
Abstract
Human activities have altered riverine silica cycling and diminished the supply of silica to the oceans, but few rivers have been intensively monitored to evaluate the magnitude of these changes. In this study we measured dissolved silica (DSi) and amorphous silica (ASi) fluxes into and out of two large, culturally-impacted natural impoundments of the upper Mississippi River, Lakes St. Croix and Pepin, USA. ASi sedimentation rates and sediment–water fluxes of DSi were calculated for each lake, and a mass-balance approach was used to determine in-lake ASi production. ASi from terrestrial phytoliths in the lake sediments was determined to be only partially available to biotic recycling, and in-lake ASi dissolution was small relative to the total silica budgets. The river reaches upstream of the two lakes were found to have abundant DSi, and riverine diatom production was found to contribute significant amounts of ASi to each lake. The average total phosphorus concentration in Lake Pepin is four times that in Lake St. Croix but ASi production in Lake Pepin is only 2.3 times higher than in Lake St. Croix, indicating that diatom growth in Pepin is limited by factors such as turbidity. Lake St. Croix currently traps about 10% of the inflowing total bioavailable silica (TSib = DSi + ASi) while Lake Pepin traps closer to 20% of its inflowing TSib, clearly demonstrating the importance of silica retention in lakes and reservoirs along the land–ocean continuum.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Rivers are the primary source of silicon (Si) to the oceans (Tréguer et al. 1995), where the nutrient is critical for the growth of diatoms, radiolarians, and other organisms. As human impacts to large river watersheds have affected global cycles of other nutrients (Caraco 1993), so too have those impacts altered the global silica cycle. Specifically, changes from eutrophication (Caraco 1993; Conley et al. 1993, 2000; Turner and Rabalais 1994), the construction of large dams (Friedl et al. 2004; Humborg et al. 2002, 2006), and landscape alterations including deforestation and agricultural expansion (Conley et al., unpublished data) have created an unnatural abundance of riverine diatoms and a rapid rate of diatom sequestration in reservoir sediments.
Rivers transport two categories of bioavailable Si (TSib): dissolved silica (DSi) and particulate amorphous silica (ASi). Rivers also transport mineral silicates that dissolve slowly on a biological timescale, so they are not considered to be bioavailable. Although silica is clearly an important nutrient, detailed datasets of riverine silica fluxes, especially those that differentiate DSi and ASi loads, are scarce. Some important exceptions include datasets for rivers in the Baltic and Black Sea catchments (Conley et al. 2000; Humborg et al. 1997, 2006; Teodoru et al. 2006). Here we present silica budgets for two large, natural riverine lakes in the watershed of the Upper Mississippi River: Lake St. Croix and Lake Pepin, USA. Lake St. Croix comprises the lowermost 37 km of the St. Croix River above its confluence with the Mississippi River. Lake Pepin, which is 24 km downstream of the St. Croix confluence, impounds 34 km of the Mississippi mainstem.
These two natural impoundments are unique in that they were formed over 9,000 years ago when glacial meltwater flow diminished and deltas from converging waterways partially blocked each river’s channel. They have existed in the same basins for millennia, and thus provide a clear picture of silica cycling without some of the complications posed by artificial reservoirs. Damming rivers disturbs the natural hydrology and geomorphology of the river channels and may affect silica cycling, such as by increasing silica input from newly inundated soils (Humborg et al. 2006). Those changes have not happened to Lake St. Croix and Lake Pepin, so silica cycle alterations observed in those lakes can be attributed to other factors, most specifically eutrophication. However, the natural riverine lakes are still good analogues for artificial reservoirs because of their short residence times and high sedimentation rates (Table 1), and because of their riverine character (majority of water, sediment and nutrient transport along a single linear flow axis). Since the Mississippi River is the primary supplier of bioavailable silica to the Gulf of Mexico (Turner and Rabalais 1994), silica budgets in these two lakes also have direct implications for this important marine ecosystem.
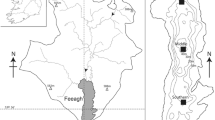
Lake St. Croix and Lake Pepin are located near one another but have very dissimilar watersheds (Fig. 1 and Table 1). The St. Croix watershed is mostly forested in the north and accommodates some agriculture and growing population centers in the south. It is a designated U.S. National Scenic Riverway managed by the National Park Service, and it has remained relatively undisturbed as compared to other large mid-continental rivers. Nonetheless, recent paleolimnological studies have shown that the total phosphorus (TP) concentration in Lake St. Croix has increased from about 20 μg l−1 before Euro-American settlement (around 1850 in this area) to 50 μg l−1 today (Edlund et al. in press).

Map of Lake St. Croix and Lake Pepin. The St. Croix River watershed (checkered) and the Lake Pepin watershed (all cross-hatched and checkered) are shown on the state map of Minnesota
In contrast, large portions of the Lake Pepin watershed are devoted to row-crop agriculture (primarily corn and soybeans). These agricultural areas have very high densities of subsurface drain tiles and ditches, which have simplified and shortened the route of water from the land surface to tributary rivers and thus changed the hydrology of the entire Mississippi River system. Moreover, a major urban center, the Twin Cities of Minneapolis and St. Paul, Minnesota, and their surrounding suburbs, lie 35 km upstream of Lake Pepin. Lake Pepin’s TP concentration before Euro-American settlement was approximately 40 μg l−1, as reconstructed from diatom analysis, and has now increased to 200 μg l−1 (Engstrom et al. in press). The St. Croix River (including Lake St. Croix) supplies about 25% of the water flow reaching Lake Pepin, and is a far smaller contributor of sediment and nutrients to Lake Pepin relative to the portions of the Mississippi that drain southern and west-central Minnesota (Kelley and Nater 2000) (Fig. 1).
These two river/lake systems are thus in close proximity to one another but reflect very different types of land-use change and consequent water quality. They provide an excellent opportunity to examine silica cycling in large rivers and impoundments, and to compare and contrast how different levels of human activity affect Si cycling in rivers and riverine lakes.
