
Abstract
Controls on chemical weathering, such as bedrock geology, runoff, and temperature, are considered to be the primary drivers of Si transport from the continents to the oceans. However, recent work has highlighted terrestrial vegetation as an important control over Si cycling. Here we show that at the regional scale (Southern New England, USA), land use/land cover (LULC) is an important variable controlling the net transport of Si from the land to the sea, accounting for at least 40% of dissolved Si (DSi) fluxes. A multiple linear regression model using average DSi fluxes from 25 rivers (>2,300 observations) shows the percent forest cover, as well as development and agricultural land use, to be significant (p < 0.05) drivers of DSi flux. This was true regardless of watershed size and lithology. Furthermore, forest cover is significantly negatively correlated, while development is significantly positively correlated, with Si concentrations and fluxes. We hypothesize that these relationships are due to several mechanisms, specifically the ability of terrestrial vegetation to store large amounts of Si within its biomass, the altered watershed hydrology that accompanies LULC change, and the capability of urban regions to serve as sources of Si to aquatic systems. Thus, we conclude that anthropogenic activities may be directly perturbing the global Si cycle through land use change and we offer a conceptual model which highlights a new approach to understanding the non-geochemical controls on Si fluxes.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
It is well documented that humans have directly increased the flux of nitrogen (N) and phosphorus (P) to coastal systems through a variety of mechanisms including fertilization, cultivation of nitrogen fixing crops, wastewater, and fossil fuel burning (Galloway 1998). While direct human impacts on N and P have appropriately received research attention, another important nutrient, silica (Si), has largely gone unexamined. While some attention has been given to indirect human impacts, such as eutrophication and the artificial lake effect on silica cycling along the land–ocean continuum (Conley et al. 2000; Humborg et al. 2000; Danielsson et al. 2008; Triplett et al. 2008), it is generally assumed that more direct human activities, such as land use change, have not altered the flux of Si to the coastal ocean.
Si is the second most abundant element in the lithosphere, coming into solution during chemical and mechanical weathering, and transported via rivers to the ocean. Rivers are the largest source of Si to marine waters, transporting approximately 80% of the total Si entering the ocean, or roughly 5.1 Tmole each year (Treguer et al. 1995). Much research has gone into understanding the controls on Si export from continents to oceans, and it is well known that several abiotic geochemical factors control these fluxes, particularly lithology, temperature, runoff, and slope (Drever 1994; Tipper et al. 2006; West et al. 2005; Gaillardt et al. 1999; Jansen et al. 2010). However, many of these studies have conflicting results and the ultimate controls over Si fluxes to marine waters remain uncertain.
The amount of Si entering marine waters directly impacts the abundance and community composition of phytoplankton species assemblages. Diatoms, the most common phytoplankton species in temperate coastal waters, require silica to form their cell walls, or frustules (Nelson et al. 1995). Diatoms require the same amounts of Si as N on a molar basis (Turner et al. 1998). In the presence of excess N and P, diatoms bloom until no more dissolved silica (DSi) is available in the water column, at which point non-diatom species bloom (Officer and Ryther 1980; Anderson et al. 2002). Thus, concentrations of Si in the marine system play a crucial role in marine trophic interactions. In addition, compared to other phytoplankton species, diatoms, and their associated carbon, are efficiently exported to the deep ocean, playing a major role in global carbon sequestration (Treguer et al. 1995).
While diatoms are typically highlighted for their Si-requiring biology, Si is also a ‘quasi-essential’ nutrient for a wide variety of terrestrial vegetation, particularly several species of grasses and trees (Epstein 1999). In fact, the same order of magnitude of Si is fixed annually on land as in the oceans (~200 Tmole year−1), highlighting the importance of the terrestrial biosphere in the global Si cycle (Conley 2002). In Si-accumulating plants, Si makes up between 1 and 10% dry weight, values higher than several other plant macronutrients, including N and potassium (K) (Epstein 1994). Si is used by vegetation to maintain cell structure and protect against environmental stressors, such as desiccation and predation (Epstein 1994).
The large quantities of Si within plant tissues results in terrestrial vegetation playing an important role in controlling the flux of Si to rivers, as vegetation can actively accumulate Si, thus delaying its export from land to water (Struyf and Conley 2009). For example, in a mostly forested watershed in Southern New England, the spring decline of river DSi concentrations was attributed to terrestrial vegetation uptake (Fulweiler and Nixon 2005). On the other hand, the biogenic Si found within plants is rapidly recycled throughout various terrestrial ecosystem pools because dissolution rates of biogenic Si can be an order of magnitude higher than mineral silicates (Bartoli and Wilding 1980; Fraysse et al. 2009). This enhanced solubility of plant phytoliths has been shown to control the concentrations of DSi in runoff waters (Street-Perott and Barker 2008). Recently, a study of extreme deforestation in Northern New England, found significant increases in the export of DSi from a watershed when the dead plant material remained on site (Conley et al. 2008). Similarly, using Ge/Si ratios, Derry et al. (2005) concluded that the vast majority of Si exported in rivers draining highly-weathered basalts in Hawaii had cycled through vegetation. All of these studies demonstrate the ability of terrestrial Si cycling to influence the rates and magnitudes of Si exported to aquatic ecosystems.
