetics 26(S7):S107–S115
Kaune W, Dovan T, Kavet R, Savitz D, Neutra R (2002a) Study of high- and low-current-configuration homes from the 1988 Denver childhood cancer study. Bioelectromagnetics 23(3):177–188
Kaune W, Miller M, Linet M, Hatch E, Kleinerman R, Wacholder S, Mohr A, Tarone R, Haines C (2002b) Magnetic fields produced by hand held hair dryers, stereo headsets, home sewing machines, and electric clocks. Bioelectromagnetics 23(1):14–25
Kavaliers M, Ossenkopp KP, Hirst M (1984) Magnetic fields abolish the enhanced nocturnal analgesic response to morphine in mice. Physiol Behav 32(2):261–264
Kolkovski S, Tandler A, Kissil GW, Gertler A (1993) The effect of dietary exogenous digestive enzymes on ingestion, assimilation, growth and survival of gilthead seabream (Sparus aurata, Sparidae, Linnaeus) larvae. Fish Physiol Biochem 12(3):203–209
Kunitz M (1947) Crystalline soybean trypsin inhibitor II. General properties. J Gen Physiol 30(4):291–310
Lai H, Carino M (1999) 60 Hz magnetic fields and central cholinergic activity: Effects of exposure intensity and duration. Bioelectromagnetics 20(5):284–289
Legros A, Corbacio M, Beuter A, Modolo J, Goulet D, Prato F, Thomas A (2012) Neurophysiological and behavioral effects of a 60 Hz, 1,800 μT magnetic field in humans. Eur J Appl Physiol 112(5):1751–1762
Mal AO (2005) Effect of the extremely low frequency, electromagnetic field on some behavioral aspects of Oreochromis Spilurus. Egypt J Aquat Res 31(2):373–382
Salzinger K, Freimark S, McCullough M, Phillips D, Birenbaum L (1990) Altered operant behavior of adult rats after perinatal exposure to a 60-Hz electromagnetic field. Bioelectromagnetics 11(2):105–116. doi:10.1002/bem.2250110202
Skauli KS, Reitan JB, Walther BT (2000) Hatching in zebrafish (Danio rerio) embryos exposed to a 50 Hz magnetic field. Bioelectromagnetics 21(5):407–410. doi:10.1002/1521-186x(200007)21:5<407::aid-bem10>3.0.co;2-v
Suzer C, Çoban D, Kamaci HO, Saka Ş, Firat K, Otgucuoğlu Ö, Küçüksari H (2008) Lactobacillus spp. bacteria as probiotics in gilthead sea bream (Sparus aurata, L.) larvae: effects on growth performance and digestive enzyme activities. Aquaculture 280(1):140–145
Walter H (1984) Proteinases: methods with hemoglobin, casein and azocoll as substrates. In: Bergmeyer H (ed) Methods of enzymatic analysis. Verlag Chemie, Weinheim, pp 270–277
Zimmerman S, Zimmerman AM, Winters WD, Cameron IL (1990) Influence of 60-Hz magnetic fields on sea urchin development. Bioelectromagnetics 11(1):37–45
Acknowledgments
This study was supported by Specialized Research Fund for the Doctoral Program of Higher Education (No. 20130142110016) and Science and Technology Project of SGCC (SG0860). The authors extend their gratitude to the China Electric Power Research Institute and Wuhan High Voltage Institute for the laboratory and experimental geomagnetic activity data.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Li, Y., Ru, B., Liu, X. et al. Effects of extremely low frequency alternating-current magnetic fields on the growth performance and digestive enzyme activity of tilapia Oreochromis niloticus . Environ Biol Fish 98, 337–343 (2015). https://doi.org/10.1007/s10641-014-0263-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-014-0263-6