Methods
Defining the silica pools
There are two primary inputs of bioavailable silica to each lake: ASi in the inflow (ASiinflow), which could potentially dissolve and be recycled in the lake, and DSi in the inflow (DSiinflow). ASi measured in these river waters and lake sediments is predominantly in the form of diatom frustules, but can also be from inorganic forms of soil ASi (Sauer et al. 2006) or siliceous phytoliths, which are ASi structures formed by terrestrial plants. The losses of silica from each lake are via the outflow, again as ASi and DSi (ASioutflow and DSioutflow), and via ASi sequestration in the sediments (ASisedimented). During the water’s residence in the lake, further diatom production occurs, transforming DSi into ASi (ASiin-lake), while some ASi re-dissolves after diatoms die (ASidissolution) (Fig. 2).

Generalized riverine lake with dissolved and amorphous silica fluxes. ASiin-lake represents ASi that formed from DSi within the lake, predominantly by diatom growth. ASidissolution represents the amount of ASi that dissolves within the lake to become DSi, and is therefore not included in the other ASi pools
Silica inflow and outflow
To determine the mass of ASi and DSi passing through each lake, silica inputs and outputs were measured directly. Water samples were collected from the Lake St. Croix inflow and outflow and from the Lake Pepin inflow weekly from 5 April to 1 November 2004 and twice each month from 1 November 2004 to 24 March 2005. Lake Pepin outflow samples were collected approximately twice a month from 9 April 2004 to 10 November 2005. No samples were collected from the Lake Pepin outflow from November to March because of dangerous ice conditions. All water samples were taken from midstream and at a 1 m depth below the surface.
Samples were refrigerated after collection and filtered within 2–3 days. The methods used to measure concentrations of ASi in water were from Krausse et al. (1983). A 125-ml subsample was passed through a 0.4 μm polycarbonate filter (some samples with high turbidity required two filters). The filtrate was saved for DSi analysis, and the filters were stored frozen in polycarbonate tubes until ASi digestion and analysis. For the ASi digestion filters were submerged in 4 ml of 0.2 M NaOH and placed in a 100°C water bath for 20 min. Filters were then rinsed with deionized water and discarded. The solutions were cooled, neutralized with 0.6 ml of 0.5 M H2SO4, diluted to 25 ml with deionized water, and refrigerated until analysis. We recommend that in future studies a time-step correction for mineral silicates be used to measure ASi concentrations in water that is highly turbid (see the “Results and Discussion” for more discussion on this topic).
Concentrations of DSi in the filtrates (river water DSi) and in the filter digestion solutions (river water ASi) were measured as SiO2 on a Lachat QuikChem 8000 colorimeter as molybdate reactive silica, with standards ranging from 0.2 to 20 mg l−1.
River discharge
River discharge was used to convert concentrations into mass fluxes. The discharge of the Mississippi River was measured by the U.S. Army Corps of Engineers (USACE) at Lock and Dam 3, near the head of Lake Pepin. The gauging station nearest Lake St. Croix is operated by the U.S. Geological Survey (USGS) 46 km upstream of the head of the lake (where samples were taken), and these flow measurements do not include the discharge of one major ungauged tributary, the Apple River. Thus the USGS data from the upstream gauging station were proportionately increased to account for the Apple River’s flow, based on correlations of historical measured Apple River flow to mainstem flow.
Total suspended solids (TSS) data were supplied by the Metropolitan Council Environmental Services (2006).
ASi sequestration in lake sediments
The flux of ASi deposited in the sediments of the two lakes were measured on sediment samples from two previous studies, with 24 to 25 sediment cores collected from each lake (Engstrom et al. in press; Triplett et al. in press). Eight cores from Lake St. Croix and 10 cores from Lake Pepin were chosen to be “primary” cores and were dated using 210Pb, 137Cs and 14C methods, and sediment accumulation rates were calculated. The sediment and ASi accumulation rates of the un-dated (secondary) cores were estimated from dated cores using magnetic susceptibility profiles for cross-correlation and chemical analyses of “top and bottom” samples in the secondary cores. To extrapolate these accumulation rates to the entire lake, the depositional area for fine-grained sediments was delineated using bathymetric maps and field observations, and that area was divided into sections, each of which was represented by one or more of the dated cores. The sum of sediment accumulation in each of the depositional areas equals the whole-lake sediment accumulation. For the purposes of this study only the recent (uppermost) rates of sediment accumulation are considered. Details of dating and calculations can be found in Engstrom et al. (in press) and Triplett et al. (in press).
Sediment ASi concentrations were measured using a time-step digestion (Conley and Schelske 2001; DeMaster 1981). Sediments were digested in 1% Na2CO3 for 5 h at 85°C in a shaking water bath. The solutions were sub-sampled at the 3-, 4- and 5-h marks to chart the progressive silica dissolution over time in order to correct for DSi contributed by mineral silicates. Sediment ASi concentrations were multiplied by the sediment accumulation rate to produce core-specific rates of ASi deposition that were then converted to whole-lake ASi accumulation rates as described above.
To determine the comparability of ASi results obtained by the NaOH digestion method (water column ASi) and the Na2CO3 time-step digestion method (sediment ASi), a small calibration experiment was performed. Seventeen water column samples of varying turbidity and ASi concentrations were split, filtered and digested with each method. The results from the two methods were similar (p = 0.65, paired p test). The NaOH method produced slightly higher results than the Na2CO3 method for samples with very low ASi (<0.5 mg l−1), while the Na2CO3 method produced higher results for samples with high levels of TSS. Therefore, water column and sediment ASi values obtained with these two different methods can be reasonably compared in a single mass balance relationship.