Historical and on-going land use changes are well known threats to biodiversity, ecosystem services, nutrient cycling, and water resources on both local and global scales (Foley et al. 2005). Globally, only 21% of indigenous forests still exist, as a result of increases in human population and the conversion of land for agricultural use (Conley et al. 2008). Locally in New England, land use change has been extreme for the past 350 years, consisting of near-complete deforestation, followed by agricultural land use, and subsequent urbanization (Foster et al. 1998). Until very recently (Conley et al. 2008; Struyf et al. 2010; Struyf and Conley in press), the impact of land use change on silica export from the terrestrial to the aquatic landscape has been ignored. Considering the rate at which land use is changing and the control that terrestrial vegetation exhibits on Si, it is likely that human perturbations of vegetative cover accompanying changes in land use is altering the rates and timing of Si as it is transported from terrestrial to aquatic ecosystems. The objective of this research was to determine the influence of dominant land use/land cover (LULC) on riverine Si export from the terrestrial to the aquatic ecosystem. We hypothesized that watershed LULC influences DSi export in rivers, impacting the magnitudes and timing of Si export during its transport from land to the coastal ocean.
Methods
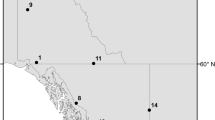
River DSi fluxes were analyzed at 25 sites throughout Southern New England from watersheds with a variety of LULC (Fig. 1; Table 1). DSi data was obtained from the United States Geological Survey (USGS) National Water Information System program (http://waterdata.usgs.gov/nwis). While all real-time streamflow gages in Connecticut (CT) and Massachusetts (MA) were considered for this analysis (175 sites throughout MA and CT), only sites with at least 15 DSi observations since 1990 were included in this study, resulting in a total of 2,370 DSi observations from 25 river sites. Watershed delineations were completed using USGS Streamstats software (http://water.usgs.gov/osw/streamstats/). LULC data was obtained from the National Land Cover Database 2001, a fifteen-class land cover classification scheme with 30 m resolution. Several LULC categories were subsequently combined and lumped into three broader categories: the ‘All Developed’ category includes Low-, Medium-, and High-Intensity Developed, the ‘All Forest’ category includes Deciduous, Conifer, and Mixed Forest and the ‘All Wetlands’ category includes both Woody and Emergent Wetlands.

Map of the 25 USGS study sites in Southern New England (Massachusetts and Connecticut) used in this study (http://waterdata.usgs.gov/nwis); Data coverage spanned the time period of 1990–2010 and in total included 2,370 values
Rivers are known to function differently depending on where they are within the longitudinal continuum (Vannote et al. 1980). For example, first-order rivers typically exhibit limited primary production due to heavy shading and, therefore, are dominated by shredders who consume course organic matter, and collectors who filter fine particles from the water column (Johnson et al. 1995). On the other hand, mid-sized rivers experience wide variability in temperatures and flow condition, and are often open to sunlight and experience high rates of primary productivity (Johnson et al. 1995). Therefore, in order to assess the impact of watershed size on the results, correlation analysis also was completed on sub-datasets that omitted the rivers with very large (>1,000 km2) and the very small (<100 km2) watersheds from the analysis. All fluxes and discharge measurements were normalized for drainage area in order to account for differences in watershed size.
New England geology is complex due to the glacial till and outwash left behind after the retreat of the Wisconsin Laurentide glacier, which covered the entire study area (Dyke and Prest 1987). While this glacial till likely contributes to some degree of soil formation, it is a mixture of various rock types and we considered it to be relatively uniform and not contributing to different Si availability across the study area. On the other hand, bedrock geology is dramatically different across the study area and no doubt contributes a great deal to soil formation and weathering products. Thus, bedrock geology was determined as a percent of each watershed in order to understand the role of lithology in determining the DSi observations. Bedrock types were combined into six main groups based on the groupings in the MA GIS data layer: mafic, granite, sedimentary, metamorphic, carbonate and calcium pelite rocks. In addition, stream temperature data was available for every corresponding DSi observation in order to discern any relationship between temperature and DSi export.
Individual regressions were completed between the 18 LULC categories described above and the average area-normalized streamflow, DSi concentrations and DSi fluxes in order to understand degree, and the manner, to which certain LULCs influence Si behavior, indicated by the slopes of the regressions and the correlation coefficients. Both streamflow and concentrations were analyzed separately from fluxes in order to better determine the mechanism for any relationship observed with Si fluxes and LULC. In addition, several multiple linear regression (MLR) models were created in order to determine the most important predictors, or variables, influencing the DSi observations. Although not included in the final model, the MLR model originally tested 27 variables, which included all 18 LULC types, number of observations, drainage area, stream temperature, and the six main geologic bedrock types for each watershed, to determine a regression line, residuals, and significance of predictors of DSi fluxes. DSi observations were transformed (Box Cox log transformation) in order to correct for non-normally distributed observations and the final model residuals were tested for and found to have normal distributions. The MLR models all passed the f-test, indicating goodness of fit and showing that the results were not due to random chance. All statistics were completed using Mathworks Matlab software with α < 0.05 to indicate significance.