ASi from terrestrial plant phytoliths
To determine whether phytolith Si confounds the ASi results, three lake sediment samples were visually examined for phytolith preservation following Na2CO3 digestion. Known masses of dry sediment were digested following the Na2CO3 method described above and were then treated with H2O2 to remove organic matter, while a set of “control” splits were treated only with H2O2. Residual sediment from each of the two treatments (Na2CO3 and H2O2-only) was neutralized, rinsed, and quantitatively mounted on microscope slides (Battarbee 1973). At least 60 phytoliths were identified on each slide using a Leica DMLP petrographic microscope at 400× magnification, and long and short dimensions for each phytolith were measured. These dimensions were used to estimate phytolith biovolume (generally assuming cylindrical shapes), and the biovolumes were converted to phytolith ASi mass using a density of 2.0 g SiO2 cm−3 (Gallego et al. 2004).
ASi dissolution in the lakes
Two methods were used to estimate the dissolution of ASi (ASidissolution) in the lakes: (1) the excess DSi that accumulated in the hypolimnion of Lake St. Croix was measured during the 2004 growing season and (2) porewater DSi gradients were measured in sediment cores from both lakes to estimate diffusion out of the sediments. For the first method, DSi and ASi concentrations and water temperature were measured throughout the water column at three sites in Lake St. Croix near the end of the summer (3 August 2004). Each sampling site was in a different sub-basin of the lake to account for spatial variability. Because Lake St. Croix stratifies during the summer, a majority of ASi dissolution in the water column or sediments is trapped in the hypolimnion, such as occurs with phosphorus (Foy 1985). This “excess” DSi was calculated by subtracting a background DSi value from the hypolimnion concentrations. The “background” was designated as the DSi concentration at the very top of the hypolimnion at each location, as the epilimnion DSi may have been depleted by diatom growth. The excess DSi concentration was multiplied by an estimated hypolimnion volume to give the mass of DSi produced by ASi dissolution during the summer stratification period, beginning in approximately late May. Finally, the dissolution rate was extrapolated to the entire year to determine the annual ASidissolution. ASi dissolution slows significantly with decreased temperature (Iler 1979), so using the summer dissolution rate provides an upper limit on the annual dissolution. Because it is relatively shallow, Lake Pepin does not stratify for significant periods of time, so the hypolimnion method could not applied there.
For the second method, sediment–water fluxes from lake sediments were estimated from surficial sediment cores collected at three locations in Lake St. Croix at the same time as the water column profiles were measured (above). Surface cores were also retrieved from two locations in Lake Pepin. All cores were collected using a HTH Teknik gravity corer and were sectioned the same day at 0.5 cm intervals and centrifuged. The supernatant was siphoned off and analyzed for DSi, as described above, to produce porewater DSi profiles. The mass of water extracted by centrifugation was measured and used to estimate sediment porosity. The Wilke–Chang equation (1955) was used to estimate a sediment diffusion coefficient for DSi, e.g. silicic acid, and Fick’s first law of diffusion (Lerman 1979) was used to calculate silica fluxes out of the sediment:
where J is DSi flux rate across the sediment–water interface (mg m−2 s−1), D0 is the diffusion of DSi in water at 20°C, ϕ is sediment porosity (Li and Gregory 1974), and dC/dz is the DSi concentration gradient across the sediment–water interface (mg m−3 m−1). For comparison, empirically derived diffusion coefficients (Applin 1987; House et al. 2000; Lerman 1979) are within 40% of the Wilke–Chang estimation.
The flux rate (J) for each Lake St. Croix core was multiplied by the depositional area in that core’s sub-basin of the lake, thereby obtaining the mass of DSi released from the sub-basin’s sediment during 1 year. For the sub-basin in which we did not collect a surface sediment core, we used the average of the flux rates obtained from cores in other sub-basins. The sub-basin mass fluxes were summed to obtain the whole-lake annual flux of DSi from sediment. This method was also applied to Lake Pepin.
Results and discussion
Inflow and outflow Si fluxes
DSi concentrations in Lake St. Croix changed gradually over the monitoring period and followed a typical seasonal pattern of low DSi in the summer and high DSi in the winter (Fig. 3) as observed in other temperate rivers (Humborg et al. 2006). The lake’s inflow DSi concentration ranged from a minimum of 7.1 mg l−1 in early June to a maximum of 18.6 mg l−1 in January, while the outflow DSi was at a minimum of 6.6 mg l−1 in late July and a maximum of 18.1 mg l−1 in February; all concentration data are reported as mg l−1 SiO2. ASi concentrations had the inverse trend of generally higher values in spring and summer (the growing season) and low values during the winter. The lake’s inflow ASi concentration reached a minimum of 0.14 mg l−1 (below our lowest calibration standard, but above the method detection limit) in late July and a maximum of 3.0 mg l−1 in early July. Excluding the two low samples in late July, the next lowest ASi concentration was 0.23 mg l−1 in early February. The outflow had a minimum ASi concentration of 0.2 mg l−1 in February and a maximum of 1.2 mg l−1 in August. The average annual DSi and ASi concentrations in the Lake St. Croix outflow were 11.1 and 0.6 mg l−1, respectively. These concentrations are higher than the world average DSi and ASi concentrations reported previously for large rivers: 9.0 mg l−1 for DSi (Tréguer et al. 1995) and 0.47 mg l−1 for biogenic silica (Conley et al. 1997).

Concentrations of amorphous silica (ASi) and dissolved silica (DSi) in the Lake St. Croix inflow and outflow from April 2004 to March 2005
DSi concentrations in Lake Pepin also changed gradually over the year with the exception of an abrupt increase from May to June (Fig. 4). Pepin’s inflow DSi concentration ranged from 4.9 mg l−1 in late April to 17.9 mg l−1 in October, while the outflow DSi ranged from 2.5 mg l−1 in May to 20.2 mg l−1 in October. In contrast to Lake St. Croix, ASi concentrations were quite variable in Lake Pepin. Inflow DSi concentrations ranged from 0.2 mg l−1 in February to 13.3 mg l−1 in September, while the outflow ranged from 0.5 mg l−1 in July to 3.4 mg l−1 in April. The annual average DSi and ASi concentration in the Lake Pepin outflow were 13.5 and 1.5 mg l−1.