Results and discussion
Streamflow and land use land cover (LULC)
The factors driving net Si transport from watersheds with various LULCs can be directly attributed to the two variables used to calculate fluxes: concentrations and streamflow. We examined both of these variables individually in order to evaluate the mechanisms driving Si export. For all of the 25 watersheds, regression analysis showed streamflow to be significantly positively correlated with development (p = 0.002 for ‘All Developed’) and significantly negatively correlated with forest cover (p = 0.0073 for ‘All Forest’) (Table 2). The correlation between streamflow and LULC is not surprising, as it is well known that land use change directly impacts watershed hydrology (Leapold 1968). Development leads to soil compaction and increased impervious surfaces, which reduce infiltration capacity on land, increasing runoff to streams and rivers (Randolph 2004; Gregory 2006). Streamflow was negatively correlated with forest cover, although less dramatically (only significant for ‘All Forests’, not individual forest types). Again, this was expected as lower streamflow in more forested sites is a result of increased water infiltration and slower runoff velocities compared to developed areas (Randolph 2004; Gregory 2006).
However, these relationships are significant only for certain LULC categories. Using the entire dataset, streamflow is only significantly correlated with developed LULC (except for “All Forest”) (Table 2). Because the smallest watersheds have the highest percent developed area, when these small rivers are removed from the dataset, streamflow is never significantly related to LULC (Table 2). In other words, the relationships observed between streamflow and LULC for the entire dataset is driven only by the rivers draining the smallest watersheds (<100 km2), as they are also the most developed.
Silica concentrations and land use land cover (LULC)
The relationships between LULC and average Si concentrations for all 25 watersheds shows a similar pattern to that of streamflow, with forest cover being significantly correlated with lower concentrations (p = 0.0323 for ‘All Forest’) and developed areas being associated with higher Si concentrations (p = 0.0424 for ‘Open Developed’). While developed areas are less strongly correlated with Si concentrations when all 25 rivers are included, removing the rivers draining the smallest watersheds (<100 km2) results in every intensity of development being highly correlated with Si concentrations (p = 0.006 for ‘All Developed’).
The close association between vegetated land covers and lower Si concentrations can likely be attributed specifically to terrestrial vegetation uptake. Terrestrial vegetation is a major sink of Si, annually sequestering the same order of magnitude of Si as is fixed annually by marine diatoms (~200 Tmole) (Conley 2002). Furthermore, several types of species can actively accumulate Si in its tissues at greater amounts than other macronutrients, such and N and potassium (K) (Epstein 1994). Si uptake by terrestrial vegetation results in less Si available for river export, as demonstrated by previous research in Southern New England that attributed spring declines in river DSi concentrations to terrestrial vegetative uptake of DSi during the growing season (Fulweiler and Nixon 2005). Likewise, two prior studies of temperate forests showed lower Si concentrations in leachate and soil porewaters under the tree species that accumulated more Si (Bartoli et al. 1983; Cornelis et al. 2010). Therefore, we hypothesize that the lower Si concentrations in rivers draining forested landscapes, compared to more developed landscapes is, at least in part, a result of terrestrial vegetation uptake and storage.
Si fluxes and land use land cover (LULC)
Because Si concentrations and streamflows were found to be significantly correlated with LULC, it is not surprising that the product of the two, Si fluxes, also shows a similar pattern (Fig. 2). In particular, forested lands export significantly less Si (p = 0.0036 for ‘All Forest’) than developed watersheds (p = 0.0285 for ‘All Developed’), as shown in the individual regressions between watershed percent LULC and average area-normalized Si fluxes (Table 2).

Relationship between Si fluxes and percent LULC. a As percent forest increases, Si fluxes decline. y = −1.4757x + 268.27, p = 0.0036, R = −0.5594. b As percent developed increase, Si fluxes increase. y = 1.2445x + 168.79, p = 0.0285, R = 0.4380. Developed land here does not include ‘Open Developed’ land
The mechanisms causing the observed Si fluxes are highlighted when the largest (>1,000 km2) and the smallest (<100 km2) watersheds are separately omitted from the correlation analysis. Si fluxes from the smallest basins are driven mostly by streamflows as a result of altered hydrology that accompanies land use change. Omitting the smallest watersheds results in the strongest correlations between Si fluxes and LULC, whereas omitting the largest watersheds results in the least significant relationships between Si fluxes and LULC. Thus, it is the rivers draining the larger watersheds (>100 km2) that are responsible for the majority of the relationships observed in the complete dataset. Furthermore, it is in these basins that Si concentrations, rather than streamflows, drive the observed fluxes. Nevertheless, regardless of watershed size, forests are always associated with less Si in rivers and developed land is always associated with more Si in rivers, as indicated by the slopes of the regressions (Table 2). These consistent relationships with LULC highlight that at any point along the longitudinal continuum of a river, LULC impacts Si fluxes in a similar manner.