Concentrations of amorphous silica (ASi) and dissolved silica (DSi) in the Lake Pepin inflow and outflow from April 2004 to March 2005
ASi comprised 12% of the Lake St. Croix inflow of TSS on average during the monitoring year (2004–2005) and was a greater proportion of the outflow’s TSS load with a mean of 21%. Lake Pepin’s TSS inflow load was on average 8% ASi, while its outflow TSS was 17% ASi.
Uncertainty levels reported for ASi concentrations in water include analytical error (2%) and potential silica contributions from phytolith dissolution during analysis (15%). The estimated error due to phytoliths is the mean of the differences between phytolith mass in digested and undigested sediments (see “ASi from phytoliths”). In addition, the largest source of error was due to the weekly or bi-weekly (i.e. non-continuous) sampling, and was estimated as the standard error of the mean. ASi varies on short time scales due to the rapid response of diatom communities to changing nutrient levels, temperature, competition and predation, so the standard errors were large (16–25%). Uncertainty for DSi concentrations includes analytical error (1%) and the error due to periodic sampling described above (16–25%).
Lake St. Croix discharge over the period of study reached a maximum of 552 m3 s−1 in early June and a minimum of 63 m3 s−1 in December, with an annual mean of 152 m3 s−1. Flow was high during the spring (April to mid-June) after which it decreased and fluctuated around a mean of 112 m3 s−1 for the remainder of the year. The period of monitoring (2004–2005) was a typical year in terms of average annual flow and seasonal patterns of flow: the mean annual flow from 1980 to 2005 was 139 m3 s−1. During the monitoring year, Lake Pepin discharge reached a maximum of 1,586 m3 s−1 in mid-June and minima of 227 and 244 m3 s−1 in August and January, respectively, with a mean annual flow of 534 m3 s−1. Flows were higher in the spring and fall and lower in late summer and winter and this was also a typical flow year: the mean annual flow between 1988 and 2007 was 614 m3 s−1. The USGS reported the level of uncertainty at the point of discharge measurement on the St. Croix River as 10%, and the USACE reported the level of uncertainty at the point of discharge measurement on the Mississippi River as 5%. An additional 5% error was estimated to accommodate the contributions of ungauged tributaries and precipitation to and evaporation from the surface of each lake.
The concentrations of DSi and ASi in each lake’s inflow and outflow were multiplied by the corresponding daily discharges to obtain daily mass fluxes of DSi and ASi into and out of the lakes. Silica fluxes for days without silica measurements were estimated as a linear interpolation between the two nearest measurements. Total bioavailable Si (TSib) flux was calculated as the sum of DSi flux plus ASi flux. A year’s worth of daily fluxes were summed to produce the annual fluxes (Table 2).
In both lakes the DSi fluxes were much larger than the ASi fluxes and neither system appeared to be DSi-limited based on concentration data. Reported threshold values for silica uptake by diatoms are well below 0.5 mg l−1 (Rabalais et al. 1996; Schelske and Stoermer 1971), and the lowest measured DSi in this study was 2.5 mg l−1 (Pepin outflow, May 2004). However, there were seasonal low points in the DSi concentrations when diatom growth (as measured by ASi) was at a peak.
ASi flux to sediment
As reconstructed from the sediment core analyses, whole-lake ASi accumulation in Lake St. Croix was 5,600 (±1,000) t yr−1 during 1990–2001. Whole-lake ASi accumulation in Lake Pepin during 1990–1996 was an order of magnitude higher at 52,000 (±9,300) t yr−1 (Engstrom et al. in press). Although the sediment cores were collected several years prior to the 2004 monitoring year, the ASi sedimentation rates calculated from those cores are an average of several years of accumulation. Because 2004 was an average flow year for both lakes, they likely received an “average” amount of sediment; therefore, the previously determined sedimentation rates provide a reasonable estimate of ASi sedimentation in 2004. The reported levels of uncertainty for the ASisedimented terms were propagated from analytical error (2%), an estimate of uncertainty from the time-step correction in the Na2CO3 digestion method (10%), and an estimate of the uncertainty of the depositional areas (10%). An additional source of uncertainty came from the dating models that produced the sediment accumulation rates (10%) (Engstrom et al. in press; Triplett et al. in press).
ASi from phytoliths
The non-mineral ASi in the water columns and sediments of Lakes St. Croix and Pepin include several distinct components of biological origin: diatom frustules, freshwater sponge spicules, testae amoeba, and siliceous phytoliths from terrestrial plants. In addition, ASi in the form of weakly or non-crystalline silica can also contribute to ASi measurements (Sauer et al. 2006). The majority of ASi produced within these lake/river systems is in the form of diatom frustules, as very few sponge spicules were observed in microscopic scans of fresh sediments. However, there can be significant input of ASi in the form of terrestrial phytoliths to each system, as might be expected for large rivers (Cary et al. 2005; Conley 2002). Fresh phytoliths from plants readily dissolve in Na2CO3 (Saccone et al. 2007). If phytoliths were dissolved in the ASi digestions (NaOH and Na2CO3 methods), it would mean that the ASi measured in this study could not be assumed to be due entirely to in situ (diatom) productivity.
To determine whether Si from inadvertant dissolution of plant phytoliths contributed to the ASi results, phytolith preservation was visually quantified in lake sediment samples prior to and following Na2CO3 digestion. The differences between phytolith masses in three paired undigested and digested samples were 11, 15 and 18%. This analysis demonstrated that the phytoliths have undergone some dissolution but a large fraction of the phytolith mass remained intact. Visual observation of the remnant sediments provided further confirmation of low dissolution, as many phytoliths were observed to have as much detail (clear edges, sharp corners, and surface patterning) as phytoliths in undigested sediment. By contrast, Saccone et al. (2006) have shown that phytoliths taken directly from plants dissolve completely in weak Na2CO3 digestions, as does ASi in many soil types. However, soils often require a much stronger base such as NaOH to dissolve more resistant ASi. For example, weak Na2CO3 extractions of Chernozems from grasslands of the Great Plains, USA were not sufficient to extract all the ASi, and strong digestions with 0.5 M NaOH extracted two to four times more ASi (Saccone et al. 2007).