In summary, we find Si fluxes to be significantly associated with LULC due to altered watershed hydrology and vegetation patterns associated with land use change. Both variables impact Si fluxes in the same manner, as both concentrations and streamflow are higher from rivers draining more developed watersheds, and lower in rivers draining more forested landscapes. Si concentrations are controlled in part by terrestrial primary productivity, and streamflow is controlled in large part by watershed infiltration capacity, or runoff. Thus, both biological and physical factors are responsible for the observed relationships with Si fluxes and land use. It appears that the factors controlling whether concentrations or streamflow dictate fluxes depends on the type of LULC, with Si fluxes from developed areas controlled more by streamflow (runoff) and Si fluxes from forested areas controlled more by concentrations (terrestrial primary production).
A special case—wetlands
Wetlands have recently been recognized for their role in exerting biological control over the Si cycle (Struyf and Conley 2009), but unlike every other major LULC in New England, wetland area was not shown to be a significantly correlated with streamflow, concentrations, nor Si fluxes. The lack of a signal from wetland cover was unexpected, considering that many species of wetland grasses are active Si accumulators and wetlands are places of dynamic Si biogeochemical cycling (Struyf et al. 2005). The lack of correlation between percent wetland area and Si fluxes may be due to the relatively small spatial coverage of this ecotone, compared to other LULC categories. Moreover, it is possible that this lack of correlation may be a result of our analysis including observations from all months of the year. This is relevant because, in New England, wetland growth and therefore active Si uptake begins in April with peak growth rates typically measured at the end of August or early September (Windham 2001; Wigand et al. 2004). When the late fall and winter months (November through March) were omitted from the analysis, wetland area within the watersheds was shown to be a significantly correlated with Si fluxes. During the April through October period, watersheds with the highest wetland area (>10%) had significantly lower DSi fluxes in the draining rivers compared to watershed with the lowest wetland area (<5%) (p = 0.035). This 7 month period includes the wetland growing season and indicated that wetlands retain Si during this period. In order to determine the mechanisms causing the observed decline in Si fluxes from wetlands, seasonal Si concentrations and streamflows were analyzed with respect to wetland area. Similar to fluxes, no significant regressions were found between percent wetland area and Si concentrations, or streamflows. Once again, the lack of relationships between wetlands and Si behavior arose because we included observations from all months of the year. When only the spring months are included in the analysis (April–June), Si concentrations were significantly lower in watersheds with the highest wetland area (>10%) compared to watersheds with the least wetland area (<5%) (p = 0.02). Thus, the periods when Si fluxes and Si concentrations are correlated with wetland area overlap with the most active growing season. On the other hand, wetland area was not related to streamflows in any season. This demonstrates that wetland Si sequestration may be the driver of the lower Si fluxes from watersheds with higher percentage of wetlands, rather than water retention.
This observed behavior of wetlands corresponds well to research that has shown wetlands to impact Si biogeochemistry in the same manner as impoundments, with wetlands acting as ‘leaky dams’ by slowing down river velocities and facilitating diatom production and sedimentation (Yelerton and Hackney 1986). Similarly, we find ‘open water’ LULC to be significantly negatively associated with Si fluxes when all but the smallest rivers are included in the regression analysis (p = 0.0278) (Table 2). This corresponds with the phenomenon that the abundance of lakes significantly reduces DSi transport (Conley et al. 2000). In addition, research in Belgium has shown freshwater wetlands to be large reservoirs of Si, as Si actively accumulates within wetland grasses and sediments (Struyf et al. 2005; Struyf and Conley 2009). While wetlands in North America have been far less studied, a recent study of a salt marsh in northern Massachusetts showed the marsh to be a source of dissolved Si, indicating that the wetlands are indeed large pools of Si in this region (Vieillard et al. 2011).
Modeling Si export at the regional scale
In order to determine the role of LULC plays in controlling Si fluxes, especially related to other factors, such as geology, several multiple linear regression (MLR) models were completed between Si fluxes, dominant LULC category, and dominant bedrock geology (Table 3). The first MLR model, which includes both LULC and geology (LULC_Geol), reveals that the majority of average DSi fluxes (R 2 = 0.94) can be explained by bedrock geology and LULC alone (p = 0.001) (Table 3). In the LULC_Geol model, the significant predictors of DSi fluxes included open, low and high-density developed land, all forest types, all agricultural types (pasture and cropland), and mafic rock. In order to determine the impact of LULC alone on Si fluxes, a second MLR model (GEOL) was built that included only bedrock. This bedrock-only MLR was again significant (p = 0.005) with an R 2 = 0.56. In order to test our hypothesis further, we created a third MLR (LULC) that included only LULC, omitting bedrock completely. The LULC model was significant (p = 0.027) with an R 2 = 0.69. Using results of all three models, a conservative estimate is that 40% of DSi fluxes can be attributed to LULC directly, although LULC could account for almost 70%, as suggested by our third model. Therefore, this MLR analysis reveals that Si export from watersheds on regional scales is driven in large part by LULC. This conclusion departs from more traditional understanding that weathering of the lithosphere is the main driver in river Si fluxes from the continents to the oceans (Bluth and Kump 1994, Jansen et al. 2010).