It is hypothesized that the more easily recyclable phytoliths have dissolved upstream in the landscape, leaving behind only the largest and sturdiest of phytoliths that could survive transport down the rivers, into the lakes and subsequently into the lake sediments. Furthermore, it is likely that the resistant phytoliths have higher levels of Al substitution at their surface and have reorganized their surface structures, which significantly inhibits dissolution (Iler 1979; Van Cappellan et al. 2002). Therefore, while phytoliths undeniably constitute a large flux of silica in these rivers, their silica is highly refractory and largely non-bioavailable to the freshwater ecosystem. Hence, all ASi measurements reported in this study are assumed to be dominated by diatom-produced ASi.
ASi dissolution
There was a clear buildup of DSi in the hypolimnion of Lake St. Croix, according to the three water columns we studied (Fig. 5). Background DSi concentrations were 7.4, 9.2 and 11.2 mg l−1 at sites 3Bs, 5Bs and 7Bs, respectively, and the corresponding excess DSi concentrations were 1.9, 3.2 and 1.2 mg l−1. Hypolimnion volumes for each sub-basin were calculated using a hypolimnion thickness of 3.0 m at all sites, as indicated by temperature profiles. Excess DSi concentrations were multiplied by the hypolimnion volumes for a total whole-lake ASidissolution of 380 t yr−1.

Water column profiles of dissolved silica (DSi), amorphous silica (ASi) and temperature at a representative Lake St. Croix monitoring site (core site 5B) on 3 August 2004. The dashed line indicates the DSi concentration used as background for estimating ASi dissolution
In the surface sediment cores, porewater DSi concentrations ranged from about 10 to 48 mg l−1 in Lake St. Croix and from 14 to 45 mg l−1 in Lake Pepin, while overlying lake water at the time of core collection was generally 13–14 mg SiO2 l−1 with one exception in Lake Pepin (core A3s, overlying water DSi concentration of 21 mg l−1). Porewater profiles showed very short, steep gradients of DSi concentration within the top 1–3 cm (Fig. 6). DSi concentrations were quite variable at depth in some cores, particularly those from Lake St. Croix. However, there were no discernible upward gradients to indicate porewaters at depth were interacting with overlying lake water. In some cores, e.g. Lake St. Croix 3Bs, the gradient was entirely within the top 0.5 cm, again demonstrating that sediment porewaters are quickly isolated from overlying lake water in these systems. In addition, ASi concentrations did not show a discernible peak at the top of any of the cores, indicating that there is no large loss of ASi from recently deposited sediments during burial.

Sediment concentrations of amorphous silica (ASi) and porewater dissolved silica (DSi) concentrations in three cores from Lake St. Croix and two cores from Lake Pepin. DSi concentrations at zero depths are those in the overlying lake water
DSi sediment–water fluxes estimated from Fick’s first law of diffusion were 4.7 × 10−4 to 1.1 × 10−3 mg m−2 s−1 in Lake St. Croix and 8.9 × 10−4 to 9.8 × 10−4 mg m−2 s−1 in Lake Pepin, which are low compared to fluxes from riverine lake sediments from the Danube (2 × 10−3 to 1 × 10−2 mg m−2 s−1; Friedrich et al. 2003) and sediments from the River Swale, UK (1.8 × 10−3 mg m−2 s−1; House et al. 2000). When converted to whole-lake annual fluxes, this method estimates that Lake St. Croix sediments released 400 t yr−1 of DSi to the water column, and Lake Pepin sediments released 2,200 t yr−1.
The two methods used to determine ASi dissolution in the lakes have significant uncertainty and thus provide only first-order estimates of ASi dissolution rates. The hypolimnion method neglects factors such as turbulence and upwelling at the boundary layer which can release silica to the epilimnion, although temperature and dissolved oxygen profiles do indicate the development of a robust hypolimnion. Similarly, the porewater gradient method may not be sensitive enough to capture all ASi dissolution at the sediment–water interface, and a study by Conley and Schelske (1989) showed that DSi fluxes measured from sediment incubation chambers were on average 2.2 times higher than DSi fluxes measured by the porewater gradient method. Nevertheless, the hypolimnion and porewater gradient estimates used here produced similar results in Lake St. Croix (380 t yr−1 by the hypolimnion method and 420 t yr−1 by the porewater method). Furthermore, even with an uncertainty level of 100% based on Conley and Schelske’s study, the ASi dissolution fluxes for both lakes as calculated by the porewater method are small in comparison to the other silica fluxes. For comparison, the DSi flux out of Lake St. Croix sediments is 9% of the ASi flux into the lake (ASiinflow, 4,500 (±1,100) t yr−1), while DSi flux out of Lake Pepin sediments is just 4% of the flux into that lake (ASiinflow, 49,500 (±11,000) t yr−1). For the final mass balance, we used the estimates from the porewater method in calculating Lake St. Croix’s silica budget, as that method was also used for Lake Pepin.