To put our results in perspective, we compared our regional average DSi flux value (69 kmol km−2 yr−1) to the continent-scale estimate by Beusen et al. (2009) and found our value to be higher than the North American average DSi flux estimate (43 kmol km−2 yr−1). Our higher value is likely due to the highly developed nature of Northeast US compared to the North American continent (Alig et al. 2004; Hobbs and Stoops 2002). In addition, the period of analysis in the two studies did not overlap; the Beusen et al. study included DSi observation from 1920 to 1990, while our study used values from 1990 to 2010. The differences between our more localized results and the large-scale estimates of Beusen et al. (2009) could be due to the major LULC changes during the last century in North America (Foster et al. 1998). In addition, the differences could be the results of changes in climate that have occurred, such as increased air temperatures (Huang et al. 2000), altered timing of peak river discharges (Hodgkins et al. 2003) and decreasing ratios of snow to total precipitation (Huntington et al. 2004). As pointed out by Beusen et al. (2009), global scale estimates may not represent localized DSi behavior. While globally, lithology, climate, and runoff may be primary factors controlling DSi fluxes from river systems, our results highlight that at regional scales, such as New England, other factors become important, particularly watershed LULC. In other words, controls on DSi may be scale-dependent in nature, with lithology and discharge being major drivers at the global scale, and LULC being an important variable at the regional and local scale.
Conceptual model of LULC controls on Si export
Combining our results with those of two other studies (Conley et al. 2008; Struyf et al. 2010), we developed a conceptual model depicting how land use change can potentially impact Si fluxes from the terrestrial to the aquatic ecosystem (Fig. 3). Initially, if land use change occurs in the form of deforestation, the most common LULCs to follow is either reforestation (sub)urbanization, or agricultural land uses, all of which impact Si cycling differently. For example, deforestation initially leads to higher Si fluxes from the watershed (Conley et al. 2008). If deforestation is followed by reforestation, active plant uptake and Si accumulation within plant biomass results in lower Si concentrations and fluxes, regardless of forest type. On the other hand, if deforestation is followed by (sub)urbanization, Si fluxes will be higher in rivers due to lack of plant uptake, high runoff from impervious surfaces, and potential urban sources of Si, such as waste water treatment plant (WWTP) effluent (Clark et al. 1992, Sferratore et al. 2006). Furthermore, if the disturbed land is used for agricultural purposes, Si fluxes may be even lower than that of the reforested landscape due to depletion of Si in the soil pool, as shown in the work by Struyf et al. (2010). Agricultural soils have been found to be significantly depleted in amorphous Si (ASi) stocks compared to continuously forest sites (Clymans et al. 2011), which could ultimately result in reduced export of DSi from agricultural watersheds. On the other hand, it could be argued that agricultural land use could increase Si fluxes due to tilling and other soil disturbance activities, especially considering that New England soils have been found to have several thousands of years of soils silica available (Conley et al. 2008). However, since we have no data to support this hypothesis, we will use the results of the most current research (Struyf et al. 2010; Clymans et al. 2011) to indicate in our model that agricultural practices lead to diminished Si fluxes over time. If land use change does not occur and mature forests are left standing, Si fluxes from mature forests will be less than those from newly aggrading forests (reforestation) due to in part to slower weathering rates from older forests (Conley et al. 2008).