Just as diatoms take DSi from the water to produce their frustules of ASi, so too may dead diatoms dissolve and return DSi to the water column. For example, 75% of sedimented ASi was dissolved and recycled by diatoms in Lough Neagh from 1994 to 1997 (Gibson et al. 2000), while over 95% of marine ASi is dissolved and recycled (Tréguer et al. 1995). In contrast, Lake St. Croix and Lake Pepin have much lower rates of ASi dissolution. Low salinity, high ambient DSi concentrations, circumneutral pH and abundant cations such as Al, which may adsorb to ASi surfaces, all act to slow the rate of diatom dissolution in these systems (Dixit et al. 2001; Hurd 1983; Icenhower and Dove 2000; Iler 1979; Ryves et al. 2006). Furthermore, these lakes are very shallow relative to the oceans or large lakes, so diatoms have a much shorter water column to traverse before they reach the sediments (Sommer 1988). Finally, the sediment accumulation rates are very high in these lakes (0.5–1.4 cm yr−1 in Lake St. Croix and 1.3–4.7 cm yr−1 in Lake Pepin), such that ASi is quickly isolated from overlying waters. This rapid burial shortens the diffusional gradient at the sediment–water interface and allows sediment porewater to more quickly come to equilibrium with the sedimented ASi, thus suppressing further dissolution (Conley et al. 1989). Other lakes and marine settings may have high diffusional fluxes out of the sediment, but in Lakes St. Croix and Pepin the process is somewhat muted.
Modern silica mass balance
It was not practical to directly measure in-lake ASi production (ASiin-lake) in these large, dynamic riverine lakes, so this flux was instead calculated by difference. Although it is possible in theory to subtract DSioutflow from DSiinflow to determine how much DSi has been absorbed from the water during its residence in the lake, the DSi fluxes are so large relative to the ASi fluxes that ASiin-lake cannot be reliably calculated by the difference in DSi.
Therefore, we chose a mass balance approach using the ASi fluxes, rather than the DSi fluxes, to solve for ASiin-lake:
Results from the mass balance calculations are shown in Table 3. After error is propagated through the mass balance, the uncertainties for the ASiin-lake terms are large, especially for Lake Pepin where ASiin-lake is 30,000 (±17,000) t yr−1. Much of that uncertainty is due to the high week-to-week variability of ASi concentrations in the lake inflows, as discussed above.
Despite the uncertainties, ASisedimented is apparently the largest silica flux in Lake St. Croix, and ASisedimented and ASiinflow are the largest in Lake Pepin. In Lake St. Croix, ASiinflow and ASiin-lake are about the same, while the Lake Pepin ASiinflow is larger than its ASiin-lake.
To directly compare the lakes to each other, the ASiin-lake results were normalized to lake surface area because Lake Pepin is much larger than Lake St. Croix. Lake Pepin is 2.3 times as productive as Lake St. Croix in terms of ASiin-lake, or diatom production, on an areal basis (Table 3).
Synthesis: amorphous silica production and trapping
The silica budgets for Lake St. Croix and Lake Pepin show that both lakes have large fluxes of DSi and an excess of DSi, such that diatom growth is likely never limited by DSi scarcity. An important consequence of the relatively low ASi production is that a small proportion of the total bioavailable silica (TSib) entering each lake is sequestered in the lake sediments. If trapping efficiency is calculated as ASisedimented divided by inflowing TSib, then today Lake St. Croix traps about 10% of the TSib, and Lake Pepin traps about 20%. That low trapping efficiency permits a majority of TSib to continue downriver toward the Gulf of Mexico. In addition, the combined amount of ASi sequestered in Lake St. Croix and Pepin sediments, 58,000 t yr−1, is only about 2% of the DSi load discharged by the Mississippi at its mouth, which is on the order of 2,500,000 t yr−1 (Kelly et al. 2001). And as we show elsewhere (Triplett et al. in preparation), ASi trapping in these lakes as a result of cultural eutrophication has increased over time.
However, these impoundments are at the far upstream reaches of the Mississippi River watershed; water must pass through 24 more navigational pools (i.e. artificial impoundments) and innumerable backwaters before reaching the Gulf of Mexico. Diatom production in the hypereutrophic Mississippi River continues to convert DSi to ASi during that passage, and sedimentation in artificial impoundments, backwaters and floodplain deposits removes additional diatom frustules (ASi) from circulation (Humborg et al. 2006). The median DSi concentration of the Mississippi River from 1996 to 2000 near its mouth in Louisiana was 6.27 mg l−1 (Kelly et al. 2001) and from 1981 to 1987 was 3.0 mg l−1 as measured by Turner and Rabalais (1991), both significantly lower than the median Lake Pepin outflow of 13.5 mg l−1 in 2004–2005. ASi sequestration in Lake St. Croix and Lake Pepin is just the first step in a long series of TSib reductions before the Mississippi reaches the Gulf of Mexico.
Our results also provide insights into the relationship between rivers and their impoundments. First, the rapid sedimentation rates in these riverine lakes, combined with high ambient DSi concentrations and low salinity, cause in-lake diatom dissolution to be small relative to other silica fluxes. While diatom dissolution is clearly an active and significant process in other environments, including in moving rivers where frustules are not buried, it is a minor factor in the silica cycling of Lake St. Croix and Lake Pepin.
Second, siliceous terrestrial phytoliths are transported by rivers in significant quantities. However, phytoliths from Lake St. Croix and Lake Pepin sediments are largely unaffected by the weak alkaline digestions (1% Na2CO3) applied here, so that the ASi measurements in this study are primarily diatom silica. Specifically, we hypothesize that Al ions have been incorporated into the surface layers of these well-traveled phytoliths thus diminishing their solubility. While these phytoliths are not bioavailable in the freshwater system (and are not included in the term TSib), they may become bioavailable upon reaching the ocean. If so, our limited sampling suggests that they would be a major source of bioavailable silica to the oceans because the quantities of phytolith Si in these lake sediments were on par with the mass of non-phytolith (or diatom) ASi.
Third, a large proportion of the suspended solid (TSS) load is ASi in both the St. Croix River (mean 12%) and the upper Mississippi River (mean 8%) (Fig. 7). The impoundments sequester much of the inflowing TSS, but diatom production is ongoing throughout the lakes so the ASi-to-TSS ratio increases downstream to a mean of 21% at Lake St. Croix’s outflow and 17% at Lake Pepin’s outflow. These ratios are much higher than values reported by Conley (1997), which were generally in the 1–10% range. The low TSS concentrations in the St. Croix (2–14 mg l−1 during the study period) help explain that lake’s high ratios, while the Mississippi’s high TSS concentrations (3–65 mg l−1) were counterbalanced by its high diatom productivity.

Concentrations of total suspended solids (TSS) and amorphous silica (ASi) in the Lake St. Croix and Lake Pepin inflows and outflows from April 2004 to March 2005
Fourth, ASi production in the St. Croix and Mississippi Rivers is of the same order of magnitude as ASi production in their respective lakes. In Lake St. Croix, ASiinflow (4,500 ± 1,100 t yr−1) from the St. Croix River is about equal to ASiin-lake (4,400 ± 1,700 t yr−1). In Lake Pepin, ASiinflow (49,500 ± 11,000 t yr−1) from the Mississippi River is twice the ASiin-lake (30,000 ± 17,000 t yr−1). The ASi production in the river channels is a reminder that efforts to manage nutrient loads, with the goal of improving the water quality of lakes and impoundments, must encompass a range of stoichiometric variables and should measure ASi as well as DSi fluxes.
Finally, the difference in river:lake productivity between these two systems (1:1 in Lake St. Croix and 2:1 in Lake Pepin) is most likely due to the higher concentrations of major nutrients (e.g. phosphorus) in the Mississippi River relative to the St. Croix River. Intensive agriculture in the watershed and municipal and industrial discharges to the river have caused the Mississippi to be extremely nutrient-rich. In fact, because the average TP concentration in Lake Pepin is four times that in Lake St. Croix, and considering that phosphorus is often cycled through multiple generations of algae during its residence in a lake, we would predict that the Pepin ASiin-lake would be at least four times that of Lake St. Croix. Instead, it is just 2.3 times more productive on an areal basis, indicating that diatom growth in Lake Pepin is severely inhibited by other factors such as light-limitation (higher turbidity) and competition from other algae that thrive in very nutrient-rich systems.
The production of diatoms and the trapping of ASi in these large riverine lake systems clearly demonstrates the importance of silica retention in lakes and reservoirs along the land–ocean continuum. More studies are needed to better quantify the trapping of ASi in rivers and to determine the role of phytoliths and diatoms in the global biogeochemical cycle of silica.
References
Applin KR (1987) The diffusion of dissolved silica in dilute aqueous solution. Geochim Cosmochim Acta 51:2147–2151
Battarbee RW (1973) A new method for estimating absolute microfossil numbers with special reference to diatoms. Limnol Oceanogr 18:647–653
Caraco NF (1993) Disturbance of the phosphorus cycle: a case of indirect effects of human activity. Trends Ecol Evol 8:51–54
Carlson RE (1977) A trophic state index for lakes. Limnol Oceanogr 22:361–369
Cary L, Alexandre A, Meunier J-D, Boeglin J-L, Braun J-J (2005) Contribution of phytoliths to the suspended load of biogenic silica in the Nyong basin rivers (Cameroon). Biogeochemistry 74:101–114
Conley DJ (2002) Terrestrial ecosystems and the global biogeochemical silica cycle. Global Biogeochem Cycles 16:1121. doi:10.1029/2002GB001894
Conley DJ, Schelske CL (1989) Processes controlling the benthic regeneration and sedimentary accumulation of biogenic silica in Lake Michigan. Archiv fur Hydrobiologie 116:23–43
Conley DJ, Schelske CL (2001) Biogenic silica. In: Smol JP, Birks HB, Last WM (eds) Tracking environmental change using lake sediments. Kluwer Academic Publishers, Dordrecht
Conley DJ, Kilham SS, Theriot EC (1989) Differences in silica content between marine and freshwater diatoms. Limnol Oceanogr 34:205–213
Conley DJ, Schelske CL, Stoermer EF (1993) Modification of the biogeochemical cycle of silica with eutrophication. Mar Ecol Prog Ser 101:179–192
Conley DJ, Wulff F, Stockenberg A, Carman R, Johnstone RW (1997) Sediment-water nutrient fluxes in the Gulf of Finland, Baltic Sea. Estuar Coast Shelf Sc 45:591–598
Conley DJ, Stalnacke P, Pitkanen H, Wilander A (2000) The transport and retention of dissolved silicate by rivers in Sweden and Finland. Limnol Oceanogr 45:1850–1853
DeMaster DJ (1981) The supply and accumulation of silica in the marine environment. Geochim Cosmochim Acta 45:1715–1732
Dixit AS, Van Cappellan P, Van Bennekom AJ (2001) Processes controlling solubility of biogenic silica and pore water build-up of silicic acid. Mar Chem 73:333–352
Edlund MB, Engstrom DR, Tripett LD, LaFrancois B, Leavitt P (in press) 20th century eutrophication of the St. Croix River (Minnesota-Wisconsin, USA) reconstructed from the sediments of its natural impoundment. J Paleolimnol
Engstrom DR, Almendinger JE, Wolin JA (in press) Historical changes in sediment and phosphorus loading to the Upper Mississippi River: mass-balance reconstructions from the sediments of Lake Pepin. J Paleolimnol
Foy RH (1985) Phosphorus inactivation in a eutrophic lake by the direct addition of ferric aluminium sulphate: impact on iron and phosphorus. Freshw Biol 15:613–629
Friedl G, Teodoru C, Wehrli B (2004) Is the Iron Gate I reservoir on the Danube River a sink for dissolved silica? Biogeochemistry 68:21–32
Friedrich J, Dinkel C, Grieder E, Radan S, Secrieru D, Steingruber S, Wehrli B (2003) Nutrient uptake and benthic regeneration in Danube Delta Lakes. Biogeochemistry 64:373–398
Gallego L, Distel RA, Camina R, Rodriguez Iglesias RM (2004) Soil phytoliths as evidence for species replacement in grazed rangelands of central Argentina. Ecography 27:725–732
Gibson CE, Wang G, Foy RH (2000) Silica and diatom growth in Lough Neagh: the importance of internal recycling. Freshw Biol 45:285–293