Conceptual model of the impact of land use change on Si fluxes. Gray boxes indicate various land use types and white boxes are the processes influencing Si cycling. Initially, if land use change occurs in the form of deforestation, the most common LULCs to follow is either reforestation, (sub)urbanization, or agricultural land uses, all of which impact Si cycling differently. Deforestation has been shown to increases Si fluxes from watersheds (Conley et al. 2008). After deforestation, development lead to increased Si fluxes, whereas reforestation and agriculture tend to result in lower Si fluxes. The thickness of the lines indicates the magnitude of change in fluxes with land use type. The thicker line from agriculture compared to forested lands indicates that agriculture leads to lower Si fluxes compared to forests (Struyf et al. 2010, Clymans et al. 2011). The thicker line from mature forests compared to newly aggrading forests (reforestation) indicates that mature forests export less Si due to slower weathering rates (Conley et al. 2008). Abbreviations: ‘WWTP’ waste water treatment plant
Initially, our results appear to differ from that of Struyf et al. (2010) and Humborg et al. (2004), who found fluxes of Si to increase with increasing percent forest cover, but agree with that of Conley et al. (2008) who found that deforestation leads to higher Si fluxes. Struyf et al. (2010) hypothesize that the difference between their findings and that of Conley et al. (2008) is a result of measurements being taken at different points in time related to when initial soil disturbance occurred; Struyf et al. (2010) conclude that their study site represents sustained long-term soil disturbance (>250 years) which explains the higher Si fluxes from forests compared to other LULCs, mostly agricultural lands. However, this hypothesis does not explain the difference with our study, as Southern New England has been highly disturbed since the 17th century. Therefore, we propose that our conclusions differ because we are comparing dissimilar LULC types. The Humborg et al. (2004) study did not include an urban land use type in the analysis and similarly, in the Scheldt basin, where Struyf et al. did their work, LULC is >80% agricultural and 11% forested, compared to our study sites, which are, on average, 8% agricultural, 54% forested and 16% developed. In other words, the Struyf et al. (2010) study mostly compares forest versus agricultural land, while our study mostly compares forest versus urban land, resulting in different ‘relative’ conclusions. Our conceptual model is complementary with the study by Struyf et al. (2010), as it shows fluxes from agricultural lands to be lower than that of forested lands, but expands upon their work by adding in the effects of urban development on Si fluxes.
Our findings that LULC is an important driver of Si export in rivers can be explained by examining Si accumulation and cycling within the terrestrial landscape. Because many species of land plants are active Si accumulators, we hypothesize that a lack of vegetation cover in developed lands essentially removes a sink of Si on the terrestrial landscape, resulting in higher export of Si to rivers in the absence of vegetation. The absence of vegetation appears to result in a more rapid delivery of Si to river waters, as observed in the more developed watersheds in this study. In addition, it is possible that more heavily developed watersheds may also act as a source of silica. Several studies have shown urban areas to be net sources of Si to rivers, as Si is found within many food items, laundry detergent, and drinking water from groundwater sources (Clark et al. 1992; Sferratore et al. 2006). From our analysis, it is unclear whether developed lands are sources of Si, or that relative to known Si sinks (vegetated landscapes), there are higher fluxes from developed regions.
Conclusion
This work identifies LULC as an important control over DSi fluxes from the continents to the sea, accounting for 40–70% of DSi fluxes. Prior work has focused on quantifying solely the geochemical controls on these fluxes, but here we demonstrate the critical nature of LULC patterns in influencing Si cycling at the land-sea interface. This analysis addresses only DSi behavior in rivers. However, significant concentrations of biogenic Si (BSi) have been found in rivers of all sizes (Humborg et al. 2006, Conley 1997). Quantifying BSi behavior in urban systems would help us better understand how anthropogenic activities impact Si biogeochemistry. Currently there is little information on biogenic Si in rivers, particularly in the Southern New England region. Future research should examine the impact of LULC change on BSi fluxes, as well as quantifying nitrogen and phosphate levels in these systems, as water column Si cycling is tied to macronutrient availability.
Scientists continue to point out the clear gap in our understanding of the linkage between the terrestrial and aquatic Si cycles (Ittekkot et al. 2006). Rivers are the central transport mechanism by which terrestrially derived Si reaches coastal systems, and therefore, is an appropriate place to focus research attention pertaining to Si cycling. The research presented here offers a new manner of understanding Si transport from terrestrial to the marine ecosystems; at regional scales, LULC appears to be an important factor influencing DSi concentrations and the ultimate export to coastal systems. In other words, anthropogenic activities are directly influencing Si cycling at the land-sea interface. While land use change is already recognized as a threat to ecosystem integrity and function at regional (Foster and Aber 2004) and global scales (Foley et al. 2005), this study confirms that land use change now must also be seen as a factor influencing Si biogeochemistry, altering the timing and magnitudes of Si fluxes from the land to coastal systems.