House WA, Denison FH, Warwick MS, Zhmud BV (2000) Dissolution of silica and the development of concentration profiles in freshwater sediments. Appl Geochem 15:425–438
Humborg C, Ittekkot V, Cociasu A, Bodungen Bv (1997) Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure. Nature 386:385–388
Humborg C, Blomqvist S, Avsan E, Bergensund Y, Smedberg E (2002) Hydrological alterations with river damming in northern Sweden: implications for weathering and river biogeochemistry. Global Biogeochem Cycles 16:12-1–12-12
Humborg C, Pastuszak M, Aigars J, Siegmund H, Morth C-M, Ittekkot V (2006) Decreased silica land-sea fluxes through damming in the Baltic Sea catchment—significance of particle trapping and hydrological alterations. Biogeochemistry 77:265–281
Hurd DC (1983) Physical and chemical properties of siliceous skeletons. In: Aston SR (ed) Silicon geochemistry and biogeochemistry. Academic Press, London
Icenhower JP, Dove PM (2000) The dissolution kinetics of amorphous silica into sodium chloride solutions: effects of temperature and ionic strength. Geochim Cosmochim Acta 64:4193–4203
Iler RK (1979) The chemistry of silica: solubility, polymerization, colloid and surface properties, and biochemistry. John Wiley & Sons, New York
Kelly VJ (2001) Concentrations and annual fluxes for selected water-quality constituents from the USGS National Stream Quality Accounting Network (NASQAN), 1996–2000. Water Resources Investigations Report 01-4255, US Geological Survey, Reston, VA
Kelley DW, Nater EA (2000) Source apportionment of lake bed sediments to watersheds in an Upper Mississippi basin using a chemical mass balance method. Catena 41:277–292
Krausse GL, Schelske CL, Davis CO (1983) Comparison of three wet-alkaline methods of digestion of biogenic silica in water. Freshw Biol 13:73–81
Lerman A (1979) Geochemical processes: water and sediment environments. John Wiley & Sons, New York
Li YH, Gregory S (1974) Diffusion of ions in sea water and in deep-sea sediments. Geochim Cosmochim Acta 38:703–714
Metropolitan Council Environmental Services (2006) Environmental Information Management System. http://www.es.metc.state.mn.us/eims/. Cited June 2006
Rabalais NN, Wiseman WJJ, Gupta BKS, Justic D, Turner RE, Dortch Q (1996) Nutrient changes in the Mississippi River and system responses on the adjacent continental shelf. Estuaries 19:386–407
Ryves DB, Battarbee RW, Juggins S, Fritz SC, Anderson NJ (2006) Physical and chemical predictors of diatom dissolution in freshwater and saline lake sediments in North America and West Greenland. Limnol Oceanogr 51:1355–1368
Saccone L, Conley DJ, Sauer D (2006) Methodologies for amorphous silica analysis. J Geochem Explor 88:235–238
Saccone L, Conley DJ, Koning E, Sauer D, Sommer M, Kaczorek D, Blecker SW, Kelly EF (2007) Assessing the extraction and quantification of amorphous silica in soils of forest and grassland ecosystems. Eur J Soil Sci 58:1446–1459
Sauer D, Saccone L, Conley DJ, Herrmann L, Sommer M (2006) Review of methodologies for extracting plant-available and amorphous Si from soils and aquatic sediments. Biogeochemistry 80:89–108
Schelske CL, Stoermer EF (1971) Eutrophication, silica depletion, and predicted changes in algal quality in Lake Michigan. Science 173:423–424
Sommer U (1988) Growth and survival strategies of planktonic diatoms. In: Sandgren CD (ed) Growth and reproductive strategies of freshwater phytoplankton. Cambridge University Press, Cambridge
Teodoru C, Dimopoulos A, Wehrli B (2006) Biogenic silica accumulation in the sediments of Iron Gate I reservoir on the Danube River. Aquat Sci 68:469–481
Tréguer P, Nelson DM, van Bennekom AJ, DeMaster DJ, Leynaert A, Queguiner B (1995) The silica balance in the world ocean: a reestimate. Science 268:375–379
Triplett LD, Engstrom DR, Edlund MB (in press) A whole-basin stratigraphic record of sediment and phosphorus loading to the St. Croix River, USA. J Paleolimnol
Turner RE, Rabalais NN (1991) Changes in Mississippi River water quality this century: implications for coastal food webs. Bioscience 41:140–147
Turner RE, Rabalais NN (1994) Coastal eutrophication near the Mississippi River Delta. Nature 368:619–621
Van Cappellan P, Dixit AS, van Beusekom J (2002) Biogenic silica dissolution in the oceans: reconciling experimental and field-based dissolution rates. Global Biogeochem Cycles 16:23
Wilke CR, Chang P (1955) Correlation of diffusion coefficients in dilute solutions. J Am Inst Chem Eng 2:264–270
Acknowledgements
We thank staff of the Limnological Research Center (LRC) at the University of Minnesota (U of MN), Jill Coleman Wasik at the St. Croix Watershed Research Station (SCWRS) for analytical assistance, Mark Edlund (SCWRS) for helpful insights, Rob Lusteck (U of MN) for initial phytolith identification, Robert Burdis (Minnesota Department of Natural Resources) for sampling assistance, and Amy Myrbo (LRC), Tony Case and two anonymous reviewers whose comments greatly improved this manuscript. DJC was supported by the European Union, Marie Curie program. Major support was provided by the Canon National Parks Science Scholars program.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Triplett, L.D., Engstrom, D.R., Conley, D.J. et al. Silica fluxes and trapping in two contrasting natural impoundments of the upper Mississippi River. Biogeochemistry 87, 217–230 (2008). https://doi.org/10.1007/s10533-008-9178-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-008-9178-7