References
Alig RJ, Kline JD, Lichtenstein M (2004) Urbanization on the US Landscape: looking ahead in the 21st century. Landsc Urban Plan 69:219–234
Anderson DM, Glibert PM, Burkholder JM (2002) Harmful algal blooms and eutrophication: nutrient sources, composition, and consequences. Estuaries 24:704–726
Bartoli F (1983) The biogeochemical cycle of silicon in two temperate forest ecosystems. Environ Biogeochem Ecol Bull 35:469–476
Bartoli F, Wilding LP (1980) Dissolution of biogenic opal as a function of its physical and chemical properties. Soil Sci Soc Am J 44:873–878
Beusen AHW, Bouwman AF, Durr HH, Dekkers ALM, Hartman J (2009) Global patterns of dissolved silica export to the coastal zone: results from a spatially explicit global model. Glob Biogeochem Cyc 23:GB0A02. doi:10.1029/2008GB003281
Bluth GJ, Kump LR (1994) Lithological and climatologic controls of river chemistry. Geochim Cosmochim Acta 58:2341–2359
Clark JF, Simpson HJ, Bopp RF, Deck B (1992) Geochemistry and loading history of phostphate and silicate in the Hudson estuary. Estuar Coast Shelf Sci 34:213–233
Clymans W, Struyf E, Govers G, Vandevenne F, Conley DJ (2011) Anthropogenic impact on amorphous silica pools in temperate soils. Biogeosciences 8:2281–2293
Conley DJ (1997) Riverine contribution of biogenic silica to the oceanic silica budget. Limnol Oceanogr 42:774–777
Conley DJ (2002) Terrestrial ecosystems and the global biogeochemical silica cycle. Glob Biogeochem Cyc 16:68.1–68.7
Conley DJ, Stalnacke P, Pitkanem H, Wilaner A (2000) The transport and retention of dissolved silicate by rivers in Sweden and Finland. Limnol Oceanogr 45:1850–1853
Conley DJ, Likens GE, Buso DC, Saccone L, Bailey SW, Johnson CE (2008) Deforestation causes increased dissolved silicate losses in the Hubbard brook experimental forest. Glob Change Bio 14:2548–2554
Cornelis JT, Ranger J, Iserentant A, Delvaux B (2010) Tree species impact the terrestrial cycle of silicon through various uptakes. Biogeochemistry 97:231–245
Danielsson A, Papush L, Rahm L (2008) Alterations in nutrient limitations—scenarios of a changing baltic sea. J Mar Sys 73:263–283
Derry LA, Kurtz AC, Ziegler K, Chadwick OA (2005) Biological control of terrestrial silica cycling and export fluxes to watersheds. Nature 433:728–731
Drever J (1994) The effect of land plants on weathering rates of silicate minerals. Geochim Cosmochim Acta 58:2325–2332
Dyke AS, Prest VK (1987) Late Wisconsinan and Holocene history of the laurentide ice sheet. Geogr Phy Quat 41:237–263
Epstein E (1994) Review: the anomaly of silicon in plant biology. Proc Natl Acad Sci 91:11–17
Epstein E (1999) Silicon. Ann Rev Plant Physiol 50:641–664
Foley et al (2005) Global consequences of land use. Science 309:570–574
Foster DR, Aber JD (2004) Forests in time; environmental consequences of 1,000 years of land use change in New England. Yale University Press, New Haven
Foster DR, Motzkin G, Slater B (1998) Land-use history as long-term broad-scale disturbance: regional forest dynamics in central New England. Ecosystems 1:96–119
Fraysse F, Pokrovsky OS, Schott J, Meunier J-D (2009) Surface chemistry and reactivity of plant phytoliths in aqueous solutions. Chem Geol 258:197–206
Fulweiler RW, Nixon SW (2005) Terrestrial vegetation and the seasonal cycle of dissolved silica in a southern New England coastal river. Biogeochemistry 74:115–130
Gaillardet J, Dupre B, Louvat P, Allegre CJ (1999) Global silicate weathering and CO2 consumption rates deduced from chemistry of large rivers. Chem Geol 159:3–30
Galloway J (1998) The global nitrogen cycle: changes and consequences. Environ Pollut 102:15–24
Gregory KJ (2006) The human role in changing river channels. Geomorphology 79:172–191
Hobbs F, Stoops N (2002) Demographic trends in the 20th Century. U.S. census bureau census 2000 special reports—series CENSR-4. US Government Printing Office, Washington, DC
Hodgkins GA, Dudley RW, Huntington TG (2003) Changes in the timing of high river flows in New England over the 20th Century. J Hydrol 278:244–252
Huang S, Pollack HN, Shen P-Y (2000) Temperature trends over the past five centuries reconstructed from borehole temperatures. Nature 403:756–758
Humborg C, Conley DJ, Rahm L, Wulff F, Cociasu A, Ittekkot V (2000) Silicon retention in river basins: far-reaching effects of biogeochemistry and aquatic food webs in coastal marine environments. Ambio 29:45–50
Humborg C, Smedberg E, Blomqvist S, Morth C-M, Brink J, Rahm L, Danielsson A, Sahlberg J (2004) Nutrient variations in boreal and subartic Swedish rivers: landscape control of land-sea fluxes. Limnol Oceangr 49:1871–1883
Humborg C, Pastuszak M, Aigars J, Siegmund H, Morth C-M, Ittekkot V (2006) Decreased silica land-sea fluxes through damming in the Baltic Sea catchment: significance of particle trapping and hydrological alterations. Biogeochemistry 77:265–281
Huntington TG, Hodgkins GA, Keim BD, Dudley RW (2004) Changes in the proportion of precipitation occurring as snow in New England (1949–2000). J Climate 17:2626–2636
Ittekkot V, Unger D, Humborg C, Tac AN (2006) The silicon cycle. Island Press, Washington, DC
Jansen N, Hartmann J, Lauerwald R, Durr HH, Kempe S, Loos S, Middelkoop H (2010) Dissolved silica mobilization in the conterminous USA. Chem Geol 270:90–109
Johnson BL, Richardson WB, Naimo TJ (1995) Past, present, and future concepts in large river ecology. BioScience 45:134–141
Leapold LB (1968) Hydrology for urban land planning—a guidebook on the hydrology effects of urban land use. United States Geologic Survey, Washington, DC
Nelson DM, Tregeur P, Brzezinski MA, Leynaert A, Queguiner R (1995) Production and dissolution of biogenic silica in the oceans: revised global estimates, comparison with regional data and relationships to biogenic sedimentation. Glob Biogeochem Cyc 9:359–372
Officer CB, Ryther JH (1980) The possible importance of silicon in marine eutrophication. Mar Ecol Prog Ser 3:83–91
Randolph J (2004) Environmental land use planning and management. Island Press, Washington, DC
Sferratore A, Garnier J, Billen G, Conley DJ, Pinault S (2006) Diffuse and point sources of silica in the Seine River watershed. Environ Sci Tech 40:6630–6635
Street-Perrott FA, Barker PA (2008) Biogenic silica: a neglected component of the coupled global continental biogeochemical cycles of carbon and silicon. Earth Surf Proc Landforms 33:1436–1457
Struyf E, Conley DJ (2009) Silica: an essential nutrient in wetland biogeochemistry. Front Ecol Environ 7:88–94
Struyf E and Conley DJ (2011) Emerging understanding of the ecosystem silica buffer. Biogeochem (in press)
Struyf E, Van Damme S, Gribsholt B, Middelburg JJ, Meire P (2005) Biogenic silica in tidal freshwater marsh sediments and vegetation (Schelde estuary Belgium). Mar Ecol Prog Ser 303:51–60
Struyf E, Smis A, Van Damme S., Garnier J, Govers G, Van Wesemael B, Conley DJ, Batelaan O, Frot E, Clymans W, Vandevenne F, Lancelot C, Goos P, Meire P (2010) Historical land use change has lowered terrestrial silica mobilization. Nat Commun. doi:10.1038/ncomms1128
Tipper ET, Bickle MJ, Galy A, West AJ, Pomiès C, Chapman HJ (2006) The short term climatic sensitivity of carbonate and silicate weathering fluxes: insight from seasonal variations in river chemistry. Geochim Cosmochim Acta 70:2737–2754
Treguer P, Nelson DM, van Bennekom AJ, DeMaster DJ, Leynaert A, Queguiner B (1995) The Silica balance in the world ocean: a reestimate. Science 268:375–379
Triplett LD, Engstrom DR, Conley DJ, Schellhaass SM (2008) Silica fluxes and trapping in two contrasting natural impoundment of the upper Mississippi River. Biogeochemistry 87:217–230
Turner RE, Qureshi N, Rabalais NN, Dortch Q, Justic D, Shaw RF, Cope J (1998) Fluctuating silicate:nitrate ratios and coastal plankton food webs. Proc Natl Acad Sci 95:13048–13051
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE (1980) The river continuum concept. Can J Fish Aqu Sci 37:130–137
West AJ, Galy A, Bickle M (2005) Tectonic and climate controls on silicate weathering. Ear Plan Sci Lett 235:211–228
Wigand C, Thursby GB, McKinney RA, Santos AF (2004) Response of spartina patens to dissolved inorganic nutrient addition in the field. J Coast Res 45:134–149
Windham L (2001) Comparison of biomass production and decomposition between phragmites australis (common reed) and spartina patens (salt hay grass) in brackish tidal marshes of New Jersey, USA. Wetlands 21:179–188
Vieillard AM, Fulweiler RW, Hughes ZJ, Carey JC (2011) The ebb and flood of dissolved and biogenic silica fluxes from a temperate salt marsh. Estuar Coast Shelf Sci. doi:10.1060/j.ecss2011.10.012
Yelerton GF, Hackney CT (1986) Flux of dissolved organic carbon and porewater through the substrate of a spartina alterniflora marsh in North Carolina. Estuar Coast Shelf Sci 22:255–267
Acknowledgments
The author would like to acknowledge the United States Geological Survey for their collection, analysis, accessibility of this data. This research was developed under STAR Fellowship Assistance Agreement no. # FP917238 awarded by the U.S. Environmental Protection Agency (EPA) to J.C. Carey. It has not been formally reviewed by EPA. The views expressed in this paper are solely those of J.C.Carey and R.W. Fulweiler, and EPA does not endorse any products or commercial services mentioned in this publication. The author would also like to thank the Department of Earth Sciences at Boston University for partial funding support of J.C Carey and Seth Binder for help with statistical analysis. This manuscript was improved through helpful comments from Andy Kurtz and two anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Carey, J.C., Fulweiler, R.W. Human activities directly alter watershed dissolved silica fluxes. Biogeochemistry 111, 125–138 (2012). https://doi.org/10.1007/s10533-011-9671-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-011-9671-2